1. Introduction
The emergence of the early Neolithic in southwestern Transylvania occurred after the critical moment of the 8.2 ka Cold Event period [
1,
2,
3,
4], when it seems that a major climatic change prevented, for a time, the northward advance of the agro-pastoral groups from the central part of the Balkans. After overcoming this climatic crisis (extreme cold oscillation of the Holocene), an event that ended around 6000 cal BC, we witness the spread of Starčevo-Criș human groups in the northern part of the Lower Danube Basin [
5].
Regarding southwestern Transylvania (western Romania), in the area located between the cities Deva and Simeria, in Hunedoara County, at a distance of ca. 15 km, in an area that does not exceed 150 km
2, were identified, during the last few years, almost 12 archaeological sites which can be linked to the earliest presences of Starčevo-Criș cultural complexes, due to the architecture of the features, material culture, and relative or absolute chronology data [
6]. The same communities had a decisive role regarding the occupation of the karst systems; no fewer than six caves from the area of Hunedoara contain archaeological layers or stray finds, with materials characteristic to the first Neolithic groups: Cerișor-Peștera Cauce, Ohaba Ponor-Peștera Bordul Mare, Nandru-Peștera Curată, Nandru-Peștera Spurcată, Balșa-Peștera Dosul Dobârlesei, and Crăciunești-Peștera Balogu/Szabó József [
5].
The appearance of Early Neolithic communities in southwestern Transylvania, as well as the establishment of the same groups in the Mureș Corridor, were motivated by several factors, such as the search for areas suitable for raising animals and grazing, as well as for fertile lands for plant cultivation, some of which were located in river meadows. Other activities (hunting, fishing, collecting molluscs, etc.) certainly had the role of supplementing the nutritional needs of the first agricultural communities.
The present study seeks to answer, from an interdisciplinary perspective, the questions regarding the beginnings of neolithization in southwestern Transylvania and to identify what the subsistence strategies were, valuing the archaeozoological and archaeobotanical data from the Starčevo-Criș site Șoimuș-Teleghi (Șoimuș commune).
Phytoliths, siliceous microresidues that form in plants, are utilised in this paper, in order to identify the vegetal resources that the neolithic communities from Șoimuș-Teleghi could have exploited. Additionally, the study will highlight, at least partially, the local vegetation signal. These plant microremains can be preserved both in modern, as well as fossilized, soil and sediments, and also archaeological contexts [
7,
8]. If for the neolithic sites of Romania, there are numerous archaeozoological studies [
9,
10,
11,
12,
13,
14,
15,
16,
17,
18,
19,
20], there are very few studies that focus on the phytolith analysis for sites from the same period [
21,
22,
23]. Very few are also studies that integrate both archaeozoological and phytolith data [
24,
25,
26,
27].
For this chronological interval and for the same area (ca. 15 km east from the site Şoimuş-Teleghi), another site was studied from a bioarchaeological point of view, namely, the site from Rapoltu Mare-Șeghi, Rapoltu Mare commune, to which we will compare from this point forward. The archaeological site of Şeghi, “also known by the names Sedi or Siediu, is located at about 3 km north from the city of Simeria and at about 1.0 km, on the southwest direction, from the village of Rapoltu Mare and from the confluence of the rivers Strei and Mureş” [
26]. The archaeological research supplied information on the chronological and cultural horizons from the Early Neolithic to the Middle Ages. “The faunal material sampled for 14C dating was analysed at “Horia Hulubei” National Institute for Research and Development in Physics and Nuclear Engineering (IFIN-HH), Bucharest-Măgurele (Romania). The obtained result gave the following date: 6897 ± 28 BP (Ro-AMS 11–11); the calibrated date (σ = 2) is 5841–5723 cal BC, with a probability of 95.4%” [
26]. New (unpublished) radiometric dates, obtained in the same 14C laboratory, show that the beginning of the site could be placed at the start of the 6th millennium cal BC.
2. Study Area and the Archaeological Context
2.1. Study Area
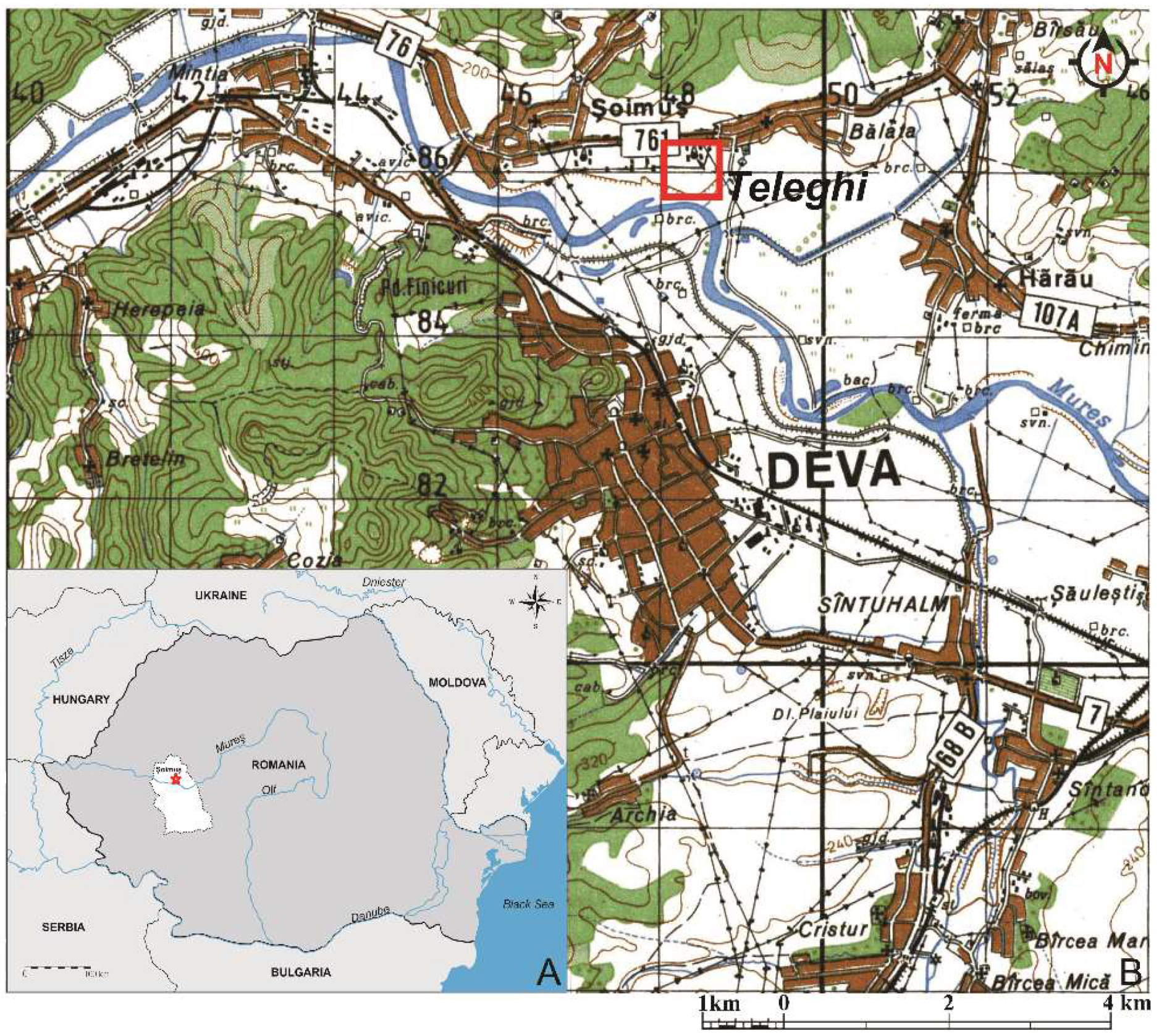
The archaeological site from Teleghi (Lat. 45°54′36.1″ N, Long. 22°54′33.0″ E, Alt. 187.8 m) is located about 2 km north of the municipality of Deva and 1.5 km southeast of Șoimuș (
Figure 1), on a natural terrace bridge dating from Pleistocene (
Figure 2), on the right side of Mureș River, with an altitude of 5–10 m above the meadow [
28]. In relation to the main water course, the site from Teleghi is located at the entrance to the depression area, with a gorge aspect, of the Mureș River valley [
29,
30,
31].
In the archaeological literature, the site from Teleghi is mentioned in the second half of the last century [
32], but it came to be known much better as a result of the rescue archaeological excavations carried out in 2011, as part of the Deva–Orăștie highway project [
33]. Both the older and more recent archaeological diggings indicate the existence of human settlements within the boundaries of the Șoimuş commune, whose chronology starts from the early Neolithic and ends during the Middle Ages [
32,
33,
34].
The fact that the human groups from prehistory, antiquity, or the Middle Ages frequented the terrace, which we currently know by the toponym Teleghi, was also a consequence of the easy access to the raw material sources. Thus, here we can mention the Upper Cretaceous sandstones and marls, also known by the name “Strate de Deva” [
35], found in the structure of the terrace. Similar rocks, in the form of deposits, are also found on the left and right side of the Mureș River valley, at the north-western edge of Deva municipality and Șoimuș village [
28]. About 2 km south of the archaeological point, Teleghi, is located the northern edge of the Poiana Ruscă Mountains, composed of volcanic rocks such as grey andesites. Such raw materials were exploited by early Neolithic communities, as shown by the geo-archaeological studies conducted on some artefacts from the oldest dwellings belonging to the Neolithic [
36,
37]. Additionally, the presence of sodium-chloride springs in the Deva–Hărău area [
28,
38,
39], at a distance between 2–4 km from the settlements of Șoimuș-Teleghi, represented an advantage for the prehistoric communities, especially since the southwestern area of Transylvania is not as rich in salt sources in comparison to the central area of the same historical province.
2.2. Archaeological Context
The first result of the rescue archaeological excavations carried out in 2011 on the section of the A1 Deva–Orăștie highway, at the southern edge of the site from Șoimuş-Teleghi, was represented by the identification and research of approximately 1000 archaeological features in an area of approximately 20,000 m
2 [
33]. The same archaeological diggings revealed that the highest intensity of habitation occurred during the Neolithic and the Bronze Ages, with more than two-thirds of the excavated features belonging to the mentioned chronological sequences.
Very interesting was the discovery, during the mechanical excavation of the site Șoimuș-Teleghi, of an early Neolithic settlement, spread over an area of approximately 3000 m
2, which is chronologically placed at the border between the 7th and 6th millennia BC [
6,
28]—a chronological interval still partially unknown, in the southwestern area of Transylvania, in comparison to later periods. From the excavated features, two possible dwellings were distinguishable: C.18a and C.18b, discovered at km 30 + 260–30 + 280 of the A1 Deva–Orăștie Highway (
Figure 2).
Feature C.18a. The oldest sample of faunal materials was discovered in the archaeological feature C.18a, at depths of 0.75–0.80 m–1–1.30 m, similar in some aspects to the architecture of pit-houses [
40,
41]. The architecture of C.18a was quite simple: it had an irregular shape, with unequal sides, the orientation of the feature being on the northwest-southeast axis. The eastern side of the pit-hut was affected by several later archaeological features, C.165a, C.165b, and C.29. Regarding the access to the Neolithic pit-feature, the excavation revealed the existence of “steps” on the western side, the short side of C.18a. The same archaeological feature had, on the south side, a bench of clay, in the middle of which was found a fragmentary, globular vessel in a small alveolus, which was reconstructed.
The archaeozoological material, sometimes well preserved, other times strongly fragmented, was spread over the entire surface of the mentioned pit-dwelling, together with numerous ceramic fragments and carved and polished stone tools [
36,
42].
In the north-western corner of C.18a, at a higher point, practically on a shelf of clay, was identified a possible area where a fire hearth worked as a result of the archaeological capture of an ash layer with very little charcoal but also due to the presence of traces from the clay surface of the hearth. On a slightly discontinuous layer of ash, not thicker than 1 cm, with a diameter of about 1 m, several costal segments of
Ovis aries/Capra hircus were caught, which, at first sight, would seem to come from the same individual, although their arrangement was not in anatomical connection. Additionally, we have to mention that some pieces of bone, based on the processing traces, were assigned to the category of tools [
43,
44,
45].
Feature C.18b. The feature was identified during the mechanical excavation of the terrace from Teleghi, at approximately a 0.50 m depth from the stepping layer of 2011, consisting of arable soil. The limits of C.18b were established based on the spread of the archaeological materials, on an area of approximately 5 × 5 m, represented by ceramics, bones, hearth fragments, and lithic pieces such as river stones and sandstone fragments. Such concentrations of archaeological material were encountered in countless situations in the early Neolithic in southeastern Europe. Thus, it is believed that some features of this kind could represent the imprint of former surface dwellings [
40], which, after abandonment, were probably transformed into areas for household waste disposal.
At the same time, the archaeological excavation revealed that the thickness of the archaeological deposits was no more than 0.20–0.25 m, but also that below C.18b, another feature was outlined, an older one, noted C.18a. Additionally, here, we emphasize that C.18b was sectioned in the eastern part by a Vinča pit-feature (C.165b) and by a ditch from the Bronze Age (C.29), and on the western side by two pits from the Bronze Age (C.17 and C.47).
In this case, also, it was noted that the osteological material is both fragmentary and very well preserved. Bone tools [
43,
44,
45], as well as objects from polished stone [
36,
42], were determined also from this archaeological context with the number C.18b, which overlaps the pit of C.18a.
3. Materials and Methods
Radiocarbon dating. From C.18a and C.18b were taken osteological materials, so that they could be subjected to 14C dating (
Table 1). The first radiocarbon results obtained at IFIN-HH Bucharest-Măgurele provided the following dates for the earliest complexes from Șoimuș-Teleghi, which we will present chronologically in the present paper.
Two other samples of faunal material, also selected from the C.18a and C.18b features, were analysed in the Beta Analytic laboratory, Miami, FL, USA.
Archaeozoology. The faunal samples collected from C.18a and C.18b during the archaeological digging are represented by animal remains, such as shells, teeth, antlers, cornular processes, and bones. These assemblages are composed of highly fragmented skeletal remains resulting mainly from the consumption of animal products, but also from the processing of hard materials of animal origin. The mammals’ bones show numerous traces of human activities, such as butchery cuts and splits, and burn marks but also traces left by the teeth of other animals (e.g., dogs). The archaeozoological samples were collected “by hand”, without sieving sediments, which could have caused the underestimation of small-sized species by losing skeletal small fragments.
After the faunal material preparation (e.g., washing, consolidation, marking), the anatomical and taxonomical identification of the remains was carried out, based on the comparison skeletal collection of the Laboratory of Archaeozoology from the Faculty of Biology, Alexandru Ioan Cuza University of Iași (Romania). Additionally, a taphonomical evaluation of the samples was performed by identifying traces of different origins (e.g., of anthropic nature, and those left by carnivores). Using the criterion of dentition (i.e., dental eruption and dental wear), as well as the degree of the bones’ ossification (i.e., epiphyseal closure), the estimation of slaughter ages was performed [
46,
47]. The quantification of the archaeozoological samples was performed by establishing the number of identified specimens (NISP) and the minimum number of individuals (MNI) for the identified taxa. MNI was estimated for each species, based on the most frequent skeletal element, after separating according to laterality (left/right) [
48]. Bones belonging to adult individuals were measured, according to the guide developed by A. von den Driesch [
49]. The wither height was estimated for sheep (
Ovis aries) using the coefficients of Teichert [
50].
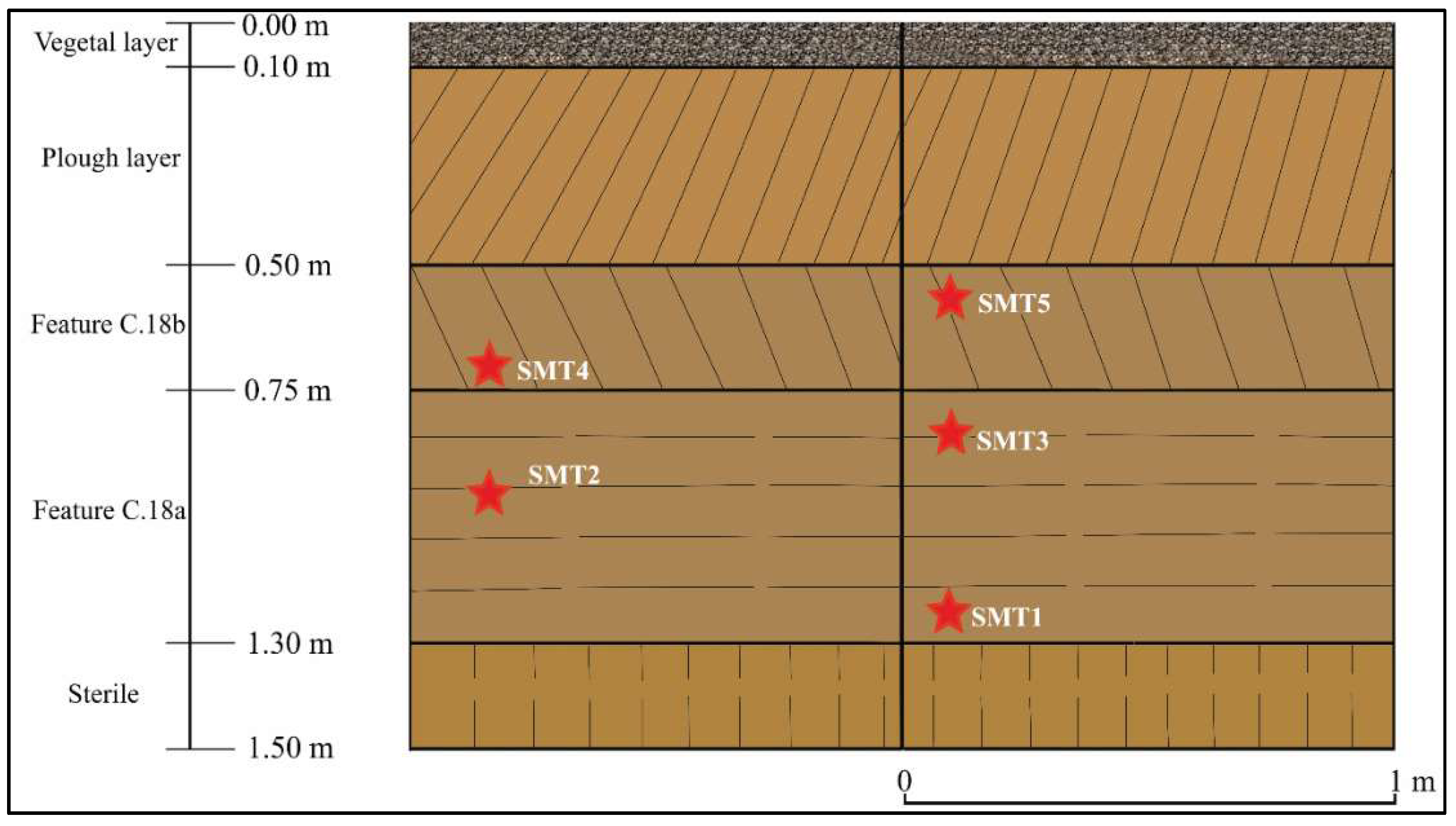
Archaeobotany. From the cultural layers were taken five sediment samples: SMT 1–3 from C.18a and SMT 4–5 from C.18b (
Figure 3 and
Figure 4). The phytolith extraction and chemical preparation were realised at the Laboratory of Bioarchaeology (Alexandru Ioan Cuza University of Iasi). The samples (almost 3 g each) were processed following a chemical protocol, adapted after Lentfer, Boyd [
51]: “clay deflocculation with distilled water under magnetic stirring, 200 μm tumbling for the removal of coarse particles; centrifugation for 2000 t. min
−1 for clay elimination; decarbonation with concentrated hydrochloric acid (33%) by heat and using an ultrasonic bath; organic matter oxidation under hot and ultrasonic action: KOH (10%), nitric acid (30%), hydrogen peroxide (30%); and phytolith densiometric separation with sodium polytungstate (density = 2.35). After cleaning, the residue was suspended in Zeiss immersion oil for mounting on glass slides” [
21]. The slides were analysed under a “Leica DMRB ™” microscope at 650× magnification. The identified phytoliths were classified according to the International Code for Phytolith Nomenclature 2.0 (International Committee on Phytolith Taxonomy) [
52]. In this study were counted at least 300 phytoliths for each sample. All identified phytoliths were included in a certain category according to ICPN [
52].
4. Results
4.1. Chronology—Occupation of Site
The first radiocarbon results obtained at IFIN-HH Bucharest–Măgurele, Romania, provided the following dates for the earliest features from Șoimuș-Teleghi, which we will present chronologically as Conventional Radiocarbon Ages (CRA values): 7356 ± 38 BP (Ro-AMS 11–2), 7216 ± 30 BP (Ro-AMS 11–4), 7130 ± 32 BP (Ro-AMS 11–1) for C.18a, 7019 ± 37 BP (Ro-AMS 11–3) for C.18b (
Table 2).
The other two faunal samples, also selected from the C.18a and C.18b features, were analysed in the Beta Analytic laboratory, Miami, FL, USA, the uncalibrated ages obtained being 7020 ± 30 BP (Beta-540964) for C.18a and 6870 ± 30 BP (Beta-540963) for C.18b (
Table 2).
The calibrated data (σ = 2) indicated for the C.18a pit-feature three distinct ranges: I. 6266 (82.9%) 6079 calBC (Ro-AMS 11–2); II. 6098 (80.7%) 6008 calBC (Ro-AMS 11–4) very close to 6066 (82.2%) 5978 calBC (Ro-AMS 11–1); III. 5987 (95.4%) 5833 calBC (Beta-540964). The first chronological interval (I) could show an in situ contamination of the samples with non-collagenous proteins or transhumification over time with humic substances already existing at the time of entering the post-depositional phase of the osteological material (which could not be removed by current laboratory procedures, leading to older apparent ages) or the (dis)placement from/in another cultural horizon. If we were to consider, even as a working hypothesis, the calibrated age (σ = 2) 6266 (82.9%) 6079 for Ro-AMS 11–2 as a plausible one, and then in C.18a we should have a horizon much older, prior to the early Neolithic, namely, “Mesolithic”. This is unlikely because we lack the archaeological contexts and lithic pieces typical of the Mesolithic in the mentioned feature, a fact also demonstrated by the archaeological excavation of about 2 hectares of the Teleghi site in 2011 [
33]. On the other hand, other (unpublished) 14C data from the same archaeological site, also from early Neolithic contexts, showed situations identical to the case of sample Ro-AMS 11–2. Some radiometric measurements of the osteological pieces indicate very “old” ages, typical of the 7th millennium cal BC, which could only be understood by an in situ contamination of the zooarchaeological materials with non-collagenous proteins or transhumification, as we have shown previously [
53,
54,
55]. The second interval (II) is much closer to archaeological contexts similar in age and characteristics of the material culture from Transylvania—such as those from Cristian I [
56,
57], Gura Baciului [
58,
59], Ocna Sibiului-Triguri [
58,
59], and Şeuşa-La Cărarea Morii [
58,
59]. Finally, the last interval (III) of C.18a should be seen through the prism of the data of C.18b, as we will see below, the calibrated absolute age representing either a moment of abandonment of C.18a or the effect of an intrusion subsequent. The last hypothesis is verified from a stratigraphic point of view, C.18a being overlapped by the pit of C.165a in the eastern part, while later both archaeological contexts were overlapped by the surface feature C.18b.
The calibrated data (σ = 2) indicated for the C. 18b feature two distinct intervals: I. 5989 (93.4%) 5802 cal BC (Ro-AMS 11–3); II. 5836 (90.2%) 5708 cal BC (Beta-540963). As shown above, the first interval (I) from C.18b is at the same chronological level as the latest date from C.18a (III), thus not excluding the belonging of the sample (Beta-540964) to C. 18b, which stratigraphically overlaps the complex C.18a. Under these circumstances, both the last interval (III) from C.18a and the earliest date from C.18b are contemporary with part of the 14C ages from Cristian I [
56,
57], but especially with the radiocarbon results obtained for the pit-dwellings from Miercurea Sibiului-Petriș [
59,
60,
61]. The second interval (II) in C.18b is at a significantly greater distance in time from the first interval (I) of the same context. This could be explained from the perspective of at least two scenarios. First of all, the sample for radiocarbon dating was taken from the surface of the archaeological complex C.18b, and, at that point, most of the archaeological pieces, due to the very high degree of fragmentation, had the appearance of “household debris”. This could explain the use of C.18b for a long period, the last stage of the feature corresponding to the idea of a space for “household waste”, from which it also emerges the possibility of the presence of later materials, which is rightly reflected by the 14C age of the second interval (II) of the feature C.18b. Secondly, it should not be overlooked that most of the archaeological materials from the surface of C.18b, such as pottery, do not indicate significant changes compared to the pieces discovered at greater depths in C.18. This is even confirmed by the results of the zooarchaeological analyses, a fact for which the age of the last interval in C.18b should be treated with great caution, new radiocarbon data being needed for the last stage of the C.18b feature.
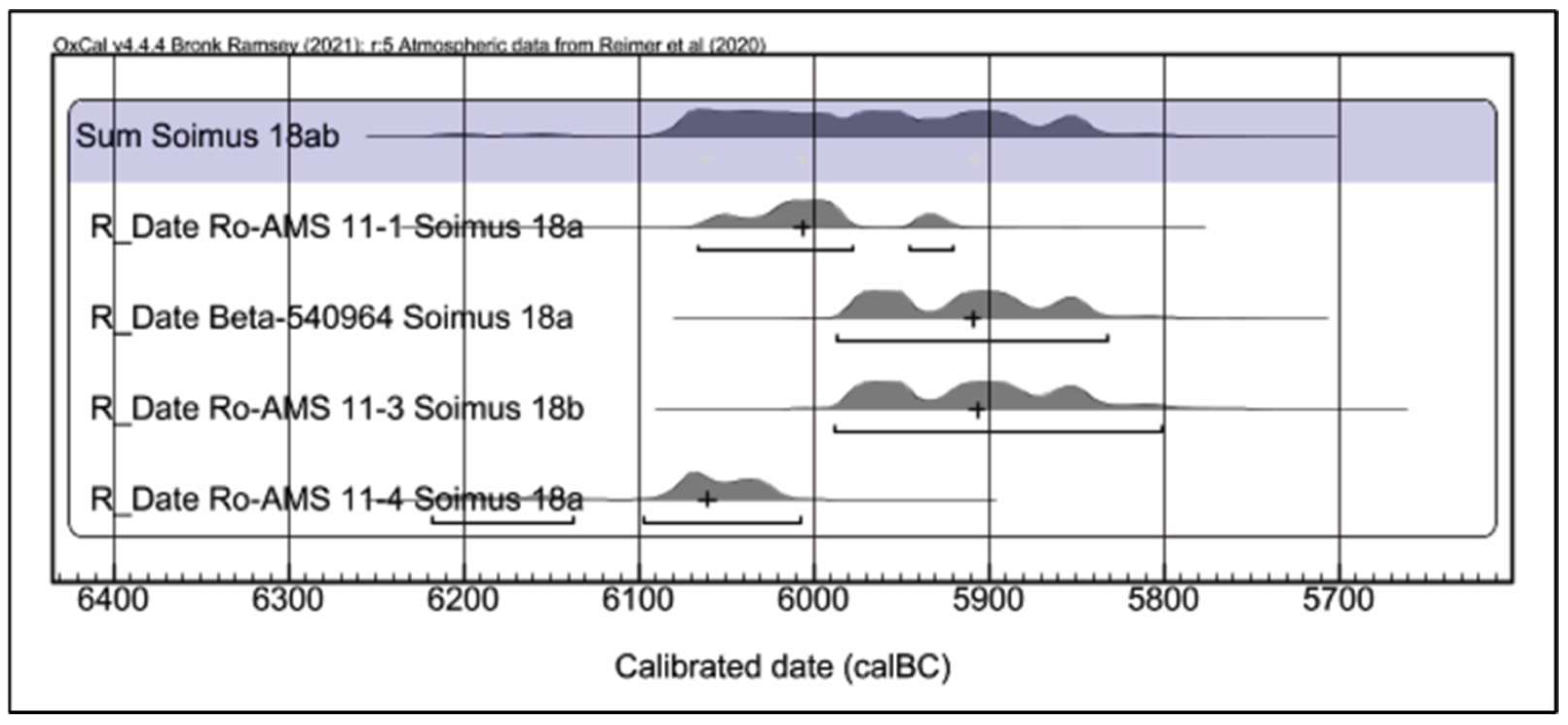
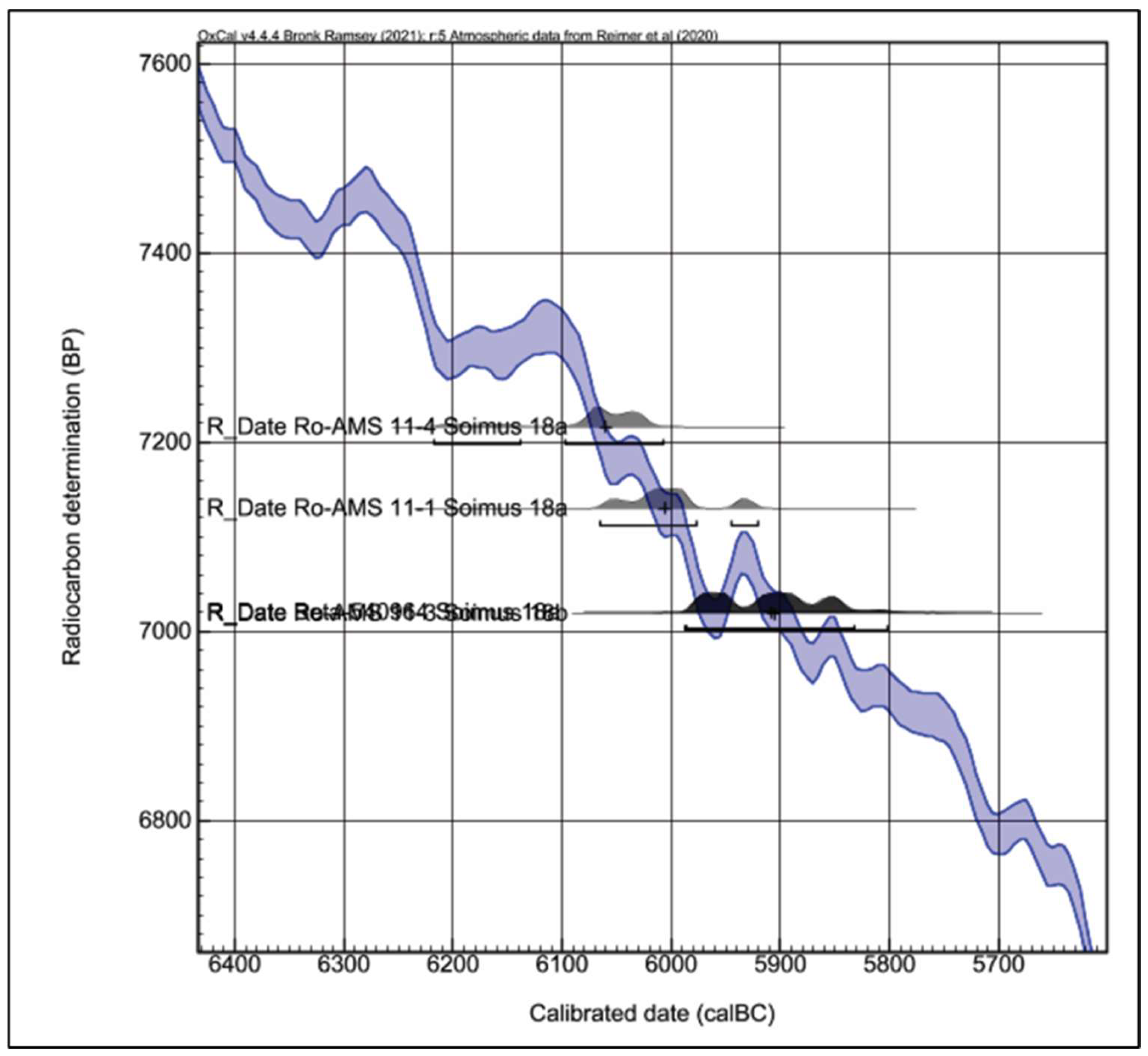
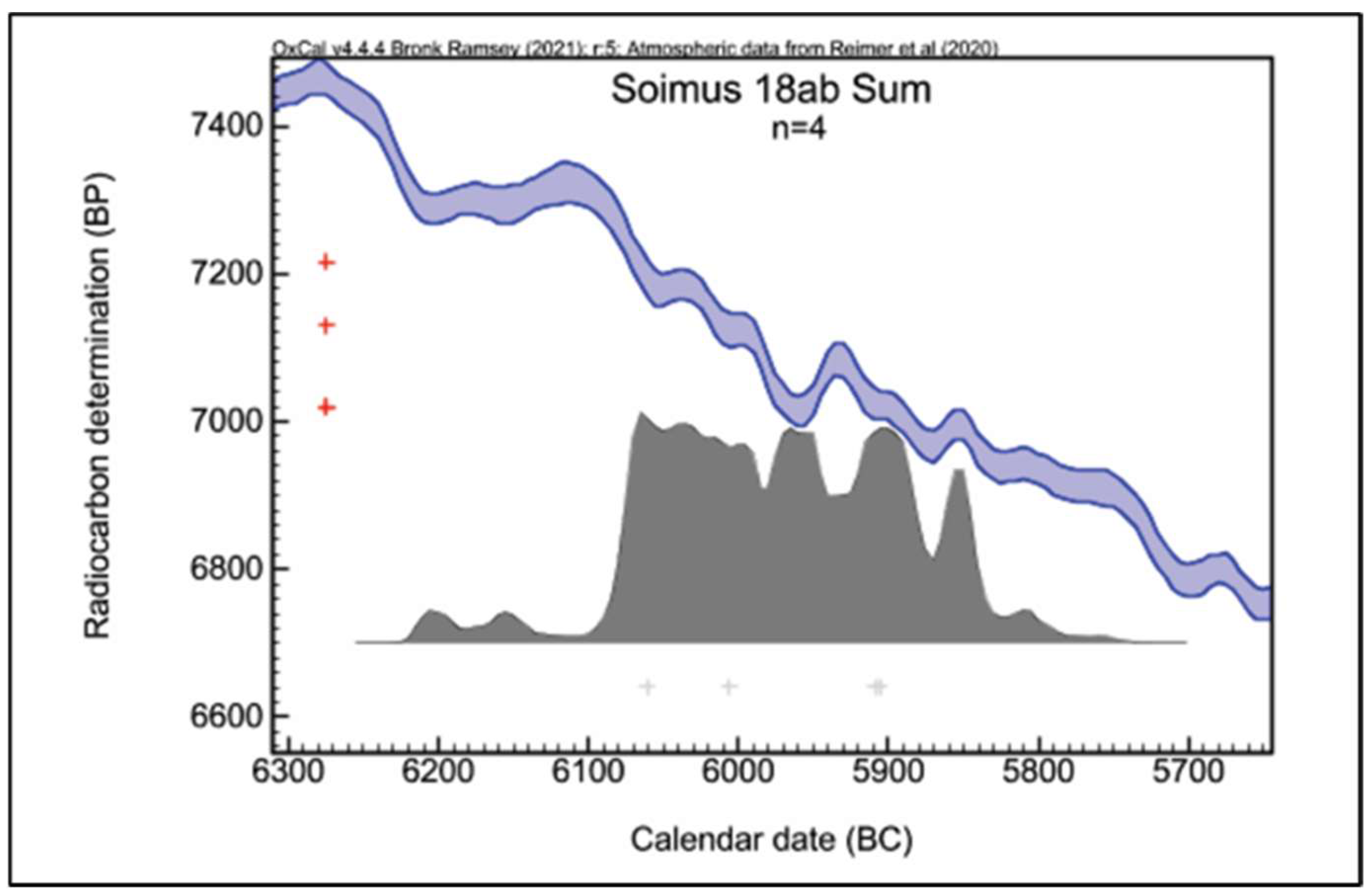
The results of applying the radiocarbon method to six samples of osteological material, all collected from C. 18a and C. 18b of the Teleghi site, eliminating the very early ages 6266 (82.9%) 6079 cal BC (Ro-AMS 11–2) or the latest 5836 (90.2%) 5708 cal BC (Beta-540963), could outline the following chronological scheme for C.18 (
Figure 5 and
Figure 6): I. Feature C. 18a corresponds to the end of the 7th millennium cal BC, respectively, and the beginning of the 6th millennium cal BC, reflected by the two close time intervals—6098 (80.7%) 6008 cal BC (Ro-AMS 11–4) and 6066 (82.2%) 5978 cal BC (Ro-AMS 11–1); II. The C. 18b feature would be placed at the beginning of the 6th millennium cal BC, as indicated by the interval 5989 (93.4%) 5802 cal BC (Ro-AMS 11–3) and the result of a sample from C.18a—5987 (95.4%) 5833 cal BC BC (Beta-540964), which would be more typical of human activity in C.18b. The sum of four usable dates from features C.18a and C.18b show that the dwelling from the feature C.18 is dated approximately between 6080 BC and 5850 BC (
Figure 5 and
Figure 7).
4.2. Archaeozoology
The studied faunal assemblages, whose quantification was carried out separately for the two features (C.18a and C.18b), are represented by household remains, as indicated by the identified butchery marks, as well as by the high degree of their fragmentation. No skeletal elements in anatomical connection were identified. Because the bones are heavily fragmented, few measurements were taken. Traces of burn, butchery (following the cutting and disarticulation process), and processing were identified on some bones; some skeletal remains have marks left by dogs’ teeth (
Table 3).
The identified remains come from domestic and wild mammals and molluscs.
Domestic mammals. In the two samples, most remains come from domestic mammals: 93.6% for C.18b and 91.6% for C.18a. The species identified in both features are cattle, sheep, goat, and pig; a dog fragment was also identified in C.18a. Pig remains are very few in both assemblages, with four fragments identified in C.18b and one fragment in C.18a. Differences are observed in terms of the proportion of cattle and sheep/goat in the two assemblages. In C.18b, cattle represent 61.6%, and sheep/goat, 30.8%. In C.18a the situation is different; sheep/goat and cattle have similar proportions (
Table 4).
A total of 640 mammalian remains were collected from C.18b, and 344 were identified at the species level. From domestic mammals came 322 fragments, which are assigned to 11 individuals, as follows: six of cattle (four mature and two immature), four of sheep/goat (two mature and two immature), and one of pig (immature) (
Table 4).
In total, 474 mammalian bone fragments were collected from C.18a, and 310 were identified at the species level. Based on 284 remains originating from domestic mammals, 12 individuals have been estimated: four of cattle (two mature and two immature), six of sheep/goat (three mature and three immature), one of pig (mature), and one of dog (mature) (
Table 4).
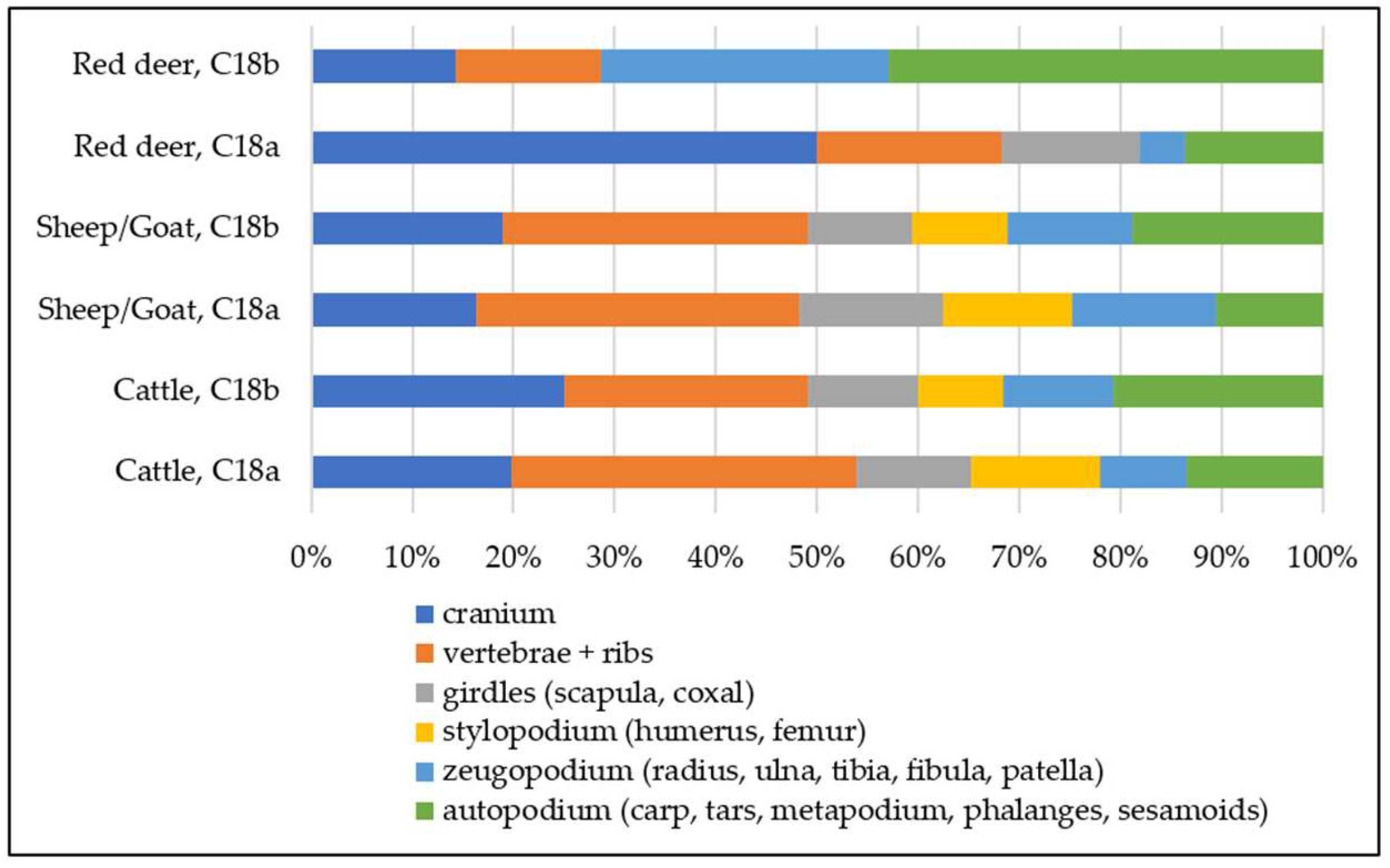
The representation of the skeletal regions in the case of the domestic species with the highest representation (i.e., cattle and sheep/goat) denotes the presence in the samples of all body parts (
Figure 8), which means that their butchering, distribution, and consumption took place within the settlement. No selection was identified for certain parts of the animal carcass.
Because the bones are heavily fragmented, few measurements were taken; due to the lack of whole long bones that generally allow the estimation of the wither height, this parameter could not be estimated for cattle, goat, pig, and dog. The wither height was estimated only for sheep (
Ovis aries), based on two astragali, the obtained values being 60.1 cm and 63.5 cm. Although wither heights obtained from astragali tend to give smaller stature information compared to other long bones, we consider that these two estimated dimensions are close to those evaluated in other early Neolithic settlements from Europe [
62].
Wild mammals. The proportion of wild mammal remains is low in the two samples. Five species were identified in C.18b and four in C.18a. The wild mammals identified are those that prefer the forest biotope (i.e., red deer, wild boar) and the forest fringe (i.e., aurochs, roe deer). We mention that we used knowledge about biotopes preferred by modern populations of identified species [
63]. In addition to the four species important in the food economy of these communities, the polecat was also identified (a fragment in C.18b), probably not having a significant contribution to the community’s diet. Most wild mammal remains are from red deer (14 fragments from C.18b and 22 from C.18a). The other species have very low frequencies within the samples (
Table 4).
Based on 22 skeletal remains of wild mammals collected from C.18b, a minimum of seven individuals were estimated: two of red deer (one mature and one immature), two of wild boar (one mature and one immature) and one each of aurochs (immature individual), roe deer (mature individual), and polecat (mature individual) (
Table 4). The distribution of red deer remains by skeletal regions indicates the presence of all body parts, indicating that the butchering and consumption of this important game species took place within the settlement (
Figure 8).
Based on 26 remains collected from the C18a, a minimum of six individuals could be estimated: three of red deer (two mature and one immature) and one mature individual for each of aurochs, wild boar, and roe deer (
Table 4). The distribution of remains in skeletal regions is relatively similar as in the case of C.18b (
Figure 8).
4.3. Archaeobotany
The analysed sediments contained numerous phytoliths and showed homogeneous assemblages. In the five analysed samples, we identified 11 morphotypes (
Figure 9): BILOBATE, RONDEL, CRENATE, SADDLE, ELONGATE ENTIRE, ELONGATE DENDRITIC, ACUTE BULBOSUS, BULLIFORM FLABELLATE, SPHEROID PSILATE, TRACHEARY, BLOCKY.
Most of the identified morphotypes are attributed to grasses.
In all samples, the
Rondel phytoliths are dominant, their percentage varying from 50.55% to 65.04%. Generally, this morphotype is associated with the Pooideae subfamily [
64,
65]. Most cereals belong to this subfamily, which includes plants with C3 metabolism, which develop in temperate biotopes.
CRENATE phytoliths indicate the existence in the studied area of taxa from the Pooideae subfamily. Their percentages vary from 2.33% to 4.44%.
The BILOBATE morphotype (between 5.83 and 8.03% of the total) was identified in all analysed samples. This is generally a characteristic of the subfamily Panicoideae [
7,
65], a subfamily that includes often C4 plants adapted to a warmer climate. Yet, subfamilies such as Arundinoideae, Pooideae, and Chloridoideae can also produce the BILOBATE morphotype [
65].
We also identified SADDLE-type phytoliths. The species of the Chloridoideae subfamily may produce this morphotype in large quantities [
7,
64,
66]; SADDLE phytoliths can also be produced by other subfamilies of Poaceae [
52]. The percentages recorded in the samples from Șoimuş are modest (maximum 0.86%).
Numerous plants can produce ELONGATE ENTIRE phytoliths. Hence, a taxonomic assignment cannot be performed [
52]. These phytoliths in Șoimuș samples vary between 8.93% and 13.99%.
The ACUTE BULBOSUS morphotype is present in up to 8.89% of our samples; it is an indicator of the Poaceae family [
52,
67,
68,
69].
ELONGATE DENDRITIC phytoliths (morphotypes from inflorescence bracts) are produced at the inflorescence level in many cereals. [
70]. However, they can also come from the wild grasses’ inflorescences [
71]. In the Șoimuș site, this morphotype was identified in all samples, reaching almost 16% in one of the samples.
The SPHEROID PSILATE morphotype is characteristic of dicot plants [
67,
72,
73,
74] and was identified in all samples, composing up to 4.17% of the total. In general, it is considered that this type of phytolith can be used as a forest indicator [
75].
The TRACHEARY morphotype was preserved modestly (0.26%) in a single sample. According to ICPN [
52], it can be found in all groups of vascular plants.
Additionally, in insignificant percentages were recorded the morphotypes BULLIFORM FLABELLATE (maximum 2.33%) and Blocky (0.29%).
5. Discussion
The discovery of the early Neolithic settlement from Şoimuş-Teleghi in southwestern Transylvania (Romania) during the rescue archaeological excavation from 2011 at the highway project Deva–Orăştie helps us to understand lifestyle sequences of the first farmers at the north of the Danube, in particular from the perspective of archaeobotany and zooarchaeology. The excavation carried out in two different features from the site Teleghi, namely, the pit-hut C.18a, which is overlapped by C.18b, a possible surface house, together with the artefact analysis and the 14C results for zooarchaeological materials, show that the beginning of the early Neolithic period in the western regions of Romania could be placed at the end of 7th millennium cal BC and at the beginning of the next one. Another confirmation for the beginning of the early Neolithic period in southwestern Transylvania, between the 7th and 6th millennium cal BC, is secured by a further archaeological site, approximately 15 km east of Şoimuş-Teleghi, more exactly, Rapoltu Mare-Şeghi. Here, the results of field surveys, test pits, and the latest 14C values confirmed good parallels between these two sites [
26], proven also by archaeobotany and archaeozoology.
Comparing these two archaeozoological samples, it can be found that in general, they are similar, but there is also a difference that could suggest a change in the behaviour of the Starčevo-Criș communities over time. Thus, in both, the remains of domestic mammals predominate, mainly cattle and sheep/goat; pig remains are almost absent; the main hunted species is red deer, and an aquatic animal resource is almost absent. The change in the sample structure over time from the older C.18a to the newer C.18b consisted of an increase in the cattle’s percentage, to the detriment of sheep/goat. However, since the analysed archaeozoological samples are relatively small and the data on the selection by age and sex are weak, we cannot evaluate the cattle use strategy.
The settlement investigated from an archaeozoological point of view must have been located in an open space, which would have provided conditions for raising herds of cattle and sheep/goat. At the same time, the forest was not at a great distance, so large wild mammals, such as red deer, could be brought to the settlement, where they would be butchered.
The archaeozoological assemblage from Rapoltu Mare-Seghi, contemporary with Soimuș-Teleghi, confirm the same characteristics: the dominance of cattle and sheep/goat, the absence of pig, and the low frequency of the hunted species [
26].
Phytolith results seem also to outline the image of a rather open space. A strong dominance of grasses is attested by phytolith assemblages. An ELONGATE DENDRITIC morphotype is abundantly found in species belonging to the tribes Triticeae and Aveneae [
52]. In an archaeological context, these phytoliths are an indicator of grass inflorescences [
71,
76,
77,
78]. Phytoliths of this type were recorded in all the samples analysed for this study, their share (up to almost 16%) suggesting an anthropogenic accumulation, which could signify the presence of cereals, an accumulation of them being quite plausible. This hypothesis can also be supported by the constant presence of CRENATE phytoliths. The accumulation could also be explained by bringing to the site grasses used for bedding or animal feeding [
79]. Quite abundant in the samples from Șoimuș-Teleghi (up to 8%), BILOBATE-type phytoliths could suggest a significant presence of some Panicoideae. This could imply the presence of spontaneous species [
23], but it is not excluded, given the context, that it also suggests the presence of cultivated species (millet—
Panicum sp). Of course, the possibility that representatives of some subfamilies, such as Arundinoideae (e.g.,
Phragmites sp.), Pooideae, and/or Chloridoideae were present in the Șoimuş-Teleghi landscape, must also be taken into account. Woody dicotyledons were part of the landscape, but the share of SPHEROID PSILATE phytoliths shows their modest presence.
The phytolith analysis from Șoimuș-Teleghi, therefore, highlights an open environment, as in the case of the Rapoltu Mare-Şeghi site [
26]. Phytolith assemblages from Rapoltu Mare-Şeghi revealed “an anthropogenic accumulation, which does not necessarily indicate the cultivation of cereals, but rather the consumption of wild grasses by the animals raised by pastoral community” [
26]. The composition of the spectra from Șoimuș-Teleghi highlighted the constant and much more significant presence of BILOBATE and CRENATE phytoliths, and the percentages of ELONGATE DENDRITIC phytoliths are slightly more important (16% compared to 11.53% at the Rapoltu site). These aspects may suggest the fact that the communities from Șoimuș-Teleghi were more sedentary than those from Rapoltu Mare, where the results revealed a relatively short-term settlement of a mobile pastoral group.
6. Conclusions
The 14C results for the Early Neolithic features C.18a and C.18b from Şoimuş-Teleghi show that at the turn of the 7th and 6th millennium cal BC, we can talk about the beginning of the Starčevo-Criş cultural complex in southwestern Transylvania, a period represented by the activity of early farmers.
Although the archaeozoological samples are relatively small, the analysed animal remains give data on the local palaeoeconomy (i.e., animal husbandry mainly practised, little hunting, and sporadic mollusc gathering), and palaeoenvironment (i.e., open spaces around the settlement needed for herding cattle and sheep/goat). The communities of the Starčevo-Criş culture from Mureş Valley probably were non-sedentary pastors, periodically searching for new grasslands, as it is indicated by the predominance of cattle and sheep/goat remains and the negligible presence of pigs.
The study of phytoliths strengthens the hypothesis outlined by the archaeozoological data. The significant presence of Poaceae plants is reflected by phytolith analyses. This brings to light an open space. An anthropogenic accumulation is noticeable, mainly due to the significant presence of ELONGATE DENDRITIC phytoliths. This does not automatically mean the cultivation of cereals by the Şoimuş-Teleghi community but can only mean their presence. We could rather think of the presence of bedding or fodder. It is also possible that the percentage of the ELONGATE DENDRITIC morphotype is due to the feeding of animals raised at Şoimuş-Teleghi with wild grasses.
Bioarchaeological data (i.e., archaeozoology and archaeobotany) are suggesting an Early Neolithic way of life centred on herding. Previous archaeozoological research on sites of the Starčevo-Criş culture, as well as our results, indicate a very low frequency of pigs, which is therefore a feature indicating the mobility of these early Neolithic groups as herders of cattle and sheep/goats.
Author Contributions
Conceptualization, M.S.S., I.A.B., and M.D.; methodology, I.A.B., O.G., D.P., and M.D.; software, O.G. and D.P.; validation, I.A.B. and L.B.; formal analysis, D.I.M., M.S.S., I.A.B., and M.D.; investigation, D.I.M., M.S.S., I.A.B., O.G., D.P., and M.D.; resources, I.A.B., O.G., and D.P.; data curation, L.B.; writing—original draft preparation, I.A.B., O.G., D.P., and M.D.; writing—review and editing, I.A.B. and M.D.; visualization, M.S.S.; supervision, L.B.; project administration, M.S.S.; funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The radiocarbon dating of the faunal remains from Şoimuş-Teleghi was conducted at the National Institute of Research Development for Physics and Nuclear Engineering “Horia Hulubei” (IFIN-HH) Bucharest–Măgurele, Romania, within the Applied Nuclear Physics Department, RoAMS Laboratory, with the help of Corina Anca Simion, Scientific Researcher, and Tiberiu Sava, Scientific Researcher, to whom we are very grateful for all the help provided, both in the work performed in the laboratory and in the interpretation of the C14 results. The radiocarbon dating at the National Institute of Research Development for Physics and Nuclear Engineering “Horia Hulubei” (IFIN-HH) Bucharest-Măgurele, Romania, was funded by HG 786/2014, republished in 2018, on the List of Facilities and Special Objectives of National Interest (IOSIN), whose expenses for the maintenance, operation, and guarding of facilities are financed from the funds allocated by the Government to the Ministry of Research and Innovation from the state budget, chapter “Basic research and research and development. Other radiometric measurements of the osteological material from the Teleghi point were conducted in the Beta Analytic laboratory from Miami, FL, USA, with the help of Wei Chu, a Postdoc Researcher at Leiden University, Netherlands, whom we must also thank for all the support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Bonsall, C. When was the Neolithic transition in the Iron Gates? A Short Walk through the Balkans: The First Farmers of the Carpathian Basin and Adjacent Regions. In Proceedings of the Conference Held at the Institute of Archaeology UCL, London, UK, 20–22 June 2005; Spataro, M., Biagi, P., Eds.; Società per la Preistoria e Protostoria della Regione Friuli-Venezia Giulia, Quaderno 12: Trieste, Italy, 2007; pp. 53–66. [Google Scholar]
- Budja, M. The 8200 calBP ‘climate event’ and the process of neolithisation in south-eastern Europe. Doc. Praehist. 2007, 34, 191–291. [Google Scholar] [CrossRef]
- Weninger, B.; Clare, L.; Gerritsen, F.; Horejs, B.; Krauss, R.; Linstädter, J.; Özbal, R.; Rohling, E.J. Neolithisation of the Aegean and Southeast Europe during the 6600–6000 cal BC period of Rapid Climate Change. Doc. Praehist. 2014, 41, 1–31. [Google Scholar] [CrossRef]
- Botić, K. Neolithisation of Sava-Drava-Danube interfluve at the end of the 6600–6000 BC period of Rapid Climate Change: A new solution to an old problem. Doc. Praehist. 2016, 43, 183–207. [Google Scholar] [CrossRef][Green Version]
- Luca, S.A.; Suciu, C.I.; Dumitrescu-Chioar, F. Cataloque of the Early Neolithic (Starčevo-Criş Culture) Settlements in Western Part of Romania—Transylvania, Banat, Crişana, Maramureş, Oltenia and Western Muntenia. In The First Neolithic Sites in Central/South-East European Transect, Early Neolithic Starčevo-Criş Sites on the Territory of Romania; Luca, S.A., Suciu, C.I., Eds.; BAR: Oxford, UK, 2011; Volume 2, pp. 79–132. [Google Scholar]
- Bărbat, I.A. Complexul Cultural Starčevo-Criş în Bazinul Mureşului Mijlociu. Ph.D. Thesis, Universitatea din Alba Iulia, Alba Iulia, Romania, 2013. [Google Scholar]
- Twiss, P.C.; Suess, E.; Smith, R.M. Morphological classification of grass phytoliths. Proced. Soil Sci. Soc. Am. 1969, 33, 109–115. [Google Scholar] [CrossRef]
- Stromberg, C.; Dunn, R.; Crifò, C.; Harris, E. Phytoliths in Paleoecology: Analytical Considerations, Current Use, and Future Directions: Reconstructing Cenozoic Terrestrial Environments and Ecological Communities. In Methods in Paleoecology: Reconstructing Cenozoic Terrestrial Environments and Ecological Communities, Vertebrate Paleobiology and Paleoanthropology; Croft, D.A., Su, D., Simpson, S.W., Eds.; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Bejenaru, L.; Stanc, S. Arheozoologia Neoliticului din Estul şi Sud-Estul României; Editura Universității “Alexandru Ioan Cuza”: Iași, Romania, 2013. [Google Scholar]
- Boghian, D.; Enea, S.C.; Ignătescu, S.; Bejenaru, L.; Stanc, M.S. Comunitățile Cucuteniene din Zona Târgului Frumos. Cercetări Interdisciplinare în Siturile de la Costești și Giurgești; Editura Universității “Alexandru Ioan Cuza”: Iași, Romania, 2014. [Google Scholar]
- Cavaleriu, R.; Bejenaru, L. Cercetări Arheozoologice Privind Cultura Cucuteni, Faza A; Editura Universității “Alexandru Ioan Cuza”: Iași, Romania, 2009. [Google Scholar]
- Bălăşescu, A.; Radu, V. Oameni şi Animale. Strategii şi Resurse la Comunităţile Preistorice Hamangia şi Boian. In Biblioteca Muzeului Naţional, Seria Cercetări Pluridisciplinare; Editura Cetatea de Scaun: București, Romania, 2004. [Google Scholar]
- Bălăşescu, A.; Radu, V.; Moise, D. Omul și mediul animal între mileniile VII-IV î.e.n. la Dunărea de Jos. In Biblioteca Muzeului Naţional, Seria Cercetări Pluridisciplinare; Editura Cetatea de Scaun: București, Romania, 2005. [Google Scholar]
- Bălăşescu, A. Arheozoologia neo-eneoliticului de pe Valea Teleormanului, Biblioteca Muzeului Național. In Seria Cercetări Puridisciplinare; Editura Mega: Cluj Napoca, Romania, 2014. [Google Scholar]
- Bindea, D. Arheozoologia Transilvaniei în Pre—Și Protoistorie; Editura Teognost: Cluj Napoca, Romania, 2008. [Google Scholar]
- El Susi, G. Vânători, Pescari şi Crescători de Animale în Banatul Mileniilor VI î. Cr.—I d.Cr.; Editra Mirton: Timişoara, Romania, 1996. [Google Scholar]
- Bejenaru, L.; Bodi, G.; Stanc, S.; Danu, M. Middle Holocene Landscape to the East of Carpathians: Bioarchaeological Considerations on the Chalcolithic Site of Hoisești (Iași County, Romania). Carpathian J. Earth Environ. Sci. 2014, 9, 121–128. [Google Scholar]
- Bejenaru, L.; Bodi, G.; Stanc, S.; Danu, M. Middle Holocene subsistence East of the Romanian Carpathians: Bioarchaeological data from the Chalcolithic site of Poduri-Dealul Ghindaru. Holocene 2018, 28, 1653–1663. [Google Scholar] [CrossRef]
- Bejenaru, L.; Stanc, S.; Cavaleriu, R. Preliminary analysis of an archaeozoological assemblage discovered in the Cucuteni B settlement from Sărata-Monteoru (Buzău County). An. Ştiinţifice Ale Univ. Al. I. Cuza Iaşi S. Biol. Anim. 2011, 57, 141–145. [Google Scholar]
- Bodi, G.; Radu Pîrnău, R.; Danu, M.; Cavaleriu, R. Cercetări Interdisciplinare în Neo-Eneoliticul Din Nord-Estul României; Editura Universității “Alexandrul Ioan Cuza”: Iași, Romania, 2013. [Google Scholar]
- Tencariu, F.A.; Delhon, C.; Vornicu, D.M.; Asăndulesei, A.; Brașoveanu, C.; Danu, M. Science Revealing Ancient Magic: Phytolith Evidence from the Early Chalcolithic Site of Isaiia (Eastern Romania). Biology 2022, 11, 1102. [Google Scholar] [CrossRef]
- Danu, M.; Messager, E.; Carozza, J.M.; Carozza, L.; Bouby, L.; Philibert, S.; Anderson, P.; Burens, A.; Micu, C. Phytolith evidence of cereal processing in the Danube Delta during the Chalcolithic period. Quat. Int. 2019, 504, 128–138. [Google Scholar] [CrossRef]
- Danu, M.; Delhon, C.; Weller, O. Could the grasses have played a role in the earliest salt exploitation? Phytoliths analysis of prehistoric salt spring from Hălăbutoaia—Ţolici (Romania). Archaeol. Anthropol. Sci. 2020, 12, 270. [Google Scholar] [CrossRef]
- Bejenaru, L.; Danu, M.; Stanc, S. Overall evaluation of biological remains discovered in the Chalcolithic site (Cucuteni culture, Vth-IVth millennia Cal B.C.) of Costeşti (Iaşi County, Romania). Int. J. Conserv. Sci. 2016, 7, 93–100. [Google Scholar]
- Danu, M.; Diaconu, V.; Bejenaru, L. Chalcolithic agropastoralism traces in the site of Răucești (Neamț county, Romania): Phytoliths and animal remains. Int. J. Conserv. Sci. 2016, 7, 1071–1080. [Google Scholar]
- Stanc, S.; Barbat, I.; Bejenaru, L.; Danu, M. Bioarchaeological evaluation of the early Neolithic site of Rapoltu Mare-Seghi (Hunedoara county, Romania). Int. J. Conserv. Sci. 2020, 11, 209–218. [Google Scholar]
- Stanc, S.; Danu, M.; Paraschiv, D.; Bejenaru, L. Bioarcheological Indicators Related to Human–Environmental Interactions in a Roman–Byzantine Settlement in Southeast Romania: Ibida Fortress. Sage Open 2020, 10, 2158244020969664. [Google Scholar] [CrossRef]
- Bărbat, I.A. Interacţiunea comunităţilor neoliticului timpuriu cu mediul înconjurător. Aşezările Starčevo-Criş de la Șoimuş (jud. Hunedoara). Sargetia S.N. 2015, 6, 9–40. [Google Scholar]
- Ujvári, I. Geografia apelor României; Editura științifică: Bucureşti, Romania, 1972. [Google Scholar]
- Grigore, M. Culoarul Mureșului. In Geografia României, III. Carpaţii Româneşti şi Depresiunea Transilvaniei; Oancea, D., Velcea, V., Eds.; Editura Academiei Republicii Socialiste România: București, Romania, 1987; pp. 426–430. [Google Scholar]
- Oncu, M. Culoarul Mureşului, Sectorul Deva-Zam, Studiu Geoecologic; Editura Focul Viu: Cluj-Napoca, Romania, 2000. [Google Scholar]
- Andrițoiu, I. Contribuţii la repertoriul arheologic al judeţului Hunedoara. Sargetia 1979, 14, 15–34. [Google Scholar]
- Schuster, C.; Petcu, R.; Petcu, R.; Heroiu, A.; Rumega, V.; Creţu, A.P.; Dimache, M.; Irimuș, L.; Dobrotă, S.; Vasilescu, D.; et al. Șoimuș, com. Șoimuș, jud. Hunedoara (Varianta de ocolire Deva-Orăștie), Punct: Șoimuș 1 (Avicola) km. 29+750-30+300, campania 2011; CCA, Editura Institutul Național al Patrimoniului: Bucuresti, Romania, 2012; pp. 291–292. [Google Scholar]
- Drașovean, F.; Rotea, M. Așezarea neolitică de la Șoimuș. Contribuţii la problemele neoliticului târziu din sud-vestul Transilvaniei. Apulum 1986, 23, 9–24. [Google Scholar]
- Popescu, D. Noi contribuţiuni în precizarea vîrstei Stratelor de Deva. Sargetia 1970, 7, 241–245. [Google Scholar]
- Lorinţ, C.R.; Bărbat, I.A. Andesites—A raw material used for manufacturing the millstones in the Early Neolithic from Hunedoara County, Romania. Preliminary results. SGEM 2014, 1, 11–18. [Google Scholar]
- Bărbat, I.A. Materii prime de origine vulcanică prezente în așezările Starčevo-Criș de pe teritoriul României: Andezitele. Caiet. C.I.V.A. 2014, 3, 9–35. [Google Scholar]
- Stoica, C. Staţiuni si Izvoare Minerale din Tinutul Hunedoarei; Consiliul Județean pentru Cultură și Artă: Deva, Romania, 1970. [Google Scholar]
- Orășeanu, I. Hidrogeologia carstului din Munții Apuseni; Editura Belvedere: Oradea, Romania, 2016. [Google Scholar]
- Lazarovici, C.M.; Lazarovici, G. Arhitectura Neoliticului şi Epocii Cuprului din România (Neoliticul, vol. 1.); Editura Trinitas: Iaşi, Romania, 2006. [Google Scholar]
- Bailey, D. Breaking the Surface. An Art/Archaeology of Prehistoric Architecture; Oxford University Press: New York, NY, USA, 2018. [Google Scholar]
- Lorinţ, C.R.; Bărbat, I.A. The petrographic and geochemical analysis results of ground stone tools discovered in the Early Neolithic settlement from Soimuș (Hunedoara County, Romania). SGEM 2015, 1, 477–484. [Google Scholar]
- Beldiman, C.; Bărbat, I.A.; Sztancs, D.M. Bone and antler artefacts dated from Early Neolithic discovered recently in South-Western Transylvania, Romania. Auvtsah 2012, 14, 43–57. [Google Scholar] [CrossRef]
- Beldiman, C.; Sztancs, D.M.; Bărbat, I.A. Bone and antler artefacts dated from Starčevo-Cris culture in Transylvania, Romania: Recent discoveries and microscopic analyses. In Archaeotechnology: Studying Technology from Prehistory to the Middle Ages; Vitezović, S., Antonović, D., Eds.; Srpsko arheološko društvo: Beograd, Serbia, 2014; pp. 43–57. [Google Scholar]
- Sztancs, D.M.; Beldiman, C.; Bărbat, I.A. Bone and Antler Artefacts Dated from Starčevo-Criș Culture in Transylvania: Recent Discoveries and Microscopic Analyses. Arheovest 2014, 1, 773–790. [Google Scholar]
- Schmid, E. Atlas of Animal Bones; Elsevier: Amsterdam, The Netherlands; London, UK; New York, NY, USA, 1972. [Google Scholar]
- Reitz, E.J.; Wing, E.S. Zooarchaeology (Cambridge Manuals in Archaeology), 2nd ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Udrescu, M.; Bejenaru, L.; Hriscu, C. Indroducere în Arheozoologie; Editura Corson: Iași, Romania, 1999. [Google Scholar]
- von den Driesch, A. A guide to the measurement of animal bones from archaeological sites. Peabody Mus. Bull. 1976, 1, 1–137. [Google Scholar]
- Teichert, M. Osteometrische Untersuchungen zur Berechnung der Widerristhohe bei Schafen. In Archaeozoological Studies; Clason, A.T., Ed.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1975; pp. 51–69. [Google Scholar]
- Lentfer, C.J.; Boyd, W.E. A comparison of three methods for the extraction of phytoliths from sediments. J. Archaeol. Sci. 1998, 25, 1159–1183. [Google Scholar] [CrossRef]
- International Committee on Phytolith Taxonomy (ICPT). International code for phytolith nomenclature (ICPN) 2.0. Ann. Bot. 2019, 124, 189–199. [Google Scholar] [CrossRef]
- Hedges, R.E.M.; van Klinken, G.J. A review of current approaches in the pretreatment of bone for radiocarbon dating by AMS. Radiocarbon 1992, 34, 279–291. [Google Scholar] [CrossRef]
- Brock, F.; Bronk Ramsey, C.; Higham, T. Quality assurance of ultrafiltered bone dating. Additional Quality Assurance Checks. Radiocarbon 2007, 49, 187–192. [Google Scholar] [CrossRef]
- Brock, F.; Higham, T.; Bronk Ramsey, C. Pre-screening techniques for identification of samples suitable for radiocarbon dating of poorly preserved bones. J. Archaeol. Sci. 2010, 37, 855–865. [Google Scholar] [CrossRef]
- Luca, S.A. Viața trăită sub zei. Situl Starčevo-Criș I de la Cristian I, județul Sibiu, România. In Studies into South-East European Prehistory, II; Karl A. Romstorfer: Suceava, Romania, 2015. [Google Scholar]
- Luca, S.A. Data about the vessels decorated with painting in the discoveries of the Early Neolithic from Cristian I, Sibiu County, Romania...and some “Mesolithic legacies and traditions” on the Danube line and north of it. Ann. D’université Valahia Targoviste. Sect. D’archéologie Et D’histoire 2020, 22, 19–46. [Google Scholar] [CrossRef]
- Biagi, P.; Spataro, M. Noi datări cu radiocarbon în așezările culturii Criș din Banat și Transilvania (România). Patrim. Banat. 2004, 3, 7–20. [Google Scholar]
- Biagi, P.; Shennan, S.; Spataro, M. Rapid Rivers and Slowseas? New data for the Radiocarbon Chronology of the Balkan Peninsula In Prehistoric Archaeology & Anthropological Theory and Education, Reports of Prehistoric Research Projects, 6–7, Salt Lake City—Karlovo; Nikolova, L., Fritz, J., Higgins, J., Eds.; International Institute of Anthropology: Salt Lake City, UT, USA, 2005; pp. 41–50. [Google Scholar]
- Luca, S.A.; Diaconescu, D.; Suciu, C.I. Archaeological Research in Miercurea Sibiului-Petriş (Sibiu County, Romania): The Starčevo-Criş Level during 1997–2005 (a preliminary report). Doc. Praehist. 2008, 25, 325–343. [Google Scholar] [CrossRef]
- Luca, S.A.; Diaconescu, D.; Suciu, C.I. Cercetările arheologice de la Miercurea Sibiului-Petriş (judeţul Sibiu, România). Nivelul Starčevo-Criş în campaniile de cercetare 1997-2005 (raport preliminar). Brukenthal Acta Musei 2008, 3, 7–46. [Google Scholar]
- Audoin-Rouzeau, F. La taille du mouton en Europe de l`antiquite aux tepms modernes. Fiches D`Osteol. Pour L`Archeol. Ser. B Mammif. 1991, 3, 3–36. [Google Scholar]
- Cotta, V.; Bodea, M.; Micu, I. Vanatul si Vanatoarea in Romania; Editura Ceres: Bucuresti, Romania, 2008. [Google Scholar]
- Mulholland, S.C. Phytoliths shape frequencies in North Dakota grasses: A comparison to general patterns. J. Archaeol. Sci. 1989, 16, 489–511. [Google Scholar] [CrossRef]
- Piperno, D.R.; Pearsall, D.M. The Silica Bodies of Tropical American Grasses: Morphology, Taxonomy, and Implications for Grass Systematics and Fossil Phytolith Identification. Smithson. Contrib. Bot. 1998, 85, 1–22. [Google Scholar] [CrossRef]
- Fredlund, G.; Tieszen, L.T. Modern phytolith assemblages from the North American Great Plains. J. Biogeogr. 1994, 21, 321–335. [Google Scholar] [CrossRef]
- Alexandre, A.; Meunier, J.D.; Lézine, A.M.; Vincens, A.; Schwartz, D. Phytoliths: Indicators of grassland dynamics during the late Holocene in intertropical Africa. Paleogeography Paleoclimatology Paleoecol. 1997, 136, 213–229. [Google Scholar] [CrossRef]
- Barboni, D.; Bremond, L.; Bonnefille, R. Comparative study of modern phytolith assemblages from inter-tropical Africa. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 246, 454–470. [Google Scholar] [CrossRef]
- Zucol, A.F.; Brea, M.; Bellosi, E.S. Phytolith analysis in Gran Barranca (central Patagonia): The middle-late Eocene. In The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia; Madden, R., Carlini, A., Vucetich, M., Kay, R., Eds.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Ball, T.; Vrydaghs, L.; Mercer, T.; Pearce, M.; Snyder, S.; Lisztes-Szabó, Z.; Petö, Á. A morphometric study of variance in articulated dendritic phytolith wave lobes within selected species of Triticeae and Aveneae. Veg. Hist. Archaeobotany 2017, 26, 85–97. [Google Scholar] [CrossRef]
- Novello, A.; Barboni, D. Grass inflorescence phytoliths of useful species and wild cereals from sub-Saharan Africa. J. Archaeol. Sci. 2015, 59, 10–22. [Google Scholar] [CrossRef]
- Albert, R.M.; Lavi, O.; Estroff, L.; Weiner, S.; Tsatskin, A.; Ronen, A.; Lev-Yadun, S. Mode of occupation of Tabun Cave, Mt Carmel, Israel during the Mousterian period: A study of the sediments and phytoliths. J. Archaeol. Sci. 1999, 26, 1249–1260. [Google Scholar] [CrossRef]
- Bozarth, S. Classification of opal phytoliths formed in selected Dicotyledons native to the Great Plains. In Phytoliths Systematics, Emerging Issue; Rapp, G., Mulholland, S., Eds.; Plenum Press: New York, NY, USA, 1992; pp. 193–214. [Google Scholar]
- Delhon, C.; Alexandre, A.; Berger, J.F.; Thiébault, S.; Brochier, J.L.; Meunier, J.D. Phytolith assemblages as a promising tool for reconstructing Mediterranean Holocene vegetation. Quat. Res. 2003, 59, 48–60. [Google Scholar] [CrossRef]
- Aleman, J.C.; Canal-Subitani, S.; Favier, C.; Bremond, L. Influence of the local environment on lacustrine sedimentary phytolith records. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 414, 273–283. [Google Scholar] [CrossRef]
- Albert, R.M.; Shahack-Gross, R.; Cabanes, D.; Gilboa, A.; Lev-Yaduin, S.; Portillo, M.; Sharon, I.; Boaretto, E.; Weiner, S. Phytolith-rich layers from the Late Bronze and Iron Ages at Tel Dor (Israel): Mode of formation and archaeological significance. J. Archaeol. Sci. 2008, 35, 57–75. [Google Scholar] [CrossRef]
- Cabanes, D.; Mallol, C.; Expósito, I.; Baena, J. Phytolith evidence for hearths and beds in the late Mousterian occupations of Esquilleu cave (Cantabria, Spain). J. Archaeol. Sci. 2010, 37, 2947–2957. [Google Scholar] [CrossRef]
- Madella, M. Understanding archaeological structures by means of phytolith analysis: A test from the Iron Age site Kilise Tepe–Turkey. In Phytoliths—Applications in Earth Science and Human History; Meunier, J.D., Colin, F., Eds.; CRC Press: Boca Raton, USA, 2011; pp. 173–182. [Google Scholar]
- Delhon, C.; Binder, D.; Verdin, P.; Mazuy, A. Phytoliths as a seasonality indicator? The example of the Neolithic site of Pendimoun, south-eastern France. Veget Hist Archaeobot 2020, 29, 229–240. [Google Scholar] [CrossRef]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}