
A Survey of Bacteria and Fungi Associated with Leaves, Rhizophylls, and Vesicles of the Carnivorous Plant Genlisea hispidula (Lentibulariaceae)



Abstract
1. Introduction
2. Materials and Methods
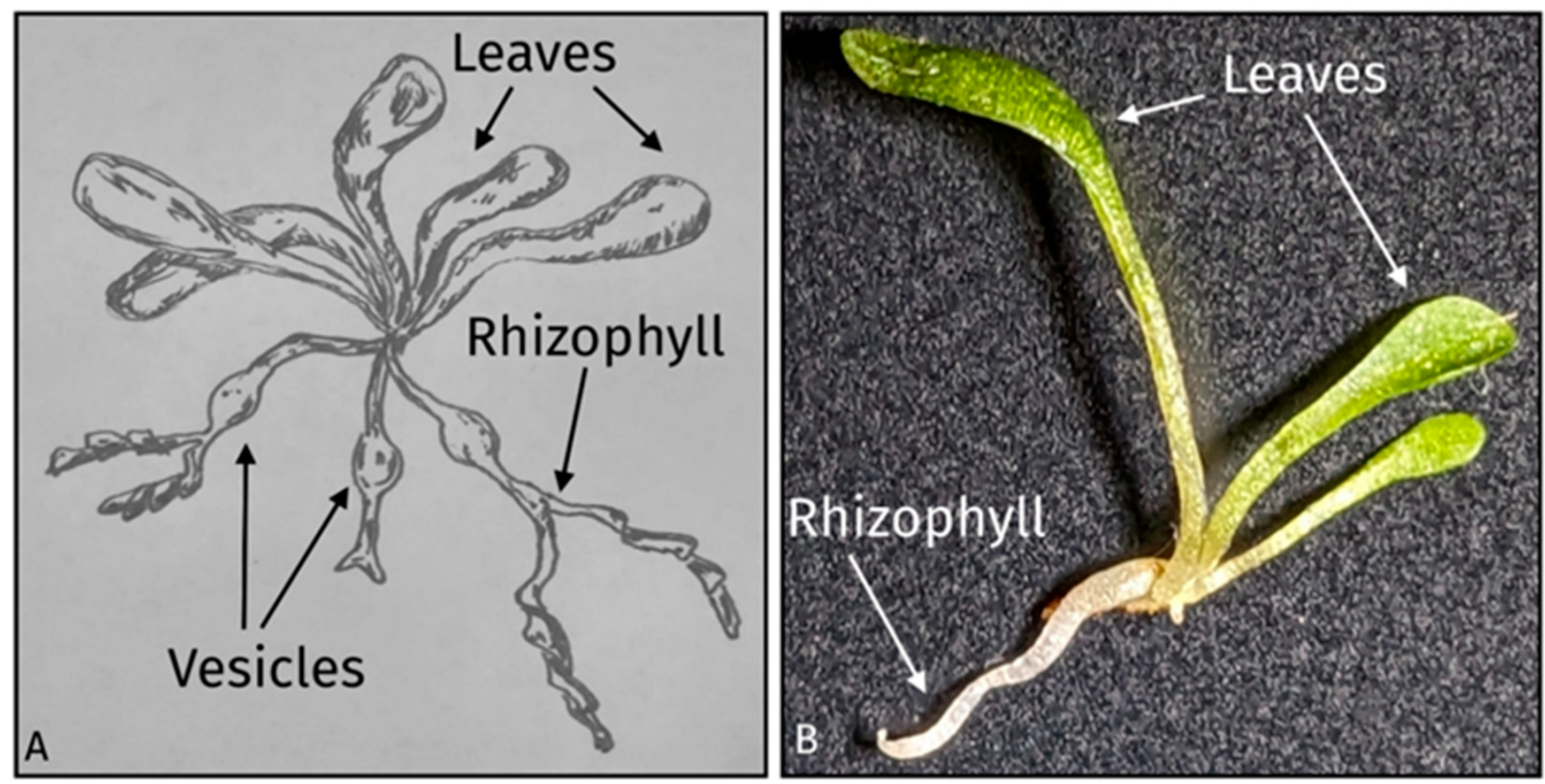
2.1. Genlisea Hispidula Plants, Growing Conditions, and Parts Examined

2.2. Tissue Preparation, Medium Inoculation, and Isolation
2.3. Molecular Identification
2.4. Unique Species Determination and Visualization
3. Results
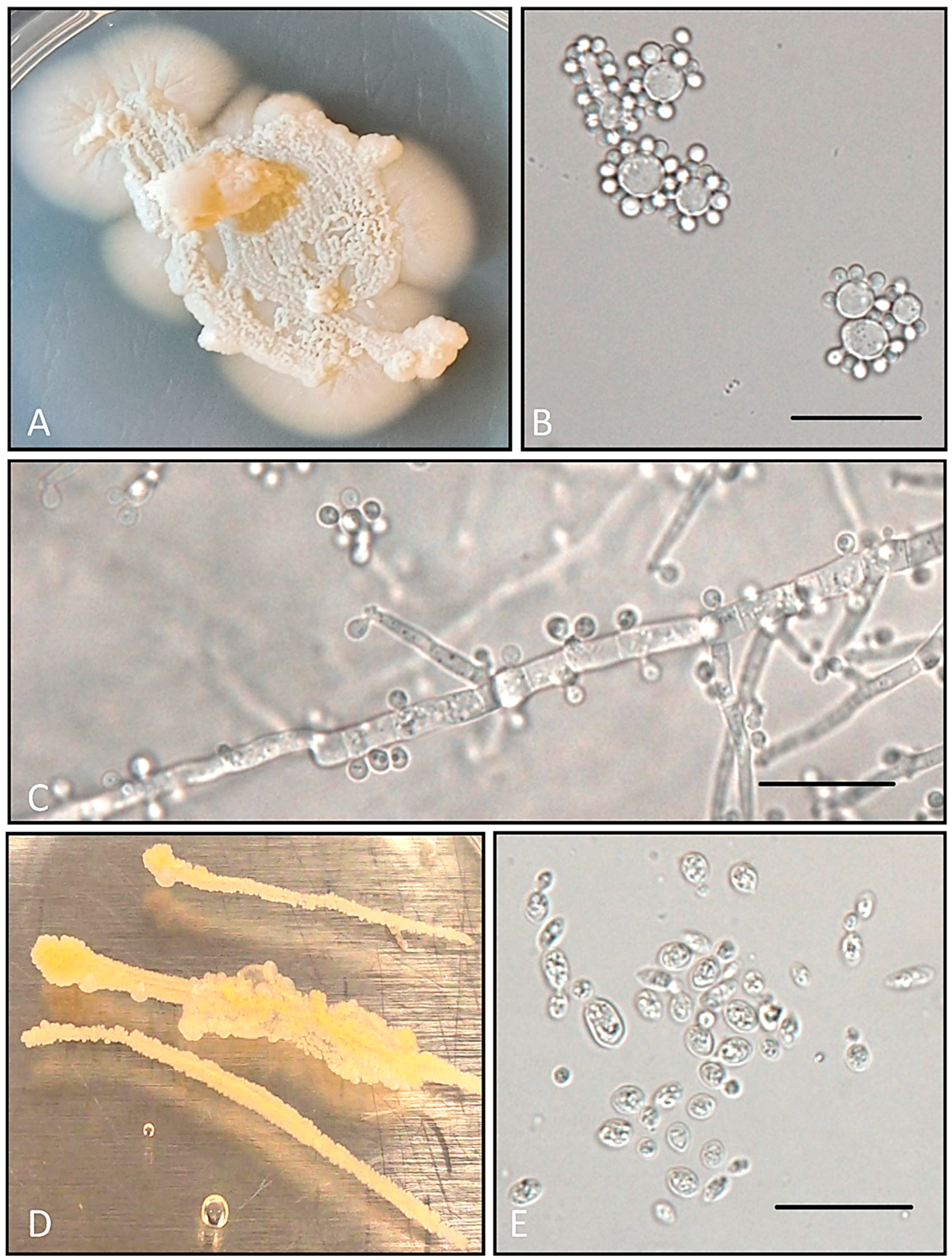
3.1. Bacteria
3.2. Fungi
3.3. Diversity and Novel Species
4. Discussion
4.1. Contribution of Bacteria and Fungi to Aid Host Digestion and Further Thoughts
4.2. Differences among G. hispidula Tissue Types
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cross, A.T.; Krueger, T.A.; Conella, P.M.; Robinson, A.S.; Fleischmann, A.S. Conservation of carnivorous plants in the age of extinction. Glob. Ecol. Conserv. 2020, 24, e01272. [Google Scholar] [CrossRef]
- Ellison, A.M.; Adamec, L. Introduction: What is a carnivorous plant? In Carnivorous Plants: Physiology, Ecology, and Evolution; Adamec, L., Ellison, A.M., Eds.; Oxford University Press: Oxford, UK, 2018. [Google Scholar] [CrossRef]
- Brewer, J.S.; Schlauer, J. Biogeography and habitats of carnivorous plants. In Carnivorous Plants: Physiology, Ecology, and Evolution; Adamec, L., Ellison, A.M., Eds.; Oxford University Press: London, UK, 2018. [Google Scholar]
- Adlassnig, W.; Koller-Peroutka, M.; Bauer, S.; Koshkin, E.; Lendl, T.; Lichtscheidl, I.K. Endocytotic uptake of nutrients in carnivorous plants. Plant J. 2012, 71, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Athauda, S.B.; Matsumoto, K.; Rajapakshe, S.; Kuribayashi, M.; Kojima, M.; Kubomura-Yoshida, N.; Iwamatsu, A.; Shibata, C.; Inoue, H.; Takahashi, K. Enzymic and structural characterization of Nepenthesin, a unique member of a novel subfamily of aspartic proteinases. Biochem. J. 2004, 381, 295–306. [Google Scholar] [CrossRef]
- Ravee, R.; Salleh, F.; Imadi, M.; Goh, H.H. Discovery of digestive enzymes in carnivorous plants with focus on proteases. PeerJ 2018, 6, e4914. [Google Scholar] [CrossRef]
- Leroy, C.; Carrias, J.F.; Céréghino, R.; Corbara, B. The contribution of microorganisms and metazoans to mineral nutrition in bromeliads. J. Plant Ecol. 2016, 9, 241–255. [Google Scholar] [CrossRef]
- Carmesin, C.F.; Fleischmann, A.S.; Klepsch, M.M.; Westermeier, A.S.; Speck, T.; Jansen, S.; Poppinga, S. Structural gradients and anisotropic hydraulic conductivity in the enigmatic eel traps of carnivorous corkscrew plants (Genilisea spp.). Am. J. Bot. 2021, 108, 2356–2370. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, A.; Schlauer, J.; Smith, S.A.; Givnish, T.J. Evolution of carnivory in angiosperms. In Carnivorous Plants: Physiology, Ecology, and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 180–193. [Google Scholar]
- Fleischmann, A. Systematics and evolution of Lentibulariaceae: II. Genlisea. In Carnivorous Plants: Physiology, Ecology, and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 81–88. [Google Scholar]
- Fischer, E.; Barthlott, W.; Seine, R.; Theisen, I. Lentibulariaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin, Germany, 2004. [Google Scholar]
- Taylor, P. The genus Genlisea. CPN 1991, 20, 20–26. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kozieradzka-Kiszkurno, M.; Świątek, P.; Darnowski, D.W. Prey attraction in carnivorous Genlisea (Lentibulariaceae). Acta Biol. Crac. Ser. Bot. 2008, 50, 87–98. [Google Scholar]
- Adamec, L. Oxygen concentrations inside the traps of the carnivourous plants Utricularia and Genlisea (Lentibulariaceae). Ann. Bot. 2007, 100, 849–856. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kozieradzka-Kiszkurno, M.; Świątek, P. Functional ultrastructure of Genlisea (Lentibulariaceae) digestive hairs. Ann. Bot. 2007, 100, 195–203. [Google Scholar] [CrossRef]
- Płachno, B.J.; Adamec, L.; Lichtscheidl, I.K.; Peroutka, M.; Adlassnig, W.; Vrba, J. Fluorescence labelling of phosphatase activity in digestive glands of carnivorous plants. Plant Biol. 2008, 8, 813–820. [Google Scholar] [CrossRef]
- Cao, H.X.; Schmutzer, T.; Scholz, U.; Pecinka, A.; Schubert, I.; Vu, G.T. Metatranscriptome analysis reveals host-microbiome interactions in traps of carnivorous Genlisea species. Front. Microbiol. 2015, 6, 526. [Google Scholar] [CrossRef] [PubMed]
- Caravieri, F.A.; Ferreira, A.J.; Ferreira, A.; Clivati, D.; de Miranda, V.F.O.; Araújo, W.L. Bacterial community associated with traps of the carnivorous plants Utricularia hydrocarpa and Genlisea filiformis. Aquat. Biol. 2014, 116, 8–12. [Google Scholar] [CrossRef]
- Koopman, M.M.; Carstens, B.C. The microbial phyllogeography of the carnivorous plant Sarracenia alata. Microb. Ecol. 2011, 61, 750–759. [Google Scholar] [CrossRef]
- Chan, X.Y.; Hong, K.W.; Yin, W.F.; Chan, K.G. Microbiome and biocatalytic bacteria in Monkey Cup (Nepenthes pitcher) digestive fluid. Sci. Rep. 2016, 6, 20016. [Google Scholar] [CrossRef]
- Płachno, B.J.; Adamus, K.; Faber, J.; Kozłowski, J. Feeding behaviour of carnivorous Genlisea plants in the laboratory. Acta Bot. Gall 2005, 152, 159–164. [Google Scholar] [CrossRef]
- Brune, A. Chapter 249-Symbiots aiding digestion. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 978–983. [Google Scholar]
- Romão, D.; Staley, C.; Ferreira, F.; Rodrigues, R.; Sabino, R.; Veríssimo, C.; Wang, P.; Sadowsky, M.; Brandão, J. Next-generation sequencing and culture-based techniques offer complementary insights into fungi and prokaryotes in beach sands. Mar. Pollut. Bull 2017, 119, 351–358. [Google Scholar] [CrossRef]
- Tang, C.Q.; Humphreys, A.M.; Fontaneto, D.; Barraclough, T.G. Effects of phylogenetic reconstruction method on the robustness of species delimitation using single-locus data. Methods Ecol. Evol. 2014, 5, 1086–1094. [Google Scholar] [CrossRef]
- Ludwig, W.; Klenk, H.P. Overview: A phylogenetic backbone and taxonomic framework for procaryotic systematics. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Boone, D.R., Castenholz, R.W., Garrity, G.M., Eds.; Springer: New York, NY, USA, 2001; pp. 49–65. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal barcoding consortium, & fungal barcoding consortium author list. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 8, 6–9. [Google Scholar]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; de Vries, M.; Verkleij, G.J.M.; Crous, P.W.; Boekhout, T.; et al. DNA barcoding analysis of more than 9000 yeasts isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Hedin, M. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef]
- Kekkonen, M.; Mutanen, M.; Kaila, L.; Nieminen, M.; Hebert, P.D. Delineating species with DNA Barcodes: A case of taxon dependent method performance in moths. PLoS ONE 2015, 10, e0122481. [Google Scholar] [CrossRef]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, N., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Raudabaugh, D.B.; Bach, E.M.; Allen, J.M.; Miller, A.N. Fungal communities of submerged fine detritus from temperate peatland and stream habitats. Aquat 2021, 86, 191–207. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.-H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Rossi-Tamisier, M.; Benamar, S.; Raoult, D.; Fournier, P.-E. Cautionary tale of using 16S rRNA gene sequence similarity values in identification of human-associated bacterial species. Int. J. Syst. Evol. Microbiol. 2015, 65, 1929–1934. [Google Scholar] [CrossRef] [PubMed]
- Sexton, W.K.; Fidero, M.; Spain, J.C.; Liang, L.; Bucalo, K.; Cruse-Sanders, J.M.; Pullman, G.S. Characterization of endophytic bacterial communities within greenhouse and field-grown rhizomes of three rare pitcher plant species (Sarracenia oreophila, S. leucophylla, and S. purpurea spp. venosa) with an emphasis on nitrogen-fixing bacteria. Plant Soil 2020, 447, 257–279. [Google Scholar] [CrossRef]
- Xiong, C.; Singh, B.K.; He, J.-Z.; Han, Y.-L.; Li, P.-P.; Wan, L.-H.; Meng, G.-Z.; Liu, S.-Y.; Wang, J.-T.; Wu, C.-F.; et al. Plant developmental stage drives the differentiation in ecological role of the maize microbiome. Microbiome 2021, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, W.E.; Creelman, R.A. Mutualism between the carnivorous purple pitcher plant and its inhabitants. Am. Midl. Nat. 1984, 112, 294–304. [Google Scholar] [CrossRef]
- Campisano, A.; Albanese, D.; Yousaf, S.; Pancher, M.; Donati, C.; Pertot, L. Temperature drives the assembly of endophytic communities’ seasonal succession. Environ. Microbiol. 2017, 19, 3353–3364. [Google Scholar] [CrossRef]
- Guzmán, J.P.S.; Nguyen, K.; Hart, S.C. Simple methods to remove microbes from leaf surfaces. J. Basic Microbiol. 2020, 60, 730–734. [Google Scholar] [CrossRef]
- Juniper, B.E.; Robins, R.J.; Joel, D.M. The Carnivorous Plants; Academic Press: London, UK, 1989. [Google Scholar]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowi, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evalulation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.E.; Brandl, M.T. Microbiology of the Phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, R.E.; Sesin, D.F.; Joshua, H.; Wilson, K.E.; Kempf, A.J.; Goklen, K.A.; Kuehner, D.; Gailliot, P.; Gleason, C.; White, R.; et al. Pneumocandins from Zalerion arboricola I. discovery and isolation. J. Antibiot. 1992, 45, 1853–1866. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barr, D.J.S. Isolation, Culture and Identification of Chytridiales, Spizellomycetales and Hyphochytriales. Zoosporic Fungi in Teaching and Research; Southeastern Publishing Corporation: Athens, GA, USA, 1987; pp. 118–120.
- Martinson, V.G. Rediscovering a forgotten system of symbiosis: Historical perspective and future potential. Genes 2020, 11, 1063. [Google Scholar] [CrossRef] [PubMed]
- Boonmak, C.; Khunnamwong, P.; Limtong, S. Yeast communities of primary and secondary peat swamp forests in southern Thailand. Antonie Van Leeuwenhoek 2020, 113, 55–69. [Google Scholar] [CrossRef]
- Aliyu, H.; Gorte, O.; Neumann, A.; Ochsenreither, K. Global transcriptome profile of the oleaginous yeast Saitozyma podzolica DSM 27192 cultuted in glucose and xylose. J. Fungi 2021, 7, 758. [Google Scholar] [CrossRef]
- Groenewald, M.; Lombard, L.; de Vries, M.; Lopez, A.G.; Smith, M.; Crous, P.W. Diversity of yeast species from Dutch garden soil and the description of six novel Ascomycetes. FEMS Yeast Res. 2018, 18, foy076. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Level | Bacteria 1 | Fungi | ||
|---|---|---|---|---|
| Filamentous Fungi 2 | Ascomycota Yeasts 3 | Basidiomycota Yeasts 3 | ||
| Phylum | 75% | - | - | - |
| Class | 78.5% | 80.9% | - | - |
| Order | 85% | 81.2% | - | - |
| Family | 86.5% | 88.5% | - | - |
| Genus | 94.5% | 94.3% | - | - |
| Species | 98.7% | 99.6% | 98.31% | 98.61% |
| Taxonomic Level | Isolate Identity | Unique bPTP Species (#) | Total Cultured Isolates (#) | Plant Tissue Type | ||
|---|---|---|---|---|---|---|
| Leaf | Rhizophyll | Vesicle | ||||
| Genus | Acidocella | 1 | 9 | Yes | Yes | Yes |
| Bacillus | 2 | 2 | Yes | Yes | ||
| Burkholderia | 8 | 13 | Yes | Yes | Yes | |
| Herminiimonas | 1 | 1 | Yes | |||
| Jatrophihabitans | 1 | 1 | Yes | |||
| Methylobacterium | 1 | 2 | Yes | |||
| Microbacterium | 1 | 4 | Yes | |||
| Novosphingobium | 7 | 7 | Yes | Yes | ||
| Paenibacillus | 1 | 1 | Yes | |||
| Paraburkholderia | 5 | 13 | Yes | Yes | ||
| Rhodopseudomonas | 2 | 5 | Yes | |||
| Family | Alcaligenaceae | 8 | 8 | Yes | Yes | Yes |
| Nitrobacteraceae | 5 | 5 | Yes | |||
| Rhodospirillaceae | 1 | 1 | Yes | |||
| Sphingomonadaceae | 1 | 1 | Yes | |||
| Order | Burkholderiales | 1 | 1 | Yes | ||
| Micrococcales | 1 | 1 | Yes | |||
| Phylum | Pseudomonadota sp. | 1 | 1 | Yes | ||
| Totals | 48 | 76 | ||||
| Taxonomic Level | Isolate Identity | Unique bPTP Species (#) | Total Cultured Isolates (#) | Plant Tissue Type | ||
|---|---|---|---|---|---|---|
| Leaf | Rhizophyll | Vesicle | ||||
| Genus | Aspergillus sp. | 1 | 2 | Yes | ||
| Cladosporium spp. | 2 | 2 | Yes | |||
| Penicillium spp. | 8 | 12 | Yes | Yes | ||
| Saitozyma spp. | 11 | 14 | Yes | Yes | Yes | |
| Talaromyces spp. | 2 | 3 | Yes | Yes | ||
| Toxicocladosporium spp. | 3 | 5 | Yes | |||
| Trichomonascus sp. | 1 | 20 | Yes | Yes | Yes | |
| Order | Eurotiales sp. | 1 | 1 | Yes | ||
| Totals | 29 | 59 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raudabaugh, D.B.; Aime, M.C. A Survey of Bacteria and Fungi Associated with Leaves, Rhizophylls, and Vesicles of the Carnivorous Plant Genlisea hispidula (Lentibulariaceae). Diversity 2024, 16, 77. https://doi.org/10.3390/d16020077
Raudabaugh DB, Aime MC. A Survey of Bacteria and Fungi Associated with Leaves, Rhizophylls, and Vesicles of the Carnivorous Plant Genlisea hispidula (Lentibulariaceae). Diversity. 2024; 16(2):77. https://doi.org/10.3390/d16020077
Chicago/Turabian StyleRaudabaugh, Daniel B., and M. Catherine Aime. 2024. "A Survey of Bacteria and Fungi Associated with Leaves, Rhizophylls, and Vesicles of the Carnivorous Plant Genlisea hispidula (Lentibulariaceae)" Diversity 16, no. 2: 77. https://doi.org/10.3390/d16020077
APA StyleRaudabaugh, D. B., & Aime, M. C. (2024). A Survey of Bacteria and Fungi Associated with Leaves, Rhizophylls, and Vesicles of the Carnivorous Plant Genlisea hispidula (Lentibulariaceae). Diversity, 16(2), 77. https://doi.org/10.3390/d16020077