
Global Potential Distribution of Sarcophaga dux and Sarcophaga haemorrhoidalis under Climate Change




Abstract
1. Introduction
2. Materials and Methods
2.1. Input Data
2.2. Environmental Niche Modeling
2.3. Model Robustness
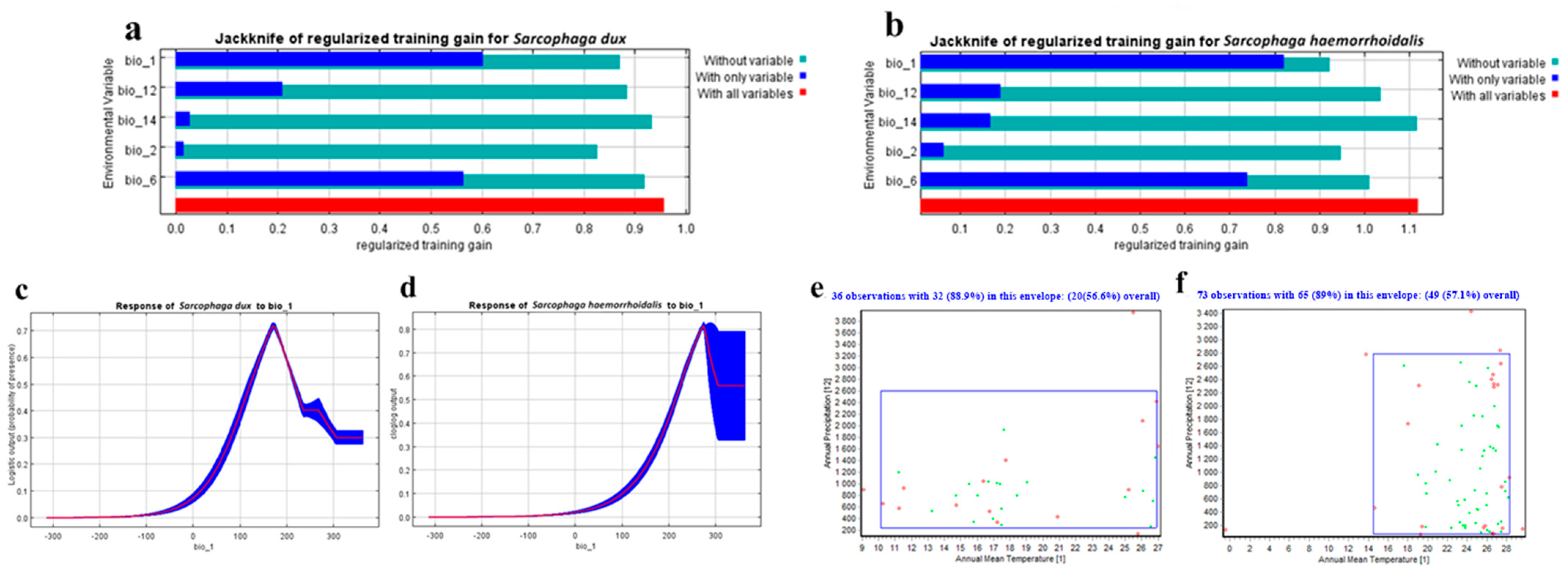
3. Results
3.1. Model Evaluation and Contribution of Environmental Covariates
3.2. Two-Dimensional Niche Analysis
3.3. Current Habitat Suitability Models
3.4. Future Habitat Suitability Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pape, T.; Blagoderov, V.; Mostovski, M.B. Order Diptera Linnaeus, 1758. Zootaxa 2011, 3148, 222–229. [Google Scholar] [CrossRef]
- Byrd, J.H.; Castner, J.L. Forensic Entomology: The Utility of Arthropods in Legal Investigations; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Hall, M.J.R.; Wall, R.; Stevens, J.R. Traumatic Myiasis: A Neglected Disease in a Changing World. Annu. Rev. Entomol. 2016, 61, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, A.; Pape, T. Sarcophagidae (Diptera) of Thailand: A Review of Current Knowledge. Zootaxa 2012, 3423, 1–73. [Google Scholar] [CrossRef]
- Mello-Patiu, C.A.; Vargas, H.A. The Importance of Flies in Pollination. J. Pollinat. Ecol. 2009, 1, 1–6. [Google Scholar] [CrossRef]
- Whitmore, D.; Wall, R. Mites and Insects in Wound Healing: An Update and Review. Trauma 2018, 20, 3–17. [Google Scholar] [CrossRef]
- Yeates, D.K.; Wiegmann, B.M. Congruence and Controversy: Toward a Higher-Level Phylogeny of Diptera. Annu. Rev. Entomol. 1999, 44, 397–428. [Google Scholar] [CrossRef]
- Zhang, D.; Yan, L.; Zhang, M.; Chu, H.; Cao, J.; Li, K.; Zhang, R. The Mitochondrial Genome of Sarcophaga dux (Diptera: Sarcophagidae), a Species with High Invasion Ability. Mitochondrial DNA Part B 2019, 4, 2284–2285. [Google Scholar] [CrossRef]
- Ebejer, M.J.; Falzon, V. A Review of the Sarcophagidae (Diptera) of Malta, with the First Record of Sarcophaga (Liopygia) ruficornis from the Maltese Islands. J. Entomol. Res. Soc. 2018, 20, 23–35. [Google Scholar]
- Hall, M.J.R.; Wall, R. Myiasis of Humans and Domestic Animals. Adv. Parasitol. 1995, 35, 257–334. [Google Scholar] [CrossRef]
- Makhubo, B.G.; Githure, J.I. Diversity and Distribution of Sarcophagid Flies (Diptera: Sarcophagidae) in Nairobi and Its Environs, Kenya. J. Insect Sci. 2013, 13, 1–10. [Google Scholar] [CrossRef]
- Sukontason, K.; Narongchai, P.; Kanchai, C.; Vichairat, K.; Sribanditmongkol, P.; Bhoopat, T.; Kurahashi, H.; Chockjamsai, M.; Piangjai, S.; Bunchu, N.; et al. Forensic entomology cases in Thailand: A review of cases from 2000 to 2006. Parasitol. Res. 2007, 101, 1417–1423. [Google Scholar] [CrossRef]
- Wells, J.D.; Greenberg, B. Courtship Behavior of the Flesh Flies (Diptera: Sarcophagidae). J. Insect Behav. 1992, 5, 185–199. [Google Scholar] [CrossRef]
- Sherman, R.A.; Hall, M.J.R.; Thomas, S. Medicinal Maggots: An Ancient Remedy for Some Contemporary Afflictions. Annu. Rev. Entomol. 2000, 45, 55–81. [Google Scholar] [CrossRef] [PubMed]
- Barták, M.; Khrokalo, L.; Verves, Y. New records, synonyms and combinations for oriental Sarcophagidae (Diptera), with updated checklists for Cambodia, India, Taiwan, Thailand and Vietnam. J. Asia Pacific Entomol. 2019, 22, 44–55. [Google Scholar]
- Bhattacharjee, D.; Halder, S. Medical Maggots: An Ancient Remedy for Wound Healing. J. Egypt. Soc. Parasitol. 2017, 47, 647–651. [Google Scholar] [CrossRef]
- Catts, E.P.; Goff, M.L. Forensic Entomology in Criminal Investigations. Annu. Rev. Entomol. 1992, 37, 253–272. [Google Scholar] [CrossRef]
- Grassberger, M.; Reiter, C. Effect of Temperature on Lucilia sericata (Diptera: Calliphoridae) Development with Special Reference to the Isomegalen- and Isomorphen-Diagram. Forensic Sci. Int. 2002, 128, 177–182. [Google Scholar] [CrossRef]
- Marino, A.; Putzolu, M.; Mazzarello, V.; Dessy, L.A. Maggot Therapy for the Treatment of Diabetic Foot Ulcers: A Meta-Analysis. J. Wound Care 2016, 25, 139–146. [Google Scholar] [CrossRef]
- Jordaens, K.; Sonet, G.; Richet, R.; Dupont, E.; Braet, Y. A Molecular Phylogenetic Analysis of Sarcophagidae (Diptera) and the Evolution of Larval Myiasis within the Calyptratae. Int. J. Parasitol. 2012, 42, 353–363. [Google Scholar] [CrossRef]
- Czajka, M.A.; Leather, S.R. A Review of Pest Control Strategies and Their Potential for Use in Sustainable and Ecologically Sound Pest Management. Crop Prot. 2016, 83, 37–45. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. In Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Nica, A.; Popescu, A.; Ibanescu, D.C. Human influence on the climate system. Curr. Trends Nat. Sci. 2019, 8, 209–215. [Google Scholar]
- Kassim, N.F.; Yaakop, S.; Idris, A.B. Effect of Temperature on Development of Sarcophaga (Liosarcophaga) dux Thomson (Diptera: Sarcophagidae) in Malaysia. Trop. Biomed. 2014, 31, 283–291. [Google Scholar]
- Matuszewski, S. An Initial Study of Insect Succession and Carrion Decomposition in Various Forest Habitats of Central Europe. Forensic Sci. Int. 2013, 231, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Dillon, M.E.; Wang, G.; Huey, R.B. Global metabolic impacts of recent climate warming. Nature 2010, 467, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Duarte, C.M.; Poloczanska, E.; Richardson, A.J.; Singer, M.C. Overstretching attribution. Nat. Clim. Chang. 2011, 1, 2–4. [Google Scholar] [CrossRef]
- Cox, J.S.H. The Role of Geographic Information Systems and Spatial Analysis in Area-Wide Vector Control Programm. In Area-wideControl of Insect Pests; Springer: Dordrecht, The Netherlands, 2007; pp. 199–209. [Google Scholar]
- Bässler, C.; Heilmann-Clausen, J.; Karasch, P.; Brandl, R.; Halbwachs, H. Climate Change Disproportionately Increases Habitat Loss for Rare Species in Fragmented Landscapes. Glob. Chang. Biol. 2020, 26, 3604–3619. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.; Yang, D. Global Warming Threatens Biodiversity and Ecosystem Services: A Review. J. Clean. Prod. 2022, 334, 130470. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Guo, X.; Zhang, Y.; Wang, Z. Climate Change and the Distributional Dynamics of the Housefly (Musca domestica) in China. Environ. Monit. Assess. 2020, 192, 761. [Google Scholar] [CrossRef]
- Sgrò, C.M.; Lowe, A.J.; Hoffmann, A.A. Building Resilience to Climate Change in Australia’s Biodiversity Hotspots. Trends Ecol. Evol. 2023, 38, 1–3. [Google Scholar] [CrossRef]
- Sukontason, K.L.; Sanit, S.; Klong-Klaew, T.; Tomberlin, J.K.; Sukontason, K. Sarcophaga (Liosarcophaga) dux (Diptera: Sarcophagidae): A flesh fly species of medical importance. Biol. Res. 2014, 47, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.K.; Mahato, S. Intra-puparial development of flesh fly Sarcophaga dux (Thomson) (Diptera, Sarcophagidae). Curr. Sci. 2016, 25, 1063–1070. [Google Scholar] [CrossRef]
- Braverman, I.; Dano, I.; Saah, D.; Gapany, B. Aural myiasis caused by flesh fly larva, Sarcophaga haemorrhoidalis. J. Oto. 1994, 23, 204–205. [Google Scholar]
- Abdel-Hafeez, E.H.; Mohamed, R.M.; Belal, U.S.; Atiya, A.M.; Takamoto, M.; Aosai, F. Human wound myiasis caused by Phormia regina and Sarcophaga haemorrhoidalis in Minia Governorate, Egypt. Parasitol. Res. 2015, 114, 3703–3709. [Google Scholar] [CrossRef]
- Haseman, L. Sarcophaga haemorrhoidalis Larvae as Parasites of the Human Intestine (Dipt.). Entomol. News. 1917, 28, 343. [Google Scholar]
- Ndueze, O.U.; Noutcha, M.A.; Umeozor, O.C.; Okiwelu, S.N. Arthropods associated with wildlife carcasses in lowland rainforest, Rivers State, Nigeria. Europ. J. Exper. Biol. 2013, 3, 111–114. [Google Scholar]
- Hosni, E.M.; Nasser, M.; Al-Ashaal, S.; Rady, M.H.; Kenawy, M.A. Modeling current and future global distribution of Chrysomya bezziana under changing climate. Sci. Rep. 2020, 10, 4947. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shaara, H.; Alashaal, S.A.; Hosni, E.M.; Nasser, M.G.; Ansari, M.J.; Alharbi, S.A. Modeling the Invasion of the Large Hive Beetle, Oplostomus fuligineus, into North Africa and South Europe under a Changing Climate. Insects 2021, 12, 275. [Google Scholar] [CrossRef] [PubMed]
- Escobar, L.E.; Lira-Noriega, A.; Medina-Vogel, G.; Peterson, A.T. Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: Use of Maxent and Niche A to assure strict model transference. Geospat. Health 2014, 9, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Hosni, E.M.; Nasser, M.; Al-Khalaf, A.A.; Al-Shammery, K.A.; Al-Ashaal, S.; Soliman, D. Invasion of the Land of Samurai: Potential Spread of Old-World Screwworm to Japan under Climate Change. Diversity 2022, 14, 99. [Google Scholar] [CrossRef]
- Hosni, E.M.; Al-Khalaf, A.A.; Nasser, M.G.; Abou-Shaara, H.F.; Radwan, M.H. Modeling the Potential Global Distribution of Honeybee Pest, Galleria mellonella under Changing Climate. Insects 2022, 13, 484. [Google Scholar] [CrossRef]
- Hosni, E.M.; Al-Khalaf, A.A.; Naguib, R.M.; Afify, A.E.; Abdalgawad, A.A.; Faltas, E.M.; Hassan, M.A.; Mahmoud, M.A.; Naeem, O.M.; Hassan, Y.M.; et al. Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS. Diversity 2022, 14, 578. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 20 March 2020).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of Climate Change on the Future of Biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A Globally Coherent Fingerprint of Climate Change Impacts across Natural Systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Díaz, S.; Settele, J.; Brondízio, E.; Ngo, H.T.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; et al. Pervasive Human-Driven Decline of Life on Earth Points to the Need for Transformative Change. Science 2019, 366, eaax3100. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating Extinction Risk from Climate Change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Ekström, J.; Åkerrén Ögren, J.; Sjöblom, T. Exact Probability Distribution for the ROC Area under Curve. Cancers 2023, 15, 1788. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar]
- Zehra, N.; Mishra, V. An entomotoxicological study on the influence of house hold toxins on the colonization of carrion by Sarcophaga haemorrhoidalis. J. Environ. Sci. Toxicol. Food Technol. 2020, 14, 40–43. [Google Scholar]
- Zhang, X.; Li, Y.; Shang, Y.; Ren, L.; Chen, W.; Wang, S.; Guo, Y. Development of Sarcophaga dux (diptera: Sarcophagidae) at constant temperatures and differential gene expression for age estimation of the pupae. J. Therm. Biol. 2020, 93, 102735. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.; Sahlén, G.; Rämert, B. The Effects of Climate Change on Forensic Entomology: A Review. J. Forensic Sci. 2020, 65, 1030–1036. [Google Scholar] [CrossRef]
- Benbow, M.E.; Fjong, S.; Nguyen, J.; Tomberlin, J.K. Microbial Ecology of Carrion Decomposition in the Context of Climate Change. Front. Ecol. Evol. 2020, 8, 24. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Variables | Description | Contribution Percentages for S. dux | Contribution Percentages for S. haemorrhoidalis |
|---|---|---|---|
| Bio 1 | Annual Mean Temperature | 55.2% | 77.1% |
| Bio 6 | Min Temperature of the Coldest Month | 16.6% | 8.1% |
| Bio 12 | Annual Precipitation | 14.2% | 6.6% |
| Bio 2 | Mean Diurnal Range (mean of monthly max temp–min temp) | 8.9% | 6.3% |
| Bio 14 | Precipitation of the Driest Month | 5.1% | 1.9% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Khalaf, A.A.; Nasser, M.G.; Hosni, E.M. Global Potential Distribution of Sarcophaga dux and Sarcophaga haemorrhoidalis under Climate Change. Diversity 2023, 15, 903. https://doi.org/10.3390/d15080903
Al-Khalaf AA, Nasser MG, Hosni EM. Global Potential Distribution of Sarcophaga dux and Sarcophaga haemorrhoidalis under Climate Change. Diversity. 2023; 15(8):903. https://doi.org/10.3390/d15080903
Chicago/Turabian StyleAl-Khalaf, Areej A., Mohamed G. Nasser, and Eslam M. Hosni. 2023. "Global Potential Distribution of Sarcophaga dux and Sarcophaga haemorrhoidalis under Climate Change" Diversity 15, no. 8: 903. https://doi.org/10.3390/d15080903
APA StyleAl-Khalaf, A. A., Nasser, M. G., & Hosni, E. M. (2023). Global Potential Distribution of Sarcophaga dux and Sarcophaga haemorrhoidalis under Climate Change. Diversity, 15(8), 903. https://doi.org/10.3390/d15080903