
Review of Harmful Algal Blooms in the Coastal Mediterranean Sea, with a Focus on Greek Waters



Abstract
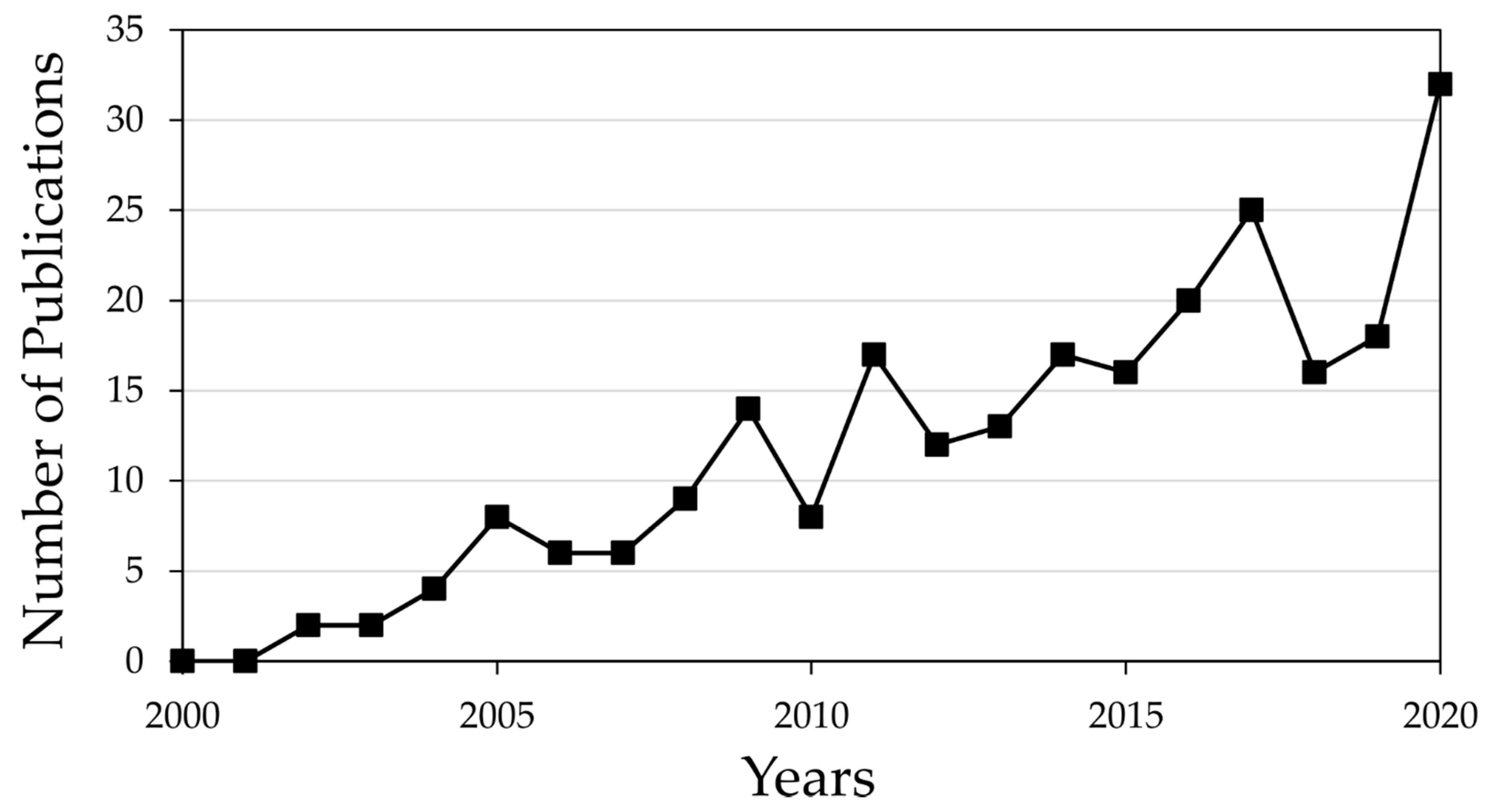
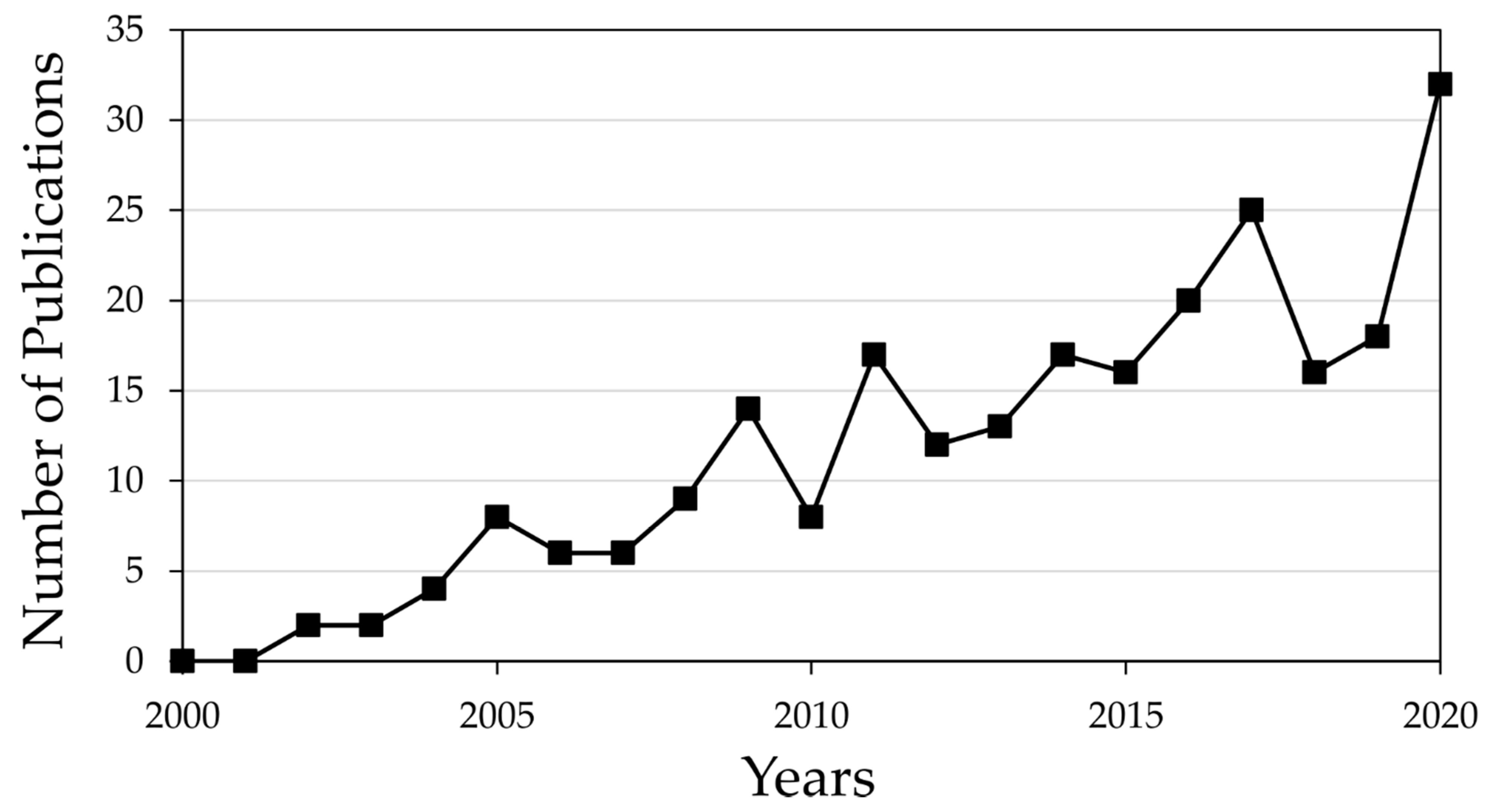
:1. Introduction
2. Literature Search Strategies
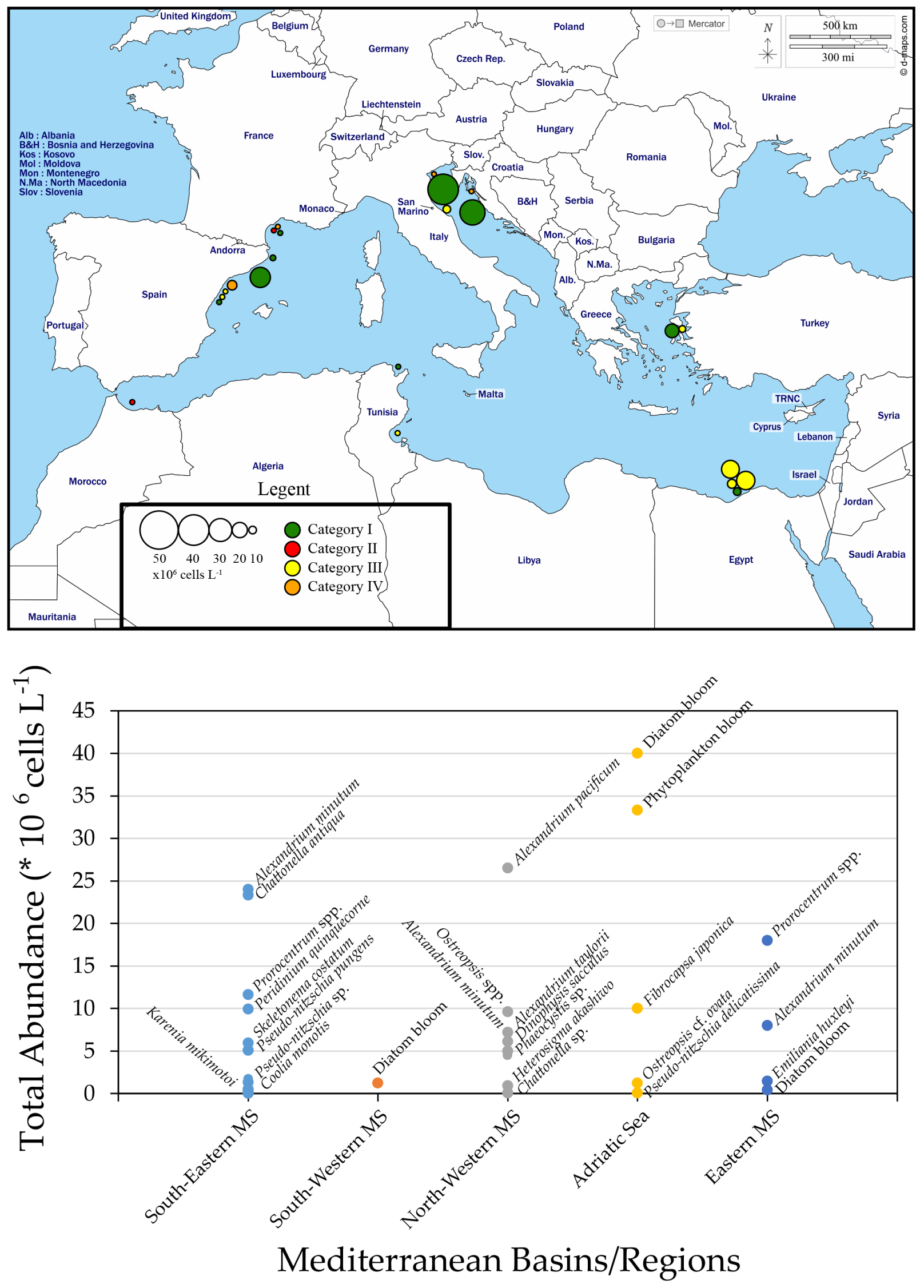
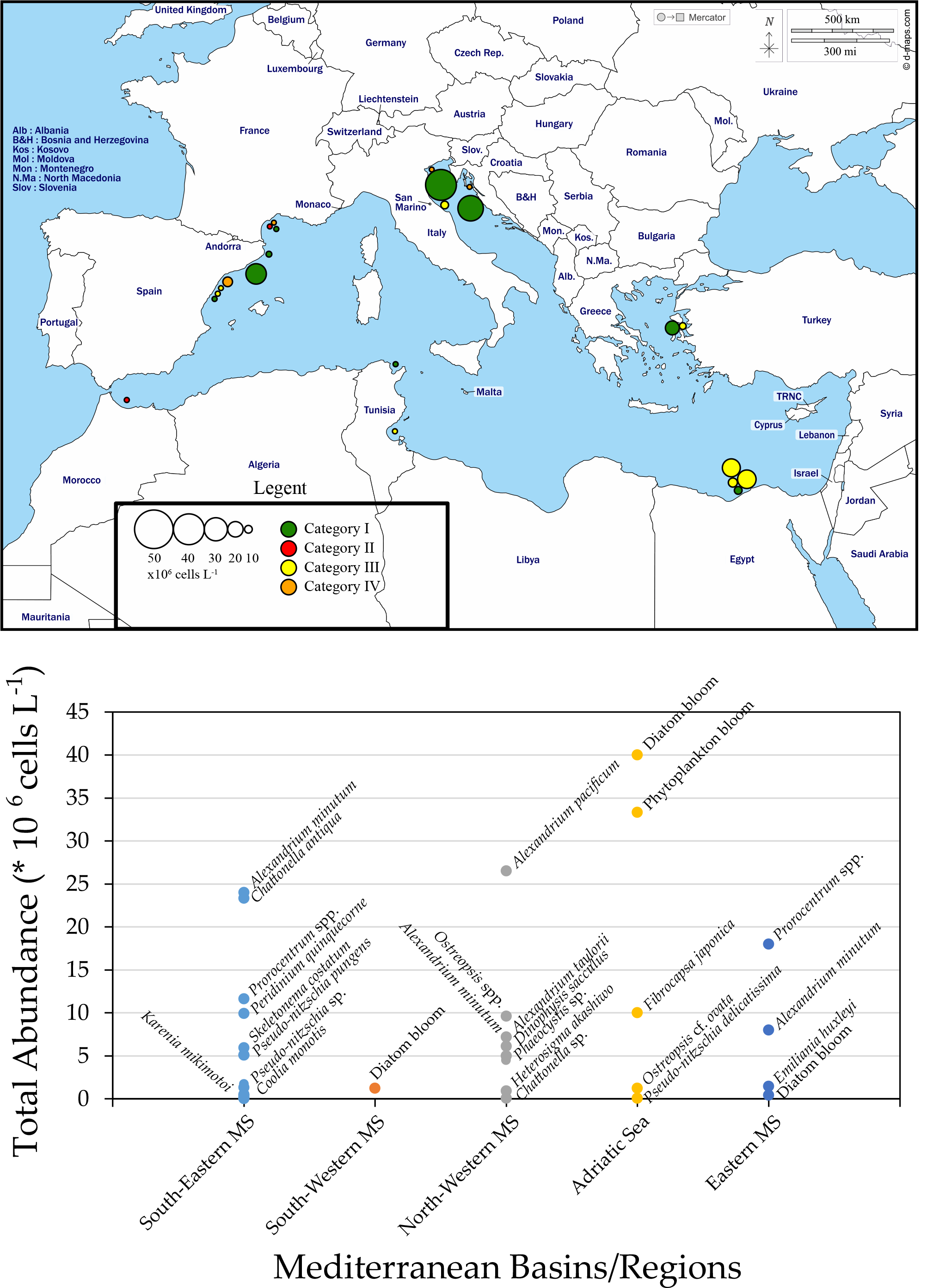
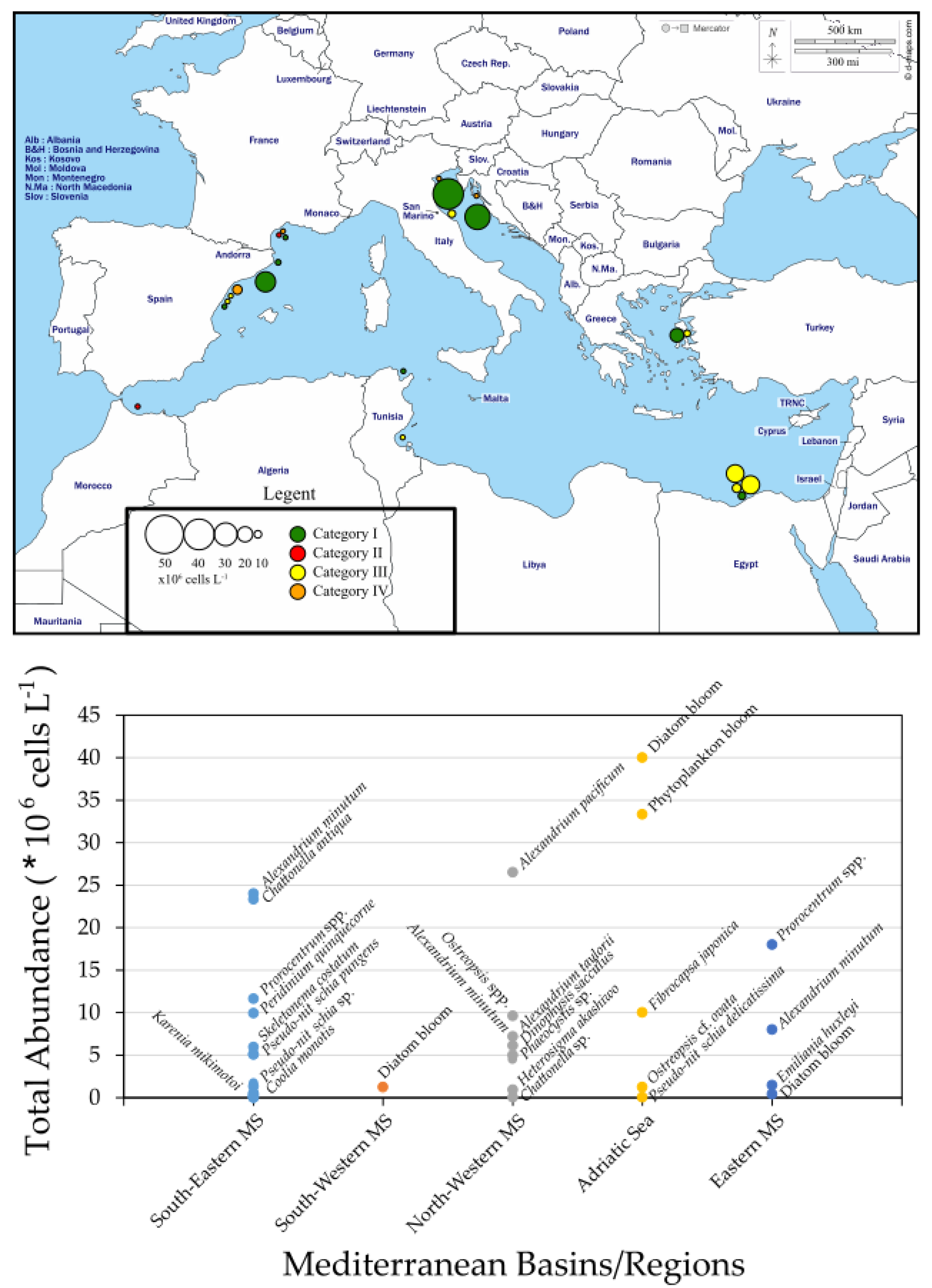
3. HABs in the Mediterranean Sea (MS) Coastal Areas
3.1. South-Eastern MS
3.2. South-Western MS
3.3. North-Western MS
3.4. Adriatic Sea
3.5. Eastern MS (Aegean Sea)
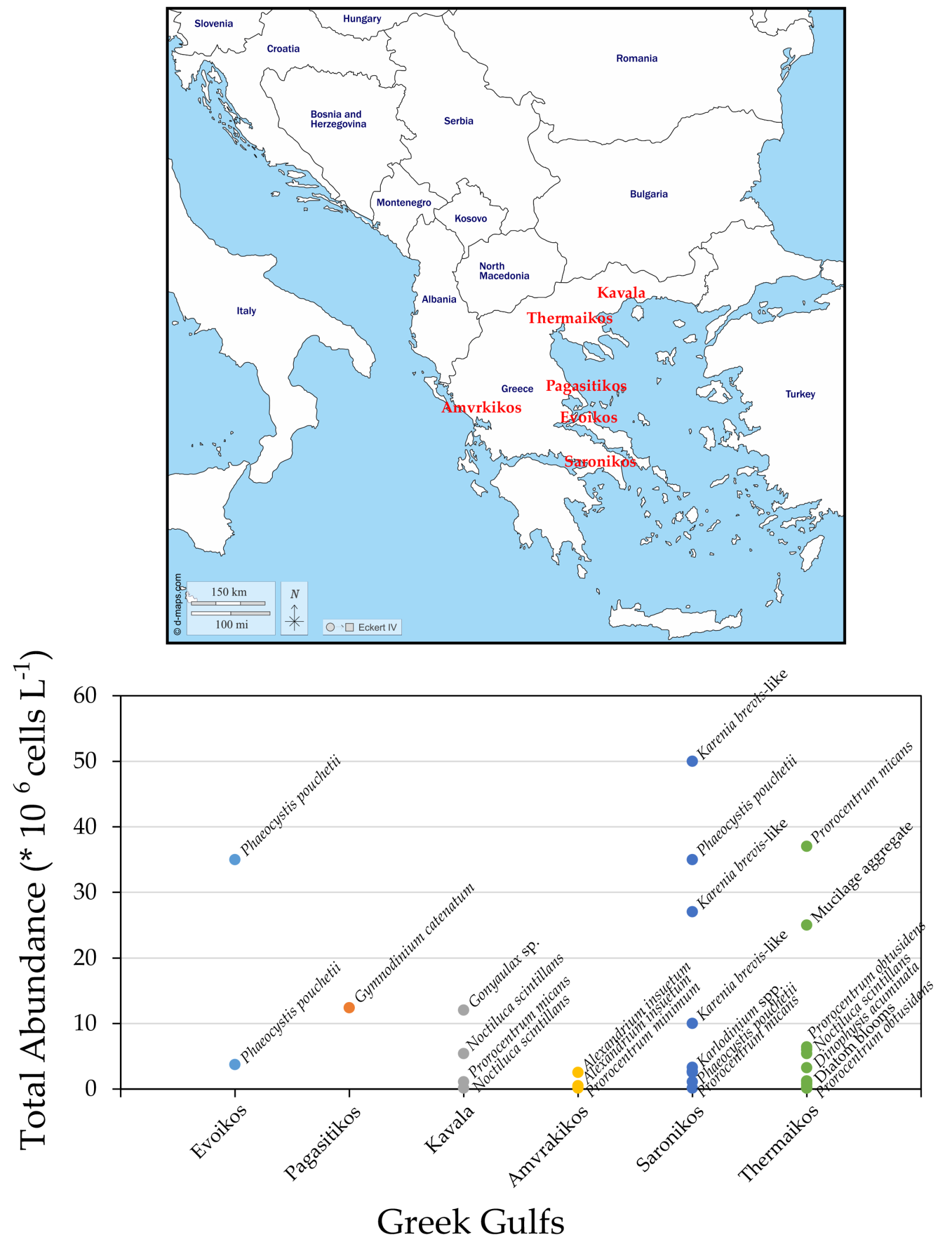
4. HABs in Coastal Areas of Greece
4.1. Saronikos Gulf
4.2. Thermaikos Gulf
4.3. Other Greek Gulfs
5. Conclusions and Perspectives
- rectification of farming and animal husbandry methods according to good agricultural practices;
- establishment of proper and complete physical/chemical wastewater treatment or/and wastewater treatment in algal ponds which enable circular bio-economy [151];
- collecting and disposing of dead coastal organisms via burial or rendering;
- more frequent, systematic, and expanded monitoring programs for harmful algae and algal toxin detection;
- development and application of advanced technologies like the -omics technologies to better understand the diversity, evolution, ecology, and dynamics of HABs;
- outreach and education of public on HAB occurrence and impacts;
- co-operation between regulatory authorities, organizations, academic institutes, researchers, and citizens to exchange relevant knowledge and best practices.
Author Contributions
Funding
Conflicts of Interest
References
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef]
- Von Glasow, R.; Jickells, T.D.; Baklanov, A.; Carmichael, G.R.; Church, T.M.; Gallardo, L.; Hughes, C.; Kanakidou, M.; Liss, P.S.; Mee, L.; et al. Megacities and large urban agglomerations in the coastal zone: Interactions between atmosphere, land, and marine ecosystems. Ambio 2013, 42, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Davidson, K.; Gowen, R.J.; Tett, P.; Bresnan, E.; Harrison, P.J.; McKinney, A.; Milligan, S.; Mills, D.K.; Silke, J.; Crooks, A.M. Harmful algal blooms: How strong is the evidence that nutrient ratios and forms influence their occurrence? Estuarine. Coast. Shelf Sci. 2012, 115, 399–413. [Google Scholar] [CrossRef]
- Guillaud, J.F.; Aminot, A.; Menesguen, A. Urban wastewater disposal and eutrophication risk assessment in the coastal zone. Water Sci. Technol. 1992, 25, 77–86. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Diaz, R.J.; Justic, D. Global change and eutrophication of coastal waters. ICES J. Mar. Sci. 2009, 66, 1528–1537. [Google Scholar] [CrossRef]
- Anderson, D.; Cembella, A.; Hallegraeff, G. Progress in understanding harmful algal blooms: Paradigm shifts and new technologies for research, monitoring, and management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, P. Design and Implementation of Some Harmful Algal Monitoring Systems; IOC Technical Series; UNESCO: Paris, France, 1996; Volume 44. [Google Scholar]
- Garcés, E.; Camp, J. Habitat changes in the Mediterranean Sea and the consequences for harmful algal blooms formation. In Life in the Mediterranean Sea: A Look at Habitat Changes; Stambler, N., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 519–541. [Google Scholar]
- Ferreira, J.G.; Andersen, J.H.; Borja, A.; Bricker, S.B.; Camp, J.; da Silva, M.C.; Garcés, E.; Heiskanen, A.S.; Humborg, C.; Ignatiades, L.; et al. Marine Strategy Framework Directive Task Group 5 report eutrophication. In JRC Scientific and Technical Reports EUR; Scientific and Technical Research Series; Zampoukas, N., Ed.; EU: Brussels, Belgium, 2010. [Google Scholar]
- Zohdi, E.; Abbaspour, M. Harmful algal blooms (red tide): A review of causes, impacts and approaches to monitoring and prediction. Int. J. Environ. Sci. Technol. 2019, 16, 1789–1806. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M. Harmful algal blooms: A global overview. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; United Nations Educational, Scientific and Cultural Organization: Paris, France, 2003. [Google Scholar]
- Pitcher, G.; Louw, D. Harmful algal blooms of the Benguela eastern boundary upwelling system. Harmful Algae 2021, 102, 101898. [Google Scholar] [CrossRef]
- Anderson, D.M.; Burkholder, J.M.; Cochlan, W.P.; Glibert, P.M.; Gobler, C.J.; Heil, C.A.; Kudela, R.M.; Parsons, M.L.; Rensel, J.E.J.; Townsend, D.W.; et al. Harmful algae blooms and eutrophication: Examining linkages from selected coastal regions of the United States. Harmful Algae 2008, 8, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.; Glibert, P.; Burkholder, J. Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Brand, L.; Compton, A. Long-term increase in Karenia brevis abundance along the Southwest Florida Coast. Harmful Algae 2007, 6, 232–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabalais, N.; Turner, E. Gulf of Mexico hypoxia: Past, present, and future. Limnol. Oceanogr. Bull. 2019, 28, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-H.; Lyubchich, V.; Glibert, P. Time series models of decadal trends in the harmful algal species Karlodinium veneficum in Chesapeake Bay. Harmful Algae 2018, 73, 110–118. [Google Scholar] [CrossRef]
- Li, J.; Glibert, P.; Gao, Y. Temporal and spatial changes in Chesapeake Bay water quality and relationships to Prorocentrum minimum, Karlodinium veneficum, and CyanoHAB events, 1991–2008. Harmful Algae 2015, 42, 1–14. [Google Scholar] [CrossRef]
- Glibert, P.; Magnien, R.; Lomas, M.; Alexander, J.; Fan, C.; Haramoto, E.; Trice, M.; Kana, T. Harmful algal blooms in the Chesapeake and Coastal Bays of Maryland, USA: Comparison of 1997, 1998, and 1999 events. Estuaries 2001, 24, 875–883. [Google Scholar] [CrossRef]
- Lu, D.; Qi, Y.; Gu, H.; Dai, X.; Wang, H.; Gao, Y.; Shen, P.; Zhang, Q.; Yu, R.; Lu, S. Causative species of harmful algal blooms in Chinese coastal waters. Algol. Stud. 2014, 145, 145–168. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, T.; Zhou, J. Historical occurrence of algal blooms in the Northern Beibu Gulf of China and implications for future trends. Front. Microbiol. 2019, 10, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, S.; Lim, W.; Lu, D.; Dai, X.; Orlova, T.; Iwataki, M. Harmful algal blooms and associated fisheries damage in East Asia: Current status and trends in China, Japan, Korea and Russia. Harmful Algae 2021, 102, 101787. [Google Scholar] [CrossRef]
- Christaki, U.; Kormas, K.; Genitsaris, S.; Georges, C.; Sime-Ngando, T.; Viscogliosi, E.; Monchy, S. Winter-Summer succession of unicellular eukaryotes in a meso-eutrophic coastal system. Microb. Ecol. 2014, 67, 13–23. [Google Scholar] [CrossRef]
- Lefebvre, A.; Guiselin, N.; Barbet, F.; Artigas, F. Long-term hydrological and phytoplankton monitoring (1992-2007) of three potential eutrophic systems in the eastern English Channel and the Southern Bight of the North Sea. ICES J. Mar. Sci. 2011, 68, 2029–2043. [Google Scholar] [CrossRef] [Green Version]
- Seuront, L.; Vincent, D.; Mitchell, J. Biologically induced modification of seawater viscosity in the eastern English Channel during a Phaeocystis globose spring bloom. J. Mar. Syst. 2006, 61, 118–133. [Google Scholar] [CrossRef]
- Schoemann, V.; Becquevort, S.; Stefels, J.; Rousseau, V.; Lancelot, C. Phaeocystis blooms in the global ocean and their controlling mechanisms: A review. J. Sea Res. 2005, 53, 43–66. [Google Scholar] [CrossRef]
- Willis, C.; Papathanasopoulou, E.; Russel, D.; Artioli, Y. Harmful algal blooms: The impacts on cultural ecosystem services and human well-being in a case study setting, Cornwall, UK. Mar. Policy 2018, 97, 232–238. [Google Scholar] [CrossRef]
- Young, N.; Sharpe, R.; Barciela, R.; Nichols, G.; Davidson, K.; Berdalet, E.; Fleming, L. Marine harmful algal blooms and human health: A systematic scoping review. Harmful Algae 2020, 98, 101901. [Google Scholar] [CrossRef]
- Nicolas, J.; Hoogenboom, R.; Hendriksen, P.; Bodero, M.; Bovee, T.; Rietjens, I.; Gerssen, A. Marine biotoxins and associated outbreaks following seafood consumption: Prevention and surveillance in the 21st century. Glob. Food Secur. 2017, 15, 11–21. [Google Scholar] [CrossRef]
- Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Walsh, C.J.; Nierenberg, K.; Clark, J.; Reich, A.; Hollenbeck, J.; Benson, J.; Cheng, Y.S.; et al. Review of Florida red tide and human health effects. Harmful Algae 2011, 10, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Manivasagan, P.; Kim, S.-K. An overview of harmful algal blooms on marine organisms. In Handbook of Microalgae: Biotechnology Advances; Kim, S.-K., Ed.; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Sha, J.; Xiong, H.; Li, C.; Lu, Z.; Zhang, J.; Zhong, H.; Zhang, W.; Yan, B. Harmful algal blooms and their eco-environmental indication. Chemosphere 2021, 274, 129912. [Google Scholar] [CrossRef]
- Da Costa, J.; De Souza, J.; Texeira, A.; Nabout, J.; Carneiro, F. Eutrophication in aquatic ecosystems: A scientometric study. Acta Limnol. Bras. 2018, 30, 2. [Google Scholar] [CrossRef] [Green Version]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [Green Version]
- Karydis, M.; Kitsiou, D. Eutrophication and environmental policy in the Mediterranean Sea: A review. Environ. Monit. Assess. 2012, 184, 4931–4984. [Google Scholar] [CrossRef]
- Bianchi, C.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea biodiversity between the legacy from the past and a future of change. In Life in the Mediterranean Sea: A Look at Habitat Changes; Stambler, N., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 519–541. [Google Scholar]
- Barale, V.; Jaquet, J.-M.; Ndiaye, M. Algal blooming patterns and anomalies in the Mediterranean Sea as derived from the SeaWiFS data set (1998–2003). Remote. Sens. Environ. 2008, 112, 3300–3313. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’ Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Zingone, A.; Escalera, L.; Aligizaki, K.; Fernandez-Tejedor, M.; Ismael, A.; Montresor, M.; Mozetič, P.; Taş, S.; Totti, C. Toxic marine microalgae and noxious blooms in the Mediterranean Sea: A contribution to the Global HAB Status Report. Harmful Algae 2021, 102, 101843. [Google Scholar] [CrossRef] [PubMed]
- Poulos, S. The Mediterranean and Black Sea marine system: An overview of its physico-geographic and oceanographic characteristics. Earth-Sci. Rev. 2020, 200, 103004. [Google Scholar] [CrossRef]
- Gazeau, F.; Smith, S.; Gentili, B.; Frankignoulle, M.; Gattuso, J.-P. The European coastal zone: Characterization and first assessment of ecosystem metabolism. Estuarine. Coast. Shelf Sci. 2004, 60, 673–694. [Google Scholar] [CrossRef]
- Malago, A.; Bouraoui, F.; Grizzetti, B.; De Roo, A. Modelling nutrient fluxes into the Mediterranean Sea. J. Hydrol. Reg. Stud. 2019, 22, 100592. [Google Scholar] [CrossRef] [PubMed]
- Fierro, P.; Valdovinos, C.; Arismendi, I.; Díaz, G.; Ruiz De Gamboa, M.; Arriagada, L. Assessment of anthropogenic threats to Chilean Mediterranean freshwater ecosystems: Literature review and expert opinions. Environ. Impact Assess. Rev. 2019, 77, 114–121. [Google Scholar] [CrossRef]
- Ferrante, M.; Sciacca, S.; Fallico, R.; Fiore, M.; Conti, G.-O.; Ledda, C. Harmful algal blooms in the Mediterranean Sea: Effects on human health. EuroMediterranean. Biomed. J. 2013, 8, 25–34. [Google Scholar] [CrossRef]
- Marampouti, C.; Buma, A.G.J.; De Boer, M.K. Mediterranean alien harmful algal blooms: Origins and impacts. Environ. Sci. Pollut. Res. 2021, 28, 3837–3851. [Google Scholar] [CrossRef]
- Penna, A.; Bertozzini, E.; Battocchi, C.; Galluzzi, L.; Giacobbe, M.; Vila, M.; Garcés, E.; Lugliè, A.; Magnani, M. Monitoring of HAB species in the Mediterranean Sea through molecular methods. J. Plankton Res. 2007, 29, 19–38. [Google Scholar] [CrossRef] [Green Version]
- Dorgham, M. Eutrophication problem in Egypt. In Eutrophication: Causes, Consequences and Control; Ansari, A., Gill, S., Lanza, G., Rast, W., Eds.; Springer Nature: Dordrecht, The Netherlands, 2011. [Google Scholar]
- Ismael, A. Coastal engineering and harmful algal blooms along Alexandria coast, Egypt. Egypt. J. Aquat. Res. 2014, 40, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Ismael, A. Succession of heterotrophic and mixotrophic dinoflagellates as well as autotrophic microplankton in the harbour of Alexandria, Egypt. J. Plankton Res. 2003, 25, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Ismael, A. First record of Coolia monotis Meunier along Alexandria coast—Egypt. Egypt. J. Aquat. Res. 2014, 40, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Mikhail, S. First monospecific bloom of the harmful raphidophyte Chatonella antiqua (Hada) Ono in Alexandria waters related to water quality and copepod grazing. Chem. Ecol. 2007, 23, 393–407. [Google Scholar] [CrossRef]
- Ayata, S.-D.; Irisson, J.-O.; Aubert, A.; Berline, L.; Dutay, J.-C.; Mayot, N.; Nieblas, A.-E.; D’Ortenzio, F.; Palmieri, J.; Reygondeau, G.; et al. Regionalisation of the Mediterranean basin, a MERMEX synthesis. Prog. Oceanogr. 2018, 163, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Béjaoui, B.; Ben Ismail, S.; Othmani, A.; Hamida, O.B.A.-B.H.; Chevalier, C.; Feki-Sahnoun, W.; Harzallah, A.; Hamida, N.B.H.; Bouaziz, R.; Dahech, S.; et al. Synthesis review of the Gulf of Gabes (eastern Mediterranean Sea, Tunisia): Morphological, climatic, physical, oceanographic, biogeochemical and fisheries features. Estuarine. Coast. Shelf Sci. 2019, 219, 395–408. [Google Scholar] [CrossRef]
- El Kateb, A.; Stalder, C.; Ruggeberg, A.; Neururer, C.; Spangenberg, J.; Spezzaferri, S. Impact of industrial phosphate waste discharge on the marine environment in the Gulf of Gabes (Tunisia). PLoS ONE 2018, 13, e0197731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdennadher, M.; Bellaaj Zouari, A.; Feki Sahnoun, W.; Walha, L.; Mahfoudi, M.; Hamza, A. A long-term study on Coolia monotis distribution from the South-East Mediterranean Sea. Cont. Shelf Res. 2020, 211, 104267. [Google Scholar] [CrossRef]
- Abdennadher, M.; Hamza, A.; Fekih, W.; Hannachi, I.; Zouari Bellaaj, A.; Nejmeddine Bradai, M.; Aleya, L. Factors determining the dynamics of toxic blooms of Alexandrium minutum during a 10-year study along the shallow Southwestern Mediterranean coasts. Estuarine. Coast. Shelf Sci. 2012, 106, 102–111. [Google Scholar] [CrossRef]
- Feki, W.; Hamza, A.; Frossard, V.; Abdennadher, M.; Hannachi, I.; Jacquot, M.; Belhassen, M.; Aleya, L. What are the potential drivers of blooms of the toxic dinoflagellate Karenia selliformis? A 10-year study in the Gulf of Gabes, Tunisia, Southwestern Mediterranean Sea. Harmful Algae 2013, 23, 8–18. [Google Scholar] [CrossRef]
- Hachani, M.; Dhib, A.; Fathalli, A.; Ziadi, B.; Turki, S.; Aleya, L. Harmful epiphytic dinoflagellate assemblages on macrophytes in the Gulf of Tunis. Harmful Algae 2018, 77, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Daly Yahia-Kefi, O.; Souissi, S.; Gómez, F.; Daly Yahia, M.N. Spatio-temporal distribution of the dominant diatom and dinoflagellate species in the Bay of Tunis (SW Mediterranean Sea). Mediterr. Mar. Sci. 2005, 6, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Accoroni, S.; Totti, C. The toxic benthic dinoflagellates of the genus Ostreopsis in temperate areas: A review. Adv. Oceanogr. Limnol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Aissaoui, A.; Dhib, A.; Reguera, B.; Ben Hassine, O.K.; Turki, S.; Aleya, L. First evidence of cell deformation occurrence during a Dinophysis bloom along the shores of the Gulf of Tunis (SW Mediterranean Sea). Harmful Algae 2014, 39, 191–201. [Google Scholar] [CrossRef]
- Garali, S.; Sahrahoui, I.; De la Iglesia, P.; Chalghaf, M.; Diogène, J.; Ksouri, J.; Sakka Hlaili, A. Factors driving the seasonal dynamics of Pseudo-nitzschia species and domoic acid at mussel farming in the SW Mediterranean Sea. Chem. Ecol. 2019, 36, 66–82. [Google Scholar] [CrossRef]
- Hlaili, A.S.; Khalifa, I.S.; Bouchouicha-Smida, D.; Garali, S.M.; Ksouri, J.; Chalghaf, M.; Bates, S.S.; Lundholm, N.; Kooistra WH, C.F.; de la Iglesia, P.; et al. Toxic and potentially toxic diatom blooms in Tunisian (SW Mediterranean) waters: Review of ten years of investigations. Adv. Environ. Res. 2016, 48, 51–69. [Google Scholar]
- Er-Raioui, H.; Bouzid, S.; Marhraoui, M.; Saliot, A. Hydrocarbon pollution of the Mediterranean coastline of Morocco. Ocean Coast. Manag. 2009, 52, 124–129. [Google Scholar] [CrossRef]
- Kessabi, M.; Elhraiki, A.; Nader, B. Contamination of urban, industrial and continental waters by chlorinated hydrocarbon pesticides along the Mediterranean coast of Morocco. Sci. Total. Environ. 1988, 71, 209–214. [Google Scholar] [CrossRef]
- Leblad, B.R.; Amnhir, R.; Reqia, S.; Sitel, F.; Daoudi, M.; Marhraoui, M.; Abdellah, M.K.O.; Veron, B.; Er-Raioui, H.; Laabir, M. Seasonal variations of phytoplankton assemblages in relation to environmental factors in Mediterranean coastal waters of Morocco, a focus on HABs species. Harmful Algae 2020, 96, 101819. [Google Scholar] [CrossRef]
- Taleb, H.; Vale, P.; Jaime, E.; Blaghen, M. Study of paralytic shellfish poisoning toxin profile in shellfish from the Mediterranean shore of Morocco. Toxicon 2001, 39, 1855–1861. [Google Scholar] [CrossRef]
- Rijal Leblad, B.; Lundholm, N.; Goux, D.; Veron, B.; Sagou, R.; Taleb, H.; Nhhala, H.; Er-Raioui, H. Pseudo-nitzschia Peragallo (Bacillariophyceae) diversity and domoic acid accumulation in tuberculate cockles and sweet clams in M’diq Bay, Morocco. Acta Bot. Croat. 2013, 72, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Sebastia, M.-T.; Rodilla, M.; Sanchis, J.-A.; Altur, V.; Gadea, I.; Falco, S. Influence of nutrient inputs from a wetland dominated by agriculture on the phytoplankton community in a shallow harbour at the Spanish Mediterranean coast. Agric. Ecosyst. Environ. 2012, 152, 10–20. [Google Scholar] [CrossRef]
- Gomez-Gutierrez, A.I.; Jover, E.; Bodineau, L.; Albaiges, J.; Bayona, J.M. Organic contaminant loads in the Western Mediterranean Sea: Estimate of Ebro river inputs. Chemosphere 2006, 65, 224–236. [Google Scholar] [CrossRef]
- Vila, M.; Masó, M. Phytoplankton functional groups and harmful algal species in anthropogenically impacted waters of the NW Mediterranean Sea. Sci. Mar. 2005, 69, 31–45. [Google Scholar] [CrossRef]
- Estrada, M.; Sole, J.; Angles, S.; Garcés, E. The role of resting cysts in Alexandrium minutum population dynamics. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 308–321. [Google Scholar] [CrossRef]
- Bravo, I.; Vila, M.; Masó, M.; Figueroa, R.-I.; Ramilo, I. Alexandrium catenella and Alexandrium minutum blooms in the Mediterranean Sea: Toward the identification of ecological niches. Harmful Algae 2008, 7, 515–522. [Google Scholar] [CrossRef]
- Vila, M.; Giacobbe, M.; Masó, M.; Gangemi, E.; Penna, A.; Sampedro, N.; Azzaro, F.; Camp, J.; Galluzzi, L. A comparative study on recurrent blooms of Alexandrium minutum in two Mediterranean coastal areas. Harmful Algae 2005, 4, 673–695. [Google Scholar] [CrossRef]
- Vila, M.; Garcés, E.; Masó, M.; Camp, J. Is the distribution of the toxic dinoflagellate Alexandrium catenella expanding along the NW Mediterranean coast? Mar. Ecol. Prog. Ser. 2001, 222, 73–83. [Google Scholar] [CrossRef]
- Satta, C.; Pulina, S.; Padedda, B.; Penna, A.; Sechi, N.; Lugliè, A. Water discoloration events caused by the harmful dinoflagellate Alexandrium taylorii Balech in a new beach of the Western Mediterranean Sea (Platamona beach, North Sardinia). Adv. Oceanogr. Limnol. 2010, 1, 259–269. [Google Scholar] [CrossRef]
- Garcés, E.; Masó, M.; Camp, J. A recurrent and localized dinoflagellate bloom in a Mediterranean beach. J. Plankton. Res. 1999, 21, 2373–2391. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Altares, M.; Casanova, A.; Fernandez-Tejedor, M.; Diogene, J.; De la Iglesia, P. Bloom of Dinophysis spp. Dominated by D. sacculus and its related diarrhetic shellfish poisoning (DSP) outbreak in Alfacs Bay (Catalonia, NW Mediterranean Sea): Identification of DSP toxins in phytoplankton, shellfish and passive samplers. Reg. Stud. Mar. Sci. 2016, 6, 19–28. [Google Scholar] [CrossRef]
- Busch, J.; Andree, K.; Diogene, J.; Fernandez-Tejedor, M.; Toebe, K.; John, U.; Krock, B.; Tillmann, U.; Cembella, A. Toxigenic algae and associated phycotoxins in two coastal embayments in the Ebro Delta (NW Mediterranean). Harmful Algae 2016, 55, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, M.; Masó, M.; Sampedro, N.; Illoul, H.; Arin, L.; Garcés, E.; Giacobbe, M.; Alvarez, J.; Camp, J. The genus Ostreopsis in the recreational waters along the Catalan Coast and Balearic Islands (NW Mediterranean Sea): Are they the origin of human respiratory difficulties? In Proceedings of the 12th International Conference on Harmful Algae, Copenhagen, Denmark, 4–8 September 2006; Moestrup, Ø., Doucette, G., Enevoldsen, H., Godhe, A., Hallegraeff, G., Luckas, B., Lundholm, N., Lewis, J., Rengefors, K., Sellner, K., et al., Eds.; International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO: Copenhagen, Denmark, 2008. [Google Scholar]
- Loureiro, S.; Garcés, E.; Fernandez-Tejedor, M.; Vaque, D.; Camp, J. Pseudo-nitzschia spp. (Bacillariophyceae) and dissolved organic matter (DOM) dynamics in the Ebro Delta (Alfacs Bay, NW Mediterranean Sea). Estuar. Coast. Shelf Sci. 2009, 83, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Pruski, A.; Buscail, R.; Bourrin, F.; Vetion, G. Influence of coastal Mediterranean rivers on the organic matter composition and reactivity of continental shelf sediments: The case of the Tet River (Gulf of Lions, France). Cont. Shelf Res. 2019, 181, 156–173. [Google Scholar] [CrossRef]
- Van Den Broeck, N.; Moutin, T. Phosphate in the sediments of the Gulf of Lions (NW Mediterranean Sea), relationship with input by the river Rhone. Hydrobiologia 2002, 472, 85–94. [Google Scholar] [CrossRef]
- Van Wambeke, F.; Heussner, S.; Diaz, F.; Raimbault, P.; Conan, P. Small-scale variability in the coupling/uncoupling of bacteria, phytoplankton and organic carbon fluxes along the continental margin of the Gulf of Lions, Northwestern Mediterranean Sea. J. Mar. Syst. 2002, 33–34, 411–429. [Google Scholar] [CrossRef]
- Belin, C.; Soudant, D.; Zouher Amzil, Z. Three decades of data on phytoplankton and phycotoxins on the French coast: Lessons from REPHY and REPHYTOX. Harmful Algae 2021, 102, 101733. [Google Scholar] [CrossRef]
- Heil, C.; Glibert, P.; Fan, C. Prorocentrum minimum (Pavillard) Schiller: A review of harmful algal bloom species of growing worldwide importance. Harmful Algae 2005, 4, 449–470. [Google Scholar] [CrossRef]
- Amzil, Z.; Sibat, M.; Chomerat, N.; Grossel, H.; Marco-Miralles, F.; Lemee, R.; Nezan, E.; Sechet, V. Ovatoxin-a and Palytoxin accumulation in seafood in relation to Ostreopsis cf. ovata blooms on the French Mediterranean coast. Mar. Drugs 2012, 10, 477–496. [Google Scholar]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemée, R.; Kantin, R.; Lasalle, Je.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health impact of unicellular algae of Ostreopsis genus blooms in the Mediterranean Sea: Experience of the French Mediterranean coast surveillance network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiroga, I. Pseudo-nitzschia blooms in the bay of Banyuls-Sur-Mer, Northwestern Mediterranean Sea. Diatom. Res. 2006, 21, 91–104. [Google Scholar]
- Penna, N.; Capellacci, S.; Ricci, F. The influence of the Po River discharge on phytoplankton bloom dynamics along the coastline of Pesaro (Italy) in the Adriatic Sea. Mar. Pollut. Bull. 2004, 48, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Totti, C.; Romagnoli, T.; Accoroni, S.; Coluccelli, A.; Pellegrini, M.; Campanelli, A.; Grilli, F.; Marini, M. Phytoplankton communities in the Northwestern Adriatic Sea: Interdecadal variability over a 30-years period (1988–2016) and relationships with meteoclimatic drivers. J. Mar. Syst. 2019, 193, 137–153. [Google Scholar] [CrossRef]
- Penna, A.; Ingarao, C.; Ercolessi, M.; Marco Rocchi, M.; Penna, N. Potentially harmful microalgal distribution in an area of the NW Adriatic coastline: Sampling procedure and correlations with environmental factors. Estuar. Coast. Shelf Sci. 2006, 70, 307–316. [Google Scholar] [CrossRef]
- Valbi, E.; Ricci, F.; Capellacci, S.; Casabianca, S.; Scardi, M.; Penna, A. A model predicting the PSP toxic dinoflagellate Alexandrium minutum occurrence in the coastal waters of the NW Adriatic Sea. Sci. Rep. 2019, 9, 4166. [Google Scholar] [CrossRef] [Green Version]
- Cerino, F.; Fornasero, D.; Kralj, M.; Giani, M.; Cabrini, M. Phytoplankton temporal dynamics in the coastal waters of the north-eastern Adriatic Sea (Mediterranean Sea) from 2010 to 2017. Nat. Conserv. 2019, 34, 343–372. [Google Scholar] [CrossRef]
- Fonda Umani, S.; Beran, A.; Parlato, S.; Virgilio, D.; Zollet, D.; De Olazabal, A.; Lazzarini, B.; Cabrini, M. Noctiluca scintillans MACARTNEY in the Northern Adriatic Sea: Long-term dynamics, relationships with temperature and eutrophication, and role in the food web. J. Plankton Res. 2004, 26, 545–561. [Google Scholar] [CrossRef] [Green Version]
- Accoroni, S.; Romagnoli, T.; Colombo, F.; Pennesi, C.; Di Camillo, C.G.; Marini, M.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; et al. Ostreopsis cf. ovatabloom in the northern Adriatic Sea during summer 2009: Ecology, molecular characterization and toxin profile. Mar. Pollut. Bull. 2011, 62, 2512–2519. [Google Scholar] [PubMed]
- Monti, M.; Minocci, M.; Beran, A.; Ivesa, L. First record of Ostreopsis cf. ovata on macroalgae in the Northern Adriatic Sea. Mar. Pollut. Bull. 2007, 54, 598–601. [Google Scholar] [CrossRef]
- Pistocchi, R.; Guerrini, F.; Pezzolesi, L.; Riccardi, M.; Vanucci, S.; Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Fattorusso, E.; Tartaglione, L.; et al. Toxin levels and profiles of microalgae from the North-Western Adriatic Sea—15 years of studies on cultured species. Mar. Drugs 2012, 10, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Pompei, M.; Mazziotti, C.; Guerrini, F.; Cangini, M.; Pigozzi, S.; Benzi, M.; Palamidesi, S.; Boni, L.; Pistocchi, R. Correlation between the presence of Gonyaulax fragilis (Dinophyceae) and the mucilage phenomena of the Emilia-Romagna coast (Northern Adriatic Sea). Harmful Algae 2003, 2, 301–316. [Google Scholar] [CrossRef]
- Pezzolesi, L.; Cucchiari, E.; Guerrini, F.; Pasteris, A.; Galletti, P.; Tagliavini, E.; Totti, C.; Pistocchi, R. Toxicity evaluation of Fibrocapsa japonica from the Northern Adriatic Sea through a chemical and toxicological approach. Harmful Algae 2010, 9, 504–514. [Google Scholar] [CrossRef]
- Vilicic, D.; Djakovac, T.; Buric, Z.; Bosak, S. Composition and annual cycle of phytoplankton assemblages in the Northeastern Adriatic Sea. Bot. Mar. 2009, 52, 291–305. [Google Scholar] [CrossRef]
- Pfannkuchen, M.; Godrijan, J.; Pfannkuchen, D.M.; Iveša, L.; Kružić, P.; Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; et al. Toxin-producing Ostreopsis cf. ovata are likely to bloom undetected along coastal areas. Environ. Sci. Technol. 2012, 46, 5574–5582. [Google Scholar] [PubMed]
- Gladan Živana, N.; Matić, F.; Arapov, J.; Skejić, S.; Bužančić, M.; Bakrač, A.; Straka, M.; Dekneudt, Q.; Grbec, B.; Garber, R.; et al. The relationship between toxic phytoplankton species occurrence and environmental and meteorological factors along the Eastern Adriatic coast. Harmful Algae 2020, 92, 101745. [Google Scholar] [CrossRef]
- Maric, D.; Kraus, R.; Godrijan, J.; Supic, N.; Djakovac, T.; Preca, R. Phytoplankton response to climatic and anthropogenic influences in the North-eastern Adriatic during the last four decades. Estuarine. Coast. Shelf Sci. 2012, 115, 98–112. [Google Scholar] [CrossRef]
- Ujević, I.; Ninčević-Gladan, Ž.; Roje, R.; Skejić, S.; Arapov, J.; Marasović, I. Domoic acid—A new toxin in the Croatian Adriatic shellfish toxin profile. Molecules 2010, 15, 6835–6849. [Google Scholar] [CrossRef] [Green Version]
- Goksel, C.; Ipbuker, C.; Bildirici, O.; Ulugtekin, N. The Aegean Sea with numbers. In The Aegean Sea Marine Biodiversity, Fisheries, Conservation and Governance; Katağan, T., Tokaç, A., Beşiktepe, Ş., Öztürk, B., Eds.; Publication No: 41; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2015. [Google Scholar]
- Bizsel, K.; Bizsel, N. Harmful Algal Blooms (HABs) and Aegean Sea. In The Aegean Sea Marine Biodiversity, Fisheries, Conservation and Governance; Katağan, T., Tokaç, A., Beşiktepe, Ş., Öztürk, B., Eds.; Publication No: 41; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2015. [Google Scholar]
- Kontas, A.; Kucuksezgin, F.; Altay, O.; Uluturhan, E. Monitoring of eutrophication and nutrient limitation in the Izmir Bay (Turkey) before and after wastewater treatment plant. Environ. Int. 2004, 29, 1057–1062. [Google Scholar] [CrossRef]
- Inanan, B.; Unluoglu, A.; Bizsel, K.; Bizsel, N. On the distribution of nuisance, potentially and obligatory toxic phytoplankton species during the red-tide (Izmir Bay, Aegean Sea). Rapp. Comm. Int. Mer. Medit. 2007, 38, 373. [Google Scholar]
- Koray, T.; Büyüki, I.K.; Parlak, H.; Gökpinar, E. Eutrophication processes and algal blooms (red-tides) in Izmir Bay. In Final Reports on Research Projects Dealing with Eutrophication and Heavy Metal Accumulation; MAP Technical Reports Series No. 104; United Nations Environment Programme: Athens, Greece, 1996. [Google Scholar]
- Ergül, H.-A.; Aksan, S.; Ipsiroglu, M. Assessment of the consecutive harmful dinoflagellate blooms during 2015 in the Izmir Bay (Marmara Sea). Acta Oceanol. Sin. 2017, 37, 91–101. [Google Scholar] [CrossRef]
- Aktan, Y.; Tufekc, V.; Tufekc, H.; Aykulu, G. Distribution patterns, biomass estimates and diversity of phytoplankton in Izmir Bay (Turkey). Estuar. Coast. Shelf Sci. 2005, 64, 372–384. [Google Scholar] [CrossRef]
- Tas, S.; Ergül, H.-A.; Balkis, N. Harmful Algal Blooms (HABs) and mucilage formations in the Sea of Marmara. In The Aegean Sea Marine Biodiversity, Fisheries, Conservation and Governance; Katağan, T., Tokaç, A., Beşiktepe, Ş., Öztürk, B., Eds.; Publication No: 42; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2016. [Google Scholar]
- Satsmadjis, J.; Friligos, N. Red tide in Greek waters. Vie Milieu/Life Environ. 1983, 33, 111–117. [Google Scholar]
- Ignatiades, L.; Gotsis-Skretas, O. A review of toxic and harmful algae in Greek coastal waters (E. Mediterranean Sea). Toxins 2010, 2, 1019–1037. [Google Scholar] [CrossRef] [Green Version]
- Ignatiades, L.; Gotsis-Skretas, O.; Metaxatos, A. Field and culture studies on the ecophysiology of the toxic dinoflagellate Alexandrium minutum (Halim) present in Greek coastal waters. Harmful Algae 2007, 6, 153–165. [Google Scholar] [CrossRef]
- Patoucheas, P.; Koukousioura, O.; Psarra, S.; Aligizaki, K.; Dimiza, M.; Skampa, E.; Michailidis, I.; Nomikou, P.; Triantaphyllou, M. Phytoplankton community structure changes during autumn and spring in response to environmental variables in Methana, Saronikos Gulf, Greece. Environ. Sci. Pollut. Res. 2021, 28, 33854–33865. [Google Scholar] [CrossRef]
- Tsiamis, K.; Panayotidis, P.; Salomidi, M.; Pavlidou, A.; Kleinteich, J.; Balanika, K.; Küpper, F. Macroalgal community response to re-oligotrophication in Saronikos Gulf. Mar. Ecol. Prog. Ser. 2013, 472, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Evangeliou, N.; Florou, H. Spatial distribution and mobility of organic carbon (POC and DOC) in a coastal Mediterranean environment (Saronikos Gulf, Greece) during 2007–2009 period. Environ. Sci. Pollut. Res. 2013, 20, 5708–5721. [Google Scholar] [CrossRef] [PubMed]
- Moncheva, S.; Gotsis-Skretas, O.; Pagou, K.; Krastev, A. Phytoplankton blooms in Black Sea and Mediterranean coastal ecosystems subjected to anthropogenic eutrophication: Similarities and differences. Estuarine. Coast. Shelf Sci. 2001, 53, 281–295. [Google Scholar] [CrossRef] [Green Version]
- Dimiza, M.D.; Koukousioura, O.; Michailidis, I.; Dimou, V.-G.; Navrozidou, V.; Aligizaki, K.; Seferlis, M. Seasonal living coccolithophore distribution in the enclosed coastal environments of the Thessalonki Bay (Thermaikos Gulf, NW Aegean Sea). Rev. Micropaléontol. 2020, 69, 100449. [Google Scholar] [CrossRef]
- Androulidakis, Y.; Kolovoyiannis, V.; Makris, C.; Krestenitis, Y.; Baltikas, V.; Stefanidou, N.; Chatziantoniou, A.; Topouzelis, K.; Moustaka-Gouni, M. Effects of ocean circulation on the eutrophication of a Mediterranean gulf with river inlets: The Northern Thermaikos Gulf. Cont. Shelf Res. 2021, 221, 104416. [Google Scholar] [CrossRef]
- Patoucheas, D.; Savvidis, Y. A hydrobiological model as a tool for the detection of Harmful Algal Bloom (HAB) episodes: Application to Thermaikos Gulf. Fresen. Environ. Bull. 2010, 19, 2100–2108. [Google Scholar]
- Nikolaidis, N.P.; Karageorgis, A.; Kapsimalis, V.; Marconis, G.; Drakopoulou, P.; Kontoyiannis, H.; Krasakopoulou, E.; Pavlidou, E.; Pagou, K. Circulation and nutrient modeling of Thermaikos Gulf, Greece. J. Mar. Syst. 2006, 60, 51–62. [Google Scholar] [CrossRef]
- Nikolaidis, G.; Koukaras, K.; Aligizaki, K.; Heracleous, A.; Kalopesa, E.; Moschandreou, K.; Tsolaki, E.; Mantoudis, A. Harmful microalgal episodes in Greek coastal waters. J. Biol. Res. 2005, 3, 77–85. [Google Scholar]
- Economou, V.; Papadopoulou, C.; Brett, M.; Kansouzidou, A.; Charalabopoulos, K.; Filioussis, G.; Seferiadis, K. Diarrheic shellfish poisoning due to toxic mussel consumption: The first recorded outbreak in Greece. Food Addit. Contam. 2007, 24, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Reizopoulou, S.; Strogyloudi, E.; Giannakourou, A.; Pagou, K.; Hatzianestis, I.; Pyrgaki, C.; Graneli, E. Okadaic acid accumulation in microfilter feeders subjected to natural blooms of Dinophysis acuminata. Harmful Algae 2008, 7, 228–234. [Google Scholar] [CrossRef]
- Mouratidou, T.; Kaniou-Grigoriadou, I.; Samara, C.; Kouimtzis, T. Detection of the marine toxin okadaic acid in mussels during a Diarrhetic Shellfish Poisoning (DSP) episode in Thermaikos Gulf, Greece, using biological, chemical and immunological methods. Sci. Total. Environ. 2006, 366, 894–904. [Google Scholar] [CrossRef]
- Koukaras, K.; Nikolaidis, G. Dinophysis blooms in Greek coastal waters (Thermaikos Gulf, NW Aegean Sea). J. Plankton. Res. 2004, 26, 445–457. [Google Scholar] [CrossRef] [Green Version]
- Genitsaris, S.; Stefanidou, N.; Moustaka-Gouni, M.; Sommer, U.; Tsipas, G. Variability and community composition of marine unicellular eukaryote assemblages in a eutrophic Mediterranean urban coastal area with marked plankton blooms and red tides. Diversity 2020, 12, 114. [Google Scholar] [CrossRef] [Green Version]
- Genitsaris, S.; Stefanidou, N.; Sommer, U.; Moustaka-Gouni, M. Phytoplankton blooms, red tides and mucilaginous aggregates in the urban Thessaloniki Bay, Eastern Mediterranean. Diversity 2019, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Aligizaki, K.; Arvaniti, E.; Nikolopoulou, I.; Kaliora, A.; Arsenakis, M. Gonyaulax spinifera bloom in Thermaikos Gulf. Harmful Algal News 2016, 52, 6–7. [Google Scholar]
- Friligos, N. Nutrient conditions in the Euboikos Gulf (West Aegean). Mar. Pollut. Bull. 1985, 16, 435–439. [Google Scholar] [CrossRef]
- Metaxatos, A.; Panagiotopoulos, C.; Ignatiades, L. Monosaccharide and aminoacid composition of mucilage material produced from a mixture of four planktonic taxa. J. Exp. Mar. Biol. Ecol. 2003, 294, 203–217. [Google Scholar] [CrossRef]
- Petihakis, G.; Triantafyllou, G.; Pollani, A.; Koliou, A.; Theodorou, A. Field data analysis and application of a complex water column biogeochemical model in different areas of a semi-enclosed basin: Towards the development of an ecosystem management tool. Mar. Environ. Res. 2005, 59, 493–518. [Google Scholar] [CrossRef]
- Theodorou, A. Ecological monitoring studies for Volos Sea outfall (Pagassitikos Gulf, Western Aegean Sea). Water Sci. Technol. 1995, 32, 273–280. [Google Scholar] [CrossRef]
- Friligos, N.; Gotsis-Skretas, O. Eutrophication and red tide in Aegean coastal waters. Toxicol. Environ. Chem. 1989, 24, 171–180. [Google Scholar] [CrossRef]
- Melidis, P.; Sylaios, G. Assessment of urban pollution footprint in a typical Mediterranean semi-enclosed gulf. Environ. Process. 2017, 4, 421–437. [Google Scholar] [CrossRef]
- Sylaios, G.; Stamatis, N.; Kallianiotis, A.; Vidoris, P. Monitoring water quality and assessment of land-based nutrient loadings and cycling in Kavala Gulf. Water Resour. Manag. 2005, 19, 713–735. [Google Scholar] [CrossRef]
- Gotsis-Skretas, O. Red tides in Greek waters. In Report of the ICES Study Group on the “Dynamics of Harmful Algal Blooms in Coastal Waters: 7–9 April 1992—Vigo, Spain; General Secretary ICES: Copenhagen, Denmark, 1992. [Google Scholar]
- Markogianni, V.; Varkitzi, I.; Pagou, K.; Pavlidou, A.; Dimitriou, E. Nutrient flows and related impacts between a Mediterranean river and the associated coastal area. Cont. Shelf Res. 2017, 134, 1–14. [Google Scholar] [CrossRef]
- Akoumianaki, I.; Papaspyrou, S.; Kormas, K.; Nikolaidou, A. Environmental variation and macrofauna response in a coastal area influenced by land runoff. Estuarine. Coast. Shelf Sci. 2013, 132, 34–44. [Google Scholar] [CrossRef]
- Varkitzi, I.; Markogianni, V.; Pantazi, M.; Pagou, K.; Pavlidou, A.; Dimitriou, E. Effect of river inputs on environmental status and potentially harmful phytoplankton in an Eastern Mediterranean coastal area (Maliakos Gulf, Greece). Mediterr. Mar. Sci. 2018, 19, 326–343. [Google Scholar] [CrossRef] [Green Version]
- Kountoura, K.; Zacharias, I. Temporal and spatial distribution of hypoxic/seasonal anoxic zone in Amvrakikos Gulf, Western Greece. Estuarine. Coast. Shelf Sci. 2011, 94, 123–128. [Google Scholar] [CrossRef]
- Ferentinos, G.; Papatheodorou, G.; Geraga, M.; Iatrou, M.; Fakiris, E.; Christodoulou, D.; Dimitriou, E.; Koutsikopoulos, C. Fjord water circulation patterns and dysoxic/anoxic conditions in a Mediterranean semi-enclosed embayment in the Amvrakikos Gulf, Greece. Estuarine. Coast. Shelf Sci. 2010, 88, 473–481. [Google Scholar] [CrossRef]
- Kountoura, K.; Zacharias, I. Trophic state and oceanographic conditions in Amvrakikos Gulf: Evaluation and monitoring. Desalination Water Treat. 2013, 51, 2934–2944. [Google Scholar] [CrossRef]
- Council Directive 91/676/EEC of 12 December 1991 Concerning the Protection of Waters against Pollution Caused by Nitrates from Agricultural Sources. Official Journal of the European Communities, No L 375/1. Available online: https://eur-lex.europa.eu/eli/dir/1991/676/oj (accessed on 31 May 2021).
- Council Directive 91/271/EEC of 21 May 1991 concerning urban waste-water treatment. Official Journal of the European Communities, No L 135/40.
- Gallardo-Rodriguez, J.; Astuya-Villalon, A.; Llanos-Rivera, A.; Avello-Fontalba, V.; Ulloa-Jofre, V. A critical review on control methods for harmful algal blooms. Rev. Aquacult. 2019, 11, 661–684. [Google Scholar] [CrossRef]
- Sutherland, D.; Ralph, P. 15 years of research on wastewater treatment high rate algal ponds in New Zealand: Discoveries and future directions. N. Z. J. Bot. 2020, 58, 334–357. [Google Scholar] [CrossRef]
- Heil, C.; Steidinger, K. Monitoring, management, and mitigation of Karenia blooms in the eastern Gulf of Mexico. Harmful Algae 2009, 8, 611–617. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HAB Categories | Description | Responsible Algae | |
|---|---|---|---|
| I | Water discoloration | HABs that principally cause harmless discoloration of seawater; result in decreased recreational value of affected area due to low water clarity; bloom can grow so dense that they cause oxygen depletion leading to death of fish and benthic invertebrates under extreme conditions in confined bays. | Dinoflagellates: Noctiluca scintillans, Ceratium spp., Prorocentrum micans, Heterocapsa triquetra, Akashiwo sanguinea, Gonyaulax polygramma, Scrippsiella trochoidea, Peridinium quinquecorne Euglenophytes: Eutreptiella spp. Haptophytes: Phaeocystis spp. Diatoms: Skeletonema costatum-complex |
| II | Toxin accumulation harmful to humans | HABs that produce toxins that accumulate in food chains causing gastrointestinal and neurological symptoms to humans and animals; such as Paralytic Shellfish Poisoning (PSP), Diarrhetic Shellfish Poisoning (DSP), Neurotoxic Shellfish Poisoning (NSP), Amnesic Shellfish Poisoning (ASP) and Ciguatera Fish Poisoning (CFP). | Dinoflagellates: Alexandrium spp., Gymnodinium catenatum, (PSP); Dinophysis acuminata, Dinophysis spp., Prorocentrum lima (DSP); Karenia brevis, Karenia spp. (NSP) Gambierdiscus spp. (CFP), Coolia spp. Diatoms: Pseudo-nitzschia spp. (ASP) |
| III | Toxin production harmful to wildlife | HABs usually non-toxic to humans but toxic to fish and invertebrates (especially in intensive aquaculture systems), e.g., by intoxication or by causing damage or clogging of the gills. | Dinoflagellates: Alexandrium tamarense-complex, Gyrodinium aureolum, Karenia mikimotoi, Karlodinium micrum, Haptophytes: Chrysochromulina polylepis Diatoms: Prymnesium parvum, Prymnesium patelliferum Raphidiophytes: Heterosigma akashiwo, Chattonella antiqua Pelagophytes: Aureococcus anophagefferens Cyanobacteria: Nodularia spumigena |
| IV | Production of aerosolized toxins | HABs that produce toxins toxic to humans, which are transferred by air in aerosols from the bloom area to the coast. | Dinoflagellates: Pfiesteria spp., Karenia brevis, Ostreopsis spp. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsikoti, C.; Genitsaris, S. Review of Harmful Algal Blooms in the Coastal Mediterranean Sea, with a Focus on Greek Waters. Diversity 2021, 13, 396. https://doi.org/10.3390/d13080396
Tsikoti C, Genitsaris S. Review of Harmful Algal Blooms in the Coastal Mediterranean Sea, with a Focus on Greek Waters. Diversity. 2021; 13(8):396. https://doi.org/10.3390/d13080396
Chicago/Turabian StyleTsikoti, Christina, and Savvas Genitsaris. 2021. "Review of Harmful Algal Blooms in the Coastal Mediterranean Sea, with a Focus on Greek Waters" Diversity 13, no. 8: 396. https://doi.org/10.3390/d13080396
APA StyleTsikoti, C., & Genitsaris, S. (2021). Review of Harmful Algal Blooms in the Coastal Mediterranean Sea, with a Focus on Greek Waters. Diversity, 13(8), 396. https://doi.org/10.3390/d13080396