
Divergence of Seed Mass in Riparian Plant Communities Dominated by Filtering and Random Processes

Abstract
:1. Introduction
2. Materials and Methods
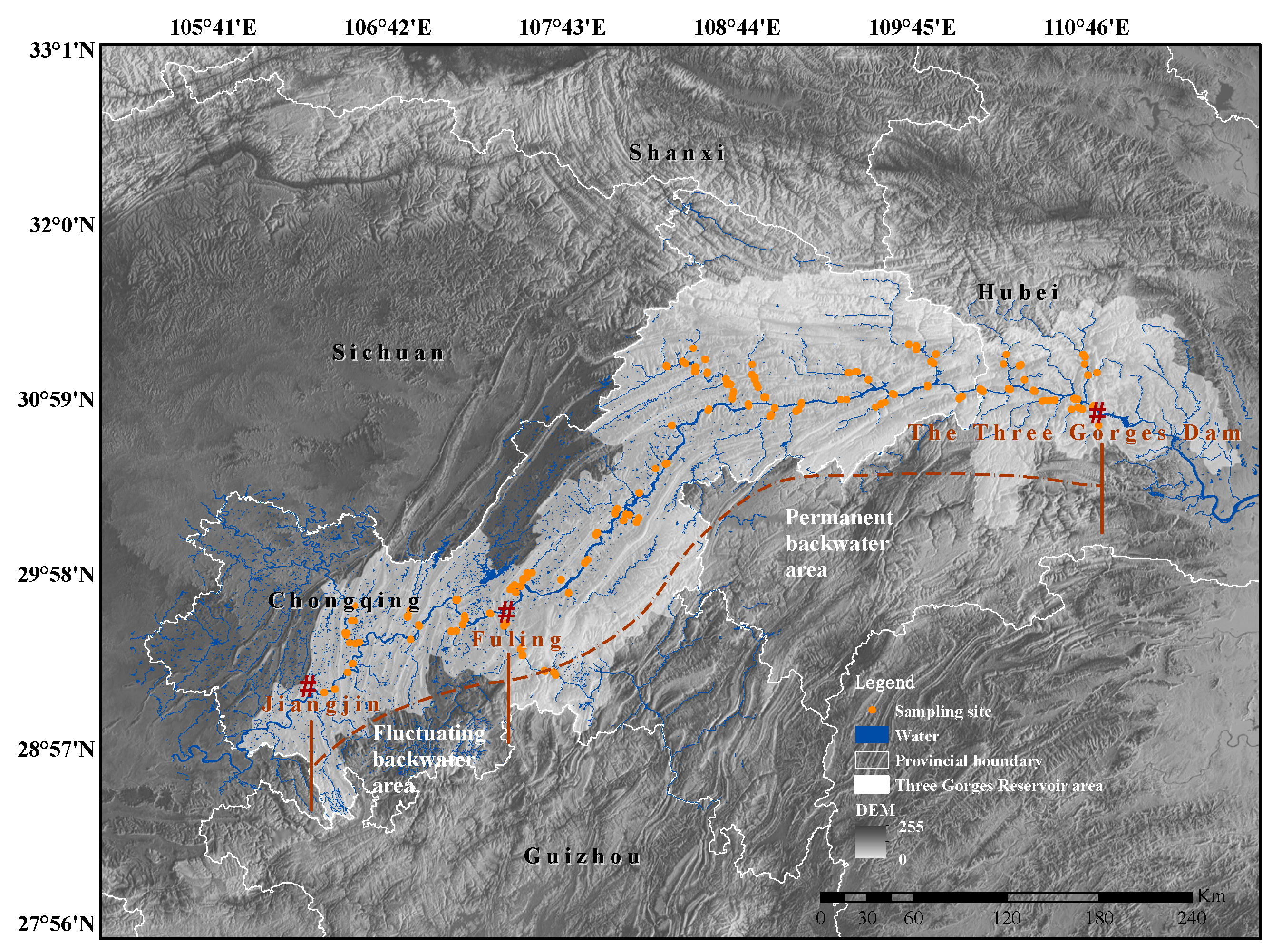
2.1. Area Description
2.2. Survey
2.3. Source of Plant Traits Data
2.4. Methods
2.4.1. Taxonomic, Phylogenetic, and Trait Diversity
2.4.2. Null Model
2.4.3. Data Analysis
3. Results
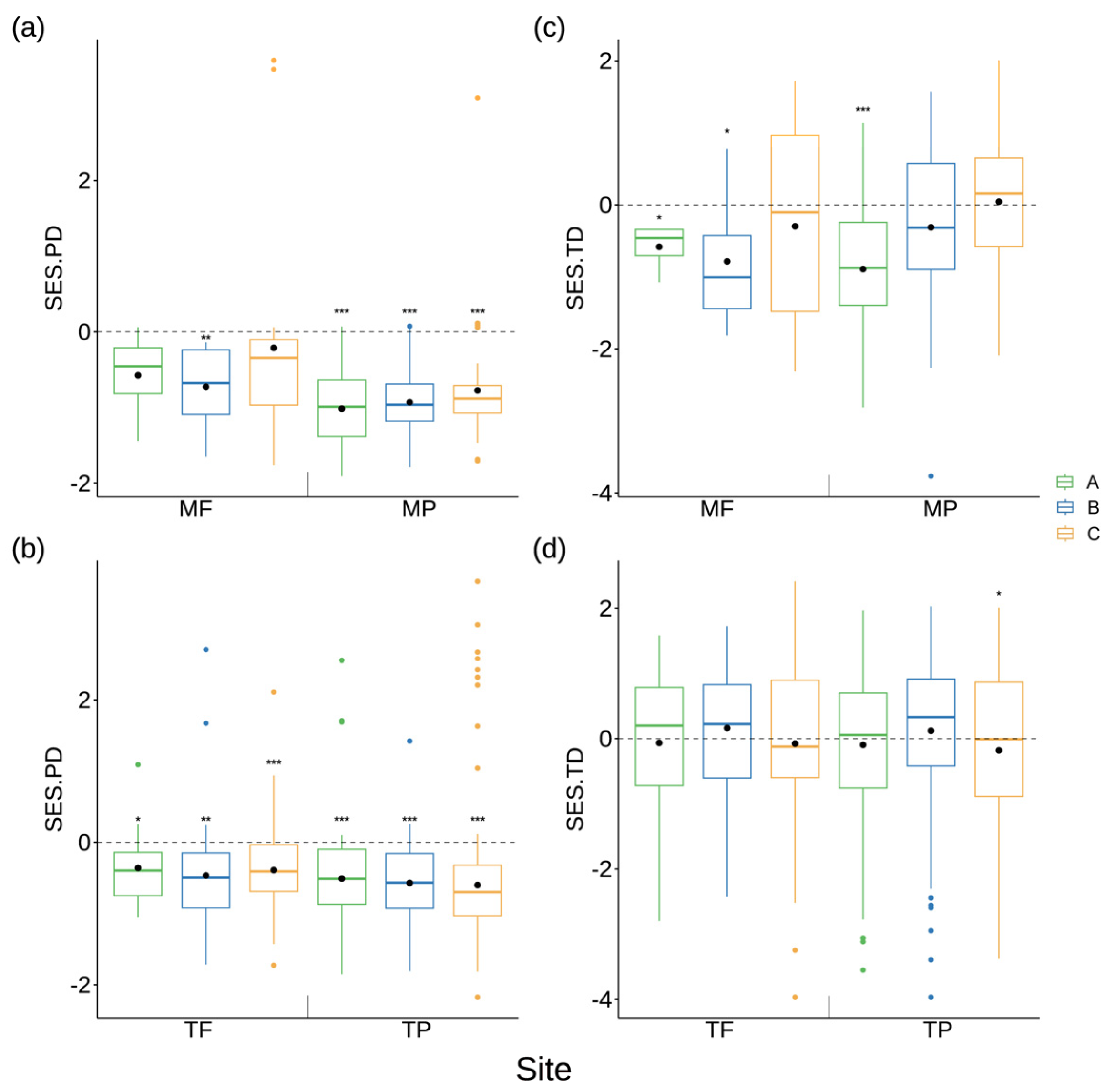
3.1. Taxonomic, Phylogenetic and Trait Diversity within the Riparian Plant Communities
3.2. Response of Plant Community Assembly to Flooding Gradient in Various Geographic Locations Using Phylogeny and Trait Analyses
3.3. Aggregation and Dispersion of Single Plant Trait in Conjunction with the Process of Community Assembly
3.4. Associations between Variation in Seed Mass and Diversity of Taxonomy and Phylogeny
4. Discussion
4.1. Strong Environmental Selection on Taxonomic and Trait Diversity within Riparian Communities in the Permanent Backwater Area of the Tributaries
4.2. Environmental Filtering and Random Processes Are the Main Assembly Rules Structuring the Riparian Plant Communities
4.3. Significant Divergence of Seed Mass in Riparian Plant Communities Dominated by Filtering and Random Assembly Processes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- McDonald, R.I.; Weber, K.F.; Padowski, J.; Boucher, T.; Shemie, D. Estimating Watershed Degradation over the Last Century and Its Impact on Water-Treatment Costs for the World’s Large Cities. Proc. Natl. Acad. Sci. USA 2016, 113, 9117–9122. [Google Scholar] [CrossRef] [PubMed]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.A.; Kembel, S.W. The Merging of Community Ecology and Phylogenetic Biology. Ecol. Lett. 2009, 12, 693–715. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, P.; Prieto, M.; Aragón, G.; Escudero, A.; Martínez, I. Critical Predictors of Functional, Phylogenetic and Taxonomic Diversity Are Geographically Structured in Lichen Epiphytic Communities. J. Ecol. 2019, 107, 2303–2316. [Google Scholar] [CrossRef]
- Loranger, J.; Munoz, F.; Shipley, B.; Violle, C. What Makes Trait–Abundance Relationships When Both Environmental Filtering and Stochastic Neutral Dynamics Are at Play? Oikos 2018, 127, 1735–1745. [Google Scholar] [CrossRef]
- Bernard-Verdier, M.; Navas, M.-L.; Vellend, M.; Violle, C.; Fayolle, A.; Garnier, E. Community Assembly along a Soil Depth Gradient: Contrasting Patterns of Plant Trait Convergence and Divergence in a Mediterranean Rangeland. J. Ecol. 2012, 100, 1422–1433. [Google Scholar] [CrossRef]
- Shipley, B.; Vile, D.; Garnier, É. From Plant Traits to Plant Communities: A Statistical Mechanistic Approach to Biodiversity. Science 2006, 314, 812–814. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carboni, M.; Si, X.; Tatsumi, S. Do Traits and Phylogeny Support Congruent Community Diversity Patterns and Assembly Inferences? J. Ecol. 2019, 107, 2065–2077. [Google Scholar] [CrossRef]
- de Bello, F.; Lavorel, S.; Hallett, L.M.; Valencia, E.; Garnier, E.; Roscher, C.; Conti, L.; Galland, T.; Goberna, M.; Májeková, M.; et al. Functional Trait Effects on Ecosystem Stability: Assembling the Jigsaw Puzzle. Trends Ecol. Evol. 2021, 36, 822–836. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Cavender-Bares, J.; Tilman, D.; Oakley, T.H. Using Phylogenetic, Functional and Trait Diversity to Understand Patterns of Plant Community Productivity. PLoS ONE 2009, 4, e5695. [Google Scholar] [CrossRef]
- Liu, C.; Wolter, C.; Xian, W.; Jeschke, J.M. Most Invasive Species Largely Conserve Their Climatic Niche. Proc. Natl. Acad. Sci. USA 2020, 117, 23643–23651. [Google Scholar] [CrossRef]
- Vetaas, O.R.; Grytnes, J.-A.; Bhatta, K.P.; Hawkins, B.A. An Intercontinental Comparison of Niche Conservatism along a Temperature Gradient. J. Biogeogr. 2018, 45, 1104–1113. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation Evaluation and Phylogenetic Diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Flynn, D.; Mirotchnick, N.; Jain, M.; Palmer, M.; Naeem, S. Functional and Phylogenetic Diversity as Predictors of Biodiversity- Ecosystem-Function Relationships. Ecology 2011, 92, 1573–1581. [Google Scholar] [CrossRef]
- Macarthur, R.; Levins, R. The Limiting Similarity, Convergence, and Divergence of Coexisting Species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Watkins, A.J.; Wilson, J.B. Local Texture Convergence: A New Approach to Seeking Assembly Rules. Oikos 2003, 102, 525–532. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and Community Ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef]
- Pearse, W.D. The Phylogenetic Structure of Ecological Communities under Change. 2012. Available online: https://core.ac.uk/download/pdf/76994895.pdf (accessed on 10 October 2023).
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community Assembly, Coexistence and the Environmental Filtering Metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Emerson, B.C.; Gillespie, R.G. Phylogenetic Analysis of Community Assembly and Structure over Space and Time. Trends Ecol. Evol. 2008, 23, 619–630. [Google Scholar] [CrossRef]
- Culmsee, H.; Leuschner, C. Consistent Patterns of Elevational Change in Tree Taxonomic and Phylogenetic Diversity across Malesian Mountain Forests. J. Biogeogr. 2013, 40, 1997–2010. [Google Scholar] [CrossRef]
- Leibold, M.A.; Chase, J.M.; Ernest, S.K.M. Community Assembly and the Functioning of Ecosystems: How Metacommunity Processes Alter Ecosystems Attributes. Ecology 2017, 98, 909–919. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Ma, M.; Ding, Z.; Wu, S.; Jia, W.; Chen, Q.; Yi, X.; Zhang, J.; Li, X.; et al. Dam-Induced Difference of Invasive Plant Species Distribution along the Riparian Habitats. Sci. Total Environ. 2022, 808, 152103. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, L.; Cao, Q.; Niu, Z.; Dai, X. Regional Climate Change and Possible Causes over the Three Gorges Reservoir Area. Sci. Total Environ. 2023, 903, 166263. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yuan, X.; Wu, S.; Zhang, G.; Liu, H.; Zhou, L.; Zhang, M. Effects of River Damming on Ground-Dwelling Arthropods along Riparian–Upland Habitats. Ecohydrology 2019, 12, e2073. [Google Scholar] [CrossRef]
- Zheng, J.; Arif, M.; Zhang, S.; Yuan, Z.; Zhang, L.; Li, J.; Ding, D.; Li, C. Dam Inundation Simplifies the Plant Community Composition. Sci. Total Environ. 2021, 801, 149827. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zeng, B.; Huang, W.; Xu, S.; Lei, S. Effects of the Three Gorges Dam on Preupland and Preriparian Drawdown Zones Vegetation in the Upper Watershed of the Yangtze River, P.R. China. Ecol. Eng. 2012, 44, 123–127. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, M.; Ran, Y.; Yi, X.; Wu, S.; Huang, P. Disentangling the Effects of Edaphic and Vegetational Properties on Soil Aggregate Stability in Riparian Zones along a Gradient of Flooding Stress. Geoderma 2021, 385, 114883. [Google Scholar] [CrossRef]
- Van Der Pijl, L. Principles of Dispersal in Higher Plants; Springer: Berlin/Heidelberg, Germany, 1982; ISBN 978-3-642-87927-2. [Google Scholar]
- Fukami, T.; Martijn Bezemer, T.; Mortimer, S.R.; Putten, W.H. Species Divergence and Trait Convergence in Experimental Plant Community Assembly. Ecol. Lett. 2005, 8, 1283–1290. [Google Scholar] [CrossRef]
- Lengyel, S.; Gove, A.D.; Latimer, A.M.; Majer, J.D.; Dunn, R.R. Convergent Evolution of Seed Dispersal by Ants, and Phylogeny and Biogeography in Flowering Plants: A Global Survey. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 43–55. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional Richness, Functional Evenness and Functional Divergence: The Primary Components of Functional Diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Wagner, H. Vegan: Community Ecology Package; R Package Version. 2.0–10; University of Helsinki; CRAN: Helsinki, Finland, 2013. [Google Scholar]
- Pearse, W.D.; Cadotte, M.W.; Cavender-Bares, J.; Ives, A.R.; Tucker, C.M.; Walker, S.C.; Helmus, M.R. Pez: Phylogenetics for the Environmental Sciences. Bioinformatics 2015, 31, 2888–2890. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 2014, 1, 1–12. Available online: https://www.researchgate.net/publication/312463190_FD_Measuring_functional_diversity_from_multiple_traits_and_other_tools_for_functional_ecology (accessed on 15 July 2023).
- Jin, Y.; Qian, H.V. PhyloMaker2: An Updated and Enlarged R Package That Can Generate Very Large Phylogenies for Vascular Plants. Plant Divers. 2022, 44, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R Tools for Integrating Phylogenies and Ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Bartomeus, I. Fundiv: Analyzing Functional Trait Diversity 2020. Available online: https://github.com/ibartomeus/fundiv (accessed on 15 July 2023).
- Chen, Z.; Yuan, X.; Roß-Nickoll, M.; Hollert, H.; Schäffer, A. Moderate Inundation Stimulates Plant Community Assembly in the Drawdown Zone of China’s Three Gorges Reservoir. Environ. Sci. Eur. 2020, 32, 79. [Google Scholar] [CrossRef]
- Zhu, K.; Chen, Y.; Zhang, S.; Lei, B.; Yang, Z.; Huang, L. Vegetation of the Water-Level Fluctuation Zone in the Three Gorges Reservoir at the Initial Impoundment Stage. Glob. Ecol. Conserv. 2020, 21, e00866. [Google Scholar] [CrossRef]
- Rong, S.; Weiqiong, D.; Xiuming, L. Distribution Pattern of Riparian Vegetation along Donghe River, a Tributary of the Three Gorges Reservoir Area. Chin. J. Ecol. 2015, 34, 2733. [Google Scholar]
- Fukami, T. Historical Contingency in Community Assembly: Integrating Niches, Species Pools, and Priority Effects. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 1–23. [Google Scholar] [CrossRef]
- Götzenberger, L.; De Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M.; et al. Ecological Assembly Rules in Plant Communities-Approaches, Patterns and Prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef]
- Ran, Y.; Wu, S.; Chen, C.; Sun, X.; Huang, P.; Ma, M.; Yi, X. Shift from Soil Chemical to Physical Filters in Assembling Riparian Floristic Communities along a Flooding Stress Gradient. Sci. Total Environ. 2022, 844, 157116. [Google Scholar] [CrossRef]
- Marteinsdóttir, B.; Svavarsdóttir, K.; Thórhallsdóttir, T.E. Multiple Mechanisms of Early Plant Community Assembly with Stochasticity Driving the Process. Ecology 2018, 99, 91–102. [Google Scholar] [CrossRef]
- Simpson, K.J.; Atkinson, R.R.L.; Mockford, E.J.; Bennett, C.; Osborne, C.P.; Rees, M. Large Seeds Provide an Intrinsic Growth Advantage That Depends on Leaf Traits and Root Allocation. Funct. Ecol. 2021, 35, 2168–2178. [Google Scholar] [CrossRef]
- Rees, M.; Venable, D.L. Why Do Big Plants Make Big Seeds? J. Ecol. 2007, 95, 926–936. [Google Scholar] [CrossRef]
- Liu, J.; Lin, F.; Shi, S.; Ayi, Q.; Liu, S.; Zeng, B. Effects of Water Level Regulation on the Seed Germination and Production of Annual Plant Xanthium Sibiricum in the Water-Level-Fluctuating-Zone of Three Gorges Reservoir. Sci. Rep. 2017, 7, 5056. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Queiroz, C.R.R.; Wilson, T.M.; Sousa, D.E.R.; Castro, M.B.; Saravia, A.; Lee, S.T.; Armién, A.G.; Barros, S.S.; Riet-Correa, F. Endemic Xanthium Strumarium Poisoning in Cattle in Flooded Areas of the Araguari River, Minas Gerais, Brazil. Toxicon 2021, 200, 23–29. [Google Scholar] [CrossRef]
- Comparative Ecology of Seed Size and Dispersal. Phil. Trans. R. Soc. Lond. B 1996, 351, 1309–1318. [CrossRef]
- Turnbull, L.A.; Philipson, C.D.; Purves, D.W.; Atkinson, R.L.; Cunniff, J.; Goodenough, A.; Hautier, Y.; Houghton, J.; Marthews, T.R.; Osborne, C.P.; et al. Plant Growth Rates and Seed Size: A Re-Evaluation. Ecology 2012, 93, 1283–1289. [Google Scholar] [CrossRef]
- Metz, J.; Liancourt, P.; Kigel, J.; Harel, D.; Sternberg, M.; Tielbörger, K. Plant Survival in Relation to Seed Size along Environmental Gradients: A Long-Term Study from Semi-Arid and Mediterranean Annual Plant Communities. J. Ecol. 2010, 98, 697–704. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Data Type | Data Classification Criteria/Units |
|---|---|---|
| Life form | Categorical variable | Phanerophyte, Chamaephyte, Hemicryptophyte, Cryptophyte, Therophyte |
| Plant height | Continuous variable | cm |
| Fruit type | Categorical variable | Berry, Pepo, Hesperidium, Drupe, Pome, Follicles, Legumes, Capsule, Silique, Achene, Grain, Samara, Schizocarp, Nut, Aggregate fruit, Collective fruit |
| Seed dispersal mode | Categorical variable | Endozoochory, Ballochory, Exozoochory, Myrmecochory, Anemochory, Hydrochory, Autochory |
| Pollination | Categorical variable | Wind pollination, Biological pollination, Self-pollination, Water pollination |
| Seed mass | Continuous variable | thousand seed weight, mg |
| Regenerative strategy | Categorical variable | W: numerous widely dispersed seeds or spores; V: lateral vegetative spread (offshoots remaining attached to parent for a considerable period); SV: seasonal regeneration by vegetative means; S: seasonal regeneration from seed; B: persistent seed bank of buried seeds or spores |
| Site | ZONE | Correlation between Seed Mass and Taxonomic Diversity | Correlation between Seed Mass and Phylogenetic Diversity |
|---|---|---|---|
| MF | A | 0.4 | 0.8 |
| MF | B | 0.558 | 0.914 ** |
| MF | C | 0.814 ** | 0.929 ** |
| MP | A | 0.681 ** | 0.968 ** |
| MP | B | 0.713 ** | 0.961 ** |
| MP | C | 0.605 ** | 0.944 ** |
| TF | A | 0.634 ** | 0.961 ** |
| TF | B | 0.812 ** | 0.974 ** |
| TF | C | 0.819 ** | 0.966 ** |
| TP | A | 0.841 ** | 0.987 ** |
| TP | B | 0.660 ** | 0.983 ** |
| TP | C | 0.670 ** | 0.956 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Huang, J.; Zou, H.; Ma, M. Divergence of Seed Mass in Riparian Plant Communities Dominated by Filtering and Random Processes. Diversity 2023, 15, 1164. https://doi.org/10.3390/d15121164
Wang W, Huang J, Zou H, Ma M. Divergence of Seed Mass in Riparian Plant Communities Dominated by Filtering and Random Processes. Diversity. 2023; 15(12):1164. https://doi.org/10.3390/d15121164
Chicago/Turabian StyleWang, Wanyu, Jinxia Huang, Hang Zou, and Maohua Ma. 2023. "Divergence of Seed Mass in Riparian Plant Communities Dominated by Filtering and Random Processes" Diversity 15, no. 12: 1164. https://doi.org/10.3390/d15121164
APA StyleWang, W., Huang, J., Zou, H., & Ma, M. (2023). Divergence of Seed Mass in Riparian Plant Communities Dominated by Filtering and Random Processes. Diversity, 15(12), 1164. https://doi.org/10.3390/d15121164