
Cultivable Endophytic Bacteria in Seeds of Dongxiang Wild Rice and Their Role in Plant-Growth Promotion

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Endophytic Bacteria
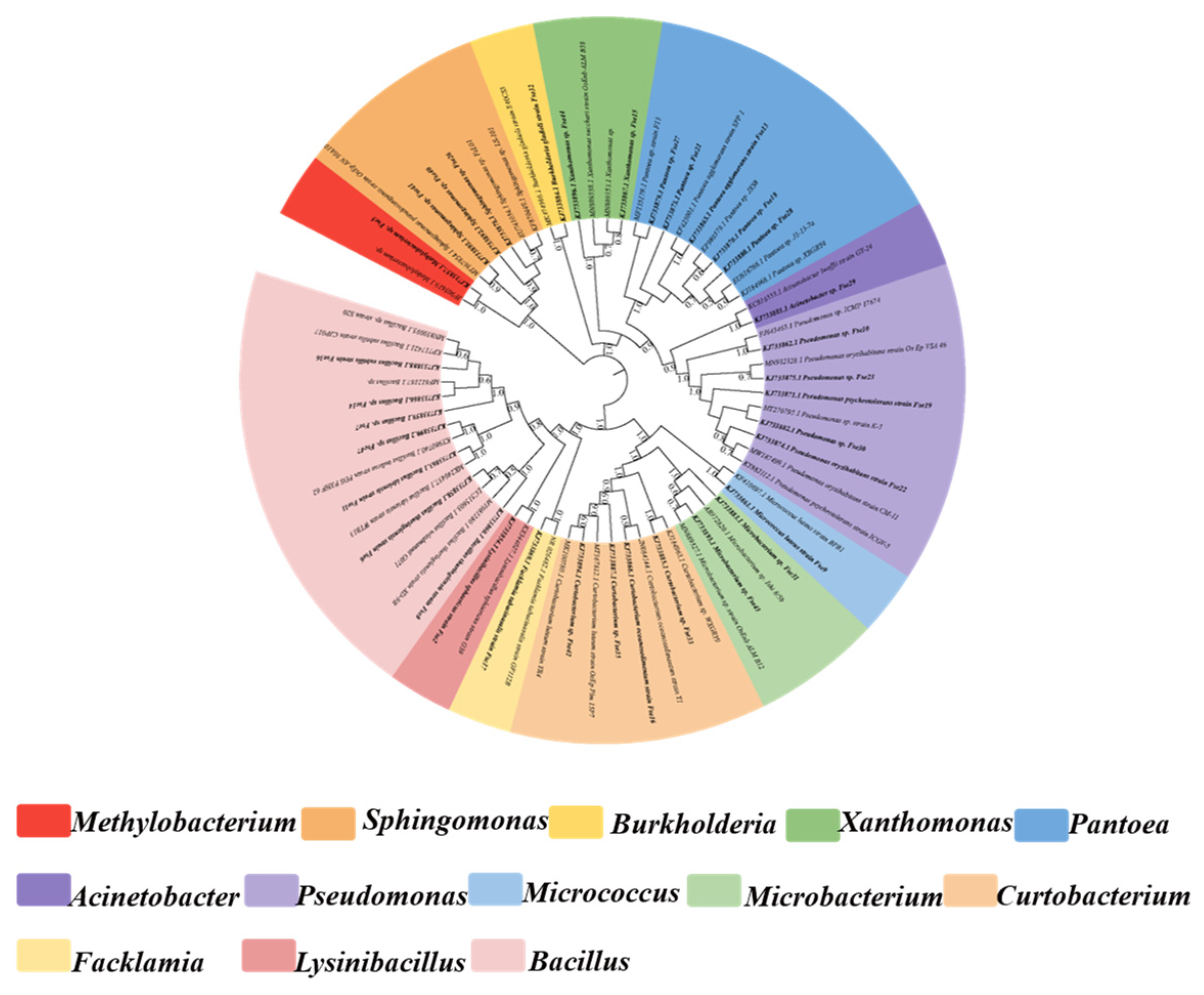
2.2. DNA Extraction, 16S rRNA Gene Amplification, Sequencing, and Strain Identification
2.3. Assay for Plant Growth-Promoting Activities
2.4. Evaluation of Plant Growth Promotion in Rice Treated with Endophytic Bacteria
2.5. Statistical Analysis
3. Results
3.1. Diversity and Community Composition of Seed Cultivable Endophytic Bacteria
3.2. Screening for PGP Traits of Isolates from Oryza Rufipogon Griff. In Vitro
3.3. Effect of Bio-Inoculum Treatment on Rice Growth-Promoting under Growth Chamber
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cordovez, V.; Dini-Andreote, F.; Carrion, V.J.; Raaijmakers, J.M. Ecology and evolution of plant microbiomes. Annu. Rev. Microbiol. 2019, 73, 69–88. [Google Scholar] [CrossRef]
- Compant, S.; Cambon, M.C.; Vacher, C.; Mitter, B.; Samad, A.; Sessitsch, A. The plant endosphere world—Bacterial life within plants. Environ. Microbiol. 2021, 23, 1812–1829. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.D.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Ali, M.; Ali, Q.; Sohail, M.A.; Ashraf, M.F.; Saleem, M.H.; Hussain, S.; Zhou, L. Diversity and taxonomic distribution of endophytic bacterial community in the rice plant and its prospective. Int. J. Mol. Sci. 2021, 22, 10165. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.N.; Yadav, N.; Dhaliwal, H.S.; Saxena, A.K. Endophytic microbes: Biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie Van Leeuwenhoek 2020, 113, 1075–1107. [Google Scholar] [CrossRef]
- Hu, J.; Wei, Z.; Weidner, S.; Friman, V.-P.; Xu, Y.-C.; Shen, Q.-R.; Jousset, A. Probiotic Pseudomonas communities enhance plant growth and nutrient assimilation via diversity-mediated ecosystem functioning. Soil. Biol. Biochem. 2017, 113, 122–129. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.d.C.; Flores, A.; Rojas-Sánchez, B.; Urtis-Flores, C.A.; Morales-Cedeño, L.R.; Valencia-Marin, M.F.; Chávez-Avila, S.; Rojas-Solis, D.; Santoyo, G. Plant growth promoting bacteria as bioinoculants: Attributes and challenges for sustainable crop improvement. Agronomy 2021, 11, 1167. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. Roles of plant endosphere microbes in agriculture—A review. J. Plant Growth Regul. 2021, 1–18. [Google Scholar] [CrossRef]
- Samreen, T.; Naveed, M.; Nazir, M.Z.; Asghar, H.N.; Khan, M.I.; Zahir, Z.A.; Kanwal, S.; Jeevan, B.; Sharma, D.; Meena, V.S.; et al. Seed associated bacterial and fungal endophytes: Diversity, life cycle, transmission, and application potential. Appl. Soil Ecol. 2021, 168, 104191. [Google Scholar] [CrossRef]
- Mitter, B.; Pfaffenbichler, N.; Flavell, R.; Compant, S.; Antonielli, L.; Petric, A.; Berninger, T.; Naveed, M.; Sheibani-Tezerji, R.; von Maltzahn, G.; et al. A new approach to modify plant microbiomes and traits by Introducing beneficial bacteria at flowering into progeny seeds. Front. Microbiol. 2017, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial endophyte colonization and distribution within plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, E.B. The seed microbiome: Origins, interactions, and impacts. Plant Soil 2018, 422, 7–34. [Google Scholar] [CrossRef]
- Rodríguez, C.E.; Mitter, B.; Barret, M.; Sessitsch, A.; Compant, S. Commentary: Seed bacterial inhabitants and their routes of colonization. Plant Soil 2017, 422, 129–134. [Google Scholar] [CrossRef]
- Johnston-Monje, D.; Lundberg, D.S.; Lazarovits, G.; Reis, V.M.; Raizada, M.N. Bacterial populations in juvenile maize rhizospheres originate from both seed and soil. Plant Soil 2016, 405, 337–355. [Google Scholar] [CrossRef] [Green Version]
- Johnston-Monje, D.; Gutiérrez, J.P.; Lopez-Lavalle, L.A.B. Seed-Transmitted Bacteria and Fungi Dominate Juvenile Plant Microbiomes. Front. Microbiol. 2021, 12, 737616. [Google Scholar] [CrossRef] [PubMed]
- Shade, A.; Jacques, M.A.; Barret, M. Ecological patterns of seed microbiome diversity, transmission, and assembly. Curr. Opin. Microbiol. 2017, 37, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.M.; Flory, E.; Koyro, H.W.; Abideen, Z.; Schikora, A.; Suarez, C.; Schnell, S.; Cardinale, M. Consistent associations with beneficial bacteria in the seed endosphere of barley (Hordeum vulgare L.). Syst. Appl. Microbiol. 2018, 41, 386–398. [Google Scholar] [CrossRef]
- Ferreira, A.; Quecine, M.C.; Lacava, P.T.; Oda, S.; Azevedo, J.L.; Araujo, W.L. Diversity of endophytic bacteria from Eucalyptus species seeds and colonization of seedlings by Pantoea agglomerans. FEMS Microbiol. Lett. 2008, 287, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Khalaf, E.M.; Raizada, M.N. Bacterial seed endophytes of domesticated cucurbits antagonize fungal and oomycete pathogens Including powdery mildew. Front. Microbiol. 2018, 9, 42. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, T.M.; Choi, B.; Kim, Y.; Kim, E. Invasive Lactuca serriola seeds contain endophytic bacteria that contribute to drought tolerance. Sci. Rep. 2021, 11, 13307. [Google Scholar] [CrossRef]
- Xu, M.; Sheng, J.; Chen, L.; Men, Y.; Gan, L.; Guo, S.; Shen, L. Bacterial community compositions of tomato (Lycopersicum esculentum Mill.) seeds and plant growth promoting activity of ACC deaminase producing Bacillus subtilis (HYT-12-1) on tomato seedlings. World J. Microb. Biot. 2014, 30, 835–845. [Google Scholar] [CrossRef]
- Xie, J.; Agrama, H.A.; Kong, D.; Zhuang, J.; Hu, B.; Wan, Y.; Yan, W. Genetic diversity associated with conservation of endangered Dongxiang wild rice (Oryza rufipogon). Genet. Resour. Crop Evol. 2010, 57, 597–609. [Google Scholar] [CrossRef]
- Qiu, B.Y.; Xiong, Y.Z.; Yu, L.Q.; Hu, B.L.; Zhang, Z.; Xie, J.K. Cluster analysis of plant morphological characters in population of Dong Xiang wild rice. Acta Agric. Jiangxi 2006, 18, 1–5. [Google Scholar] [CrossRef]
- Li, L.; Chen, H.; Mao, D. Pyramiding of rapid germination loci from Oryza Sativa cultivar ‘Xieqingzao B’ and cold tolerance loci from Dongxiang wild rice to increase climate resilience of cultivated rice. Mol. Breed. 2019, 39, 85. [Google Scholar] [CrossRef]
- Qi, W.D.; Chen, H.P.; Yang, Z.Z.; Hu, B.L.; Luo, X.D.; Ai, B.; Luo, Y.; Huang, Y.; Xie, J.K.; Zhang, F.T. Systematic characterization of long non-coding RNAs and their responses to drought stress in Dongxiang wild rice. Rice Sci. 2020, 27, 21–31. [Google Scholar] [CrossRef]
- Cui, L.L.; Li, L.; Han, X.X.; Xing, J.J.; Li, L.L.; Yang, Z.G.; Luo, X.J.; Li, D.; Xie, L.L.; Cao, M.L. Cloning and analysis of candidate RPS2 in Dongxiang wild rice. J. Hunan Agric. Univ. 2009, 35, 111–115. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, P.; Cui, F.; Zhang, F.; Luo, X.; Xie, J. Transcriptome Analysis of Salt Stress Responsiveness in the Seedlings of Dongxiang Wild Rice (Oryza rufipogon Griff.). PLoS ONE. 2016, 11, e0146242. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.X.; Peng, G.X.; Wang, E.T.; Yan, H.; Yuan, Q.H.; Zhang, W.; Lou, X.; Wu, H.; Tan, Z.Y. Diverse endophytic nitrogen-fixing bacteria isolated from wild rice Oryza rufipogon and description of Phytobacter diazotrophicus gen. nov. sp. nov. Arch. Microbiol. 2008, 189, 431–439. [Google Scholar] [CrossRef]
- Chaudhary, H.J.; Peng, G.; Hu, M.; He, Y.; Yang, L.; Luo, Y.; Tan, Z. Genetic diversity of endophytic diazotrophs of the wild rice, Oryza alta and identification of the new diazotroph, Acinetobacter oryzae sp. nov. Microb. Ecol. 2012, 63, 813–821. [Google Scholar] [CrossRef]
- Borah, M.; Das, S.; Bora, S.S.; Boro, R.C.; Barooah, M. Comparative assessment of multi-trait plant growth-promoting endophytes associated with cultivated and wild Oryza germplasm of Assam, India. Arch. Microbiol. 2021, 203, 2007–2028. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Wang, Y.; Zeng, Q.; Yan, R.; Zhang, Z.; Zhu, D. Diversity and plant growth promoting activities of the cultivable rhizobacteria of Dongxiang wild rice (Oryza rufipogon). Biodivers. Sci. 2011, 19, 476–484. [Google Scholar]
- Chen, Z.Y.; Liu, J.; Yang, X.P.; Liu, M.; Wang, Y.; Zhang, Z.B.; Zhu, D. Community composition and diversity of cultivable endophytic bacteria isolated from Dongxiang wild rice. Biodivers. Sci. 2019, 27, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, B.L.; Li, X.X.; Zhang, Z.B.; Yan, R.M.; Yang, H.L.; Zhu, D. Phylogenetic diversity of culturable endophytic fungi in Dongxiang wild rice (Oryza rufipogon Griff), detection of polyketide synthase gene and their antagonistic activity analysis. Fungal Biol. 2015, 119, 1032–1045. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Deng, Y.M.; Xiong, Y.Y.; Wang, Y.; Yan, R.M.; Zhu, D. Isolation of endophytic actinomycetes from dongxiang wild rice (oryza rufipogon) and analysis of secondary metabolite of active strain s123. Microbiol. China 2015, 42, 1662–1670. [Google Scholar]
- Zeng, Q.G.; Luo, F.; Zhang, Z.B.; Yan, R.M.; Zhu, D. Phosphate solubilizing rhizospherebacterial t21 isolated from Dongxiang wild rice species promotes cultivated rice growth. Appl. Mech. Mater. 2011, 108, 167–175. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evolut. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-De los Santos, P.; Bustillos-Cristales, R.; Caballero-Mellado, J. Burkholderia, a genus rich in plant-associated nitrogen fixers with wide environmental and geographic distribution. Appl. Environ. Microbiol. 2001, 67, 2790–2798. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.K.; Verma, M. Modified microplate method for rapid and efficient estimation of siderophore produced by bacteria. 3 Biotech 2017, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Mehta, S.; Nautiyal, C.S. An efficient method for qualitative screening of phosphate-solubilizing bacteria. Curr. Microbiol. 2001, 43, 51–56. [Google Scholar] [CrossRef]
- Ribeiro, C.M.; Cardoso, E.J.B.N. Isolation, selection and characterization of root-associated growth promoting bacteria in Brazil Pine (Araucaria angustifolia). Microbiol. Res. 2012, 167, 69–78. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plantarum. 2003, 118, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porra, R.J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth. Res. 2002, 73, 149–156. [Google Scholar] [PubMed]
- Zhang, J.; Zhang, C.W.; Yang, J.; Zhang, R.J.; Gao, J.S.; Zhao, X.; Zhao, J.J.; Zhao, D.F.; Zhang, X.X. Insights into Endophytic Bacterial Community Structures of Seeds Among Various Oryza sativa L. Rice Genotypes. J. Plant Growth Regul. 2019, 38, 93–102. [Google Scholar]
- Liu, Y.; Xu, P.; Yang, F.; Li, M.; Yan, H.; Li, N.; Zhang, X.; Wang, W. Composition and diversity of endophytic bacterial community in seeds of super hybrid rice ‘Shenliangyou 5814’ (Oryza sativa L.) and its parental lines. Plant Growth Regul. 2018, 87, 257–266. [Google Scholar] [CrossRef]
- Okunishi, S.; Sako, K.; Mano, H.; Imamura, A.; Morisaki, H. Bacterial flora of endophytes in the maturing seed of cultivated rice (Oryza sativa). Microbes Environ. 2005, 20, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Mano, H.; Tanaka, F.; Watanabe, A.; Kaga, H.; Okunishi, S.; Morisaki, H. Culturable surface and endophytic bacterial flora of the maturing seeds of rice plants (Oryza sativa) cultivated in a paddy field. Microbes Environ. 2006, 21, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Cottyn, B.; Debode, J.; Regalado, E.; Mew, T.W.; Swings, J. Phenotypic and genetic diversity of rice seed-associated bacteria and their role in pathogenicity and biological control. J. Appl. Microbiol. 2009, 107, 885–897. [Google Scholar] [CrossRef]
- Cottyn, B.; Regalado, E.; Lanoot, B.; De Cleene, M.; Mew, T.W.; Swings, J. Bacterial populations associated with rice seed in the tropical environment. Phytopathology 2001, 91, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Rosenblueth, M.; Martinez-Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, G.; Kanaujia, N.; Bafana, A. Functional and phylogenetic diversity of root-associated bacteria of Ajuga bracteosa in Kangra valley. Microbiol. Res. 2012, 167, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant-microbe interactions. Anton. Leeuw. Int. J. Gen. 2014, 106, 85–125. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.; Xu, J.; Acosta, K.; Poulev, A.; Lebeis, S.; Lam, E. Bacterial production of indole related compounds reveals their role in association between duckweeds and endophytes. Front. Chem. 2018, 6, 265. [Google Scholar] [CrossRef] [Green Version]
- Ruiza, D.; Agaras, B.; de Werrab, P.; Wall, L.G.; Valverde, C. Characterization and screening of plant probiotic traits of bacteria isolated from rice seeds cultivated in Argentina. J. Microbiol. 2011, 49, 902–912. [Google Scholar] [CrossRef]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Al-Hosni, K.; Kang, S.M.; Seo, C.W.; Lee, I.J. Indoleacetic acid production and plant growth promoting potential of bacterial endophytes isolated from rice (Oryza sativa L.) seeds. Acta Biol. Hung. 2017, 68, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-solubilizing microorganisms: Mechanism and their role in phosphate solubilization and uptake. J. Soil Sci. Plant Nut. 2020, 21, 49–68. [Google Scholar] [CrossRef]
- Varga, T.; Hixson, K.K.; Ahkami, A.H.; Sher, A.W.; Barnes, M.E.; Chu, R.K.; Battu, A.K.; Nicora, C.D.; Winkler, T.E.; Reno, L.R.; et al. Endophyte-promoted phosphorus solubilization in Populus. Front. Plant Sci. 2020, 11, 567918. [Google Scholar] [CrossRef]
- Hameed, A.; Yeh, M.W.; Hsieh, Y.T.; Chung, W.C.; Lo, C.T.; Young, L.S. Diversity and functional characterization of bacterial endophytes dwelling in various rice (Oryza sativa L.) tissues, and their seed-borne dissemination into rhizosphere under gnotobiotic P-stress. Plant Soil 2015, 394, 177–197. [Google Scholar] [CrossRef]
- Nag, P.; Shriti, S.; Das, S. Microbiological strategies for enhancing biological nitrogen fixation in nonlegumes. J. Appl. Microbiol. 2020, 129, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, S.H.; Gururani, M.A.; Chun, S.-C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Walitang, D.I.; Kim, K.; Madhaiyan, M.; Kim, Y.K.; Kang, Y.; Sa, T. Characterizing endophytic competence and plant growth promotion of bacterial endophytes inhabiting the seed endosphere of rice. BMC Microbiol. 2017, 17, 209. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-producing rhizobacteria as a promising tool for empowering plants to cope with iron limitation in saline soils: A review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Yang, J.H.; Liu, H.X.; Zhu, G.M.; Pan, Y.L.; Xu, L.P.; Guo, J.H. Diversity analysis of antagonists from rice-associated bacteria and their application in biocontrol of rice diseases. J. Appl. Microbiol. 2008, 104, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Loaces, I.; Ferrando, L.; Scavino, A.F. Dynamics, diversity and function of endophytic siderophore-producing bacteria in rice. Microb. Ecol. 2011, 61, 606–618. [Google Scholar] [CrossRef]
- Sultana, S.; Alam, S.; Karim, M.M. Screening of siderophore-producing salt-tolerant rhizobacteria suitable for supporting plant growth in saline soils with iron limitation. J. Agric. Food Res. 2021, 4, 100150. [Google Scholar] [CrossRef]
- Ghavami, N.; Alikhani, H.A.; Pourbabaei, A.A.; Besharati, H. Effects of two new siderophore-producing rhizobacteria on growth and iron content of maize and canola plants. J. Plant. Nutr. 2017, 40, 736–746. [Google Scholar] [CrossRef]
- Murali, M.; Gowtham, H.G.; Singh, S.B.; Shilpa, N.; Aiyaz, M.; Niranjana, S.R.; Amruthesh, K.N. Bio-prospecting of ACC deaminase producing rhizobacteria towards sustainable agriculture: A special emphasis on abiotic stress in plants. Appl. Soil Ecol. 2021, 168, 104142. [Google Scholar] [CrossRef]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013, 366, 93–105. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Hernandez, A.G.; Glick, B.R.; Rossi, M.J. The extreme plant-growth-promoting properties of Pantoea phytobeneficialis MSR2 revealed by functional and genomic analysis. Environ. Microbiol. 2020, 22, 1341–1355. [Google Scholar] [CrossRef] [PubMed]
- Silambarasan, S.; Logeswari, P.; Valentine, A.; Cornejo, P. Role of Curtobacterium herbarum strain CAH5 on aluminum bioaccumulation and enhancement of Lactuca sativa growth under aluminum and drought stresses. Ecotox. Environ. Saf. 2019, 183, 109573. [Google Scholar] [CrossRef]
- Bourles, A.; Guentas, L.; Chalkiadakis, E.; Majorel, C.; Juillot, F.; Cavaloc, Y.; Burtet-Sarramegna, V.; Medevielle, V.; Jourand, P.; Amir, H. New Caledonian ultramafic conditions structure the features of Curtobacterium citreum strains that play a role in plant adaptation. Can. J. Microbiol. 2019, 65, 880–894. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolation Medium | Numbers of Isolates | Genera |
|---|---|---|
| NA | 14 | Microbacterium, Bacillus, Pseudomonas, Pantoea, Xanthomonas, Facklamia, Acinetobacter, Burkholderia |
| DNA | 13 | Methylobacterium, Pantoea, Curtobacterium, Microbacterium, Micrococcus, Bacillus, Xanthomonas, Sphingomonas |
| BPA | 6 | Bacillus, Microbacterium, Sphingomonas, Curtobacterium, Pseudomonas |
| R2A | 6 | Bacillus, Microbacterium, Pseudomonas, Pantoea, Curtobacterium, Lysinibacillus |
| TSA | 8 | Bacillus, Microbacterium, Curtobacterium, Pantoea, Pseudomonas |
| Strain Code | GenBank Number | Closest Relative from GenBank | Max. Identity | ACC Deaminase | N-Fixation | IAA (μg mL−1) | Siderophore | Phosphate Solubilisation (μg mL−1) |
|---|---|---|---|---|---|---|---|---|
| Fse5 | KJ733857 | Methylobacterium sp. | 99 | 6.13 ± 0.27 | − | 33.72 ± 1.28 | + | 576.34 ± 24.63 |
| Fse26 | KJ733878 | Sphingomonas sp. | 99 | - | + | 22.86 ± 1.66 | + | 27.70 ± 6.15 |
| Fse40 | KJ733892 | Sphingomonas sp. | 99 | - | + | 22.45 ± 2.73 | − | - |
| Fse41 | KJ733893 | Sphingomonas sp. | 99 | - | + | 16.75 ± 1.76 | − | - |
| Fse32 | KJ733884 | Burkholderia gladioli | 100 | 16.91 ± 1.38 | − | 18.56 ± 0.69 | + | 339.07 ± 10.28 |
| Fse10 | KJ733862 | Pseudomonas sp. | 99 | 8.22 ± 0.68 | + | 36.76 ± 2.87 | + | 187.85 ± 12.88 |
| Fse23 | KJ733875 | Pseudomonas sp. | 99 | - | + | 59.63 ± 6.18 | − | 85.29 ± 8.02 |
| Fse30 | KJ733882 | Pseudomonas sp. | 100 | - | + | 45.08 ± 1.81 | − | 297.09 ± 32.93 |
| Fse19 | KJ733871 | Pseudomonas psychrotolerans | 100 | - | + | 18.87 ± 1.28 | + | 135.77 ± 3.86 |
| Fse22 | KJ733874 | Pseudomonas oryzihabitans | 100 | 7.61 ± 0.72 | + | 19.06 ± 4.46 | − | 30.47 ± 2.40 |
| Fse24 | KJ733876 | Pseudomonas oryzihabitans | 100 | - | + | 33.02 ± 1.38 | − | 120.46 ± 11.67 |
| Fse15 | KJ733867 | Xanthomonas sp. | 100 | - | − | 15.09 ± 1.22 | − | 78.73 ± 5.15 |
| Fse20 | KJ733872 | Xanthomonas sp. | 100 | - | − | 20.89 ± 1.53 | − | - |
| Fse37 | KJ733889 | Xanthomonas sacchari | 100 | - | − | 14.01 ± 0.35 | − | 207.06 ± 9.16 |
| Fse44 | KJ733896 | Xanthomonas sacchari | 100 | - | − | 54.29 ± 2.41 | − | 16.25 ± 4.15 |
| Fse21 | KJ733873 | Pantoea sp. | 100 | - | + | 72.97 ± 1.87 | − | 384.56 ± 29.09 |
| Fse18 | KJ733870 | Pantoea sp. | 99 | - | + | 89.70 ± 5.17 | − | 13.13 ± 2.95 |
| Fse25 | KJ733877 | Pantoea ananatis | 99 | 10.21 ± 0.94 | + | 47.25 ± 0.41 | − | 500.32 ± 56.05 |
| Fse27 | KJ733879 | Pantoea sp. | 99 | 15.41 ± 0.85 | + | 81.24 ± 2.01 | − | 280.90 ± 7.77 |
| Fse28 | KJ733880 | Pantoea sp. | 99 | 19.51 ± 1.64 | + | 90.11 ± 5.90 | + | 302.54 ± 8.80 |
| Fse13 | KJ733865 | Pantoea agglomerans | 99 | 13.23 ± 0.97 | + | 48.14 ± 2.73 | + | 193.74 ± 2.55 |
| Fse29 | KJ733881 | Acinetobacter sp. | 99 | - | − | 27.18 ± 0.98 | + | 209.67 ± 23.52 |
| Fse1 | KJ733853 | Bacillus subtilis | 99 | 12.24 ± 1.02 | − | 51.41 ± 2.04 | − | - |
| Fse36 | KJ733888 | Bacillus subtilis | 99 | - | − | 29.28 ± 4.17 | + | 43.83 ± 3.99 |
| Fse7 | KJ733859 | Bacillus velezensis | 100 | - | − | 6.02 ± 0.35 | − | 18.46 ± 3.85 |
| Fse14 | KJ733866 | Bacillus sp. | 100 | 8.12 ± 0.56 | − | 30.45 ± 1.83 | + | 102.29 ± 8.28 |
| Fse47 | KJ733899 | Bacillus sp. | 100 | - | − | - | + | 8.62 ± 1.45 |
| Fse6 | KJ733858 | Bacillus thuringiensis | 100 | - | − | 27.11 ± 1.35 | + | 92.75 ± 12.27 |
| Fse8 | KJ733860 | Bacillus thuringiensis | 100 | - | − | 3.37 ± 0.25 | − | 5.88 ± 0.69 |
| Fse11 | KJ733863 | Bacillus idriensis | 100 | 12.25 ± 1.25 | − | - | − | - |
| Fse2 | KJ733854 | Lysinibacillus sphaericus | 100 | 5.77 ± 0.44 | − | 8.66 ± 0.18 | + | - |
| Fse17 | KJ733869 | Facklamia tabacinasalis | 100 | - | − | 5.53 ± 0.21 | + | 61.53 ± 1.34 |
| Fse3 | KJ733855 | Microbacterium sp. | 100 | - | + | 7.88 ± 0.14 | − | 4.63 ± 0.79 |
| Fse4 | KJ733856 | Microbacterium sp. | 100 | - | + | 34.35 ± 2.35 | − | 29.09 ± 0.46 |
| Fse12 | KJ733864 | Microbacterium sp. | 100 | - | + | 6.16 ± 0.95 | − | 509.96 ± 39.41 |
| Fse31 | KJ733883 | Microbacterium sp. | 100 | - | + | 8.45 ± 0.46 | − | 151.88 ± 8.61 |
| Fse34 | KJ733886 | Microbacterium sp. | 100 | - | + | 54.52 ± 1.95 | − | 24.75 ± 4.31 |
| Fse43 | KJ733895 | Microbacterium sp. | 100 | - | + | 28.83 ± 3.65 | − | - |
| Fse46 | KJ733898 | Microbacterium sp. | 100 | - | − | 15.70 ± 2.78 | − | 60.49 ± 7.94 |
| Fse9 | KJ733861 | Micrococcus luteus | 100 | 2.12 ± 0.18 | − | 18.77 ± 0.69 | + | - |
| Fse33 | KJ733885 | Curtobacterium sp. | 100 | - | + | 8.58 ± 1.23 | − | 14.69 ± 1.91 |
| Fse35 | KJ733887 | Curtobacterium sp. | 99 | 20.72 ± 1.89 | + | 86.72 ± 5.40 | + | 112.35 ± 3.11 |
| Fse38 | KJ733890 | Curtobacterium sp. | 99 | 14.32 ± 0.23 | + | 72.94 ± 7.46 | − | - |
| Fse39 | KJ733891 | Curtobacterium sp. | 99 | - | + | 42.23 ± 5.49 | − | - |
| Fse42 | KJ733894 | Curtobacterium sp. | 100 | 12.12 ± 0.18 | + | 25.42 ± 2.26 | − | 6.90 ± 1.66 |
| Fse45 | KJ733897 | Curtobacterium sp. | 100 | - | + | 18.35 ± 0.33 | − | 84.77 ± 11.66 |
| Fse16 | KJ733868 | Curtobacterium oceanosedimentum | 100 | 15.74 ± 0.26 | + | 9.09 ± 0.34 | + | 191.90 ± 29.50 |
| Treatments | 3 Days | 5 Days | 7 Days | ||
|---|---|---|---|---|---|
| Germination Rate (%) | Germination Rate (%) | Germination Rate (%) | Sprout Length (cm) | Root Length (cm) | |
| Control | 47.57 ± 0.66 c | 70.13 ± 1.98 c | 94.62 ± 1.52 b | 1.40 ± 0.18 a | 1.28 ± 0.15 a |
| Fse28 | 56.44 ± 1.01 a | 85.52 ± 2.11 a | 96.67 ± 0.49 a | 2.19 ± 0.28 b | 2.53 ± 0.49 b |
| Fse35 | 50.86 ± 1.14 b | 80.63 ± 2.09 b | 95.33 ± 0.54 b | 2.05 ± 0.29 b | 2.07 ± 0.24 b |
| Days | Treatment | Shoot Length (cm) | Root Length (cm) | Dry Weight (mg) | Chlorophyll Content (mg g−1) |
|---|---|---|---|---|---|
| 10 d | control | 14.35 ± 0.22 a | 6.60 ± 0.28 a | 18.80 ± 2.29 a | 0.57 ± 0.15 a |
| Fse28 | 18.84 ± 0.69 bc | 8.92 ± 1.46 a | 28.34 ± 1.17 b | 1.82 ± 0.15 bc | |
| Fse32 | 15.89 ± 0.45 b | 7.35 ± 1.49 a | 25.86 ± 2.47 b | 1.11 ± 0.11 b | |
| 20 d | control | 26.04 ± 0.40 a | 9.06 ± 0.69 a | 70.80 ± 9.53 a | 1.68 ± 0.14 a |
| Fse28 | 32.22 ± 1.00 bc | 12.60 ± 1.12 b | 168.87 ± 32.21 b | 2.74 ± 0.37 b | |
| Fse32 | 29.55 ± 1.44 b | 11.63 ± 1.42 b | 118.79 ± 15.23 b | 2.41 ± 0.12 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Liu, T.; Zhang, X.; Xie, J.; Wang, Y.; Yan, R.; Jiang, Y.; Zhu, D. Cultivable Endophytic Bacteria in Seeds of Dongxiang Wild Rice and Their Role in Plant-Growth Promotion. Diversity 2021, 13, 665. https://doi.org/10.3390/d13120665
Zhang Z, Liu T, Zhang X, Xie J, Wang Y, Yan R, Jiang Y, Zhu D. Cultivable Endophytic Bacteria in Seeds of Dongxiang Wild Rice and Their Role in Plant-Growth Promotion. Diversity. 2021; 13(12):665. https://doi.org/10.3390/d13120665
Chicago/Turabian StyleZhang, Zhibin, Tingting Liu, Xiao Zhang, Jing Xie, Ya Wang, Riming Yan, Yumei Jiang, and Du Zhu. 2021. "Cultivable Endophytic Bacteria in Seeds of Dongxiang Wild Rice and Their Role in Plant-Growth Promotion" Diversity 13, no. 12: 665. https://doi.org/10.3390/d13120665
APA StyleZhang, Z., Liu, T., Zhang, X., Xie, J., Wang, Y., Yan, R., Jiang, Y., & Zhu, D. (2021). Cultivable Endophytic Bacteria in Seeds of Dongxiang Wild Rice and Their Role in Plant-Growth Promotion. Diversity, 13(12), 665. https://doi.org/10.3390/d13120665