Abstract
The Eocene-Oligocene Transition (EOT) was one of the most profound climate changes in the Cenozoic era, characterized by global cooling around 34 million years ago. This time period also witnessed major faunal turnovers, such as the “Mongolian Remodeling” of Asia, characterized by the dominance of rodents and lagomorphs after the EOT. Previous studies have primarily focused on overall faunal change across the EOT. Here, we examined one genus, the earliest ctenodactylid Karakoromys, based on rich fossils from continuous sections at Ulantatal, Nei Mongol, magnetostragraphically dated to latest Eocene-Early Oligocene. Based on a systematic paleontological study of these fossils, we recognized four species of Karakoromys (Karakoromys decussus, K. arcanus, K. chelkaris, and K. conjunctus sp. nov.), indicating a relatively high diversity of the most primitive ctenodactylids during the latest Eocene-Early Oligocene (~34.9–30.8 Ma). The turnover of ctenodactylids primarily occurred during a regional aridification event around 31 Ma rather than during the EOT cooling event, suggesting that regional precipitation variation in the semi-arid area may have played a more important role than global temperature change in the evolution of early ctenodactylids.
1. Introduction
The Eocene–Oligocene transition (EOT) marked a major climate shift in the Cenozoic era from a largely ice-free greenhouse world to an icehouse world around 34 million years ago (Ma). This event was characterized by a rapid decrease of nearly 5 °C in global mean temperature, the formation of a permanent ice sheet in Antarctica, and a sea level drop of nearly 80 m [1,2,3,4,5,6,7]. Around the same time, major faunal turnover events occurred in various continents, including the “Grande Coupure” in Europe and the “Mongolian Remodeling” in Asia, while the change in North America was less pronounced. These faunal turnovers were generally considered to be coupled with the EOT cooling event [8,9,10,11,12,13]. Previous studies have primarily focused on the overall faunal change across the EOT but paid less attention to the evolution of individual groups [11,12,13].
Ctenodactyloidea is one of the most common rodent groups in the Paleogene of Asia, with only the clade Ctenodactylidae surviving the EOT and continuing to the present [14,15,16,17]. Karakoromys, the earliest member of the family Ctenodactylidae, was named by Matthew and Granger [18] based on a pair of mandibles from the Hsanda Gol Formation in the Tsagaan Nuur Basin of Mongolia. Since then, this genus has been documented in many Early Oligocene fossil sites across East and Central Asia (Figure 1a). These areas include, but are not limited to, Shunkht, South Gobi Province [19]; Boongeen Gol, Ulan Ganga, and Khatan Khayrkhan, Gobi Altai Province [20]; Valley of Lakes [21,22,23,24,25,26,27], Mongolia; Saint Jacques [14], Qianlishan District [14,28], and Ulantatal (including Kekeamu) [14,29,30,31,32,33], Nei Mongol; Lanzhou Basin, Gansu [34]; Burqin Basin, Xinjiang [13,35,36]; and Zaisan Basin, Kazakhstan [37]. Although most Karakoromys fossils were found to come from Lower Oligocene strata, cf. Karakoromys was also found at the K10 site in the Late Eocene Aksyir Svita in the Zaysan Basin [37]. In addition, the lowest fossil level of Kekeamu with Karakoromys was once considered to be the earliest Oligocene [14,30], but our recent biostratigraphic and magnetostratigraphic studies have shown that Kekeamu fauna originate from the latest Eocene, which occurred about 34.9 Ma [32,33].

Figure 1.
(a) Distribution of Karakoromys fossil sites. The yellow circle represents Ulantatal, and the red squares represent other fossil sites. Sites 1–5 are from Mongolia: 1. Shunkht; 2. Valley of Lakes; 3. Ulan Ganga; 4. Khatan Khayrkhan; 5. Boongeen Gol; Sites 6–10 are from China: 6. Saint Jacques; 7. Qianlishan; 8. Ulantatal; 9. Lanzhou; 10. Burqin; Sites 11–13 are from Kazakhstan: 11. Zaisan; 12. Kyzyl Kak; 13. Chelkar-Teniz Lake. (b) Satellite image of the Ulantatal area (8) showing four sections and two previous fossil sites. Abbreviations: KM, Kekeamu section; SJ, Shangjing section; UTM, Ulantatal Main Section; SZT, Shaozengtu section. (c) Simplified lithologic column of the lower part of the Ulantatal sequence with line segments representing different sections (Adapted with permission from Ref. [38]. 2021, Wasiljeff et al.). Only the lower part of UTM is shown.
Since 2009, our research group has carried out extensive annual surveys and excavation in the Ulantatal area, Nei Mongol [32,33]. Hundreds of well-preserved Karakoromys specimens have been collected at a high chronologic resolution, providing ideal material for further understanding the taxonomy, stratigraphic distribution, and diversification of this group, as well as its response to global and regional climatic changes.
2. Geological Setting
The Ulantatal fossil site is 42 km northwest of Bayanhot Town, Alxa Zuoqi, Nei Mongol (Figure 1). It is located in the Bayanhot Basin where Meso-Cenozoic continental red clastic rocks are widely deposited with the absence of Upper Cretaceous, Paleocene, and Lower Eocene rocks [39,40,41]. Upper Eocene–Late Miocene strata are exposed in the Ulantatal area, which possess particularly well-exposed Oligocene strata that are rich in mammalian fossils.
The Ulantatal Formation is primarily composed of massive yellowish-brown-reddish clayey siltstones or siltstones, with occasional thin intercalations of parallel and cross-laminated fine-sand lenses in the bottom and middle. It total thickness is over 100 m. Based on the massive structure and bimodal grain-size distribution, the Ulantatal sediments are interpreted to be primarily Eolian in origin [42]. The strata of the Ulantatal Formation are not significantly deformed, with beds dipping gently at about 1–2 degrees towards the west-southwest (260°). The Ulantatal Formation unconformably overlies the Eocene Chaganbulage (Qaganbulag) Formation and is unconformably overlain by the Miocene Wuertu Formation. Based on paleomagnetic reversal stratigraphy and biostratigraphical correlation, the Ulantatal Formation is found to span a temporal range of ca. 34.9 to 27 Ma ([32,33,42]).
For the convenience of stratigraphic investigations and fossil collection, we informally divided the Ulantatal area into four sections: Kekeamu (KM), Shangjing (SJ), Ulantatal Main Section (UTM), and Shaozengtu (SZT) (Figure 1b). The Karakoromys fossils studied here were primarily collected from the KM and SJ sections, with sporadic specimens taken from the lower part of the UTM. The Karakoromys fossils span 0–60 m of the Ulantatal composite profile, corresponding to a time interval of 34.9–30.8 Ma (Figure 1c).
3. Materials and Methods
3.1. Materials
The materials examined in this study consisted of 273 jaw fragments or isolated teeth, along with several broken skulls associated with mandibles (see Table S1). These specimens were collected either from the surface or by excavation, and all had GPS coordinates.
3.2. Determining the Age of Fossils
The precise stratigraphic levels of fossils were directly measured in the field for most specimens, but not for all. To estimate the levels of specimens without recorded stratigraphic levels, we mapped these specimens on the software Google Earth Pro v.7.3 using their GPS coordinates. Then, their stratigraphic levels were estimated based on the elevation model of Google Earth Pro and the geomorphic location of the fossil spots, constrained by adjacent upper and lower fossil spots with stratigraphic known levels. We found that the GPS coordinates and our estimations of fossil spot levels were reliable, as verified by a re-examination of the levels of some specimens in the field. The numerical age of each fossil spot was then estimated based on the age-depth model developed with magnetostratigraphic data [33].
3.3. Systematic Paleontological Study
The terminology used for the structures of the skull and mandible followed those of Wible et al. [43]. The dental terminology primarily followed Wang [14]. In the descriptions, frequency of a character variant is expressed as a ratio. The measurements of tooth length and width, as well as the description of sinuses (sinusids) on the tooth, are indicated in Figure 2. The depth of the sinus (sinusids) was defined as the distance from top to bottom (and is not marked in Figure 2). Tooth measurements were conducted using an ocular micrometer on a Zeiss Stemi 2000 microscope with a precision of 0.02 mm. Photographs of specimen were taken using the JSM-6100 scanning electron microscope (SEM) at the Key Laboratory of Vertebrate Evolution and Human Origins of the Chinese Academy of Sciences. All fossil specimens were housed at the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP).
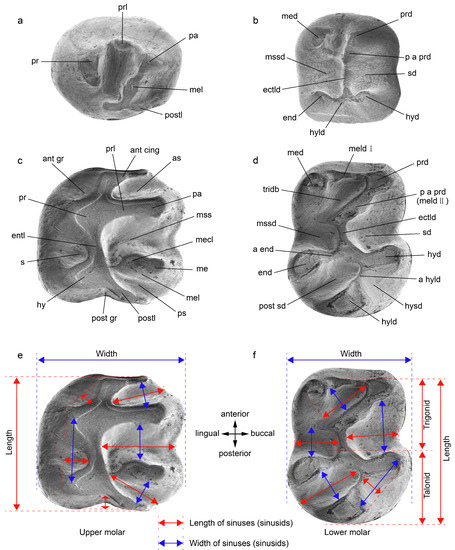
Figure 2.
Terminology for occlusal dental morphology of Ctenodactylidae (Adapted with permission from Ref. [14]. 1997, Wang). (a) left upper premolar, (b) right lower premolar, (c) left upper molar, (d) right lower molar. (e,f) Definition of lengths and widths of teeth and structures. Upper teeth: ant cing, anterior cingulum (=anteroloph); ant gr, anterior groove; as, anterosinus; entl, entoloph; hy, hypocone; me, metacone; mecl, metaconule; mel, metaloph; mss, mesosinus; pa, paracone; postl, posteroloph (=posterior cingulum); post gr, posterior groove; pr, protocone; prl, protoloph; ps, posterosinus; s, sinus. Lower teeth: a end, arm of entoconid; a hyld, arm of hypoconulid; ectld, ectolophid; end, entoconid; hyd, hypoconid; hyld, hypoconulid; hysd, hyposinusid; med, metaconid; meld I, metalophid I; meld II, metalophid II; mssd, mesosinusid; p a prd, posterior arm of protoconid; post sd, posterosinusid; prd, protoconid; sd, sinusid; tridb, trigonid basin.
3.4. Statistics
To determine if there were size differences between species, we ran one-way analyses of variance (ANOVA) for the length and width of M2/m2. Multiple comparisons were performed using both Fisher’s least-significant difference test (LSD) and Scheffe’s test. Analyses were performed with the SPSS 20.0 [44]. The boxplots and bivariate graphs were plotted in Past 4.11 [45].
3.5. Diversity Analysis
To study the community structure and succession of Karakoromys, we compiled the number of specimens for each species in 0.4-million-year time bins. The duration of time bins was chosen after giving consideration to both the temporal resolution and the number of specimens in each time bin. Aside from Karakoromys, Euryodontomys, Tataromys, Alashania and Bounomys were also found to occur in the lower part of the Ulantatal Formation. Euryodontomys was considered the sister group of Karakoromys and was included in the same subfamily Karakoromyinae, while Tataromys, Alashania, and Bounomys were thought to be relatively distantly related to the Karakoromys [14,16]. Given the close phylogenetic relationship between Euryodontomys ampliatus and Karakoromys and their occurrence in the same stratigraphic set, we also included unpublished specimens of E. ampliatus in the statistical analyses. We calculated the percentage of specimens of each species as the relative abundance and in plotted time series graphs of relative abundance. The median of each time bin was used as the age of that time bin. Data from the time interval 34.7–34.3 Ma were lacking, as fossils had not yet been collected from the corresponding strata.
Stratigraphically constrained cluster analysis (CONISS) was used to demarcate stages of community succession. This is a multivariate method for the quantitative definition of stratigraphic zones and is widely used in the zoning of microfossils such as spores and pollens [46]. Analysis was performed with Tilia 2.6.1 [47].
We counted the species richness of the two main subfamilies, Karakoromyinae and Tataromyinae, of the Ctenodactylidae during the period of 34.9–30.6 Ma, including unpublished new materials and new species of Tataromyinae. We used species richness as our metric of diversity and plotted diversity curves based on the range-through assumption at a chronologic resolution of 0.2 million years [48].
3.6. Institutional Abbreviations
The following institutions were involved in this research: AMNH, American Museum of Natural History (New York); IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (Beijing); and PIN, Paleontological Institute, Russian Academy of Sciences (Moscow).
4. Systematic Paleontology
Rodentia Bowdich, 1821 [49]
Ctenodactyloidea Tullberg, 1899 [50]
Ctenodactylidae Gervais, 1853 [51]
Karakoromyinae Wang, 1994 [52]
Karakoromys Matthew and Granger, 1923 [18]
Type species: Karakoromys decessus Matthew and Granger,1923 [18]
Included species: Karakoromys arcanus (Shevyreva, 1971) [53], K. chelkaris (Shevyreva, 1971) [53], K. conjunctus, new species.
Geological age: latest Eocene to Early Oligocene.
Geographic distribution: Saint Jacques, Qianlishan District, Ulantatal (including Kekeamu), Nei Mongol; Burqin Basin, Xinjiang, China; Shunkht, Boongeen Gol; Ulan Ganga, Khatan Khayrkhan; Valley of Lakes, Mongolia; Zaisan Basin, Shalkarteniz (Chelkar-Teniz Lake); Kyzyl Kak, Kazakhstan.
Emended diagnosis: Small-sized ctenodactylids. Cheek teeth brachydont and wider than Tataromyinae; main cusps on upper molars distinct, hypocone equal or close in size to protocone, enlarged metaconule merging into developed metaloph, metaloph more obliquely forward pointing toward protocone or sinus on M2 and M3. The connection between metaloph and hypocone absent to present, longitudinal entoloph usually low and short, V-shaped sinus nearly symmetrical and transverse; on lower molars, trigonid equal to or longer than talonid, straight posterior arm of protoconid extending directly from protoconid at a large angle to metalophid I (greater than 30° or even 45°), lingual end of the posterior arm of protoconid absent, present or developed and joining metaconid, trigonid basin open to closed, entoconid conical with weak arm of entoconid, transversely elongated hypoconulid with long lingual tail, hyposinusid short and shallow with steep bottom, m3 trigonid obviously wider than talonid with reduced hypoconid and hypoconulid, hyposinusid usually reduced or absent, and the absence of hyposinusid resulting in continuous ridge fused by hypoconid and hypoconulid.
Remarks: Karakoromys has long been considered to include only one species, K. decessus. Wang [14,52] suggested that the genera Terrarboreus and Woodomys, established by Shevyreva [53], should be considered as synonyms of Karakoromys. Additionally, the scholar that the species, Terrarboreus arcanus, Woodomys chelkaris, and W. dimetron, should be reclassified as Karakoromys decessus. The recent discovery of new material from Ulantatal brings our attention back to the two genera established by Shevyreva [53]. Terrarboreus and Woodomys share many similarities with Karakoromys, such as the presence of transverse sinus and longitudinal entoloph on the upper molars and location of the lingual opening of the trigonid basin on the lower molars. We agree with Wang’s suggestion [14,52] that these two genera are junior synonyms of Karakoromys. However, there are some notable differences at the species level. For instance, the metaloph on the upper molars of T. arcanus is incomplete and points towards the protocone, which is markedly different from that of K. decessus, as emphasized by Dashzeveg and Meng [15]. Combined with other characteristics of our new materials (see the comparison below for details), we consider the species to be valid and revise it to Karakoromys arcanus. Similarly, Woodomys chelkaris has distinct characteristics, such as a very low tooth crown height on lower molars, a short posterior arm of protoconid that extends posteriorly without a lingual end and the conical hypoconid and entoconid being equal in size on the p4, and the complete absent of a hyposinusid on the m3. These characteristics are obviously different from those of Karakoromys decessus. Additionally, Woodomys chelkaris is also obviously smaller than K. decessus (Table 1, Figure 3). Therefore, we consider Woodomys chelkaris to be a valid species different from Karakoromys decessus and revise it as K. chelkaris. However, W. dimetron named by Shevyreva [54] is consistent in both morphology and size with Karakoromys decessus, so is therefore considered to be a junior synonym of K. decessus. Based on the new materials from Ulantatal, Karakoromys can be divided into four species: K. decessus, K. arcanus, K. chelkaris, and K. conjunctus sp. nov. The Karakoromys chelkaris is significantly smaller than the other Karakoromys species. Karakoromys decessus is marked by the complete metaloph, which connects the hypocone on upper molars, and by the posterior arm of the protoconid, which possesses a short lingual end on lower molars. Karakoromys arcanus is characterized by the incomplete metaloph on M1-2 and the absence of the lingual end of the posterior arm of protoconid on lower molars. Karakoromys conjunctus differs from K. decessus or K. arcanus in that the connection between the metaloph and hypocone is more developed, the entoloph is longer and higher on upper molars, and the lingual end of the posterior arm of protoconid bends towards or connects to the metaconid, forming a semi-closed or closed trigonid basin on lower molars.

Table 1.
Summary statistics of tooth measurements of Karakoromys species.
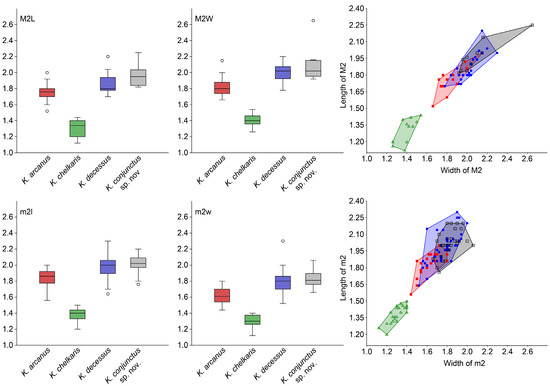
Figure 3.
Boxplots and bivariate graphs of the length and width of the M2 and m2 for each species of Karakoromys. Abbreviations: M2L, length of M2; M2W, width of M2; m2l, length of m2; width of m2. Red squares represent K. arcanus; green triangles represent K. chelkaris; blue dots represent K. decessus; grey squares represent K. conjunctus sp. nov. The colors in the boxplots correspond to the colors of each species in the bivariate graphs. The boxplots were plotted in Past 4.11 [45] with “Outliers” and “Interpolation” ticked. The 25–75 percent quartiles are drawn using a box. The median is shown with a horizontal line inside the box. The whiskers are drawn from the top of the box up to the largest data point less than 1.5 times the box height from the box (the “upper inner fence”), and similarly below the box. Values outside the inner fences are shown as circles (“outliers”). The measurements are in mm.
4.1. Karakoromys decessus Matthew et Granger, 1923 [18]
(Figure A1, Figure A2, Figure A3 and Figure A4, Table S1)
Karakoromys decessus: Matthew and Granger, 1923: 6–7, Figure 7. Wood, 1977: 125, Figure 3P.
Karakoromys decessus (partim): Kowalski, 1974: 166–167, pl. XLIX, Figures 3, 4 and 7 [20]. Wang, 1994: 38–40, Figure 4A,C [52]. Wang, 1997: 49–54, Figures 30 and 32A,C [14]. Höck et al., 1999: 115–117, Figure 20/4 [27]. Vianey-Liaud et al., 2006: 121–123, Figure 5.1 [31]. Schmidt-Kittler et al., 2007: 175–180, 212, Figures 3 and 4. Pl. 1: Figure G–J [23]. Daxner-Höck et al., 2017: Figure 45a–i [25].
Woodomys dimetron: Shevyreva, 1994. 116, Figure 11 [54].
Holotype: AMNH 19070, a pair of mandibles with both left and right p4–m3 and left incisor; Hsanda Gol Formation, red beds, Loh, Tsagan Nor basin, Mongolia; Early Oligocene.
Referred specimens: IVPP V 32087–32133, 107 specimens (see Table S1).
Locality and Horizon: KM and SJ section and lower part of UTM of Ulantatal area, Alxa Zuoqi, Nei Mongol, China; The lower part of the Ulantatal Formation (level 20–56 m).
Geological age: early Early Oligocene (33.3–30.9 Ma).
Emended diagnosis: On M1/2, metaloph connecting with hypocone but weak, lingual end of mesosinus bending backward but not united with posterosinus, longitudinal entoloph low and short. On DP4, metaloph connected with hypocone. On lower molars, lingual end of the posterior arm of protoconid pointing straight to the lingual side and neither bending towards nor connecting with the metaconid, trigonid basin open lingually, arm of entoconid transversely connecting to the midpoint of the ectolophid or slightly behind.
Measurements: see Table 1.
Description:
Small ctenodactylid. The incisive foramen is large, the zygomatic process of the maxilla is anterior to P4, the palate is wide and its width between two dental rows is wider than the width of the cheek teeth. The mandible is sciurognathous, the ventral masseteric crest extends forward to below m1, and the mental foramen is located below p4. Dental formula 1.0.1.3/1.0.1.3, but a juvenile individual has two upper deciduous teeth (DP3 and DP4) and a lower one (dp4). Its bilateral dentitions are parallel, with brachydont and relatively wide cheek teeth. P4 possesses two roots, a smaller anterobuccal root and a larger posterolingual one. DP4 and M1–3 all possess three roots: a large lingual root and two small buccal roots. DP3 has a single root. Both dp4 and p4–m3 have two roots. Specimens V 32127 and V 32128 both preserve occluded maxillae and mandibles, providing direct evidence for matching of the upper and lower dentition of K. decessus.
The P4 has an oval outline, with conical protocone and paracone. The protocone is slightly larger and lower than the paracone. The protoloph is complete and high, while the metaloph is usually incomplete and low, with the lingual end being either free (3/16) or connected to the posteroloph (12/16). The posterior arm of protocone connects the posteroloph (9/16) or does not (7/16). The anteroloph is generally absent (11/14); when present, it is low and weak. The posteroloph is weak and lacks a lingual part.
The M1 is nearly square in its occlusal surface, and its width is either equal to or slightly wider than its length. The four main cusps are distinct. The V-shaped protocone is slightly larger than the hypocone. The conical paracone is slightly anterior to the protocone. The nearly transverse protoloph is low and narrow in the middle. The metaconule merges into the metaloph, although it does without any obvious cusp contour and it is still recognizable. The metaloph weakly connects with the hypocone. The hypocone is slightly anteroposteriorly compressed and located more lingually than the protocone. The entoloph is low and short. The anteroloph is much lower than the protocone and there is no distinct anterocone. The anterior groove is weak, and the posterior groove is not obvious. The anterosinus and mesosinus are obviously longer than the posterosinus. The lingual end of the mesosinus bends posteriorly but does not join the posterosinus. The transverse sinus is almost symmetrical, V-shaped, and opposite to the mesosinus. The direction of striations on the enamel is anterolingually oblique, indicating the direction of mastication.
The M2 is almost the same as M1 in overall morphology, except for minor differences: it is slightly larger, has a squarer crown profile (still slightly wider than long), longer protoloph and metaloph, more transverse protoloph, more aligned hypocone and protocone on the lingual side, larger opening of the anterosinus, and wider and longer mesosinus. Its anterior groove is less obvious or absent, and it has a weak posterior groove.
The M3 is similar to M1/M2 in morphology, but smaller in size. The occlusal outline has much shorter posterior side than the anterior one due to its possession of a much smaller and less lingually located hypocone. The posteroloph and posterosinus are both shorter. The posterior groove is somewhat distinct, and the sinus is shorter, albeit with a larger opening.
The DP3 has single conical cusp and is much smaller in size than DP4. The tip of DP3 is situated lower than the anteroloph of DP4.
The DP4 is molariform and generally similar to the upper molars. The lingual side is obviously shorter than the buccal one, the anteroloph is isolated and low, the main cusps are more conical, and the entoloph is lower and shorter.
The p4 is longer than it is wide. The posterior arm of protoconid is either connected (6/15) or not connected (9/15) to the metaconid. The longitudinal ectolophid is either low and weak (12/17) or absent (5/17). In the former case, it is located centrally (9/12), or slightly buccally in the latter (3/12). The hypoconid is usually present (14/15) and smaller than the entoconid, and usually conical (10/14), but several have low transverse ridges (4/14). The hypoconulid is reduced to a weak protrusion on a transverse ridge between entoconid and hypoconid (4/14), or it is absent (10/14).
On the m1, the trigonid is obviously longer and slightly narrower than talonid. The anterior cingulum is absent. The metaconid is conical. The straight posterior arm of protoconid extends directly from protoconid at a large angle to the metalophid I (greater than 30°). The lingual end of the posterior arm of the protoconid points straight to the lingual side, neither bending towards nor connecting with the metaconid, so that the trigonid basin opens lingually. The lingual end of the posterior arm of protoconid can be long, short, or occasionally absent. The ectolophid extends obliquely from the lingual part of the posterior arm of protoconid to the hypoconid. The entoconid is conical and is nearly transversely connected to the midpoint of the ectolophid (10/22) or slightly behind (12/22). The hypoconid is slightly compressed anteroposteriorly, with a somewhat forward buccal end. The hypoconulid is a transversely elongated crescent with a longer transverse lingual tail. The short and weak arm of the hypoconulid connects to the hypoconid more buccally. The sinusid is deeper and larger than the mesosinusid or posterosinusid. The hyposinusid is short and shallow. In moderately abraded specimens, the buccal main cuspids are more heavily abraded than the lingual main cuspids, resulting in the metaconid and entoconid tending to be higher than the protoconid and hypoconid. On the occlusal surface, there is a wear groove extending obliquely from the mesosinusid to the hyposinusid.
The m2 is similar to m1 in morphology, but it is larger. The trigonid and talonid have roughly equal width, and the lingual end of the posterior arm of protoconid is always more developed and longer than that in m1. The arm of hypoconulid is slightly higher than that in m1, and the hyposinusid is shorter.
The m3 has a talonid that is obviously narrower than the trigonid, with a short arm of entoconid, less transversely expanded hypoconid, and a significantly reduced conical hypoconulid. The hyposinusid is either short and shallow or not obvious at all.
The dp4 is molariform and similar to the lower molars in basic form. It is smaller in size than molars, with a narrower trigonid compared to the talonid. The main cuspids are more distinct and the lophids are lower. In some specimens, the entoconid is isolated.
Remarks:
The holotype of K. decessus (AMNH 19070) is a pair of mandibles from the Hsanda Gol Formation [18], and no occluded maxilla and mandible were found for a long time. The new materials of the maxillae associated with the mandibles (V 32127 and V 32128, Figure A1) from Ulantatal confirm the existence of K. decessus in this region and demonstrate that its upper molars have significantly different morphologies from those attributed to K. arcanus. Specifically, the metaloph of K. decessus is complete and weakly connected to the hypocone, whereas the metaloph of K. arcanus is incomplete and does not connect to the hypocone. Additionally, some specimens (V 10576) from Kekeamu previously classified as K. decessus by Wang [14], with an incomplete metaloph, clearly do not belong to K. decessus and should be reassigned to K. arcanus.
A mandible fragment (V 13558) from the lower part of the Paoniuquan Formation in the Danghe area of Gansu, China, was identified by Wang and Qiu [55] as Karakoromys decessus. Its teeth are more lophodont, the posterior arm of protoconid does not extend posteriorly, and the trigonid is significantly shorter than the talonid, which is obviously different from Karakoromys. Though slightly wider, its overall morphology is similar to that of Tataromys parvus; therefore, we assigned it as Tataromys cf. parvus. Additionally, two molars from the HY1203 locality of the Hongyazi area in western Gansu, China, were identified as Karakoromys decessus and Karakoromys sp., respectively [56]. The former (V 18883) has an ectolophid extending anterobuccally to the protoconid, which is obviously different from Karakoromys. The latter (V 18884) has an ectolophid located slightly buccally, the arm of the entoconid is weak, the lingual end of the hypoconulid is more anteriorly located, and the opening of the posterosinusid is narrow, which is unlike that of Karakoromys. The accurate classification of these two specimens requires further research in the future.
4.2. Karakoromys arcanus (Shevyreva, 1971) [53]
(Figure A5 and Figure A6, Table S1)
Terrarboreus arcanus Shevyreva, 1971: 81–83, Figure 7 [53]. Wood, 1977: 123, Figure 2Q [57].
Woodomys chelkaris (partim): Shevyreva, 1971: 83–85, Figure 8e [53].
Karakoromys decessus Wang et al., 1981: 28–29 [28].
Karakoromys decessus (partim): Wang, 1994: 38–40, Figure 4B [52]. Wang, 1997: 49–54, Figures 31 and 32B [14]. Vianey-Liaud et al., 2006: 122, Figure 5.3 [31]. Schmidt-Kittler et al., 2007: 175–180, Figures 6 and 7 [23]. Daxner-Höck et al., 2010: 353–354, 358–359, Figure 3.1 [24].
Karakoromys cf. decessus (partim): Wang and Wang, 1991: 67–70 [30].
?Karakoromys decessus Huang, 1985: 36–37 [29].
Holotype: PIN, no. 478/386, skull fragment with a part of the zygomatic arch and a complete row of upper cheek teeth; Chelkar-Teniz Lake (=Shalkarteniz), Yrgyz District, Aktobe Region, Kazakhstan; Early Oligocene.
Referred specimens: IVPP V 32058–32086, 72 specimens (see Table S1).
Locality and Horizon: KM and SJ section of Ulantatal area, Alxa Zuoqi, Nei Mongol, China; the lower part of the Ulantatal Formation (level 0–46 m).
Geological age: latest Eocene to early Early Oligocene (34.9–31.4 Ma).
Emended diagnosis: The metaloph on M1/2 is incomplete and points to either sinus or protocone. The mesosinus joins the posterosinus to form a U-shaped valley. On DP4, the metaloph is also incomplete. On lower molars, the posterior arm of protoconid smoothly connects with ectolophid lacking a lingual end, and there is a wide lingual opening of the trigonid basin (trumpet-shaped opening). The arm of entoconid usually extends obliquely forward and connects with the anterior part of the ectolophid.
Measurements: see Table 1.
Description:
Small ctenodactylid. The structures of maxilla and mandible are similar to K. decessus. The dental formula and basic teeth form are the same as thaose of K. decessus, but some stable features distinguish them from K. decessus. The root pattern of cheek teeth is the same as that of K. decessus.
The P4 has an oval outline, with a conical protocone and paracone. The protoloph is complete and high, whereas the metaloph, which is low, is incomplete and not connected to the posteroloph. The posterior arm of protocone joins the posteroloph (1/5) or does not (4/5). The anteroloph is absent, and the posteroloph is weak without a lingual part.
The M1 is nearly square and slightly wider than it is long. The main cusps are distinct. The protocone is equal to or slightly larger than hypocone. The paracone is conical, slightly anteriorly positioned and smaller than the protocone. The transverse protoloph is complete and slightly lower in the middle. The metaconule merges into the developed metaloph without obvious cusp contour; however, it still has an enlarged body at its location and is slightly larger than the metacone. The metaloph is incomplete and not connected to the protocone or hypocone, pointing to the sinus or protocone. The hypocone is more lingually situated than the protocone. The entoloph is short and low. The anteroloph extending from the protocone is obviously lower than the protocone and usually lacks a distinct anterocone. Both the anterior and the posterior grooves are not obvious. The anterosinus reaches or slightly exceeds half of the tooth width. The mesosinus joins the posterosinus to form a continuous U-shaped valley. The sinus is transverse and nearly symmetrically V-shaped. Both the direction of striations on the enamel band and the wear groove indicates a posterobuccal–anterolingual chewing direction.
The M2 has the same overall structures as M1, but displays some differences. It is larger in size, and the protoloph is more transverse. The metaloph is longer and more oblique, pointing towards the protocone. The hypocone is slightly more buccally situated and is aligned with the protocone on the lingual side, and the mesosinus is longer.
The M3 is equal to or smaller than M2 in size; it is similar to M2 in basic form, but different from M1 or M2 in the following ways: the posterior side of crown is shorter than anterior one due to the reduced hypocone and the inwardly receding metacone; the paracone is slightly backward and transversely aligned with the protocone; a weak connection occasionally exists between the metaloph and the hypocone; the posterosinus and posteroloph are both shorter and the latter extends more posteriorly with a cusplet shape; the posterior groove is slightly distinct; and the sinus is shorter with a wider opening.
The DP3 is smaller than DP4 and has a single conical cusp.
The DP4 is molariform and wider than it is long, being similar in basic form to the upper molars. However, it differs from the upper molars in that the buccal side of the crown is significantly longer than the lingual one, the main cusps are more conical, and the entolophs are lower and shorter, respectively. The anteroloph is isolated and very low, and the metaloph is more transverse. Additionally, the metaloph is incomplete.
The p4 is slightly longer than it is wide. The protoconid and metaconid are conical. The posterior arm of protoconid is connected with the metaconid, and the longitudinal ectolophid is low and located slightly lingually. The hypoconid and entoconid are conical or ridged. The hypoconulid is usually reduced to a small protrusion on the transverse ridge between the entoconid and the hypoconid, or is absent. However, in specimen V 32072.11 (Figure A6g), the hypoconulid of p4 is distinct and isolated, located behind the entoconid and hypoconid, and slightly lower than the latter two cuspids (similar to the case in IVPP V 10576.25).
On m1, the trigonid is equal to or slightly shorter than the talonid and slightly narrower than talonid. The main cuspids are distinct. The anterior cingulum is absent. The metaconid is conical. The straight posterior arm of protoconid extends directly from protoconid at a large angle to the metalophid I (greater than 30°) and is connected with the ectolophid without a lingual end, resulting in a lingual side of the trigonid basin that is a wide trumpet-shaped opening. The entoconid is also conical. The arm of the entoconid usually connects to the anterior part of the ectolophid. The hypoconid is slightly compressed anteroposteriorly, with a slightly forward buccal end. The hypoconulid is a transversely elongated crescent with a longer transverse lingual tail. The arm of hypoconulid is short and weak, with a slightly buccal connection to the hypoconid. The sinusid is deeper and larger than the mesosinusid or posterosinusid. The hyposinusid is short and shallow with a steep bottom. Finally, there is a wear groove on the occlusal surface extending from the mesosinusid to the hyposinusid.
The m2 is basically the same as m1 except for the following differences: the size is larger, the trigonid is as wide as the talonid and the width-to-length ratio of tooth is larger, the arm of hypoconulid is slightly higher, the hyposinusid is shorter.
The m3 is similar to m2 in basic form and size, but different from m1 or m2 in the following respects: the talonid is obviously narrower than trigonid; the entoconid is smaller and more conical; the arm of entoconid is so short that entoconid is close to the ectolophid; the arm of hypoconulid is usually developed and high; the hyposinusid is usually reduced or occasionally absent; and when absent, the hypoconid and hypoconulid merge as one continuous ridge.
The dp4 is molariform, similar to lower molars in basic form. The size is much smaller, the trigonid is significantly narrower than talonid, the main cuspids are more distinct with weak lophids, the arm of entoconid is weak or absent, and when present, it is transversely connected to the hypoconid.
Remarks:
The holotype of K. arcanus (PIN, no. 478/386) is a skull fragment with upper cheek teeth [53]. Mandibles and maxillae of matching size collected from the same strata in Ulantatal have been included in this species, although no occluded specimens have been found. The mandibles of K. decessus, K. conjunctus and K. chelkaris can be easily excluded from the candidate mandible of K. arcanus, as K. decessus and K. conjunctus both have occluded mandibles and maxillae and as K. chelkaris is obviously smaller.
K. arcanus differs from K. decessus in several ways, including having a slightly smaller size (Table 1, Table 2 and Table 3, Figure 3). The incomplete metaloph on M1/2 points to the sinus or protocone, and a mesosinus joins the posterosinus to form a continuous U-shaped valley. On DP4, the metaloph is incomplete. On lower molars, the posterior arm of protoconid lacks a lingual end and smoothly connects the ectolophid, meaning that the lingual side of the trigonid basin is a wide trumpet-shaped opening. The arm of entoconid usually extends obliquely forward, and its connection point with the ectolophid is usually anterior to the midpoint of the ectolophid.
Table 2.
Results of the one-way ANOVAs on tooth size. Significance at p ≤ 0.05 is indicated in bold. SS: sum of squares, MS: mean square, df: degree of freedom, M2: length of M2: M2W, width of M2, m2l: length of m2; m2w: width of m2.
Table 3.
Post hoc pairwise comparisons of tooth size between species of Karakoromys. Significance at p ≤ 0.05 is indicated in lightface font when both Fisher’s least-significant difference test (LSD) and Scheffe’s test are significant, in bold font when only Scheffe’s test is not significant, and in bold font with an underline when neither Fischer’s LSD test nor Scheffe’s test is significant. M2L, length of M2; M2W, width of M2; m2l, length of m2; m2w, width of m2.
4.3. Karakoromys chelkaris (Shevyreva, 1971) [53]
(Figure A7 and Figure A8, Table S1)
Woodomys chelkaris (partim): Shevyreva, 1971: 83–85, figure 8a–d [53]. Wood, 1977: 125, figure 3Q [57].
Karakoromys decessus (partim): Wang, 1994: 38–40, figure 4D [52]. Wang, 1997: 49–54, figure 32D [14]. Schmidt-Kittler et al., 2007: 175–180, figure 2 [23].
Holotype: PIN, no. 478/387, left mandible with incisor and p4-m2; Chelkar-Teniz Lake (=Shalkarteniz), Yrgyz District, Aktobe Region, Kazakhstan; Early Oligocene.
Referred specimens: IVPP V 32134–32166, 50 specimens (see Table S1).
Locality and Horizon: KM and SJ section of Ulantatal area, Alxa Zuoqi, Nei Mongol, China; The lower part of the Ulantatal Formation (level 0–49 m).
Geological age: latest Eocene to early Early Oligocene (34.9–31.2 Ma).
Diagnosis: The smallest species of Karakoromys with lowest cheek tooth crown height. On P4, posterior arm of protocone absent. On upper molars, incomplete metaloph usually transversely pointing to hypocone or slightly anteriorly oriented, mesosinus uniting with posterosinus to form U-shaped valley, entoloph short and low, transverse sinus U-shaped or blunt V-shaped. On M3, metaloph oblique and thinner with its height lower than hypocone. On p4, posterior arm of protoconid short and extend more posteriorly, lingual end of the posterior arm of protoconid absent, conical hypoconid equal in size to entoconid. On lower molars, larger trigonid basin nearly as deep as mesosinusid, arm of entoconid usually thin and weak. On m3, conical entoconid usually elongated anteroposteriorly, hyposinusid absent, hypoconid and hypoconulid merging into one continuous ridge.
Measurements: see Table 1.
Description:
It is the smallest and has the lowest tooth crown height among all species of Karakoromys, with lower cusps (cuspids), lophs (lophids) and shallower sinuses (sinusids). The structures of maxilla and mandible are similar to those of K. decessus. The root pattern is the same as that of K. decessus, except for the cases of DP3 and DP4, which find no fossil.
The P4 has an oval outline with a conical protocone and paracone. The protoloph is low. The metaloph is low and incomplete, extending toward but not joining the posteroloph. The posterior arm of protocone and the anteroloph are both absent, and the posteroloph is weak without a lingual part.
The M1 is nearly square in shape and its width is equal to (5/11), slightly wider (4/11) or occasionally slightly narrower (2/11) than its length. The main cusps on M1 are distinct. The hypocone is equal to the protocone in size and the former is slightly located towards the lingual side. The paracone is conical, slightly anteriorly located and smaller than the protocone. The protoloph is slightly oblique to the anterobuccal side. The metaconule merges into the developed metaloph without any obvious cusp contour, and it is lower than the metacone. The metaloph is incomplete without connecting to the protocone or hypocone, and usually transversely points to the hypocone (5/8) or slightly forward (3/8). The entoloph is short and low. The anteroloph extending from the protocone is obviously lower than the protocone or protoloph and usually does not have a distinct anterocone. The anterior groove is weak, and the posterior groove is not obvious. The anterosinus reaches or slightly exceeds half of the tooth width. The mesosinus unites with the posterosinus to form a continuous U-shaped valley. The transverse sinus is U-shaped or blunt V-shaped, and nearly symmetrical. A shallow oblique wear groove extends from the posterosinus to the sinus and the direction of the wear groove is consistent with the one of striations.
The M2 is larger than M1. Both the protoloph and metaloph are longer and the former is more transverse; the metaloph is more obliquely forward, pointing toward the protocone (6/13) or sinus (7/13). The hypocone slightly retreats toward the buccal, being aligned with the protocone on the lingual side, and the mesosinus is longer than that in M1.
The M3 is usually around the same size as M2 (2/10) or slightly smaller (7/10); occasionally, it is even smaller than M1 (1/10). Its shape is similar to that of M1 or M2, but with a shortened posterior side. The hypocone is obviously smaller than protocone and located further toward the buccal side. Both the protoloph and metaloph are thinner and lower than that on M2. This especially the case for the latter, which is even lower than that of the hypocone. The metaloph is also more oblique, resulting in the formation of a larger angle with the protoloph; conversely, the entoloph is short and weak with a reduced sinus. The mesosinus and the posterosinus are wide and shallow, and the posteroloph is shorter and extends more posteriorly than that on M2.
The p4 is slightly longer than it is wide. The protoconid and metaconid are conical and isolated from each other, whereas the entoconid and hypoconid are slightly compressed anteroposteriorly and connected by a weak transverse ridge. The posterior two cusps are significantly smaller and lower than the anterior two. The posterior arm of protoconid extends more posteriorly and lacks a lingual end connecting to the metaconid (13/13). The nearly longitudinal ectolophid is absent (1/13) or thin and weak (12/13) connected to the hypocone and located slightly buccally. The hypoconulid is usually absent (12/13) and occasionally extends transversely from the hypoconid in a ridge shape (1/13).
On m1, the trigonid is equal to (3/19) or slightly longer than the talonid (16/19). The anterior cingulum is absent. The metaconid and entoconid are distinct. The straight posterior arm of protoconid extends directly from the protoconid at a large angle to the metalophid I (greater than 30° or even 45°), with (4/18) or without a lingual end (14/18). The trigonid basin opens lingually, and the ectolophid is long and centered, extending from the lingual end of the posterior arm of the protoconid to the hypoconid in the anterolingual–posterobuccal direction. The entoconid is conical. The arm of entoconid is usually thin and weak, joining slightly obliquely to the midpoint of the ectolophid (8/19), or nearly transversely behind the midpoint of the ectolophid (11/19). The hypoconid is slightly compressed anteroposteriorly, with a slightly forward buccal end. The hypoconulid is a transversely elongated crescent with a longer transverse lingual tail and which occasionally has a shorter buccal tail. The short and weak arm of hypoconulid connects to the hypoconid in more buccally. The sinusid is equal to or slightly larger than the mesosinusid, and both are wide and shallow. The trigonid basin opens posterolingually to the mesosinusid. The bottom of the trigonid basin is connected with the bottom of the mesosinusid. The posterosinusid is relatively shallow, with a wide lingual opening. The hyposinusid is short and shallow with a steep bottom. On some moderately abraded specimens, there is a very shallow and wide wear groove on the occlusal surface extending from the mesosinusid to the hyposinusid on some moderately abraded specimens.
The m2 is larger than m1. The length of the trigonid is either equal to (15/24) or slightly shorter than the talonid (9/24), and its width is as wide as the talonid. The trigonid basin is large and deep, being almost as deep as the mesosinusid. The lingual end of the posterior arm of protoconid is usually developed (18/24) and occasionally absent (6/24). The arm of entoconid is usually thinner and weaker (22/24): sometimes it is absent (2/24); the arm of entoconid extends transversely (3/22), more obliquely forward in such a way as to connect to the midpoint of the ectolophid (4/22), or more forward (11/22), and even connects to the lingual end of the posterior arm of protoconid (4/22). Additionally, the arm of hypoconulid is more developed and higher.
The m3 has an obviously narrower talonid than trigonid, and the small entoconid is usually elongated anteroposteriorly with anterior and posterior accessory crests (12/17). The arm of entoconid is reduced to a low, slender, weak ridge or line connecting to the front of ectolophid (4/17), to the lingual end of the posterior arm of protoconid (5/17), or is completely absent (8/17). The hypoconid is shortened transversely, and the hypoconulid is reduced. The hyposinusid is completely absent while the arm of the hypoconulid is developed, meaning that the hypoconid and hypoconulid merge into one continuous ridge (17/17). One specimen known as V 32160.2 (Figure A8h) has a long ridge that protrudes from the protoconid to the lingual side in a way that is anterior to the metalophid I on m3; which may be the anterior cingulum, the occurrence of which is accompanied by a longer anterosinusid.
The dp4 is molariform, which is similar to lower molars in its basic form, the main differences being that the tooth is much narrower and significantly smaller, the lophids are much lower, and protocone is slightly larger than hypocone.
Remarks:
The holotype of K. chelkaris (PIN, no. 478/387) is only a fragmentary mandible [53], and no specimens of occluded upper and lower jaws have been found. However, some maxillae have been assigned to this species. The upper and lower molars here assigned to K. chelkaris are consistent in size and morphology, with both being significantly smaller than those of the other three species of Karakoromys. Additionally, they both show the lowest crowns and weakest lophs (lophids) in Karakoromys.
K. chelkaris shares certain features with Gobiomys, including a low metaloph extending obliquely forward on M3, a weak arm of the entoconid on the lower molars, and an entoconid longitudinal extension with anterior and posterior accessory crests on m3 without a hyposinusid. However, K. chelkaris obviously has the synapomorphies of Karakoromys, including the presence of entoloph, the metaconule merging into the metaloph on upper molars, the developed ectolophid extending anterolingually, and the arm of entoconid, which usually extends obliquely forward and connects to the ectolophid on lower molars. In contrast, the features of Gobiomys, such as the weak or absent entoloph, a distinct metaconule on upper molars, and short and weak ectolophid on lower molars, remain relatively primitive. Therefore, K. chelkaris is classified as Karakoromys rather than Gobiomys.
K. chelkaris differs from K. decessus and K. arcanus in that the size is significantly smaller (Table 1, Table 2 and Table 3, Figure 3). On P4, the posterior arm of protocone is absent. On M3, the protoloph and metaloph are thinner and lower, especially the latter, and they are even lower than the hypocone. The metaloph is also more oblique, meaning that it forms a larger angle with the protoloph. On p4, the lingual end of the posterior arm of protoconid is absent. On the lower molars, the arm of the entoconid is thinner and weaker. The trigonid basin is larger and deeper, almost as deep as the mesosinusid. On m3, the entoconid is a smaller cone that is usually elongated anteroposteriorly, with anterior and posterior accessory crests. The hyposinusid is absent, and the hypoconid and hypoconulid merges into one continuous ridge. Additionally, K. chelkaris differs from K. decessus in that the metaloph is incomplete on upper molars, and is different from K. arcanus in that the lingual end of the posterior arm of protoconid is present.
4.4. Karakoromys conjunctus sp. nov.
(Figure A9 and Figure A10, Table S1)
Holotype: IVPP V 32180, a broken skull associated with mandibles; SJ section of Ulantatal area, Alxa Zuoqi; Nei Mongol, China; the lower part of the Ulantatal Formation, Early Oligocene.
Paratype: IVPP V 32181, a broken skull with mandibles (juvenile); V 32167.2, left maxilla associated with mandibles; V 32175.1, left maxilla with M1-3; V 32172, left mandible with p4–m3; V 32195.1, a pair of mandibles with complete dentition; V 32170, left mandible with p4 and m2–3; V 32178, left mandible with m1–2; V 32174.1, left mandible with m1–2; V 32169, left mandible with incisor and dp4–m2 (juvenile); and V 32194, left m2. Except for V 32195.1 from the top of KM section of Ulantatal area and V 32178 from the base of UTM section of Ulantatal area, the localities and horizons of other paratypes are the same as those seen for the holotype.
Referred specimens: IVPP V 32167.1, 32168, 32171, 32173, 32174.2, 32175.2–3, 32176–32177, 32179, 32182–32193, 32195.2; total 33 specimens (see Table S1).
Locality and Horizon: KM and SJ section and lower part of UTM of Ulantatal area, Alxa Zuoqi, Nei Mongol, China; the lower part of the Ulantatal Formation (level 30–60 m).
Geological age: early Early Oligocene (32.5–30.8 Ma)
Etymology: Conjunctus, Latin, connected. The name indicates that in species K. conjunctus, the lingual end of the posterior arm of protoconid connects or tends to connect to the metaconid.
Diagnosis: On upper molars, strong connection between metaloph and hypocone, entoloph longer and higher than that of other Karakoromys species, sinus U-shaped or blunt V-shaped. M3 more square than other species of Karakoromys due to relatively larger hypocone. On lower molars, lingual end of the posterior arm of protoconid developed and bending toward or connecting to the metaconid, leading to closed or semi-closed trigonid basin, arm of entoconid slightly obliquely or nearly transversely connecting to midpoint of ectolophid.
Measurements: see Table 1.
Description
The new species is close to Karakoromys decessus in size. The structures of maxilla and mandible are similar to K. decessus. The root pattern of cheek teeth is the same as that of K. decessus.
The P4 has oval outline with the conical protocone and paracone. The protoloph is complete. The metaloph is completely connected to the posteroloph. The posterior arm of protocone connects the posteroloph (2/3) or not (1/3). The anteroloph is absent, whereas the posteroloph is obvious, with a long buccal part but without a lingual part.
The M1 has nearly equal length and width. The main cusps are distinct. The protocone is equal to the hypocone in size. The paracone is conical and slightly smaller than the protocone. The protoloph is slightly oblique to the anterobuccal side. The enlarged metaconule merges into the developed metaloph with a low cusp that is slightly larger than that of the metacone. The metaloph is complete and usually strongly connected to the hypocone. The hypocone is slightly compressed anteroposteriorly and is more lingually located than the protocone. The entoloph is more developed, longer and higher than that of other Karakoromys species. The anteroloph extending from the protocone is lower than the protocone and usually without distinct anterocone. The anterior groove is weak, and the posterior groove is not obvious. The posterosinus is slightly shorter than the anterosinus, and the latter reaches or slightly exceeds half of the tooth width. The mesosinus is wide and long, and its lingual end bends posteriorly but does not join the posterosinus. The sinus is transversely U-shaped or bluntly V-shaped, short and wide, nearly symmetrical and is opposite to the mesosinus. The protocone and hypocone are usually lower than paracone and metacone after wear. A shallow oblique wear groove extends from the posterosinus to the sinus and the direction of the wear groove is consistent with the striations.
Compared to M1, M2 is larger, both the protoloph and metaloph are longer, the former is more transverse, and the connection between the metaloph and the hypocone is thicker and higher. The metaconule is more obvious and higher than that seen in M1, with an anterior edge protruding forward. The hypocone slightly retreats toward the buccal end, aligned with protocone on the lingual side. The entoloph is longer and higher than that in M1. The sinus becomes wider and shorter, showing a wide U-shape, the mesosinus is longer, and the anterior groove is less obvious, possessing a weak posterior groove.
M3 has a narrower posterior part. Its hypocone is more conical and slightly smaller than its protocone, and recedes toward the buccal side. The connection between the metaloph and hypocone is shorter and thinner than that seen in M1/2. The sinus is very short transversely; both the posterosinus and posteroloph are short, with the latter extending more posteriorly.
The DP3 is very small with a conical cusp.
The DP4 is molariform and wider than it is long. It is basically similar to the upper molars. The crown with the lingual side is obviously shorter than the buccal one; the main cusps are conical and the entoloph is short; the paracone is more anteriorly located than the protocone with more oblique protoloph; the anteroloph is isolated and low; the metaloph is curved but not connected to the posteroloph; and, without the metaconule, both the anterior groove and posterior groove are absent.
The p4 is longer than it is wide. The main cuspids are conical. The posterior arm of protoconid is connected with the metaconid (2/8) or not (6/8). The longitudinal ectolophid is weak and located slightly buccally (6/8) or slightly lingually (2/8). The hypoconid is developed (8/8) and smaller than entoconid, conical (7/8), or a low transverse ridge (1/8). The hypoconulid is reduced to a weak protrusion (2/8) or absent (6/8).
The m1 has a trigonid that is equal to (4/16) or slightly longer than the talonid (12/16), and narrower than the talonid. The anterior cingulum is absent. The metaconid and entoconid are conical. The straight posterior arm of protoconid extends directly from the protoconid at a large angle to the metalophid I (greater than 30° or even 45°). The posterior arm of the protoconid has a developed lingual end which bends toward (6/15) or connects with (9/15) the metaconid, resulting in a semi-closed (6/15) or closed (9/15) trigonid basin. The ectolophid extends obliquely from the lingual end of the posterior arm of protoconid to the hypoconid. The arm of entoconid is slightly obliquely (10/16) or nearly transversely (6/16) connected to the midpoint of the ectolophid. The hypoconid is slightly compressed anteroposteriorly with a somewhat forward buccal end. The hypoconulid is a transversely elongated crescent with a longer transverse lingual tail. The short and weak arm of the hypoconulid connects to the hypoconid more buccally. The sinusid is obliquely U-shaped and deeper and larger than the mesosinusid or posterosinusid. The hyposinusid is short and shallow, with a steep valley bottom. The lingual cuspids are higher than the buccal ones due to differential abrasion. On some moderately abraded specimens, there is a very shallow wear groove on the occlusal surface extending from the mesosinusid to the hyposinusid.
The m2, similar to m1, has a developed lingual end of the posterior arm of protoconid that bends or connects to the metaconid. However, m2 is larger than m1, and the trigonid is either equal to or slightly narrower than the talonid. Additionally, the lingual end of the posterior arm of protoconid more frequently connects to the metaconid (15/22), the proportion of the transverse arm of entoconid is higher than that in m1 (10/23), and the arm of hypoconulid is more developed and higher than that in m1.
The m3 is similar in morphology and size to m2, but differs from m1 and m2 in several aspects. The talonid is obviously narrower than the trigonid, the lingual end of the posterior arm of protoconid is short, and neither curved towards nor connected to the metaconid in a few specimens (5/18). The entoconid is smaller and more conical, and the arm of entoconid is often thin (11/17) but it can be thick in some cases (6/17). The hypoconid is shorter transversely, with a more forward buccal end. The hypoconulid is significantly reduced and more conical, with a shorter or absent transverse lingual tail. The hyposinusid is usually reduced or occasionally absent, and in cases where it is absent, the hypoconid and hypoconulid merge into one continuous ridge.
The dp4 is molariform and differs from the lower molars in that the size is much smaller, the trigonid is significantly narrower than the talonid with the close protoconid and metaconid, the main cuspids are more distinct with lower lophids, and the arm of entoconid connects obliquely forward to the ectolophid.
Remarks:
Three specimens, V 32180, V 32181, and V 32167.2, have occluded upper and lower jaws, providing reliable evidence for matching isolated upper and lower jaws (Figure A9a–c).
Although there are no apparent differences in size between K. conjunctus and K. decessus (Table 1, Table 2 and Table 3, Figure 3), there are some differences in morphology. K. conjunctus differs from K. decessus in that its upper molars have a more developed connection between the metaloph and hypocone, a longer and higher entoloph, a wide U-shape or blunt V-shape sinus, and a relatively square-shaped M3 with a larger hypocone. Its lower molars have a lingual end of the posterior arm of protoconid that bends towards or connects to the metaconid, forming a semi-closed or closed trigonid basin, and the arm of entoconid extends obliquely forward more often.
Compared to K. arcanus, K. conjunctus is slightly larger in size (Figure 3) and has stronger lophs or lophids. In particular, on P4, the metaloph is completely connected to the obvious posteroloph. On the upper molars, the entoloph is longer and higher, the metaloph is complete and strongly connects to hypocone, and the sinus is wider and shorter, showing a wide U-shape or a blunt V-shape. On lower molars, the trigonid is slightly longer than the talonid more often, and the lingual end of the posterior arm of protoconid bends toward or connects to the metaconid.
Compared to K. chelkaris, K. conjunctus is significantly larger (Table 1, Table 2 and Table 3, Figure 3), and the cheek tooth crown is higher, with more developed lophs (lophids). On P4, the metaloph is complete and the posterior arm of protocone is present. On upper molars, the metaloph strongly connects to the hypocone, and the entoloph is longer and higher. On M3, the tooth is relatively square, and the protoloph and metaloph are stronger and higher, with the latter being more transverse. On lower molars, the lingual end of the posterior arm of protoconid bends towards or connects to the metaconid, the arm of entoconid is stronger, and the trigonid basin is obviously shallower than the mesosinusid. On m3, the entoconid is more conical without anterior and posterior accessory crests, and the hyposinusid is usually present.
5. Discussion
5.1. Stratigraphic Distribution and Biochronological Significance of Karakoromys
The Karakoromys fossils found in the Ulantatal area in the past were primarily from the KM01 and UTL1 sites [30,31,32]. KM01 is situated at the bottom of the Ulantatal Formation, and UTL1 is located at the top of the lower part of the Ulantatal Formation (Figure 4). Huang [29] also reported a mandible of Karakoromys from the Ulantatal Formation, but did so without providing GPS coordinates or detailed stratigraphic information. Furthermore, no fossils were previously found in the 50 m-thick strata between KM01 and UTL1. Our new materials of Karakoromys come from the lower part of the Ulantatal Formation, with a composite profile interval of 0–60 m and have an age estimation of 34.9–30.8 Ma (Figure 4).
K. arcanus and K. chelkaris appeared before the Eocene–Oligocene Boundary (EOB), and spanned from 34.9 to 31.3 Ma (stratigraphic range of 0–48 m). On the other hand, K. decessus and K. conjunctus appeared after the EOB, with the former emerging slightly earlier, from 33.3 to 30.9 Ma (stratigraphic range of 20–56 m), and the latter arising slightly later, from 32.5 to 30.8 Ma (stratigraphic range of 30–60 m). The earliest occurrence of K. decessus (33.3 Ma) was very close to the EOB of 33.9 Ma, so its first appearance datum (FAD) can be roughly taken as a biochronological marker for the onset of the Oligocene in Asia. Additionally, K. decessus is widely distributed in the Early Oligocene strata of Central Asia, with its common occurrence ranging from 32.5 to 31.3 Ma (Figure 5h).

Figure 4.
The stratigraphic ranges of Karakoromys species from Ulantatal with magnetic polarity column. (a) GTS2004 [58]. (b–d) Simplified lithologic column with magnetic polarity sequence (partial) and schematic sections of the lower part of the Ulantatal sequence (modified from Wasiljeff et al. [38]). (e) Stratigraphic ranges of Karakoromys species from Ulantatal. Black dots represent the stratigraphic levels of Karakormys fossils. KM01 and UTL1 are fossil horizons of Ulantatal that were previously studied by Wang and Wang [30] and Vianey-Liaud et al. [31], respectively.
5.2. Karakoromys Diversity in Response to Climatic Change
5.2.1. The Diversity of Karakoromys and Related Taxa
In modern ecology, community diversity is measured based on species list and the population size of each species [59]. However, in paleontology and paleoecology, differences in fossil preservation and uneven sampling can lead to variation in the number of specimens collected from different time intervals or strata. This makes it difficult to directly compare the population size of various species among different strata. Relative abundance, which converts the number of individual of species into a percentage, eliminates the influence of different sample sizes among different strata on population size to some extent and is therefore widely used in deep-time evolution studies of ancient communities [60,61,62,63,64,65].

Figure 5.
The diversity of Karakoromys (including E. ampliatus) and climate change in Ulantatal during 34.9–30.5 Ma. (a) Global benthic δ18O record [5]. (b) The soil organic carbon (SOC) content. (c) Weathering intensity as indicated by the ratio of feldspars to quartz (Kf + Pl)/Q. (d) Mean annual precipitation (MAP). (b–d) All modified from Wasiljeff et al. [42], (b,c) reveal the changes in aridity of the Ulantatal area. (e) Number of specimens of Karakoromys (including E. ampliatus) through time for 0.4 Ma time bins. (f–j) The relative abundance (percentage of specimens) of each species in Karakoromys (including E. ampliatus) through time for 0.4 Ma time bins. (k) Cluster analysis of time bins. (l,m) are the lengths of M2 and m2, respectively. Light blue dots, K. arcanus; green dots, K. chelkaris; yellow dots, K. decessus; orange dots, K. conjunctus sp. nov.; gray dots, E. ampliatus. (n) Species richness of ctenodactylids with range-through assumption through time for 0.2 Ma time bins. n-1, Karakoromyinae (blue dots); n-2, Tataromyinae (orange dots); n-3, total (gray dots). (o) Schematic diagram of the diversity evolution of Karakoromyinae and Tataromyinae. The red arrows indicate the main changes in the ctenodactylids. The two blue horizontal bands mark global glaciation events Oi1 and Oi2, respectively, while the orange horizontal band marks the Early Oligocene Aridification Event (EOAE) in Central Asia [42,66,67,68,69].
Cluster analysis for community succession [46] suggests that the Karakoromys-Euryodontomys assemblage can be divided into four evolutionary stages (Figure 5k). During stage 1 (34.9–33.3 Ma), only K. arcanus and K. chelkaris are present, with the former being dominant. In stage 2 (33.3–32.5 Ma), K. decessus appears, but with a relatively low abundance, and K. chelkaris surpasses K. arcanus in relative abundance, becoming the dominant species. In stage 3 (32.5–31.7 Ma), K. conjunctus appears with a relatively low abundance. K. decessus becomes the dominant species, with a higher abundance than K. arcanus and K. chelkaris. In stage 4 (31.7–30.9 Ma), the relative abundance of K. arcanus and K. chelkaris is extremely low. However, E. ampliatus appears and quickly surpasses K. decessus and K. conjunctus, becoming the dominant species (Figure 5f–j).
Cluster analysis also reveals that the fourth stage forms a separate group, distinct from the previous three stages, (Figure 5k), indicating a significant change in the Karakoromys-Euryodontomys assemblage (from 31.7 to 31.3 Ma). This change is manifested by a shift in the dominant species at the genus level, with K. decessus decreasing and E. ampliatus increasing.
Body size is a fundamental ecological parameter of mammals and many studies have confirmed the correlation between tooth size and body size in mammals [70]. Tooth size analysis shows no significant change in Karakoromoyinae from 34.9 to 31.7 Ma, covering a period of three million years. However, after 31.7 Ma, there is a significant increase in size resulting from the emergence of larger-sized Euryodontomys, suggesting that the Karakoromoyinae may have undergone a significant ecological shift around that time (Figure 5l,m).
Diversity curves show that from 34.9 to 33.4 Ma, the species richness of Karakoromyinae and Tataromyiane remained stable without any obvious changes; from 33.4 to 33.0 Ma, the species richness of both Karakoromyinae and Tataromyiane increased significantly; from 33.0 to 31.8 Ma, the species richness generally remained stable, with only a small increase at 32.4 Ma; from 31.8 to 30.7 Ma, a significant turnover occurred, with the species richness of Tataromyinae increasing, while the species richness of Karakoromyinae decreased (Figure 5n,o).
5.2.2. The Possible Relationship between Karakoromys Diversity and Climatic Change
In evolutionary biology, it has long been hypothesized that both biotic (Red Queen hypothesis) and abiotic factors (Court Jester hypothesis) shape evolution [71,72]. Our extensive sampling and detailed chronologic data on ctenodactylids at Ulantatal over a time period of major climatic change provide an opportunity for exploring the relative contribution of the two hypotheses in the evolutionary history of this lineage.
The Eocene–Oligocene transition (EOT) occurred in two steps, with the second step (~33.8–33.6 Ma) representing the major ice-sheet expansion known as the Oi1 glaciation [67,73,74] and caused an expansion of open habitats in Asia [13,75]. However, there appears no extinction of Karakoromys species in Ulantatal during this significant global cooling event (Figure 5f,g,n). A possible explanation for the survival of Karakoromys and other ctenodactylids may lie in their ecology. Extant ctenodactylids live in the deserts of Africa and are adapted to arid environments [14,76]. Our previous research on the skull morphology of Oligocene ctenodactylids showed that Oligocene ctenodactylids may have had adaptations to arid and open environments [77], thus increasing their chance to survive the EOT cooling event.
From 33.4 to 33.0 Ma, there was a significant increase in diversity of the ctenodactylids, which postdated the EOT by 0.3–0.7 million years. Interestingly, there was also a delay of approximately 0.5 million years between the Post-Grande Coupure fauna and the Oi1 glaciation in the Hampshire Basin in Europe [12], and the faunal turnover in the Shapaja section in South America also lagged behind the Oi1 glaciation [78]. Although occurring slightly later than the EOT, the increase in diversity of ctenodactylids is possibly a response to the EOT, and the expansion of open habitats [13,75] may have provided new ecological niches for the diversification of ctenodactylids after the EOT.
Palaeoclimatic reconstruction of the Ulantatal area shows a stable, semi-arid regime during 33.0–31.8 Ma period, accompanied by a stable global mean temperature (Figure 5a–d). Despite the climatic stability, K. decessus surpassed K. chelkaris in relative abundance and became the dominant species during that period, suggesting a species interaction or other ecological processes at play.
An aridification event occurred in Ulantatal area around 31 Ma (Figure 5b,c) [42]. This was also recorded in Mongolia, and is referred to as the Early Oligocene Aridification Event (EOAE) [68,69]. Although research in the Lanzhou Basin southwest to the Ulantatal area did not emphasize this aridification event, its chemical weathering records show an obvious “cold-dry event” in the Basin around 31.7 Ma [74]. These records collectively suggest that the aridification around 31 Ma was likely a widespread event in Central Asia. The turnover of ctenodactylids during 31.8–30.8 Ma was well coupled with this aridification event (Figure 5b,c,n). Small and less lophodont Karakoromys species were replaced by larger and more lophodont Tataromyinae species. Higher degrees of lophodonty in the cheek teeth of rodents may have greater efficiency in grinding hard or rough foods [11,79,80] and are thus better adapted to arid environments. Another study on lagomorphs in Ulantatal also revealed a turnover of the Ordolagus lineage around 30.5 Ma [81], and there was also a record of a faunal turnover around 31 Ma in the Valley of Lakes, Mongolia [68]. Notably, E. ampliatus experienced a rapid population increase in the early stages of the aridification event, outnumbering K. decessus to become the dominant species of Karakoromyinae. This indicates that E. ampliatus had a better adaptation to arid environments than Karakoromys. However, as aridification intensified, the population of E. ampliatus also rapidly decreased and eventually went extinct together with Karakoromys after the peak of the aridification event around 30.8 Ma (Figure 5j).
From the responses of ctenodactylids to different climatic events, it appears that regional precipitation variation may have had a greater impact on the diversity of ctenodactylids than global temperature change. Palaeoprecipitation reconstruction showed that the mean annual precipitation (MAP) of the Ulantatal area during 34.9–30.8 Ma was around 400 mm (Figure 5d) [42], within the range of semi-arid regions [82]. Many ecological studies have shown that precipitation is the main controlling factor of ecosystems in semi-arid regions [83,84,85,86], which may explain why the aridification event around 31 Ma had a greater impact on the diversity of ctenodactylids in Ulantatal than the EOT cooling event.
In the evolutionary history of the ctenodactylids, the turnover at Ulantatal around 31 Ma seems to have been primarily influenced by environmental change, echoing the Court Jester hypothesis. However, during the long, stable semi-arid period between the EOT and the aridification event, biotic pressures (such as interspecific competition) may have been the main driver of community succession of ctenodactylids, which favors the Red Queen hypothesis [71,72]. Further explorations of the evolution dynamic need more data at larger scales.
6. Conclusions
Four species of Karakoromys, namely Karakoromys decessus, K. arcanus, K. chelkaris, and K. conjunctus sp. nov., were recognized from Ulantatal, showing the relatively high diversity of this most primitive ctenodactylids during the latest Eocene-Early Oligocene.
The chronological distribution of Karakoromys is at the latest Eocene to Early Oligocene. The first appearance datum (FAD) of K. decessus (33.3 Ma) is very close to the Eocene–Oligocene boundary (EOB) and is recommended as the biochronological marker for the beginning of the Oligocene in Asia.
Ctenodactylids survived the EOT cooling event without any extinction of the Karakoromys species. From 34.9 to 30.8 Ma, there were two distinct changes in the ctenodactylids fauna in the Ulantatal area: the first was an initial increase in diversity of ctenodactylids during 33.4–33.0 Ma after EOT, and the second was a notable turnover of ctenodactylids during 31.8–30.8 Ma coinciding with the regional aridification event in Central Asia. Regional precipitation variation in the semi-arid area may have had a greater impact on the diversity of ctenodactylids than global temperature change.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15060744/s1, Table S1: List of specimens; Table S2: Measurements of Karakoromys teeth (raw data).
Author Contributions
Conceptualization, Z.Z.; methodology, Z.Z., Q.L., B.W. and R.X.; validation, Z.Z. and Q.L.; formal analysis, Z.Z., Q.L., B.W. and R.X.; investigation, Z.Z., Q.L., B.W. and R.X.; resources, Z.Z. and Q.L.; data curation, Z.Z. and R.X.; writing—original draft preparation, R.X.; writing—review and editing, Z.Z., Q.L. and B.W.; visualization, R.X.; supervision, Z.Z. and Q.L.; project administration, Z.Z.; funding acquisition, Z.Z. and Q.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Strategic Priority Research Program of Chinese Academy of Sciences (XDB26000000), the National Natural Science Foundation of China (42072023, 41472003), and the Special Fund for Fossil Excavation and Preparation of the Chinese Academy of Sciences.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data generated during this study are included in this published article/supplementary material; further inquiries can be directed to the corresponding authors.
Acknowledgments
We would like to thank Wei Zhou, Ping Wang, Shijie Li, Qi Li, Yongfu Wang, Yongxing Wang, Yan Liu, Lihua Wang, Jian Wang, Haidan Ma, Lu Li, Zhongyun Liu, Fangyuan Mao, Haibing Wang, Yanxin Gong, Xinyue Zhang, Chenhui Jiao, Jiawen Liu, Qian Wang, Xiuqin Wang, Arya Farjand for their assistance in the fieldwork; Banyue Wang and Wuyang Xiong for providing some important references; Jian Wang for his helpful discussion; Wei Zhou and Shijie Li for preparation of the specimens. Finally, we would also like to thank the three reviewers for their constructive suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1, Figure A2, Figure A3, Figure A4, Figure A5, Figure A6, Figure A7, Figure A8, Figure A9 and Figure A10. Photographs of specimens with annotations.

Figure A1.
Karakoromys decessus from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a1–a4). IVPP V 32127, a fragment of skull associated with lower jaws (only dentitions are displayed): (a1) right P4–M3 (part); (a2) left P4–M1 (part); (a3) left p4–m1 (part); (a4) right p4–m3; b1–2. V 32128, upper jaw associated with lower one (note that the two are on different sides): (b1) right P4–M3; (b2) left p4–m3. Note: Upper case letters represent the upper jaw teeth, lower case letters represent the lower jaw teeth. The same applies below.

Figure A2.
Karakoromys decessus from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a) V 32129, left upper jaw with M1–2; (b) V 32117.3, left upper jaw with M1–3; (c) V 32119.1, left upper jaw with P4–M2 (part); (d) V 32120.1, left upper jaw with DP4–M3. (e) V 32131.2, left upper jaw with DP3–M2. (f) V 32120.3, left upper jaw with DP4–M3. (g) V 32113.2, right upper jaw with M1–3. (h) V 32118.1, right upper jaw with P4–M2. (i) V 32121.1, right upper jaw with M1–2 (only dentition is displayed). (j) V 32120.2, right upper jaw with DP3–M3.

Figure A3.
Karakoromys decessus from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a) V 32113.1, right lower jaw with p4–m3; (b) V 32116.4, right lower jaw with m1–3; (c1,c2) V 32117.1, a pair of lower jaws with both left and right dp4–m2; (d1,d2) V 32122, a pair of lower jaws with both left and right dp4–m1; (e) V 32114, right lower jaw with dp4–m2; (f) V 32123, right lower jaw with m1–3.

Figure A4.
Karakoromys decessus from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a1,a2) V 32119.3, a pair of lower jaws with both left and right m1–3; (b) V 32115.1, right lower jaw with p4–m3; (c) V 32125, left lower jaw with p4–m3; (d1,d2) V 32131.1, a pair of lower jaws with left p4-m3 and right p4, m2–3.

Figure A5.
Karakoromys arcanus from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a) V 32085.1, paired upper jaws with both left and right P4–M3; (b) V 32083, left upper jaw with M1–2. (c) V 32084, left upper jaw with M1–2; (d1,d2) V 32086.2, a pair of upper jaws with right M1-3 and left P4–M2; (e) V 32076.2, left upper jaw with P4–M3; (f) V 32075.1, left upper jaw with P4–M1.

Figure A6.
Karakoromys arcanus from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a) V 32071, left lower jaw with p4–m2; (b) V 32076.3, right lower jaw with dp4–m2 (only dentition is displayed); (c) V 32075.2, right lower jaw with m1–3; (d) V 32074, right lower jaw with m1–2; (e) V 32073, right lower jaw with m1–2; (f) V 32058, right lower jaw with p4–m2 (only dentition is displayed); (g) V 32072.11, left lower jaw with dp4–m1.

Figure A7.
Karakoromys chelkaris from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a) V 32163.1, a fragment of skull with left P4–M3 (only left upper jaw is displayed); (b) V 32163.2, left upper jaw with P4–M3; (c) V 32161.1, a fragment of skull with left M1–3 (only dentition is displayed); (d) V 32164.2, left upper jaw with M1–2; (e) V 32166.1, broken skull with right P4-M3 and left M2–3 (only upper jaws are displayed); (f) V 32166.2, right upper jaw with M1–3; (g) V 32162.2, right upper jaw with P4–M3; (h) V 32159, right lower jaw with m1–3; (i) V 32163.3, right lower jaw with m1–3; (j) V 32157, right lower jaw with m1–2; (k) V 32154, right lower jaw with m2–3; (l) V 32155, right lower jaw with dp4–m2.

Figure A8.
Karakoromys chelkaris from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a1,a2) V 32166.4, a pair of lower jaws with both left and right p4–m3; (b1,b2) V 32152, a pair of lower jaws with left m1-3 and right p4–m3; (c) V 32166.5, left lower jaw with m1–3; (d) V 32158.1, left lower jaw with m1–3; (e) V 32162.1, left lower jaw with p4–m3; (f) V 32153, left lower jaw with p4–m3; (g) V 32164.1, left lower jaw with p4–m1; (h) V 32160.2, left lower jaw with m1–3; (i) V 32156, left lower jaw with m2–3; (j) V 32166.6, left lower jaw with m2–3.

Figure A9.
Karakoromys conjunctus sp. nov. from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a1–a3) V 32180, a broken skull associated with lower jaws: (a1) upper jaws with right M1 (part)–M3 and left P4–M2; (a2) left lower jaw with p4–m1; (a3) right lower jaw with p4–m3 (Holotype). (b1–b3) V 32181, a broken skull associated with lower jaws (juvenile individual): (b1) upper jaws with right M1–3 and left DP3–M3; (b2) left lower jaw with m1–3; (b3) right lower jaw with m1 (part)–m3. (c1–c3) V 32167.2, upper jaw associated with lower ones: (c1) left upper jaw with P4–M3; (c2) left lower jaw with p4–m3; (c3) right lower jaw with P4–M2; (d) V 32175.1, left upper jaw with M1–3.

Figure A10.
Karakoromys conjunctus sp. nov. from Ulantatal Formation of Ulantatal area, Alxa Zuoqi, Inner Mongolia. (a1,a2) V 32195.1, a pair of lower jaws with both left and right p4–m3; (b) V 32173, left lower jaw with p4–m3; (c) V 32172, left lower jaw with p4–m3; (d) V 32170, left lower jaw with p4, m2–3; (e) V 32195.2, left lower jaw with m1–3; (f) V 32174.1, left lower jaw with m1–2; (g) V 32178, left lower jaw with m1–2; (h) V 32177, left lower jaw with m2–3 (part); (i) V 32194, left m2; (j) V 32179.2, right lower jaw with m1–2; (k) V 32169, right lower jaw with dp4–m2; (l) V 32176, right lower jaw with m1–3.
References
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef]
- Liu, Z.; Pagani, M.; Zinniker, D.; DeConto, R.; Huber, M.; Brinkhuis, H.; Shah, S.R.; Leckie, R.M.; Pearson, A. Global cooling during the Eocene-Oligocene climate transition. Science 2009, 323, 1187–1190. [Google Scholar] [CrossRef]
- Hren, M.T.; Sheldon, N.D.; Grimes, S.T.; Collinson, M.E.; Hooker, J.J.; Bugler, M.; Lohmann, K.C. Terrestrial cooling in Northern Europe during the eocene-oligocene transition. Proc. Natl. Acad. Sci. USA 2013, 110, 7562–7567. [Google Scholar] [CrossRef] [PubMed]
- Colwyn, D.A.; Hren, M.T. An abrupt decrease in Southern Hemisphere terrestrial temperature during the Eocene–Oligocene transition. Earth Planet. Sci. Lett. 2019, 512, 227–235. [Google Scholar] [CrossRef]
- Westerhold, T.; Marwan, N.; Drury, A.J.; Liebrand, D.; Agnini, C.; Anagnostou, E.; Barnet, J.S.; Bohaty, S.M.; De Vleeschouwer, D.; Florindo, F. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 2020, 369, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.G.; Browning, J.V.; Schmelz, W.J.; Kopp, R.E.; Mountain, G.S.; Wright, J.D. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records. Sci. Adv. 2020, 6, eaaz1346. [Google Scholar] [CrossRef]
- Hutchinson, D.K.; Coxall, H.K.; Lunt, D.J.; Steinthorsdottir, M.; de Boer, A.M.; Baatsen, M.; von der Heydt, A.; Huber, M.; Kennedy-Asser, A.T.; Kunzmann, L.; et al. The Eocene-Oligocene transition: A review of marine and terrestrial proxy data, models and model–data comparisons. Clim. Past 2021, 17, 269–315. [Google Scholar] [CrossRef]
- Berggren, W.A.; Prothero, D.R. Eocene-Oligocene Climatic and Biotic Evolution: An Overview. In Eocene-Oligocene Climatic and Biotic Evolution; Berggren, W.A., Prothero, D.R., Eds.; Princeton University Press: Princeton, NJ, USA, 1992; pp. 1–28. [Google Scholar]
- Dashzeveg, D. Asynchronism of the main mammalian faunal events near the Eocene-Oligocene boundary. Tert. Res. 1993, 14, 141–149. [Google Scholar]
- Prothero, D.R.; Heaton, T.H. Faunal stability during the early Oligocene climatic crash. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1996, 127, 257–283. [Google Scholar] [CrossRef]
- Meng, J.; McKenna, M.C. Faunal turnovers of Palaeogene mammals from the Mongolian Plateau. Nature 1998, 394, 364–367. [Google Scholar] [CrossRef]
- Hooker, J.J.; Collinson, M.E.; Sille, N.P. Eocene-Oligocene mammalian faunal turnover in the Hampshire Basin, UK: Calibration to the global time scale and the major cooling event. J. Geol. Soc. 2004, 161, 161–172. [Google Scholar] [CrossRef]
- Sun, J.; Ni, X.; Bi, S.; Wu, W.; Ye, J.; Meng, J.; Windley, B.F. Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene Boundary in Asia. Sci. Rep. 2014, 4, 7463. [Google Scholar] [CrossRef] [PubMed]
- Wang, B. The Mid-Tertiary Ctenodactylidae (Rodentia, Mammalia) of Eastern and Central Asia. Bull. Am. Mus. Nat. Hist. 1997, 234, 1–88. [Google Scholar]
- Dashzėvėg, D.; Meng, J. New Eocene Ctenodactyloid Rodents from the Eastern Gobi Desert of Mongolia and a Phylogenetic Analysis of Ctenodactyloids Based on Dental Features. Am. Mus. Novit. 1998, 3246, 1–20. [Google Scholar]
- Vianey-Liaud, M.; Gomes Rodrigues, H.; Marivaux, L. A new Oligocene Ctenodactylinae (Rodentia: Mammalia) from Ulantatal (nei Mongol): New insight on the phylogenetic origins of the modern Ctenodactylidae. Zool. J. Linn. Soc. 2010, 160, 531–550. [Google Scholar] [CrossRef]
- Li, Q.; Meng, J. New Ctenodactyloid Rodents from the Erlian Basin, Nei Mongol, China, and the Phylogenetic Relationships of Eocene Asian Ctenodactyloids. Am. Mus. Novit. 2015, 3828, 1–20. [Google Scholar] [CrossRef]
- Matthew, W.D.; Granger, W. Nine new rodents from the Oligocene of Mongolia. Am. Mus. Novit. 1923, 102, 1–10. [Google Scholar]
- Dashzeveg, D. [New data on the stratigraphy of middle Oligocène sédiments of the M.P.R.] Novyye dannyye o stratigrafii sredneoligotsenovkh otlozheniy MNR. Sovmest. Sov. Mong. Nauch. Issled. Geol. Exped. Tr. 1970, 2, 43. (In Russian) [Google Scholar]
- Kowalski, K. Middle Oligocene rodents from Mongolia. In Results of the Polish-Mongolian Palaeontological Expédition, Part V.; Kielan-Jaworowsk, Z., Ed.; Palaeontologia Polonica; Polish Academy of Sciences: Warsaw, Poland, 1974; Volume 30, pp. 147–178. [Google Scholar]
- Mellett, J.S. The Oligocene Hsanda Gol Formation, Mongolia: A revised faunal list. Am. Mus. Novit. 1968, 2318, 1–16. [Google Scholar]
- Daxner-Höck, G.; Höck, V.; Badamgarav, D.; Furtmüller, G.; Frank, W.; Montag, O.; Schmid, H.P. Cenozoic stratigraphy based on a sediment-basalt association in Central Mongolia as requirement for correlation across Central Asia. In Proceedings of the Biochronologie Mammalienne du Cénozoique en Europe et Domaines Reliés, Montpellier, France, 14 April 1997; pp. 163–176. [Google Scholar]
- Schmidt-Kittler, N.; Vianey-Liaud, M.; Marivaux, L. Oligocene-Miocene Vertebrates from the Valley of Lakes (Central Mongolia): Morphology, phylogenetic and stratigraphic implications 6. The Ctenodactylidae (Rodentia, Mammalia). Ann. Nat. Mus. Wien 2007, 108, 173–215. [Google Scholar]
- Daxner-Höck, G.; Badamgarav, D.; Erbajeva, M. Oligocene stratigraphy based on a sediment-basalt association in Central Mongolia (Taatsiin Gol and Taatsiin Tsagaan Nuur area, Valley of Lakes): Review of a Mongolian-Austrian project. Vertebr. PalAsiat. 2010, 48, 348. [Google Scholar]
- Daxner-Hock, G.; Badamgarav, D.; Barsbold, R.; Bayarmaa, B.; Erbajeva, M.; Gohlich, U.B.; Harzhauser, M.; Hock, E.; Hock, V.; Ichinnorov, N.; et al. Oligocene stratigraphy across the Eocene and Miocene boundaries in the Valley of Lakes (Mongolia). Paleobiodivers. Paleoenviron. 2017, 97, 111–218. [Google Scholar] [CrossRef]
- Oliver, A.; Sanisidro, O.; Baatarjav, B.; Niiden, I.; Daxner-Hock, G. Diversification rates in Ctenodactylidae (Rodentia, Mammalia) from Mongolia. Paleobiodivers. Paleoenviron. 2017, 97, 51–65. [Google Scholar] [CrossRef]
- Höck, V.; Daxner-Höck, G.; Schmid, H.P.; Badamgarav, D.; Frank, W.; Furtmüller, G.; Montag, O.; Barsbold, R.; Khand, Y.; Sodov, J. Oligocene-Miocene sediments, fossils and basalts from the Valley of Lakes (Central Mongolia): An integrated study. Mitt. Österr. Geol. Ges. 1999, 90, 83–125. [Google Scholar]
- Wang, B.; Chang, J.; Meng, X.; Chen, J. Stratigraphy of the Upper and Middle Oligocene of Qianlishan District, Nei Mongol (Inner Mongolia). Vertebr. PalAsiat. 1981, 19, 26–34. [Google Scholar]
- Huang, X. Middle Oligocene Ctenodactylids (Rodentia, Mammalia) of Ulantatal, Nei Mongol. Vertebr. PalAsiat. 1985, 23, 27–38. [Google Scholar]
- Wang, B.; Wang, P. Discovery of Early Medial Oligocene Mammalian Fauna From Kekeamu, Alxa Left Banner, Nei Mongol. Vertebr. PalAsiat. 1991, 29, 64–71. [Google Scholar]
- Vianey-Liaud, M.; Schmidt-Kittler, N.; Marivaux, L. The Ctenodactylidae (Rodentia) from the Oligocene of Ulantatal (Inner Mongolia, China). Palaeovertebrata 2006, 34, 111–206. [Google Scholar]
- Zhang, Z.; Liu, Y.; Wang, L.; Kaakinen, A.; Wang, J.; Mao, F.; Tong, Y. Lithostratigraphic context of Oligocene mammalian faunas from Ulantatal, Nei Mongol, China. Comptes Rendus Palevol 2016, 15, 903–910. [Google Scholar] [CrossRef]
- Wasiljeff, J.; Kaakinen, A.; Salminen, J.M.; Zhang, Z. Magnetostratigraphic constraints on the fossiliferous Ulantatal sequence in Inner Mongolia, China: Implications for Asian aridification and faunal turnover before the Eocene-Oligocene boundary. Earth Planet. Sci. Lett. 2020, 535, 116125. [Google Scholar] [CrossRef]
- Qiu, Z.; Wang, B.; Qiu, Z.; Xie, G.; Xie, J.; Wang, X. Recent advances in study of the Xianshuihe Formation in Lanzhou Basin. In Evidence for Evolution—Eassays in Honor of Prof. Chungchien Young on the Hundredth Anniversary of His Birth; Tong, Y., Zhang, Y., Wu, W., Li, J., Shi, L., Eds.; China Ocean Press: Beijing, China, 1997; pp. 177–192. [Google Scholar]
- Wu, W.; Meng, J.I.N.; Ye, J.I.E.; Ni, X. Propalaeocastor (Rodentia, Mammalia) from the Early Oligocene of Burqin Basin, Xinjiang. Am. Mus. Novit. 2004, 3461, 1–16. [Google Scholar] [CrossRef]
- Jie, Y.; Jin, M.; Wen-Yu, W.; Xi-Jun, N. Late Eocene-Early Oligocene Lithological and Biological Stratigraphy in the Burqin Region of Xinjiang. Vertebr. PalAsiat. 2005, 43, 49–60. [Google Scholar]
- Emry, R.J.; Lucas, S.G.; Tyutkova, L.; Wang, B. The Ergilian-Shandgolian (Eocene-Oligocene) transition in the Zaysan basin, Kazakhstan. Bull. Carnegie Mus. Nat. Hist. 1998, 34, 298–312. [Google Scholar]
- Wasiljeff, J.; Zhang, Z. Stratigraphical significance of Ulantatal sequence (Nei Mongol, China) in refining the latest Eocene and Oligocene terrestrial regional stages. Vertebr. PalAsiat. 2021, 60, 42–53. [Google Scholar] [CrossRef]
- Tang, X.; Feng, Q.; Li, D. Tectonic features and evolution of Bayanhot Basin in western Inner Mongolia. Oil Gas Geol. 1990, 2, 127–135. [Google Scholar]
- Wang, P.; Wang, B. Tertiary system and its fauna of Alxa area, Nei Mongol, China. Northwest Geosci. 1998, 19, 1–37. [Google Scholar]
- Zhao, J.; Zhou, Y.; Wang, K.; Li, X.; Ma, Z.; Ruan, Z.; Wang, D.; Xue, R. Provenance and paleogeography of Carboniferous–Permian strata in the Bayanhot Basin: Constraints from sedimentary records and detrital zircon geochronology. Geosci. Front. 2021, 12, 101088. [Google Scholar] [CrossRef]
- Wasiljeff, J.; Salminen, J.M.; Stenman, J.; Zhang, Z.; Kaakinen, A. Oligocene moisture variations as evidenced by an aeolian dust sequence in Inner Mongolia, China. Sci. Rep. 2022, 12, 5597. [Google Scholar] [CrossRef]
- Wible, J.R.; Wang, Y.; Li, C.; Dawson, M.R. Cranial Anatomy and Relationships of a New Ctenodactyloid (Mammalia, Rodentia) from the Early Eocene of Hubei Province, China. Ann. Carnegie Mus. 2005, 74, 91–150. [Google Scholar] [CrossRef]
- SPSS. IBM SPSS Statistics for Windows, Version 20.0; SPSS Inc. Ltd.: Chicago, IL, USA, 2013. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: https://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 5 April 2023).
- Grimm, E.C. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Grimm, E.C. Tilia and Tiliagraph; Springfield, Illinois State Museum: Springfield, IL, USA, 1991; pp. 1–101. [Google Scholar]
- Hammer, Ø.; Harper, D.A. Paleontological Data Analysis, 1st ed.; Blackwell Publishing: Oxford, UK, 2006; pp. 1–351. [Google Scholar]
- Bowdich, T.E. An Analysis of the Natural Classifications of Mammalia: For the Use of Students and Travellers; Printed by J. Smith: Paris, France, 1821; pp. 51–61. [Google Scholar]
- Tullberg, T. Ueber das System der Nagethiere: Eine Phylogenetische Studie; Akademische Buchdruckerei: Herborn, Germany, 1899. [Google Scholar]
- Gervais, M. Description ostéologique de l’Anomalurus et remarques sur la classification naturelle des rongeurs. Ann. Sci. Nat. Zool. 1853, 20, 238–246. [Google Scholar]
- Wang, B. The Ctenodactyloidea of Asia. Natl. Sci. Mus. Monogr. 1994, 8, 35–47. [Google Scholar]
- Shevyreva, N. New middle Oligocene rodents of Kazakhstan and Mongolia. Acad. Sci. USSR Trans. Paleontol. Inst. 1971, 130, 70–86. [Google Scholar]
- Shevyreva, N. New rodents (Rodentia, Mammalia) from the lower Oligocene of the Zaisan depression (Eastern Kazakhstan). Paleontol. Zhurnal 1994, 1, 111–126. [Google Scholar]
- Wang, B.; Qiu, Z. Discovery of early Oligocene mammalian fossils from Danghe area, Gansu, China. Vertebr. PalAsiat. 2004, 42, 130–143. [Google Scholar]
- Li, Q.; Wang, X.; Xie, G.; Yin, A. Oligocene-Miocene mammalian fossils from Hongyazi Basin and its bearing on tectonics of Danghe Nanshan in Northern Tibetan Plateau. PLoS ONE 2013, 8, e82816. [Google Scholar] [CrossRef]
- Wood, A.E. The evolution of the rodent family Ctenodactylidae. J. Palaeontol. Soc. India 1977, 20, 120–137. [Google Scholar]
- Gradstein, F.M.; Ogg, J.G.; Smith, A.G. A Geologic Time Scale 2004; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Sun, R. Principles of Animal Ecology (Translated from Chinese); Beijing Normal University Publishing Group: Bejing, China, 2006; p. 386. [Google Scholar]
- Hadly, E.A. Influence of late-Holocene climate on northern Rocky Mountain mammals. Quat. Res. 1996, 46, 298–310. [Google Scholar] [CrossRef]
- Wagner, P.J.; Kosnik, M.A.; Lidgard, S. Abundance distributions imply elevated complexity of post-Paleozoic marine ecosystems. Science 2006, 314, 1289–1292. [Google Scholar] [CrossRef]
- Canales, M.L.; Henriques, M.H. Foraminifera from the Aalenian and the Bajocian GSSP (Middle Jurassic) of Murtinheira section (Cabo Mondego, West Portugal): Biostratigraphy and paleoenvironmental implications. Mar. Micropaleontol. 2008, 67, 155–179. [Google Scholar] [CrossRef]
- Western, D.; Behrensmeyer, A.K. Bone assemblages track animal community structure over 40 years in an African savanna ecosystem. Science 2009, 324, 1061–1064. [Google Scholar] [CrossRef] [PubMed]
- Elshanawany, R.; Mohamed, Y.; Ibrahim, M.I.A. Paleontological Records of Holocene Environmental Changes offshore, Egypt. J. Ocean Univ. China 2019, 18, 1382–1394. [Google Scholar] [CrossRef]
- Cole, S.R.; Wright, D.F.; Ausich, W.I.; Koniecki, J.M. Paleocommunity composition, relative abundance, and new camerate crinoids from the Brechin Lagerstätte (Upper Ordovician). J. Paleontol. 2020, 94, 1103–1123. [Google Scholar] [CrossRef]
- Miller, K.G.; Wright, J.D.; Fairbanks, R.G. Unlocking the Ice House: Oligocene-Miocene oxygen isotopes, eustasy, and margin erosion. J. Geophys. Res. Solid Earth 1991, 96, 6829–6848. [Google Scholar] [CrossRef]
- Coxall, H.K.; Wilson, P.A.; Pälike, H.; Lear, C.H.; Backman, J. Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature 2005, 433, 53–57. [Google Scholar] [CrossRef]
- Harzhauser, M.; Daxner-Hock, G.; Lopez-Guerrero, P.; Maridet, O.; Oliver, A.; Piller, W.E.; Richoz, S.; Erbajeva, M.A.; Neubauer, T.A.; Gohlich, U.B. Stepwise onset of the Icehouse world and its impact on Oligo-Miocene Central Asian mammals. Sci. Rep. 2016, 6, 36169. [Google Scholar] [CrossRef]
- Richoz, S.; Baldermann, A.; Frauwallner, A.; Harzhauser, M.; Daxner-Hock, G.; Klammer, D.; Piller, W.E. Geochemistry and mineralogy of the Oligo-Miocene sediments of the Valley of Lakes, Mongolia. Paleobiodivers. Paleoenviron. 2017, 97, 233–258. [Google Scholar] [CrossRef]
- Hopkins, S.S.B. Estimation of body size in fossil mammals. In Methods in Paleoecology: Reconstructing Cenozoic Terrestrial Environments and Ecological Communities; Croft, D.A., Su, D.F., Simpson, S.W., Eds.; Springer: Cham, Switzerland, 2018; pp. 7–22. [Google Scholar]
- Barnosky, A.D. Distinguishing the effects of the Red Queen and Court Jester on Miocene mammal evolution in the northern Rocky Mountains. J. Vertebr. Paleontol. 2001, 21, 172–185. [Google Scholar] [CrossRef]
- Žliobaitė, I.; Fortelius, M.; Stenseth, N.C. Reconciling taxon senescence with the Red Queen’s hypothesis. Nature 2017, 552, 92–95. [Google Scholar] [CrossRef]
- Ladant, J.-B.; Donnadieu, Y.; Lefebvre, V.; Dumas, C. The respective role of atmospheric carbon dioxide and orbital parameters on ice sheet evolution at the Eocene-Oligocene transition. Paleoceanography 2014, 29, 810–823. [Google Scholar] [CrossRef]
- Ao, H.; Dupont-Nivet, G.; Rohling, E.J.; Zhang, P.; Ladant, J.B.; Roberts, A.P.; Licht, A.; Liu, Q.; Liu, Z.; Dekkers, M.J.; et al. Orbital climate variability on the northeastern Tibetan Plateau across the Eocene-Oligocene transition. Nat. Commun. 2020, 11, 5249. [Google Scholar] [CrossRef] [PubMed]
- Gomes Rodrigues, H.; Marivaux, L.; Vianey-Liaud, M. Expansion of open landscapes in Northern China during the Oligocene induced by dramatic climate changes: Paleoecological evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 358–360, 62–71. [Google Scholar] [CrossRef]
- Sousa, F.V. Biogeography and Phylogenetic Position of a Sahara-Sahel Mountain Endemic, Felovia Vae (Ctenodactylidae). Master’s Dissertation, University of Porto, Porto, Portugal, 2015. [Google Scholar]
- Xu, R.; Li, Q. New skulls of ctenodactyloids from the Early Oligocene of Ulantatal, Nei Mongol, China. Vertebr. PalAsiat. 2020, 58, 305–327. [Google Scholar]
- Antoine, P.-O.; Yans, J.; Castillo, A.A.; Stutz, N.; Abello, M.A.; Adnet, S.; Custódio, M.A.; Benites-Palomino, A.; Billet, G.; Boivin, M.; et al. Biotic community and landscape changes around the Eocene–Oligocene transition at Shapaja, Peruvian Amazonia: Regional or global drivers? Glob. Planet. Chang. 2021, 202, 103512. [Google Scholar] [CrossRef]
- Butler, P. Homologies of molar cusps and crests, and their bearing on assessments of rodent phylogeny. In Evolutionary Relationships among Rodents: A Multidisciplinary Analysis; Springer: New York, NY, USA, 1985; pp. 381–401. [Google Scholar]
- Ferrusquia-Villafranca, I.; Flynn, L.J.; Ruiz-Gonzalez, J.E.; Torres-Hernandez, J.R.; Martinez-Hernandez, E. New Eocene rodents from northwestern Oaxaca, southeastern Mexico, and their paleobiological significance. J. Vertebr. Paleontol. 2018, 38, e1514615. [Google Scholar] [CrossRef]
- Angelone, C.; Zhang, Z. Climate change and evolution of early lagomorphs (Mammalia): A study perspective based on new materials of Ordolagus from Nei Mongol (northern China). Vertebr. PalAsiat. 2021, 59, 138. [Google Scholar]
- Lukens, W.E.; Stinchcomb, G.E.; Nordt, L.C.; Kahle, D.J.; Driese, S.G.; Tubbs, J.D. Recursive partitioning improves paleosol proxies for rainfall. Am. J. Sci. 2019, 319, 819–845. [Google Scholar] [CrossRef]
- Fischer, R.; Turner, N.C. Plant productivity in the arid and semiarid zones. Annu. Rev. Plant Physiol. 1978, 29, 277–317. [Google Scholar] [CrossRef]
- Zheng, S.; Shangguan, Z. Spatial patterns of foliar stable carbon isotope compositions of C3 plant species in the Loess Plateau of China. Ecol. Res. 2006, 22, 342–353. [Google Scholar] [CrossRef]
- Zhao, X.; Tan, K.; Zhao, S.; Fang, J. Changing climate affects vegetation growth in the arid region of the northwestern China. J. Arid Environ. 2011, 75, 946–952. [Google Scholar] [CrossRef]
- Peng, J.; Dong, W.; Yuan, W.; Zhang, Y. Responses of grassland and forest to temperature and precipitation changes in Northeast China. Adv. Atmos. Sci. 2012, 29, 1063–1077. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).