
Transition from Fire-Dependent Open Forests: Alternative Ecosystem States in the Southeastern United States


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
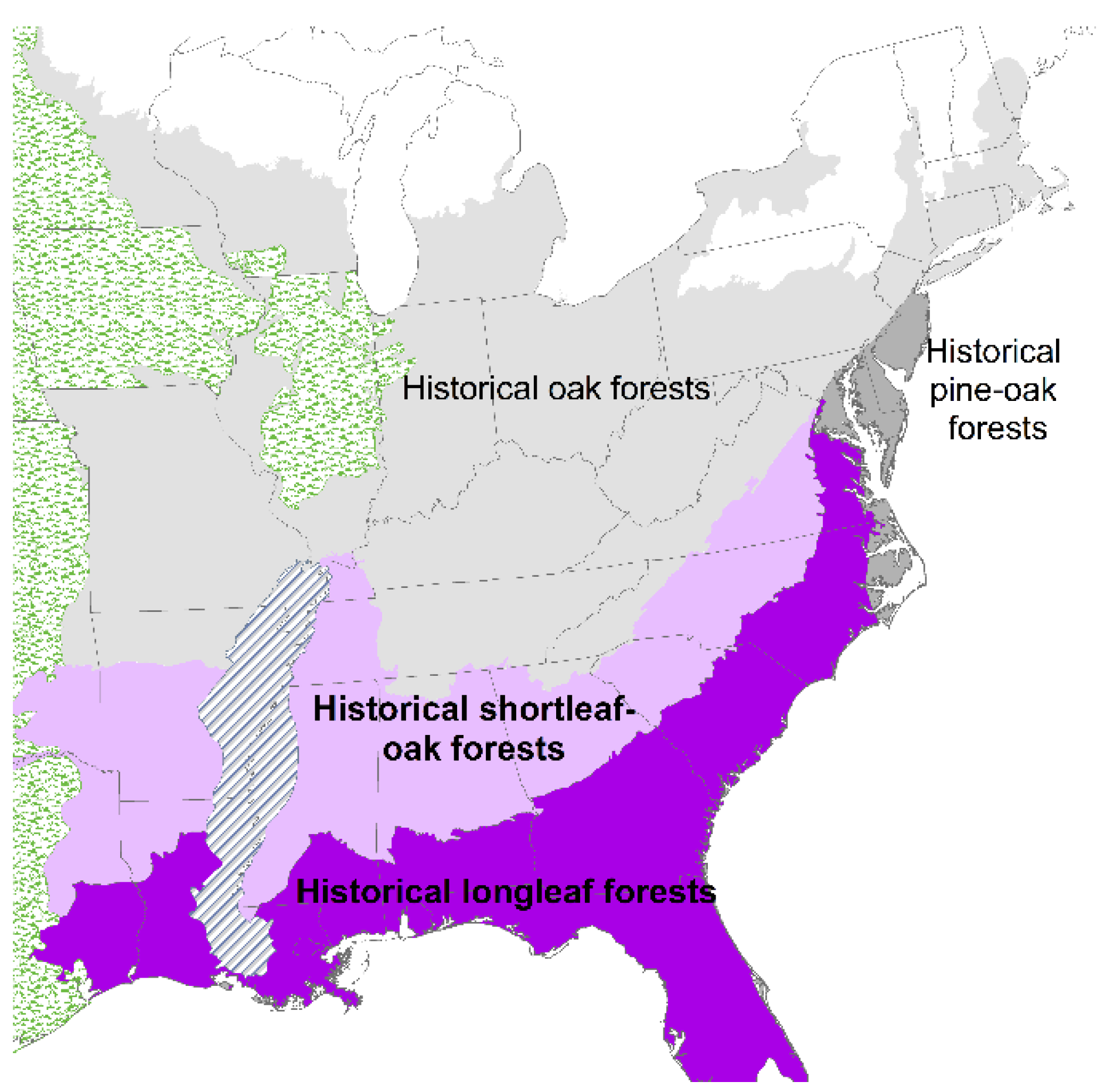
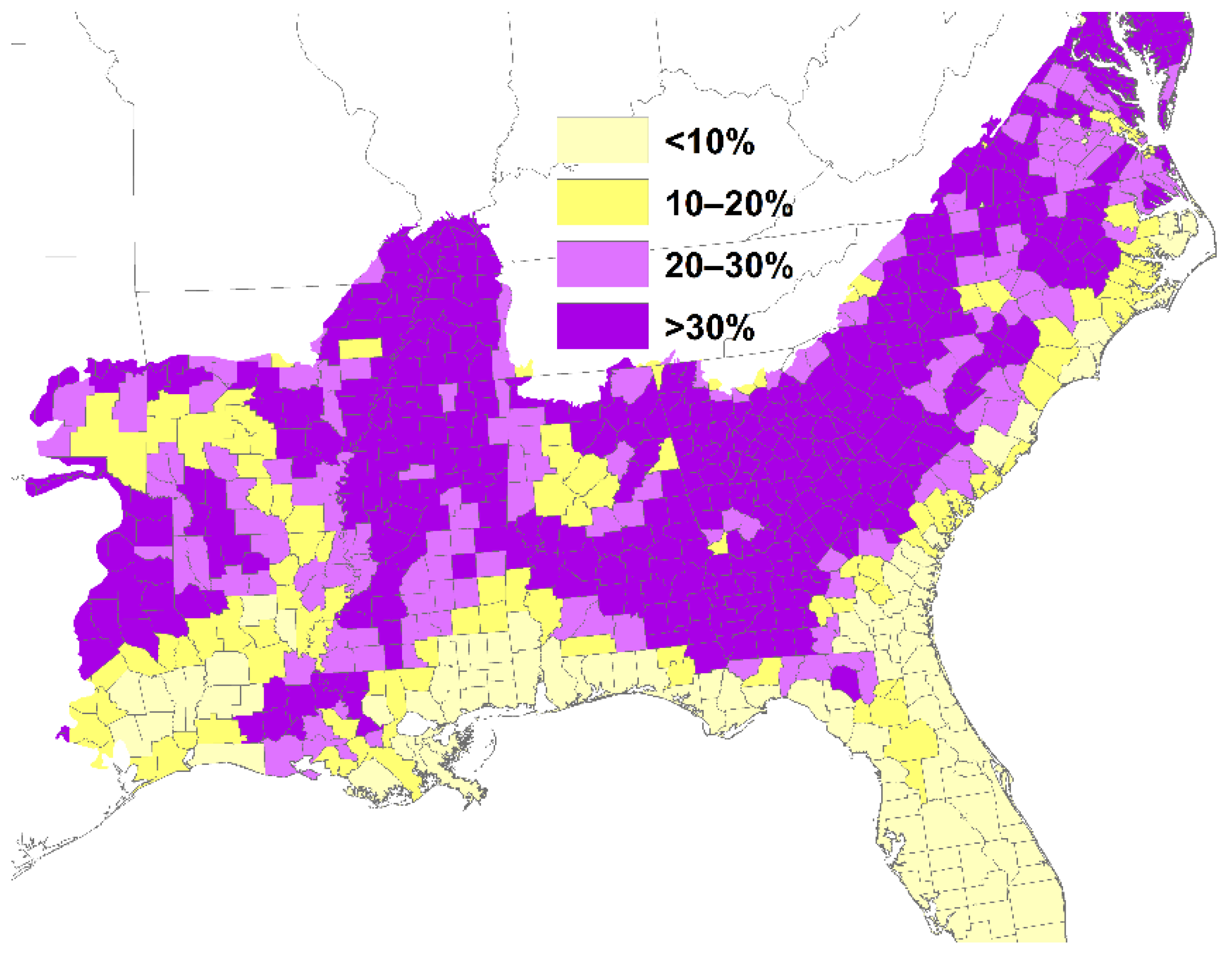
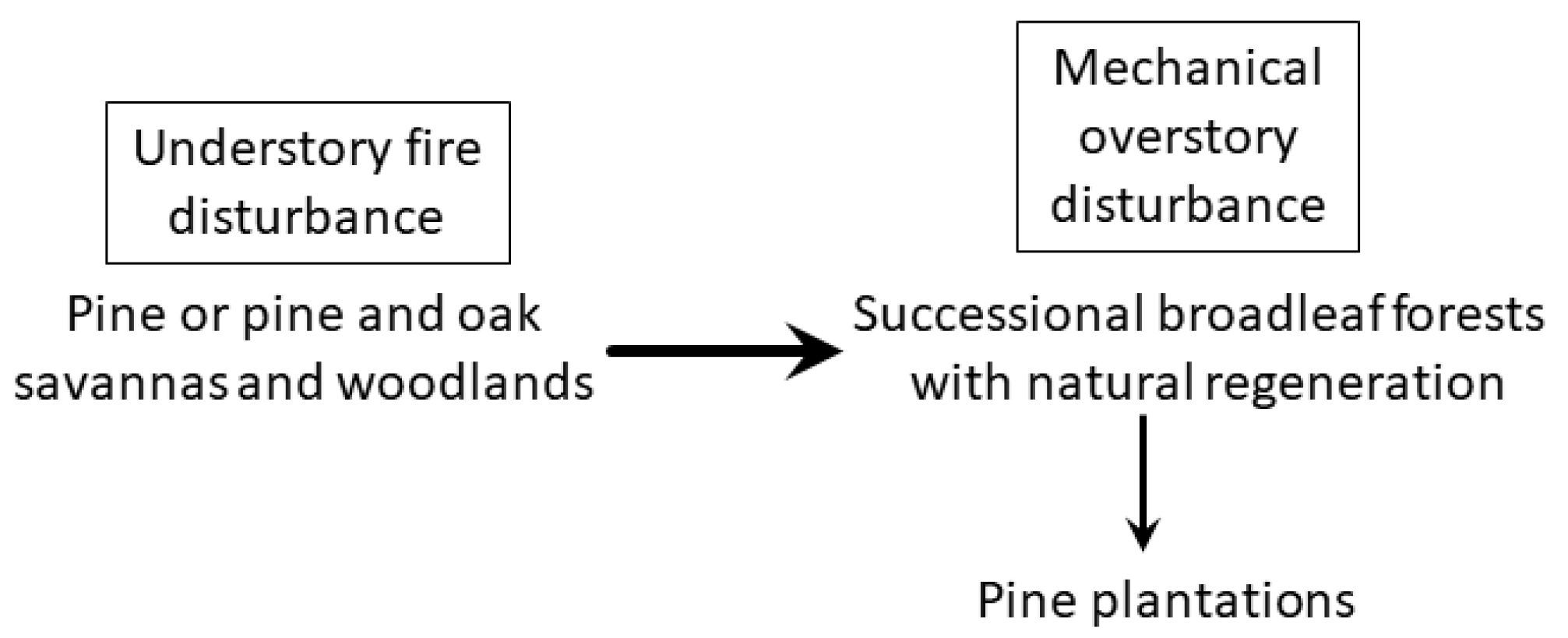
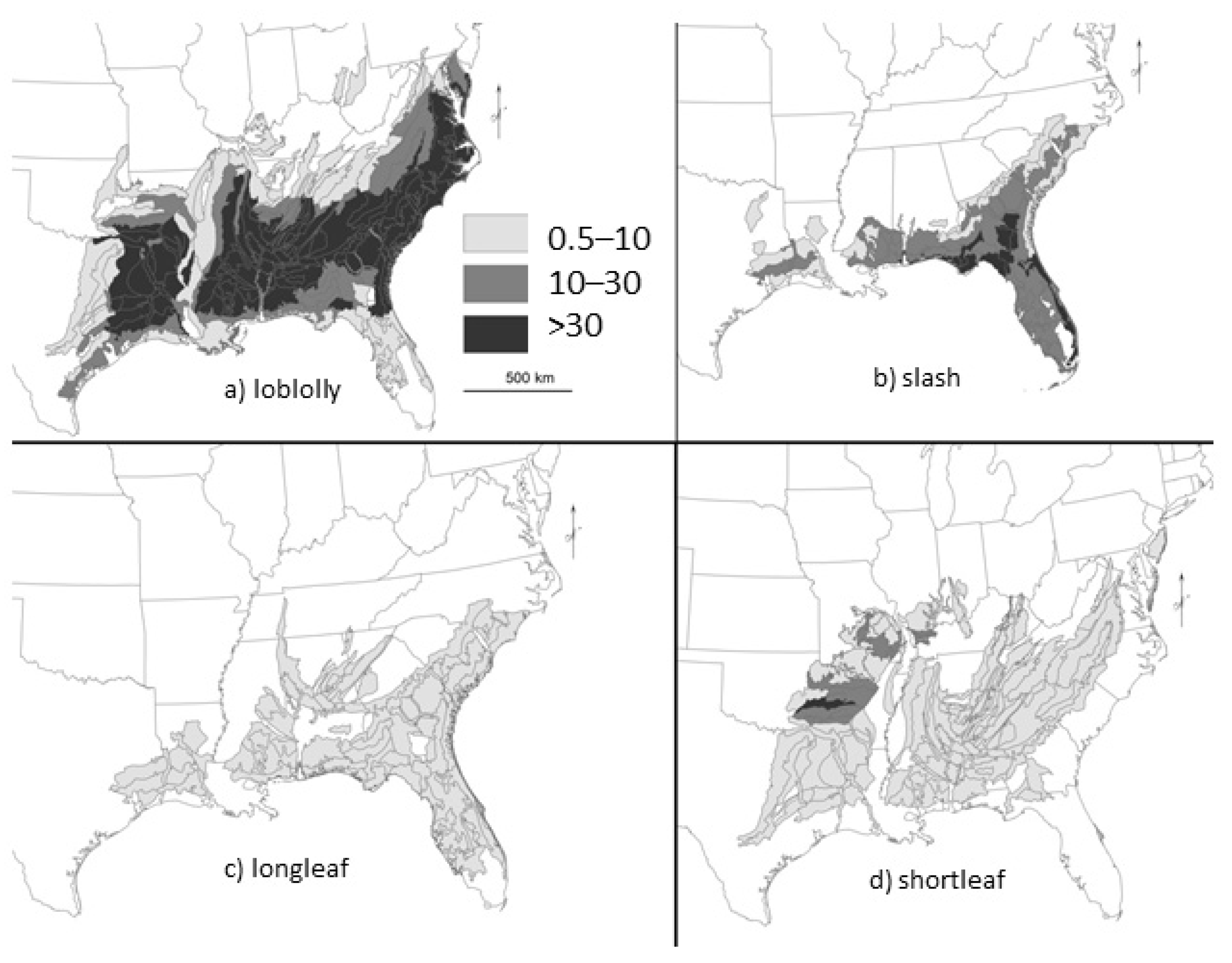
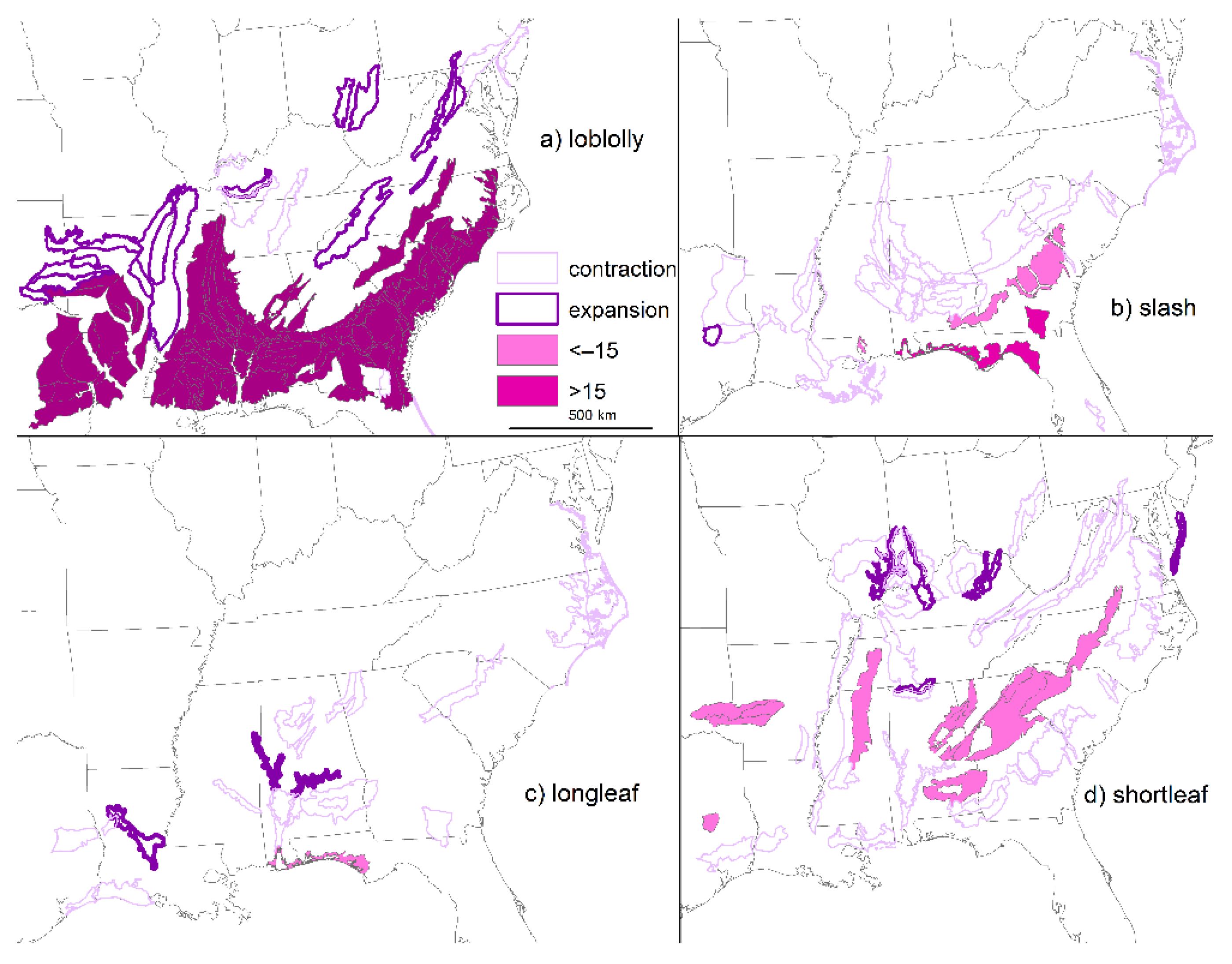
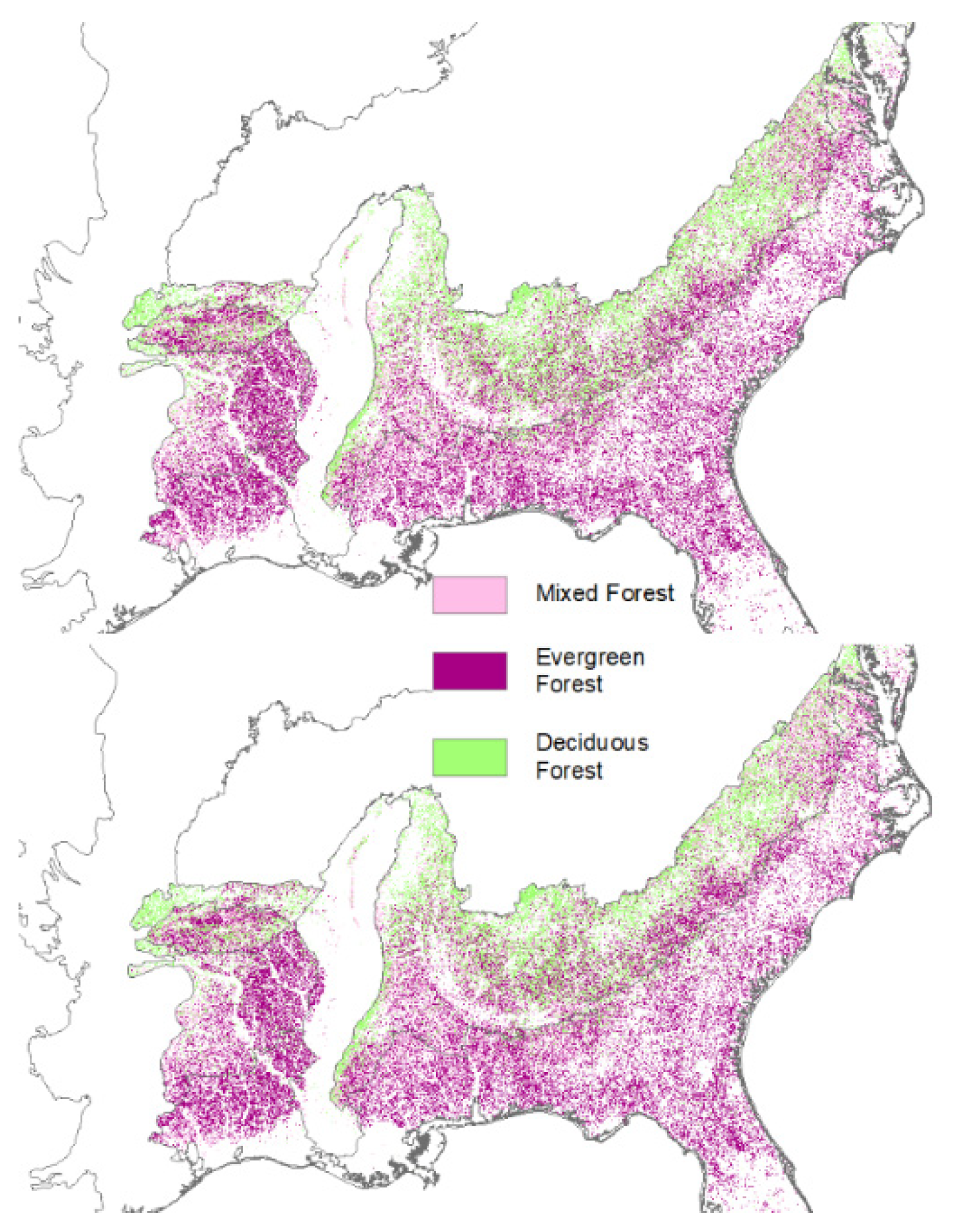
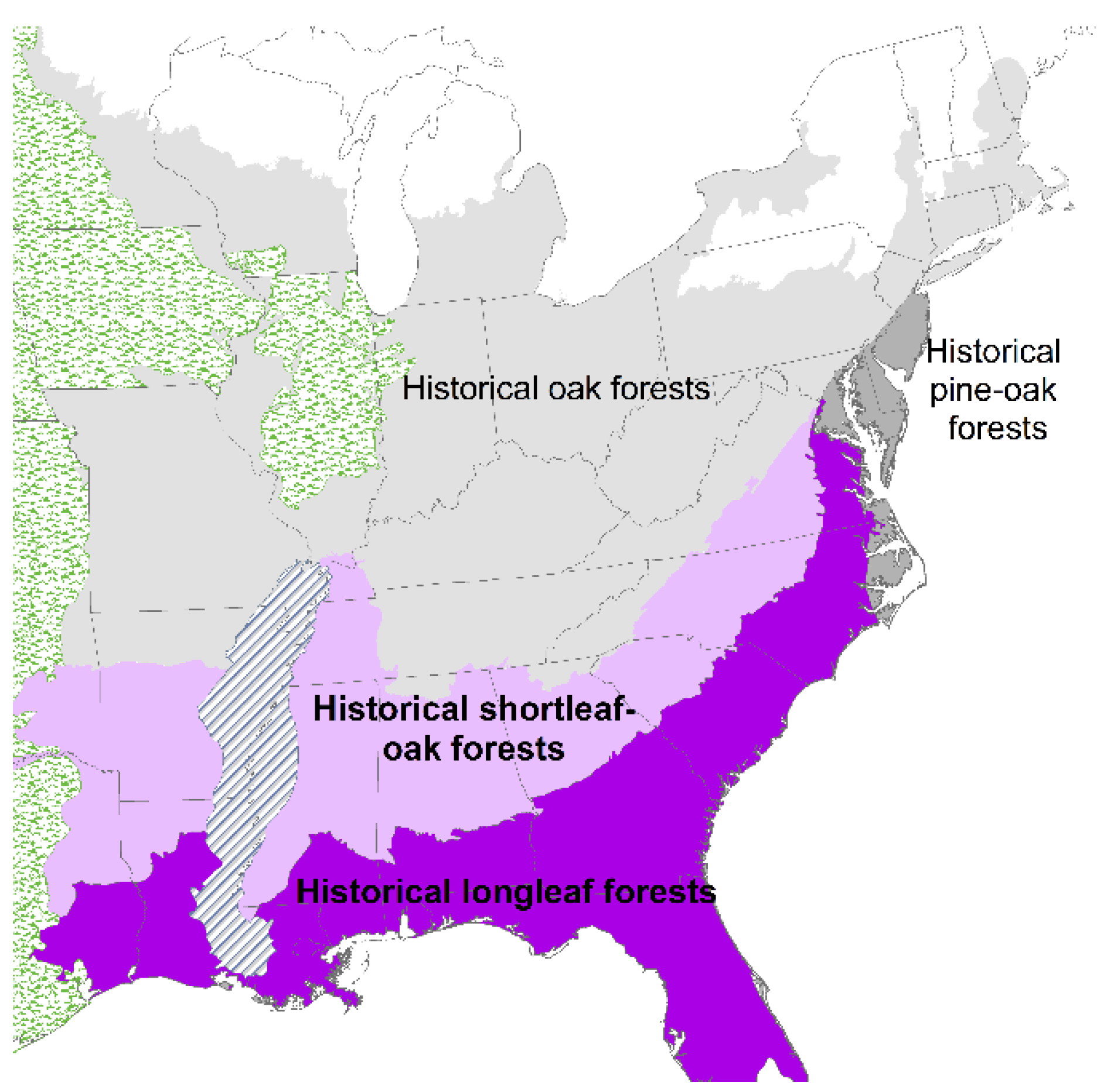
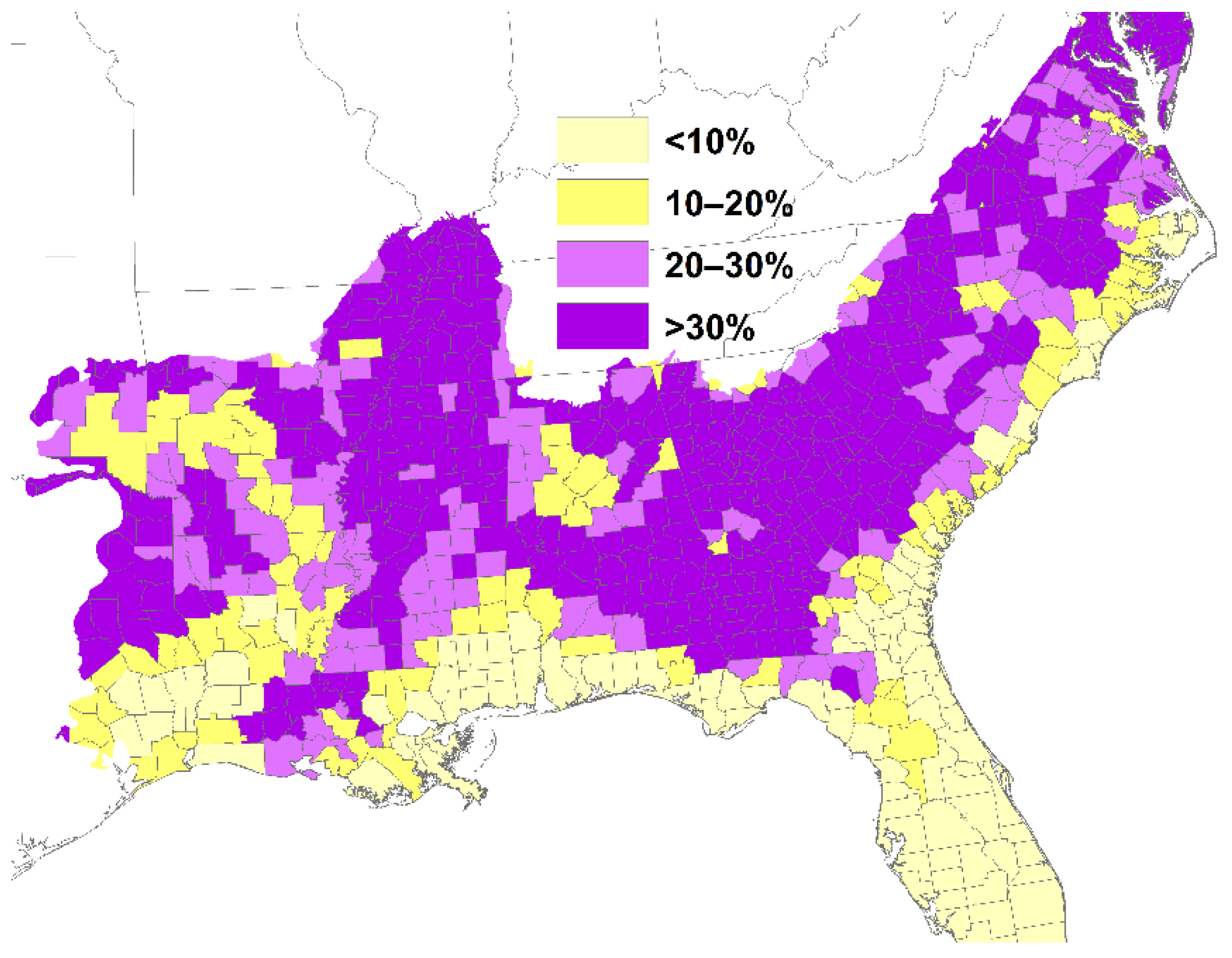
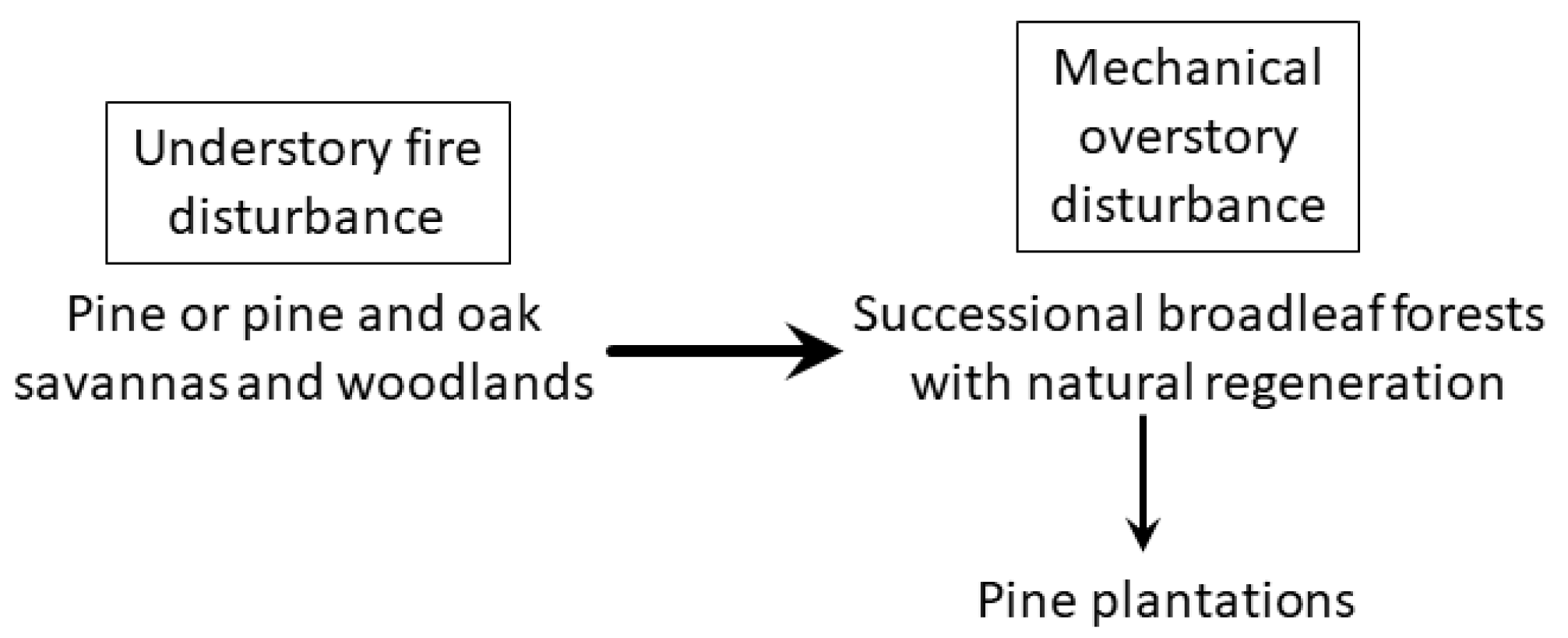
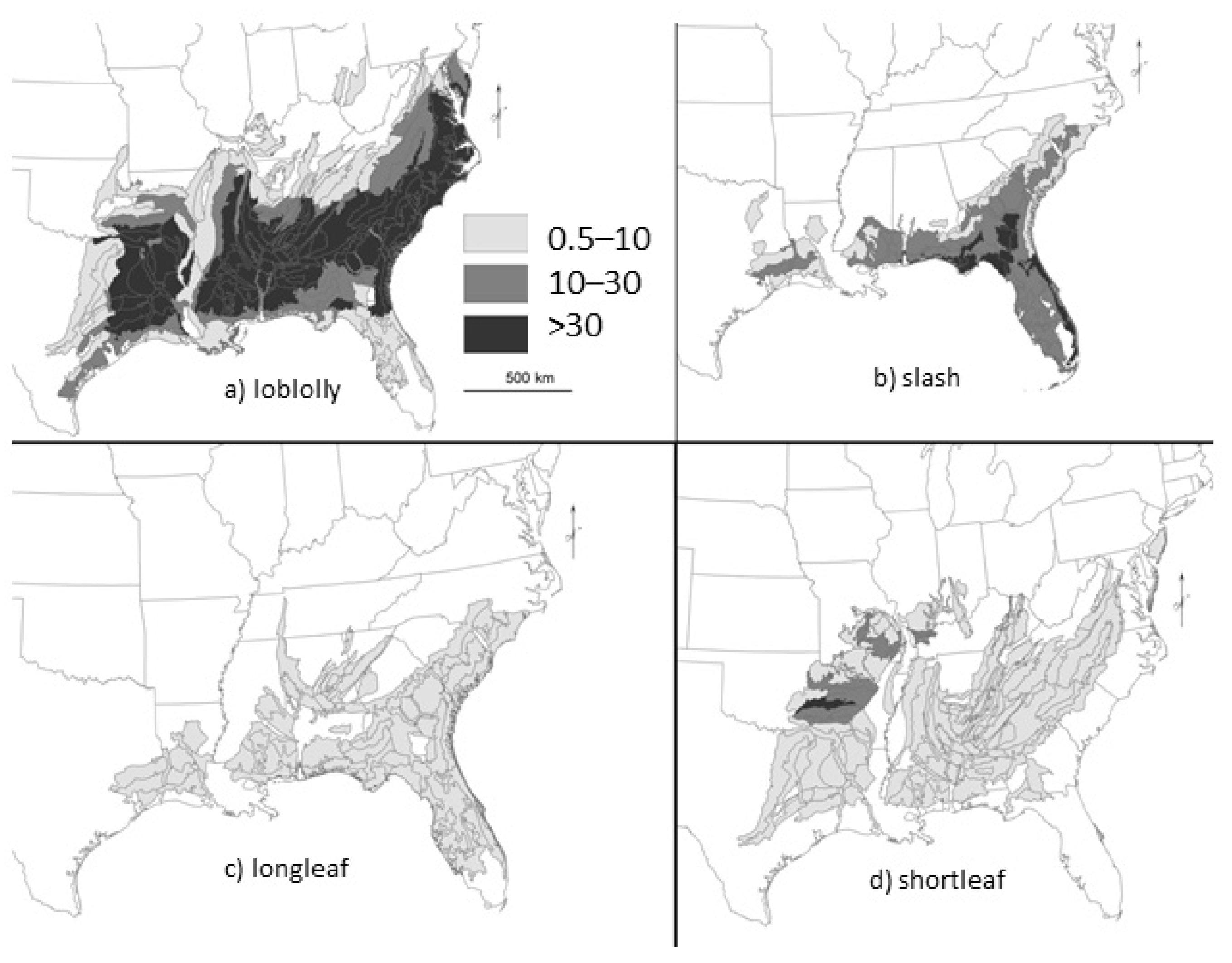
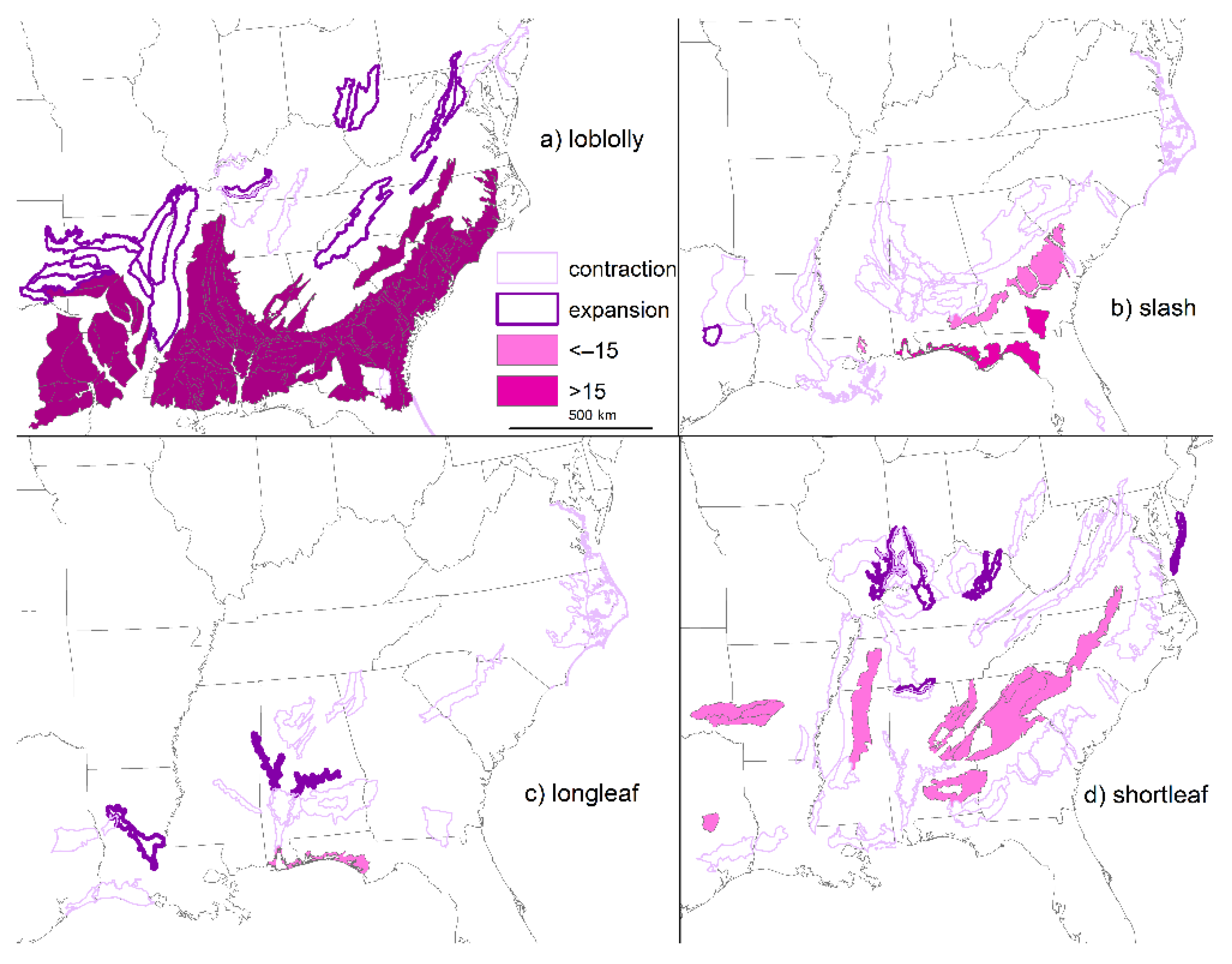
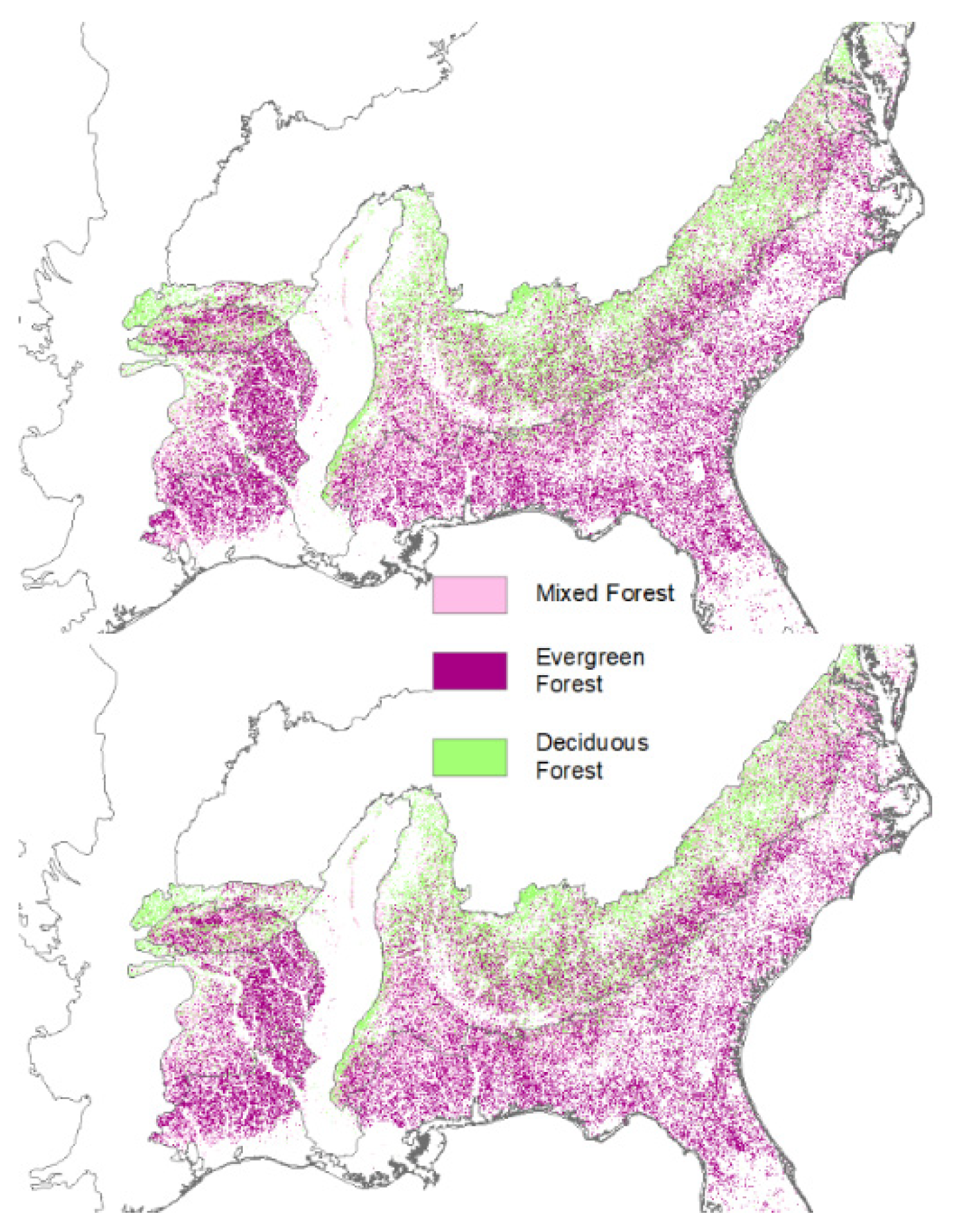
2. Loss of Fire-Dependent Pine and Pine-Oak Forests and Replacement by Broadleaf Forests and Pine Plantations
3. Implications for Management and Conservation
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Lear, D.H.; Carroll, W.; Kapeluck, P.; Johnson, R. History and restoration of the longleaf pine-grassland ecosystem: Implications for species at risk. For. Ecol. Manag. 2005, 211, 150–165. [Google Scholar] [CrossRef]
- Richardson, D.M.; Rundel, P.W.; Jackson, S.T.; Teskey, R.O.; Aronson, J.; Bytnerowicz, A.; Wingfield, M.J.; Procheş, Ş. Human Impacts in Pine Forests: Past, Present, and Future. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 275–297. [Google Scholar] [CrossRef] [Green Version]
- Procheş, Ş.; Wilson, J.R.U.; Richardson, D.M.; Rejmánek, M. Native and naturalized range size in Pinus: Relative importance of biogeography, introduction effort and species traits. Glob. Ecol. Biogeogr. 2012, 21, 513–523. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.J.; Jack, S.B.; Engstrom, R.T. Silviculture that sustains: The nexus between silviculture, frequent prescribed fire, and conservation of biodiversity in longleaf pine forests of the southeastern United States. Can. J. For. Res. 2006, 36, 2724–2736. [Google Scholar] [CrossRef] [Green Version]
- Bragg, D.C.; Hanberry, B.B.; Hutchinson, T.F.; Jack, S.B.; Kabrick, J.M. Silvicultural options for open forest management in eastern North America. For. Ecol. Manag. 2020, 474, 118383. [Google Scholar] [CrossRef]
- Noss, R.F. Forgotten Grasslands of the South: Washington, D.C.; Island Press: Washington, DC, USA, 2013; 320p. [Google Scholar]
- Stoddard, H.L. Use of fire in pine forests and game lands of the deep Southeast. In Proceedings of the 1st Tall Timbers Fire Ecology Conference; Tall Timbers: Tallahassee, FL, USA, 1962; pp. 31–42. [Google Scholar]
- Hanberry, B.B.; Bragg, D.C.; Alexander, H.D. Open forest ecosystems: An excluded state. For. Ecol. Manag. 2020, 472, 118256. [Google Scholar] [CrossRef]
- Peet, R.K. Ecological classification of longleaf pine woodlands. In Longleaf Pine Ecosystems: Ecology, Silviculture, and Restoration; Jose, S., Jokela, E., Miller, D., Eds.; Springer: New York, NY, USA, 2006; pp. 51–93. [Google Scholar]
- Semenova-Nelsen, T.A.; Platt, W.J.; Patterson, T.R.; Huffman, J.; Sikes, B.A. Frequent fire reorganizes fungal communities and slows decomposition across a heterogeneous pine savanna landscape. New Phytol. 2019, 224, 916–927. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Platt, W.J. Effects of long-term fire exclusion on tree species composition and stand structure in an old-growth Pinus palustris (longleaf pine) forest. Plant Ecol. 1999, 140, 15–26. [Google Scholar] [CrossRef]
- Brown, H. Wildland burning by American Indians in Virginia. Fire Manag. Today 2000, 60, 29. [Google Scholar]
- Fowler, C.; Konopik, E. The history of fire in the southern United States. Human Ecol. Rev. 2007, 14, 165–176. [Google Scholar]
- Coughlan, M.R. Errakina: Pastoral Fire Use and Landscape Memory in the Basque Region of the French Western Pyrenees. J. Ethnobiol. 2013, 33, 86–104. [Google Scholar] [CrossRef]
- Coughlan, M.R.; Nelson, D.R. Influences of Native American land use on the Colonial Euro-American settlement of the South Carolina Piedmont. PLoS ONE 2018, 13, e0195036. [Google Scholar] [CrossRef] [Green Version]
- Hanberry, B.B.; Abrams, M.D.; Arthur, M.A.; Varner, J.M. Reviewing fire, climate, deer, and foundation species as drivers of historically open oak and pine forests and transition to closed forests. Front. For. Glob. Chang. 2020, 3, 56. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, J.M.; Birdsey, R.; McCullough, K.; He, L.; Deng, F. Age structure and disturbance legacy of North American forests. Biogeosciences 2011, 8, 715–732. [Google Scholar] [CrossRef] [Green Version]
- Lorimer, C.G. Historical and ecological roles of disturbance in eastern North American forests: 9000 Years of change. Wildl. Soc. Bull. 2001, 29, 425–439. [Google Scholar]
- Elsner, J.B.; Jagger, T.H.; Liu, K.-B. Comparison of Hurricane Return Levels Using Historical and Geological Records. J. Appl. Meteorol. Clim. 2008, 47, 368–374. [Google Scholar] [CrossRef]
- Burkhart, H.E.; Haney, H.L., Jr.; Newberry, J.D.; Leuschner, W.A.; Morris, C.L.; Reed, D.D. Evaluation of thinning for reduction of losses from southern pine beetle attack in loblolly pine stands. South. J. Appl. For. 1986, 10, 105–108. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Duncan, S.L. Range of Variability in Southern Coastal Plain Forests: Its Historical, Contemporary, and Future Role in Sustaining Biodiversity. Ecol. Soc. 2009, 14, 17. [Google Scholar] [CrossRef] [Green Version]
- Dickens, E.D.; Moorhead, D.J. A Guide to Thinning Pine Plantations; University of Georgia: Athens, GA, USA, 2015; Available online: https://bugwoodcloud.org/bugwood/productivity/pdfs/Guide_thinning_pine_plantations.pdf (accessed on 25 July 2021).
- Hanberry, B.B.; Thompson, F.R., III. Open forest management for early successional birds. Wildl. Soc. Bull. 2019, 43, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Hanberry, B. Defining the Historical Northeastern forested boundary of the Great Plains Grasslands in the United States. Prof. Geogr. 2019, 72, 1–8. [Google Scholar] [CrossRef]
- Oswalt, C.M.; Cooper, J.A.; Brockway, D.G.; Brooks, H.W.; Walker, J.L.; Connor, K.F.; Oswalt, S.N.; Conner, R.C. History and Current Condition of Longleaf Pine in the Southern United States; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2012.
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of Fire Regime and Habitat on Tree Dynamics in North Florida Longleaf Pine Savannas. Ecol. Monogr. 1995, 65, 441–476. [Google Scholar] [CrossRef]
- Stambaugh, M.C.; Guyette, R.P.; Marschall, J.M. Longleaf pine (Pinus palustris Mill.) fire scars reveal new details of a frequent fire regime. J. Veg. Sci. 2011, 22, 1094–1104. [Google Scholar] [CrossRef]
- Delcourt, H.R. Presettlement vegetation of the North of Red River Land District, Louisiana. Castanea 1976, 41, 122–139. [Google Scholar]
- Frost, C.C. Four centuries of changing landscape patterns in the longleaf pine ecosystem. In The Longleaf Pine Ecosystem: Ecology, Restoration, and Management, Proceedings of the Tall Timbers Fire Ecology Conference, No. 18, Tallahassee, FL, USA, 3–6 November 1993; Hermann, S.M., Ed.; Tall Timbers Research: Tallahassee, FL, USA, 1993; pp. 17–43. [Google Scholar]
- Predmore, S.A. Presettlement Forests and Fire in Escambia and Covington Counties, Alabama. Master’s Thesis, University of Alabama, Auburn, AL, USA, 2007. [Google Scholar]
- Bragg, D.C.; Bragg, H.A. Historical and contemporary environmental context for the Saline-Fifteen site (3BR119). Ark. Archeol. 2016, 55, 1–30. [Google Scholar]
- Hanberry, B.B.; Brzuszek, R.F.; Foster, H.T.; Schauwecker, T.J. Recalling open old growth forests in the Southeastern Mixed Forest province of the United States. Écoscience 2019, 26, 11–22. [Google Scholar] [CrossRef]
- Dayton, P.K. Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo Sound, Antarctica. In Conservation Problems in Antarctica, Proceedings of the Colloquium on Conservation Problems in Antarctica, Blacksburg, VA, USA, 10–12 September 1971; Parker, B.C., Ed.; Allen Press: Lawrence, KS, USA, 1972; pp. 81–96. [Google Scholar]
- Mohr, C.T.; Roth, F. The Timber Pines of the Southern United States; US Government Printing Office: Washington, DC, USA, 1897.
- Mattoon, W.R. Life History of Shortleaf Pine; US Department of Agriculture: Washington, DC, USA, 1915.
- Burns, R.M.; Honkala, B.H. Silvics of North America: 1. Conifers; Agriculture Handbook 654; USDA Forest Service: Washington, DC, USA, 1990.
- Conner, R.C.; Hartsell, A.J. Forest area and conditions. In Southern Forest Resource Assessment; Wear, D.N., Greis, J.G., Eds.; GTR SRS-53; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 357–402. [Google Scholar]
- Maizel, M.; White, R.D.; Root, R.; Gage, S.; Stitt, S.; Osborne, L.; Muehlbach, G. Historical Interrelationships between Population Settlement and Farmland in the Conterminous United States, 1790 to 1992. 1998. Available online: https://archive.usgs.gov/archive/sites/landcover.usgs.gov/luhna/chap2.html (accessed on 21 May 2020).
- Oswalt, C.M. Broad Scale Inventory of Shortleaf Pine. 2013. Available online: http://shortleafpine.net/why-shortleaf/fia-data (accessed on 17 January 2021).
- Frost, C.C. History and future of the longleaf pine ecosystem. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2006; pp. 9–48. [Google Scholar]
- Haynes, R.W. Forest management in the 21st century: Changing numbers, changing context. J. For. 2002, 100, 38–43. [Google Scholar]
- Martin, W.H.; Boyce, S.G. Introduction: The southeastern setting. In Biodiversity of the Southeastern United States: Lowland Terrestrial Communities; Martin, W.H., Boyce, S.G., Echternacht, A.C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 1993; pp. 1–46. [Google Scholar]
- Prestemon, J.P.; Abt, R.C. Timber products supply and demand. In Southern Forest Resource Assessment; Wear, D.N., Greis, J.G., Eds.; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 299–324. [Google Scholar]
- Oswalt, S.N.; Smith, W.B.; Miles, P.D.; Pugh, S.A. Forest Resources of the United States, 2017: A Technical Document Supporting the Forest Service 2020 RPA Assessment; U.S. Department of Agriculture, Forest Service, Washington Office: Washington, DC, USA, 2019.
- Landers, B.J.L.; Lear, D.H.V.; Boyer, W.D. The longleaf pine forests of the Southeast: Requiem or renaissance. J. For. 1995, 93, 39–44. [Google Scholar]
- South, D.B.; Johnson, E.E.; Hainds, M.J.; VanderSchaaf, C.L. Restoring longleaf pine on an agricultural site by planting alternating rows of slash pine: A case study. South. J. Appl. For. 2012, 36, 118–125. [Google Scholar] [CrossRef]
- Forest Inventory and Analysis. 2021. Available online: https://apps.fs.usda.gov/fia/datamart/CSV/datamart_csv.html (accessed on 30 January 2021).
- Bechtold, W.A.; Patterson, P.L. The Enhanced Forest Inventory and Analysis Program—National Sampling Design and Estimation Procedures; USDA Southern Research Station: Asheville, NC, USA, 2005.
- Hanberry, B.; Hansen, M.H. Latitudinal range shifts of tree species in the United States across multi-decadal time scales. Basic Appl. Ecol. 2015, 16, 231–238. [Google Scholar] [CrossRef]
- Ecomap. National Hierarchical Framework of Ecological Units; USDA Forest Service: Washington, DC, USA, 1993.
- Hanberry, B.B.; Hanberry, P.; Riffell, S.K.; DeMarais, S.; Jones, J.C. Bird assemblages of intensively established pine plantations in Coastal Plain Mississippi. J. Wildl. Manag. 2012, 76, 1205–1214. [Google Scholar] [CrossRef]
- Homer, C.; Dewitz, J.; Jin, S.; Xian, G.; Costello, C.; Danielson, P.; Gass, L.; Funk, M.; Wickham, J.; Stehman, S.; et al. Conterminous United States land cover change patterns 2001–2016 from the 2016 National Land Cover Database. ISPRS J. Photogramm. Remote Sens. 2020, 162, 184–199. [Google Scholar] [CrossRef]
- Hanberry, B.B. Recent shifts in shade tolerance and disturbance traits in forests of the eastern United States. Ecol. Process. 2019, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Zachrisson, O. Influence of forest fires on the north Swedish boreal forest. Oikos 1977, 29, 22–32. [Google Scholar] [CrossRef]
- Seischab, F.K.; Bernard, J.M. Pitch pine (Pinus rigida Mill.) communities in central and western New York. Bull. Torrey Bot. Club 1991, 118, 412–423. [Google Scholar] [CrossRef]
- Minnich, R.A.; Barbour, M.G.; Burk, J.H.; Fernau, R.F. Sixty Years of Change in Californian Conifer Forests of the San Bernardino Mountains. Conserv. Biol. 1995, 9, 902–914. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Covington, W.W.; Moore, M.M. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecol. Appl. 1997, 7, 895–908. [Google Scholar] [CrossRef]
- Taylor, A.H. Fire regimes and forest changes in mid and upper montane forests of the southern Cascades, Lassen Volcanic National Park, California, U.S.A. J. Biogeogr. 2000, 27, 87–104. [Google Scholar] [CrossRef]
- Welch, N.; Waldrop, T.; Buckner, E. Response of southern Appalachian table mountain pine (Pinus pungens) and pitch pine (P. rigida) stands to prescribed burning. For. Ecol. Manag. 2000, 136, 185–197. [Google Scholar] [CrossRef]
- Pinto, F.; Romaniuk, S.; Ferguson, M. Changes to preindustrial forest tree composition in central and northeastern Ontario, Canada. Can. J. For. Res. 2008, 38, 1842–1854. [Google Scholar] [CrossRef]
- Brūmelis, G.; Strazds, M.; Eglava, Ž. Stand structure and spatial pattern of regeneration of Pinus sylvestris in a natural treed mire in Latvia. Silva Fenn. 2009, 43, 767–781. [Google Scholar] [CrossRef] [Green Version]
- Fulé, P.Z.; Korb, J.E.; Wu, R. Changes in forest structure of a mixed conifer forest, southwestern Colorado, USA. For. Ecol. Manag. 2009, 258, 1200–1210. [Google Scholar] [CrossRef]
- Bekker, M.F.; Taylor, A.H. Fire disturbance, forest structure, and stand dynamics in montane forests of the southern Cascades, Thousand Lakes Wilderness, California, USA. Écoscience 2010, 17, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Konstantinov, A.V. Changes in resilience to fire disturbance in lowland pine forest ecosystems. Biol. Bull. 2011, 38, 974–979. [Google Scholar] [CrossRef]
- Rubiales, J.M.; Morales-Molino, C.; Álvarez, S.G.; García-Antón, M. Negative responses of highland pines to anthropogenic activities in inland Spain: A palaeoecological perspective. Veg. Hist. Archaeobot. 2011, 21, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Hanberry, B.B.; Dey, D.C. Historical range of variability for restoration and management in Wisconsin. Biodivers. Conserv. 2019, 28, 2931–2950. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Kabrick, J.M.; He, H.S.; Palik, B. Historical trajectories and restoration strategies for the Mississippi River Alluvial Valley. For. Ecol. Manag. 2012, 280, 103–111. [Google Scholar] [CrossRef]
- Means, D.B. Vertebrate faunal diversity in longleaf pine savannas. In Longleaf Pine Ecosystems: Ecology, Management, and Restoration; Jose, S., Jokela, E., Miller, D., Eds.; Springer: New York, NY, USA, 2006; pp. 155–213. [Google Scholar]
- Steen, D.A.; Conner, L.M.; Smith, L.L.; Provencher, L.; Hiers, J.K.; Pokswinski, S.; Helms, B.S.; Guyer, C. Bird assemblage response to restoration of fire-suppressed longleaf pine sandhills. Ecol. Appl. 2013, 23, 134–147. [Google Scholar] [CrossRef]
- Kirkman, L.K.; Coffey, K.L.; Mitchell, R.J.; Moser, E.B. Ground cover recovery patterns and life-history traits: Implications for restoration obstacles and opportunities in a species-rich savanna. J. Ecol. 2004, 92, 409–421. [Google Scholar] [CrossRef]
- Means, D.B. Longleaf pine forest, going, going,… In Eastern Old-Growth Forests: Prospects for Rediscovery and Recovery; Davis, M.B., Ed.; Island Press: Washington, DC, USA, 1996; pp. 210–229. [Google Scholar]
- Darracq, A.K.; Boone, W.W., IV; McCleery, R.A. Burn regime matters: A review of the effects of prescribed fire on vertebrates in the longleaf pine ecosystem. For. Ecol. Manag. 2016, 378, 214–221. [Google Scholar] [CrossRef]
- America’s Longleaf Restoration Initiative. 2020. Available online: http://www.americaslongleaf.org (accessed on 17 January 2017).
- Shortleaf Pine Initiative. 2021. Available online: http://www.shortleafpine.net (accessed on 17 January 2021).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanberry, B.B. Transition from Fire-Dependent Open Forests: Alternative Ecosystem States in the Southeastern United States. Diversity 2021, 13, 411. https://doi.org/10.3390/d13090411
Hanberry BB. Transition from Fire-Dependent Open Forests: Alternative Ecosystem States in the Southeastern United States. Diversity. 2021; 13(9):411. https://doi.org/10.3390/d13090411
Chicago/Turabian StyleHanberry, Brice B. 2021. "Transition from Fire-Dependent Open Forests: Alternative Ecosystem States in the Southeastern United States" Diversity 13, no. 9: 411. https://doi.org/10.3390/d13090411
APA StyleHanberry, B. B. (2021). Transition from Fire-Dependent Open Forests: Alternative Ecosystem States in the Southeastern United States. Diversity, 13(9), 411. https://doi.org/10.3390/d13090411