
Evaluation of the Potential Seed Dispersal Effectiveness of Malus sieversii (Lebed.) M. Roem. by Cattle


Abstract
:1. Introduction
2. Materials and Methods
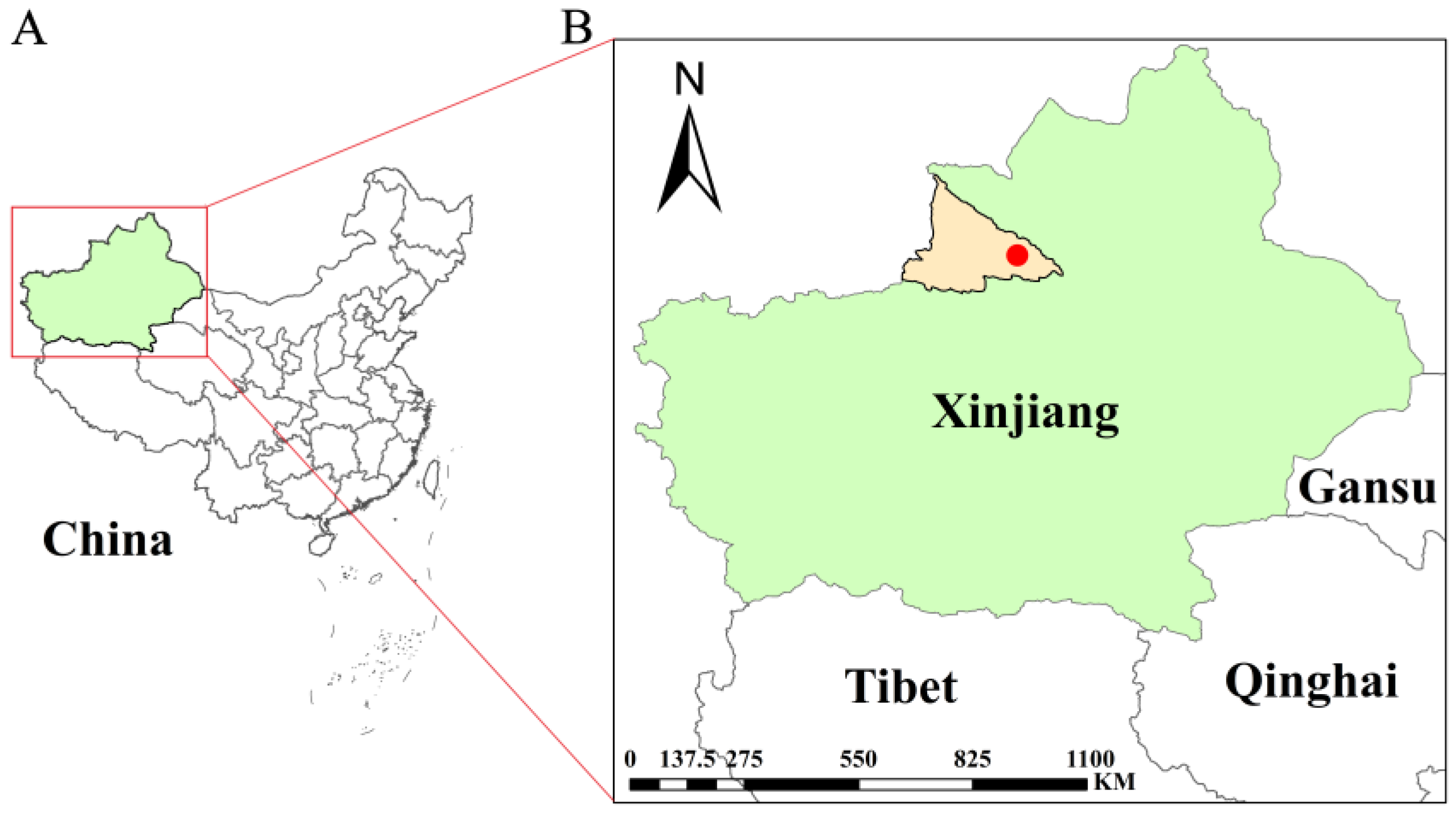
2.1. Study Site
2.2. Survey of Animals Visiting Ground Fruits
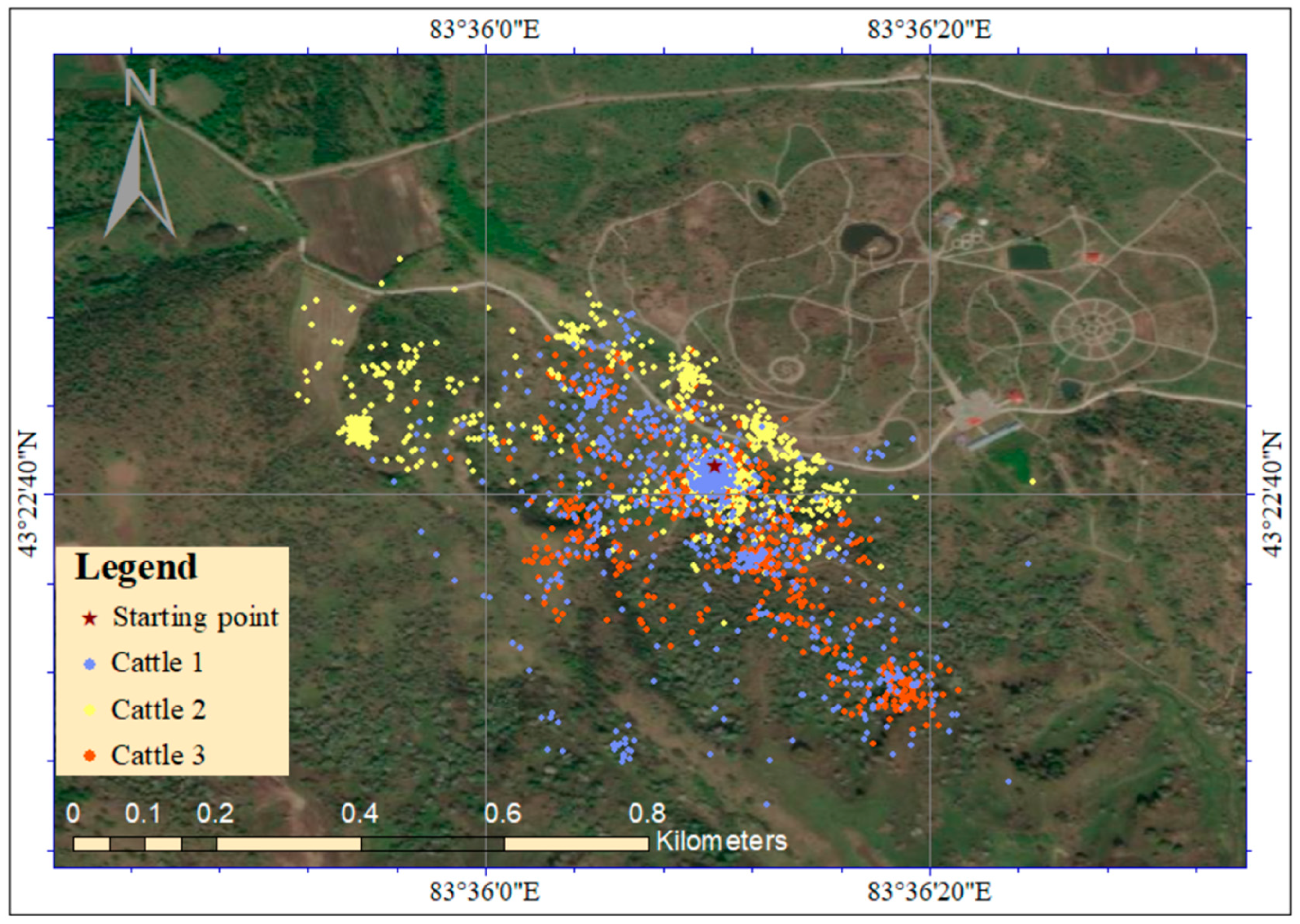
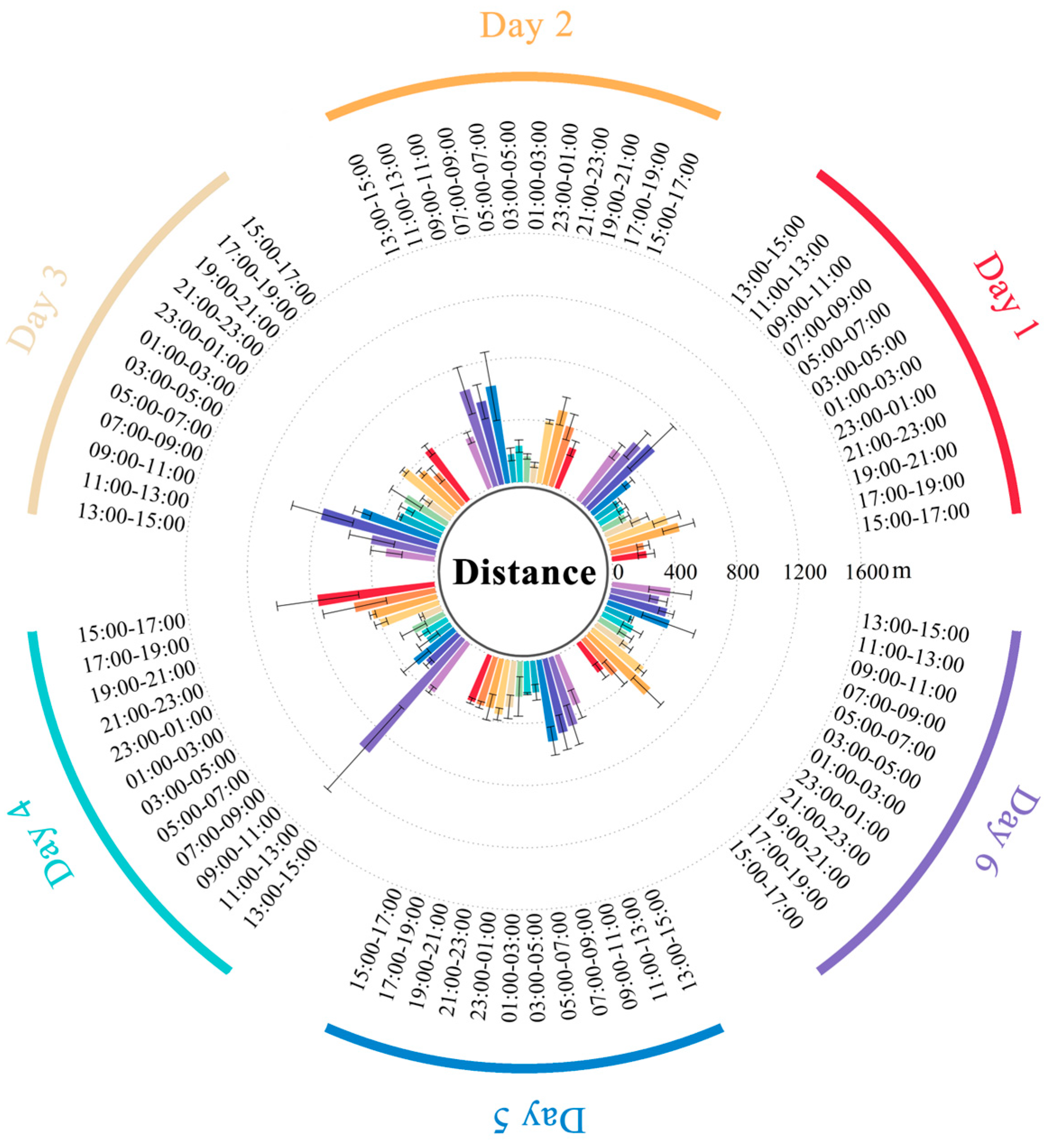
2.3. Estimation of Seed Dispersal Distance by Cattle
2.4. Germination Trials
2.4.1. Seed Germination Experiment in Laboratory
2.4.2. Seed Germination Experiment in Wild
2.5. Statistical Analyses
3. Results
3.1. Animals Visiting Ground Fruits
3.2. Seed Dispersal Distance by Cattle
3.3. Seed Germination
3.4. Seedling Emergence and Survival
4. Discussion
4.1. Seed Dispersal Quantity of Cattle
4.2. Seed Dispersal Quality of Cattle
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Jordano, P.; Forget, P.-M.; Lambert, J.E.; Böhning-Gaese, K.; Traveset, A.; Wright, S.J. Frugivores and seed dispersal: Mechanisms and consequences for biodiversity of a key ecological interaction. Biol. Lett. 2011, 7, 321–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.C.; Smith, T.B. Closing the seed dispersal loop. Trends Ecol. Evol. 2002, 17, 379–386. [Google Scholar] [CrossRef]
- Mouissie, A.M.; Lengkeek, W.; Diggelen, R.V. Estimating adhesive seed-dispersal distances: Field experiments and correlated random walks. Funct. Ecol. 2005, 19, 478–486. [Google Scholar] [CrossRef]
- Casper, B.B.; Heard, S.B.; Apanius, V. Ecological correlates of single-seededness in a woody tropical flora. Oecologia 1992, 90, 212–217. [Google Scholar] [CrossRef]
- Traveset, A.; Robertson, A.W.; Rodríguez-Pérez, J. A Review on the Role of Endozoochory in Seed Germination. In Seed Dispersal: Theory and Its Application in a Changing World; CABI Pub.: Wallingford, UK, 2007; pp. 78–103. ISBN 978-1-84593-165-0. [Google Scholar]
- Brochet, A.L.; Guillemain, M.; Fritz, H.; Gauthier-Clerc, M.; Green, A.J. Plant dispersal by teal (Anas crecca) in the Camargue: Duck guts are more important than their feet: Plant dispersal by teal. Freshw. Biol. 2010, 55, 1262–1273. [Google Scholar] [CrossRef]
- Vander Wall, S.B.; Beck, M.J. A comparison of frugivory and scatter-hoarding seed-dispersal syndromes. Bot. Rev. 2012, 78, 10–31. [Google Scholar] [CrossRef]
- McConkey, K.R. Seed dispersal by primates in Asian habitats: From species, to communities, to conservation. Int. J. Primatol. 2018, 39, 466–492. [Google Scholar] [CrossRef]
- Zwolak, R.; Sih, A. Animal personalities and seed dispersal: A conceptual review. Funct. Ecol. 2020, 34, 1294–1310. [Google Scholar] [CrossRef]
- Naniwadekar, R.; Mishra, C.; Datta, A. Fruit resource tracking by hornbill species at multiple scales in a tropical forest in India. J. Trop. Ecol. 2015, 31, 477–490. [Google Scholar] [CrossRef]
- Butler, H.C.; Johnson, S.D. Seed dispersal by monkey spitting in Scadoxus (Amaryllidaceae): Fruit selection, dispersal distances and effects on seed germination. Austral. Ecol. 2022, 47, 1029–1036. [Google Scholar] [CrossRef]
- Bascompte, J.; Jordano, P. Plant-animal mutualistic networks: The architecture of biodiversity. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 567–593. [Google Scholar] [CrossRef]
- Jordano, P.; García, C.; Godoy, J.A.; García-Castaño, J.L. Differential contribution of frugivores to complex seed dispersal patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 3278–3282. [Google Scholar] [CrossRef]
- Spiegel, O.; Nathan, R. Incorporating dispersal distance into the disperser effectiveness framework: Frugivorous birds provide complementary dispersal to plants in a patchy environment. Ecol. Lett. 2007, 10, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Calviño-Cancela, M.; Martín-Herrero, J. Effectiveness of a varied assemblage of seed dispersers of a fleshy-fruited plant. Ecology 2009, 90, 3503–3515. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, D.P.; Morris, W.F.; Jordano, P. Interaction frequency as a surrogate for the total effect of animal mutualists on plants: Total effect of animal mutualists on plants. Ecol. Lett. 2005, 8, 1088–1094. [Google Scholar] [CrossRef]
- Schupp, E.W.; Jordano, P.; Gómez, J.M. A general framework for effectiveness concepts in mutualisms. Ecol. Lett. 2017, 20, 577–590. [Google Scholar] [CrossRef]
- Haurez, B.; Tagg, N.; Petre, C.-A.; Brostaux, Y.; Boubady, A.; Doucet, J.-L. Seed dispersal effectiveness of the western lowland gorilla (Gorilla gorilla gorilla) in Gabon. Afr. J. Ecol. 2018, 56, 185–193. [Google Scholar] [CrossRef]
- Quintero, E.; Pizo, M.A.; Jordano, P. Fruit resource provisioning for avian frugivores: The overlooked side of effectiveness in seed dispersal mutualisms. J. Ecol. 2020, 108, 1358–1372. [Google Scholar] [CrossRef]
- Cárdenas, S.; Echeverry-Galvis, M.Á.; Stevenson, P.R. Seed dispersal effectiveness by oilbirds (Steatornis caripensis) in the Southern Andes of Colombia. Biotropica 2021, 53, 671–680. [Google Scholar] [CrossRef]
- Jerozolimski, A.; Ribeiro, M.B.N.; Martins, M. Are tortoises important seed dispersers in Amazonian forests? Oecologia 2009, 161, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A. Animal-mediated seed dispersal in India: Implications for conservation of India’s biodiversity. Biotropica 2022, 54, 1320–1330. [Google Scholar] [CrossRef]
- Pérez-Méndez, N.; Jordano, P.; Valido, A. Downsized mutualisms: Consequences of seed dispersers’ body-size reduction for early plant recruitment. Perspect. Plant Ecol. 2015, 17, 151–159. [Google Scholar] [CrossRef]
- Naniwadekar, R.; Rathore, A.; Shukla, U.; Chaplod, S.; Datta, A. How far do Asian forest hornbills disperse seeds? Acta Oecologica 2019, 101, 103482. [Google Scholar] [CrossRef]
- Li, N.; Zhong, M.; Leng, X.; Wang, A.; Fang, S.B.; An, S.Q. Seed dispersal effectiveness of plant by frugivores: A review. Chin. J. Ecol. 2015, 34, 2041–2047. [Google Scholar] [CrossRef]
- Camargo, P.H.S.A.; Martins, M.M.; Feitosa, R.M.; Christianini, A.V. Bird and ant synergy increases the seed dispersal effectiveness of an ornithochoric shrub. Oecologia 2016, 181, 507–518. [Google Scholar] [CrossRef]
- González-Castro, A.; Calviño-Cancela, M.; Nogales, M. Comparing seed dispersal effectiveness by frugivores at the community level. Ecology 2015, 96, 808–818. [Google Scholar] [CrossRef]
- Schupp, E.W.; Jordano, P.; Gómez, J.M. Seed dispersal effectiveness revisited: A conceptual review. New Phytol. 2010, 188, 333–353. [Google Scholar] [CrossRef]
- Chu, J.Y.; Feng, L.J.; Hou, Y.X.; Lu, B.; Wang, Q.; Zhou, L.; Wang, J. Analysis on population damage of Malus sieversii. Non Wood For. Res. 2022, 40, 265–273. [Google Scholar] [CrossRef]
- Duan, N.; Bai, Y.; Sun, H.; Wang, N.; Ma, Y.; Li, M.; Wang, X.; Jiao, C.; Legall, N.; Mao, L.; et al. Genome re-sequencing reveals the history of apple and supports a two-stage model for fruit enlargement. Nat. Commun. 2017, 8, 249. [Google Scholar] [CrossRef]
- Lamboy, W.F.; Yu, J.; Forsline, P.L.; Weeden, N.F. Partitioning of allozyme diversity in wild populations of Malus sieversii L. and implications for germplasm collection. J. Am. Soc. Hortic. Sci. 1996, 121, 982–987. [Google Scholar] [CrossRef]
- Yang, M.; Li, F.; Long, H.; Yu, W.; Yan, X.; Liu, B.; Zhang, Y.; Yan, G.; Song, W. Ecological distribution, reproductive characteristics, and in situ conservation of Malus sieversii in Xinjiang, China. HortScience 2016, 51, 1197–1201. [Google Scholar] [CrossRef]
- Mierkamili, M.; Liu, Z.Q.; Ma, X.D.; Zhang, H.X.; Tian, Z.P. Survival status, problems and conservation strategies of Malus sieversii. Guihaia 2021, 41, 2100–2109. [Google Scholar] [CrossRef]
- Cornille, A.; Giraud, T.; Smulders, M.J.M.; Roldán-Ruiz, I.; Gladieux, P. The domestication and evolutionary ecology of apples. Trends Genet. 2014, 30, 57–65. [Google Scholar] [CrossRef]
- Panyushkina, I.; Mukhamadiev, N.; Lynch, A.; Ashikbaev, N.; Arizpe, A.; O’Connor, C.; Abjanbaev, D.; Mengdibayeva, G.; Sagitov, A. Wild apple growth and climate change in southeast Kazakhstan. Forests 2017, 8, 406. [Google Scholar] [CrossRef]
- Cui, X.N.; Liu, D.G.; Liu, A.H. Research progress in integrated management of Agrilus mali. Plant Prot. 2015, 41, 16–23. [Google Scholar]
- Shan, Q.; Wang, Z.; Ling, H.; Zhang, G.; Yan, J.; Han, F. Unreasonable human disturbance shifts the positive effect of climate change on tree-ring growth of Malus sieversii in the origin area of world cultivated apples. J. Clean. Prod. 2021, 287, 125008. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Xu, J.; Shi, X.J. Effects of different light intensities on seed germination and seedling growth of Malus sieversii. J. Xinjiang Agric. Univ. 2021, 44, 401–406. [Google Scholar] [CrossRef]
- Zhang, H.X.; Wen, Z.B.; Wang, Q. Population genetic structure of Malus sieversii and environmental adaptations. Chin. J. Plant Ecol. 2022, 46, 1098–1108. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Xu, J.; Shi, X.J. Responses of seedling growth and biomass allocation of Malus sieversii to precipitation amount and precipitation interval. Arid Zone Res. 2023, 40, 102–110. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Z.; Bai, S.; Lv, Y.; Shi, X.; Tan, D. Recovery and germination of Malus sieversii (Ledeb.) M. Roem. (Rosaceae) seeds after ingestion by cattle, horses, and sheep. Sustainability 2022, 14, 13930. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, Y.M.; Zhou, X.B. Ecological stoichiometry of surface soil nutrient and its influencing factors in the wild fruit forest in Yili region, Xinjiang, China. Chin. J. Appl. Ecol. 2016, 27, 10. [Google Scholar] [CrossRef]
- Zhang, X.S. On the eco-geographical characters and the problems of classification of the wild fruit-tree forest in the Ili valley of Sinkiang. J. Integr. Plant Biol. 1973, 2, 239–253. [Google Scholar]
- Yan, G.R. Study on the wild fruit trees and its conservation of Tianshan mountain in Xinjiang. Wild Plant Res. 2001, 20, 13–14. [Google Scholar]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World; Johns Hopkins University Press: Baltimore, MD, USA, 2005; ISBN 978-0-8018-8221-0. [Google Scholar]
- Campos, C.M.; Campos, V.E.; Miguel, F.; Cona, M.I. Management of protected areas and its effect on an ecosystem function: Removal of Prosopis flexuosa seeds by mammals in Argentinian drylands. PLoS ONE 2016, 11, e0162551. [Google Scholar] [CrossRef] [PubMed]
- Miguel, F.; Cona, M.I.; Campos, C.M. Seed removal by different functional mammal groups in a protected and grazed landscape of the Monte, Argentina. Seed Sci. Res. 2017, 27, 174–182. [Google Scholar] [CrossRef]
- Prasad, S.; Pittet, A.; Sukumar, R. Who really ate the fruit? A novel approach to camera trapping for quantifying frugivory by ruminants. Ecol. Res. 2010, 25, 225–231. [Google Scholar] [CrossRef]
- Blake, S.; Wikelski, M.; Cabrera, F.; Guezou, A.; Silva, M.; Sadeghayobi, E.; Yackulic, C.B.; Jaramillo, P. Seed dispersal by Galápagos tortoises. J. Biogeogr. 2012, 39, 1961–1972. [Google Scholar] [CrossRef]
- Qin, W.; Xiao, Y.Q.; Yan, J.J.; Ma, Y. Comparative study on fruit and seed morphology of 17 types of Malus sieversii (Led.) Roem. J. Xinjiang Agric. Univ. 2014, 37, 373–378. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier/AP: San Diego, CA, USA, 2014; ISBN 978-0-12-416677-6. [Google Scholar]
- Liu, Z.Q.; Dong, H.G.; Yu, T.; Chen, W.M. Study on the germination characteristics of Malus sieversii seeds and the field transplanting of seedlings of different seedling ages. J. Anhui Agric. Sci. 2021, 49, 54–56. [Google Scholar] [CrossRef]
- Zhang, Z.Z. Study on Sexual Regeneration Limitation of Malus sieversii Population. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2022. [Google Scholar]
- Campos, C.M.; Velez, S.; Miguel, M.F.; Papú, S.; Cona, M.I. Studying the quantity component of seed dispersal effectiveness from exclosure treatments and camera trapping. Ecol. Evol. 2018, 8, 5470–5479. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.G.N.; Koroiva, R.; Cassimiro, C.A.L.; Batista, F.R.C. Endangered globose cactus Melocactus lanssensianus P. J. Braun depends on lizards for effective seed dispersal in the Brazilian Caatinga. Plant Ecol. 2021, 222, 14. [Google Scholar] [CrossRef]
- Jordano, P. Fruits and Frugivory. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Pub.: Wallingford, UK, 2000; pp. 125–166. ISBN 0-85199-432-6. [Google Scholar]
- Castañeda, I.; Doherty, T.S.; Fleming, P.A.; Stobo-Wilson, A.M.; Woinarski, J.C.Z.; Newsome, T.M. Variation in red fox Vulpes vulpes diet in five continents. Mammal Rev. 2022, 52, 328–342. [Google Scholar] [CrossRef]
- Cain, M.L.; Milligan, B.G.; Strand, A.E. Long-distance seed dispersal in plant populations. Am. J. Bot. 2000, 87, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Schupp, E.W. Quantity, quality and the effectiveness of seed dispersal by animals. Vegetatio 1993, 1, 15–29. [Google Scholar] [CrossRef]
- Zhao, G.; Xu, Z.X.; Li, D.S. A review about grazing behavior of domestic ruminants. J. Inn. Mong. Agric. Univ. Nat. Sci. Ed. 2000, 2, 109–116. [Google Scholar] [CrossRef]
- Hou, L.L.; Wang, X.; Zhang, X.; Yan, Y.C.; Yan, R.R.; Cheng, L.; Xin, X.P. The effect of grazing intensity on beef cattle’s behavior. Acta Agrestia Sin. 2021, 29, 1974–1982. [Google Scholar]
- Traveset, A. Effect of seed passage through vertebrate frugivores’ guts on germination: A review. Perspect. Plant Ecol. Evol. Syst. 1998, 1, 151–190. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, C.C.; Baskin, J.M.; Heshmati, S.; Mirfazeli, M.S. A meta-analysis of the effects of frugivory (endozoochory) on seed germination: Role of seed size and kind of dormancy. Plant Ecol. 2018, 219, 1283–1294. [Google Scholar] [CrossRef]
- Illescas-Gallegos, E.; Rodríguez-Trejo, D.A.; Villanueva-Morales, A.; Borja-de La Rosa, M.A.; Ordóñez-Candelaria, V.R.; Ortega-Aragón, L.A. Factors influencing physical dormancy and its elimination in two legumes. Rev. Chapingo Ser. Cienc. For. Am. 2021, 27, 413–429. [Google Scholar] [CrossRef]
- Carlo, T.A.; García, D.; Martínez, D.; Gleditsch, J.M.; Morales, J.M. Where do seeds go when they go far? Distance and directionality of avian seed dispersal in heterogeneous landscapes. Ecology 2013, 94, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.M.; García, D.; Martínez, D.; Rodriguez-Pérez, J.; Herrera, J.M. Frugivore behavioural details matter for seed dispersal: A multi-species model for Cantabrian thrushes and trees. PLoS ONE 2013, 8, e65216. [Google Scholar] [CrossRef]
- Campos, C.M.; Campos, V.E.; Mongeaud, A.; Borghi, C.E.; De Los Ríos, C.; Giannoni, S.M. Relationships between Prosopis flexuosa (Fabaceae) and cattle in the Monte desert: Seeds, seedlings and saplings on cattle-use site classes. Rev. Chil. Hist. Nat. 2011, 84, 289–299. [Google Scholar] [CrossRef]
- Venier, P.; Carrizo García, C.; Cabido, M.; Funes, G. Survival and germination of three hard-seeded Acacia species after simulated cattle ingestion: The importance of the seed coat structure. S. Afr. J. Bot. 2012, 79, 19–24. [Google Scholar] [CrossRef]
- Bayandala; Mishanbieke, J. Study on the effect of distance restriction on the survival of Malus Sieversii seedlings in Xinjiang. Hunan Agric. Sci. 2019, 11, 75–77. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Species | Total Visits | Visits with Fruit Removal | Fruits Removed (%) | Visiting Behaviors | Consumption Time |
|---|---|---|---|---|---|
| Bos taurus | 477.33 ± 67.66 a | 315.00 ± 56.71 a | 96.67 ± 3.33 a | Passing, fruit consumption, resting | All day |
| Equus caballus caballus | 53.00 ± 17.24 ab | 36.00 ± 10.58 ab | 76.67 ± 4.41 ab | Passing, fruit consumption | All day |
| Sus scrofa | 40.00 ± 2.65 abc | 29.67 ± 2.60 ab | 93.33 ± 3.33 a | Passing, fruit consumption | 22:00–8:00 |
| Capreolus pygargus | 10.67 ± 1.76 bc | 4.33 ± 1.20 b | 15.00 ± 2.89 b | Passing, fruit consumption | 21:00–12:00 |
| Meles meles | 9.67 ± 3.48 bc | 5.00 ± 1.00 b | 8.33 ± 1.67 b | Passing, fruit consumption | 22:00–8:00 |
| Vulpes vulpes | 7.67 ± 4.18 c | 0 | 0 | Passing | Not |
| Treatment | Habitat | Emergence (%) | Survival (%) |
|---|---|---|---|
| C | Forest understory | 24.45 ± 2.22 | 55.56 ± 5.56 |
| W | Forest edges | 36.67 ± 1.92 * | 100.00 ± 0 ** |
| Forest gaps | 53.33 ± 1.92 ** | 100.00 ± 0 ** | |
| Forest understory | 27.78 ± 1.11 | 44.44 ± 9.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, S.; Xu, J.; Lv, Y.; Shi, X.; Tan, D. Evaluation of the Potential Seed Dispersal Effectiveness of Malus sieversii (Lebed.) M. Roem. by Cattle. Diversity 2023, 15, 1205. https://doi.org/10.3390/d15121205
Bai S, Xu J, Lv Y, Shi X, Tan D. Evaluation of the Potential Seed Dispersal Effectiveness of Malus sieversii (Lebed.) M. Roem. by Cattle. Diversity. 2023; 15(12):1205. https://doi.org/10.3390/d15121205
Chicago/Turabian StyleBai, Shilin, Jiang Xu, Yaya Lv, Xiaojun Shi, and Dunyan Tan. 2023. "Evaluation of the Potential Seed Dispersal Effectiveness of Malus sieversii (Lebed.) M. Roem. by Cattle" Diversity 15, no. 12: 1205. https://doi.org/10.3390/d15121205
APA StyleBai, S., Xu, J., Lv, Y., Shi, X., & Tan, D. (2023). Evaluation of the Potential Seed Dispersal Effectiveness of Malus sieversii (Lebed.) M. Roem. by Cattle. Diversity, 15(12), 1205. https://doi.org/10.3390/d15121205