Demographic Traits Variation in a Pyrenean Newt (Calotriton asper) among Lacustrine and Stream Populations


Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Begon, M.; Harper, J.L.; Townsend, C.R. Ecology. Individuals, Populations and Communities; Blackwell Scientific Publications: London, UK, 1986. [Google Scholar]
- Sorci, G.; Clobert, J.; Belichon, S. Phenotypic plasticity of growth and survival in the common lizard Lacerta vivipara. J. Anim. Ecol. 1996, 65, 781–790. [Google Scholar] [CrossRef]
- Wilbur, H.M.; Collins, J.P. Ecological aspects of amphibian metamorphosis: Nonnormal distributions of competitive ability reflect selection for facultative metamorphosis. Science 1973, 182, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Stearns, S.C. Life history evolution: Successes, limitations, and prospects. Naturwissenschaften 2000, 87, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Adolph, S.C.; Porter, W.P. Growth, seasonality, and lizard life histories: Age and size at maturity. Oikos 1996, 77, 267–278. [Google Scholar] [CrossRef]
- Oromi, N.; Valbuena-Ureña, E.; Soler-Membrives, A.; Amat, F.; Camarasa, S.; Carranza, S.; Sanuy, D.; Denoël, M. Genetic structure of lake and stream populations in a Pyrenean amphibian (Calotriton asper) reveals evolutionary significant units associated with paedomorphosis. J. Zool. Syst. Evol. Res. 2018, 57, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics; Longman: Harlow, UK, 1996. [Google Scholar]
- Colleoni, E.; Denoël, M.; Padoa-Schioppa, E.; Scali, S.; Ficetola, G.F. Rensch’s rule and sexual dimorphism in salamanders: Patterns and potential processes. J. Zool. 2014, 293, 143–151. [Google Scholar] [CrossRef]
- Selander, R.K. Sexual dimorphism and differential niche utilization in birds. Condor 1966, 68, 113–151. [Google Scholar] [CrossRef]
- Dobson, F.; Wigginton, J.D. Environmental influences on the sexual dimorphism in body size of western bobcats. Oecologia 1996, 108, 610–616. [Google Scholar] [CrossRef]
- Frafjord, K.; Stevy, I. The red fox in Norway: Morphological adaptation or random variation in size? Z. Säugetierkunde 1998, 63, 16–25. [Google Scholar]
- Miaud, C.; Merilä, J. Local adaptation or environmental induction? Causes of population differentiation in alpine amphibians. Biota 2001, 2, 31–50. [Google Scholar]
- Clergue-Gazeau, M.; Martinez-Rica, J.P. Les différents biotopes de l’urodèle pyrénéen, Euproctus asper. Bull. Soc. Hist. Nat. Toulouse 1978, 114, 461–471. [Google Scholar]
- Bruce, R.C. Life histories. In Reproductive Biology and Phylogeny of Urodela; Sever, D.M., Ed.; Science Publisher: Enfield, CT, USA, 2003; Volume 1, pp. 477–526. [Google Scholar]
- Serra-Cobo, J.; Uiblein, F.; Martinez-Rica, J.P. Variation in sexual dimorphism between two populations of the Pyrenean salamander Euproctus asper from ecologically different mountain sites. Belg. J. Zool. 2000, 130, 39–45. [Google Scholar]
- Camarasa, S.; Amat, F.; Sanuy, D.; Oromi, N. The occurrence of facultative paedomorphosis in a lacustrine population of the Pyrenean newt (Calotriton asper) morphology and age structure. J. Nat. Hist. 2018, 52, 33–34. [Google Scholar] [CrossRef]
- Oromi, N.; Amat, F.; Sanuy, D.; Carranza, S. Life history trait differences between a lake and a stream-dwelling population of the Pyrenean brook newt (Calotriton asper). Amphib. Reptil. 2014, 35, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Montori, A. Skeletochronological results in the pyrenean newt Euproctus asper (Dugès, 1852) from one prepyrenean population. Ann. Sci. Nat. Zool. Paris 1990, 13, 209–211. [Google Scholar]
- Guerrero-Campo, J. Variación altitudinal y dimorfismo sexual en la coloración y biometría del tritón pirenaico (Euproctus asper, amphibia-salamandridae). Lucas Mallada 1995, 7, 213–225. [Google Scholar]
- Miaud, C.; Guillaume, O. Variation in age, body size and growth among surface and cave-dwelling populations of the Pyrenean newt, Euproctus asper (Amphibia; Urodela). Herpetologica 2005, 61, 241–249. [Google Scholar] [CrossRef]
- Valbuena-Ureña, E.; Oromi, N.; Soler-Membrives, A.; Carranza, S.; Amat, F.; Camarasa, S.; Denoël, M.; Guillaume, O.; Sanuy, D.; Loyau, A.; et al. Jailed in the mountains: Genetic diversity and structure of an endemic newt species across the Pyrenees. PLoS ONE 2018, 13, e0200214. [Google Scholar] [CrossRef] [Green Version]
- Puig, M.A.; Ventura, M. Arroyos y ríos de Alta Montaña. Boletín Life+limnopirineus 2016, 3, 2–4. [Google Scholar]
- Catalan, J.; Camarero, L.; Felip, M.; Pla, S.; Ventura, M.; Buchaca, T.; Bartumeus, F.; De Mendoza, G.; Miró, A.; Casamayor, E.O.; et al. High mountain lakes: Extreme hábitats and witnesses of environmental chances. Limnetica 2006, 25, 551–584. [Google Scholar]
- Miaud, C. La squelettochronologie chez les Triturus (Amphibiens, Urodeles) à partir d’une étude de T. alpestris, T. helveticus et T. cristatus du sud-est de la France. In Tissus Durs et Age Individuel des Vertébrés; Baglinière, J.L., Castanet, J., Conand, F., Meunier, F.J., Eds.; ORSTOM-INRA: Paris, France, 1991. [Google Scholar]
- Amat, F.; Oromi, N.; Sanuy, D. Body size, population size, and age structure of adult Palmate newts (Lissotriton helveticus) in Pyrenean Lakes. J. Herpetol. 2010, 44, 313–319. [Google Scholar] [CrossRef]
- Morrison, C.; Hero, J.M. Altitudinal variation in growth and development rates of Litoria chloris and Litoria pearsoniana tadpoles in southeast Queensland, Australia. J. Herpetol. 2003, 37, 59–64. [Google Scholar] [CrossRef] [Green Version]
- JMP, Version Pro 14; SAS Institute Inc.: Cary, NC, USA, 1989–2019.
- Elser, J.J.; Wu, C.; González, A.L.; Shain, D.H.; Smith, H.J.; Sommaruga, R.; Williamson, C.E.; Brahney, J.; Hotaling, S.; Vanderwall, J.; et al. Key rules of life and the fading cryosphere: Impacts in alpine lakes and streams. Glob. Chang. Biol. 2020, 26, 6644–6656. [Google Scholar] [CrossRef] [PubMed]
- Roff, D. Trade-offs between growth and reproduction: An analysis of the quantitative genetic evidence. J. Evol. Biol. 2000, 13, 434–445. [Google Scholar] [CrossRef]
- Lucati, F.; Poignet, M.; Miró, A.; Trochet, A.; Aubret, F.; Barthe, L.; Bertrand, R.; Buchaca, T.; Calvez, O.; Caneret, J.; et al. Multiple glacial refugia and contemporary dispersal shape the genetic structure of an endemic amphibian from the Pyrenees. Mol. Ecol. 2020, 29, 2904–2921. [Google Scholar] [CrossRef] [PubMed]
- Jalut, G.; Montserrat-Marti, J.; Fontugne, M.; Delibrias, G.; Vilaplana, J.M.; Julia, R. Glacial to interglacial vegetation changes in the northern and southern Pyrénées: Deglaciation, vegetation cover and chronology. Quat. Sci. Rev. 1992, 11, 449–480. [Google Scholar] [CrossRef]
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.G.; Cosson, J.F. Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, H.H. Evolution of facultative paedomorphosis in salamanders. Quat. Rev. Biol. 1994, 69, 205–221. [Google Scholar] [CrossRef]
- Denoël, M. Terrestrial versus aquatic foraging in juvenile Alpine newts (Triturus alpestris). Écoscience 2004, 11, 404–409. [Google Scholar] [CrossRef]
- Winandy, L.; Darnet, E.; Denoël, M. Amphibians forgo aquatic life in response to alien fish introduction. Anim. Behav. 2015, 109, 209–216. [Google Scholar] [CrossRef]
- Montori, A.; Richter-Boix, A.; Franch, M.; Santos, X.; Garriga, N.; Llorente, G.A. Natural fluctuations in a stream dwelling newt as a result of extreme rainfall: A 21-year survey of a Calotriton asper population. Basic Appl. Herpetol. 2012, 26, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Montori, A.; Llorente, G.A.; Richter-Boix, A. Habitat features affecting the small-scale distribution and longitudinal migration patterns of Calotriton asper in a Pre-Pyrenean population. Amphib. Reptil. 2008, 29, 371–381. [Google Scholar] [CrossRef]
- Clergue-Gazeau, M.; Beetschen, J.C. Contribution à l’étude de la reproduction de l’Urodèle Euproctus asper (Dugès) dans les Pyrénées centrales et Ariègeoises. Ann. Limnol. Int. J. Limnol. 1966, 2, 217–226. [Google Scholar] [CrossRef]
- Miaud, C.; Guyétant, R.; Faber, H. Age, size and growth of the alpine newt, Triturus alpestris (Urodela, Salamandridae), at high altitude and a review of life-history trait variation throughout its range. Herpetologica 2000, 56, 135–144. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Habitat | Locality | Latitude | Longitude | Altitude | n |
|---|---|---|---|---|---|
| Stream | Peramola | 42°4′17.17″ N | 1°16′30.98″ E | 592 | 28 |
| San Juan de la Peña | 42°30′26.94″ N | 0°40′0.16″ W | 1215 | 38 | |
| Sarvisé | 42°34′51.90″ N | 0°4′17.70″ W | 1222 | 38 | |
| Oto | 42°35′47.47″ N | 0°7′42.56″ W | 916 | 44 | |
| Benasc | 42°38′2.23″ N | 0°34′25.21″ E | 1600 | 37 | |
| Barranc de Acherito | 42°52′27.81″ N | 0°43′18.64″ W | 1525 | 47 | |
| Lake | Espeluciecha | 42°47′18.06″ N | 0°25′47.93″ W | 1955 | 52 |
| Acherito | 42°52′45.52″ N | 0°42′27.48″ W | 1872 | 44 | |
| Perramó | 42°38′19.29″ N | 0°30′1.14″ E | 2254 | 71 |
| Age (Years) | SVL (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Habitat | Locality | Sex | n | Minimum | Maximum | Median | Minimum | Maximum | Mean ± ES |
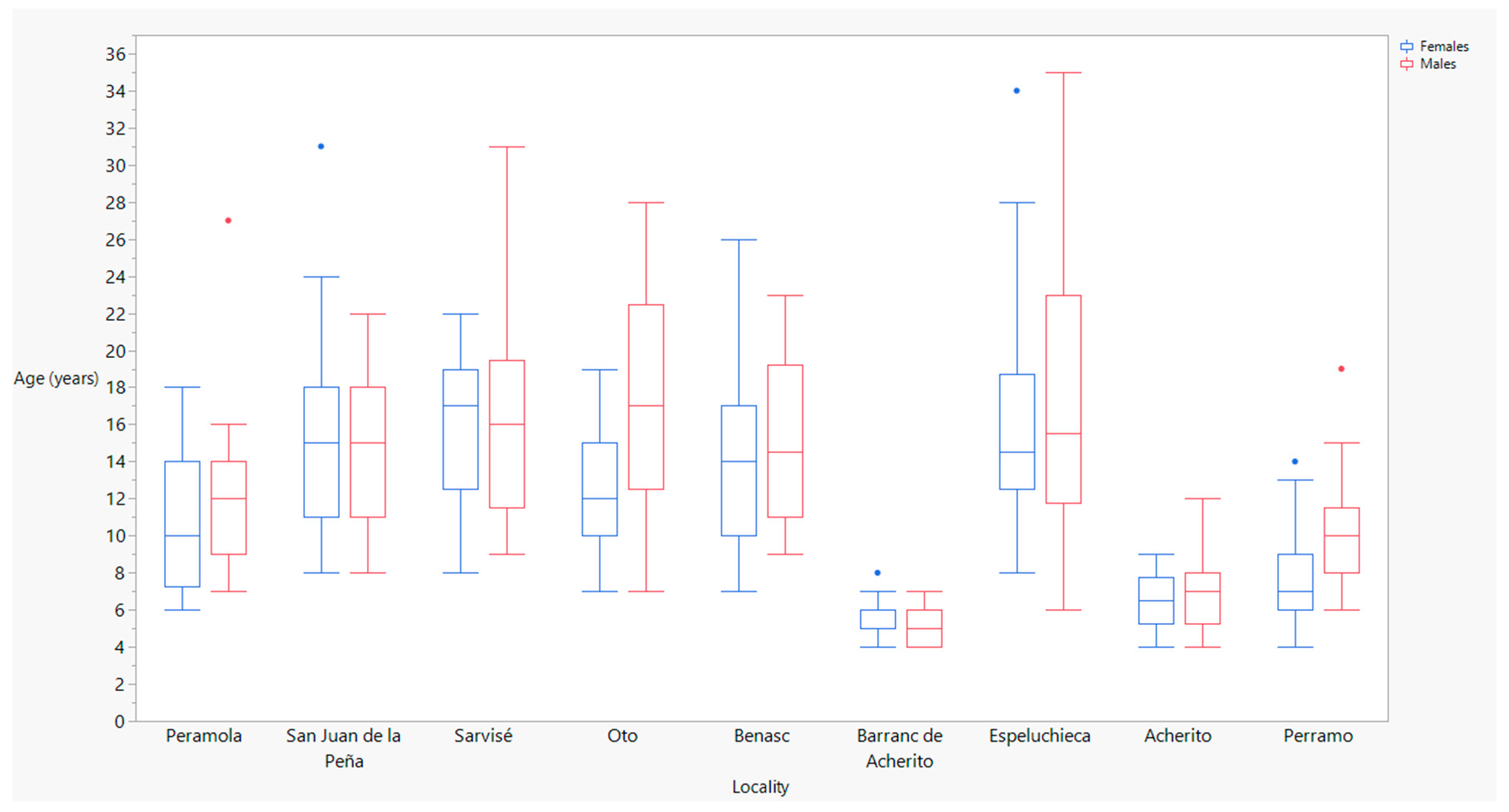
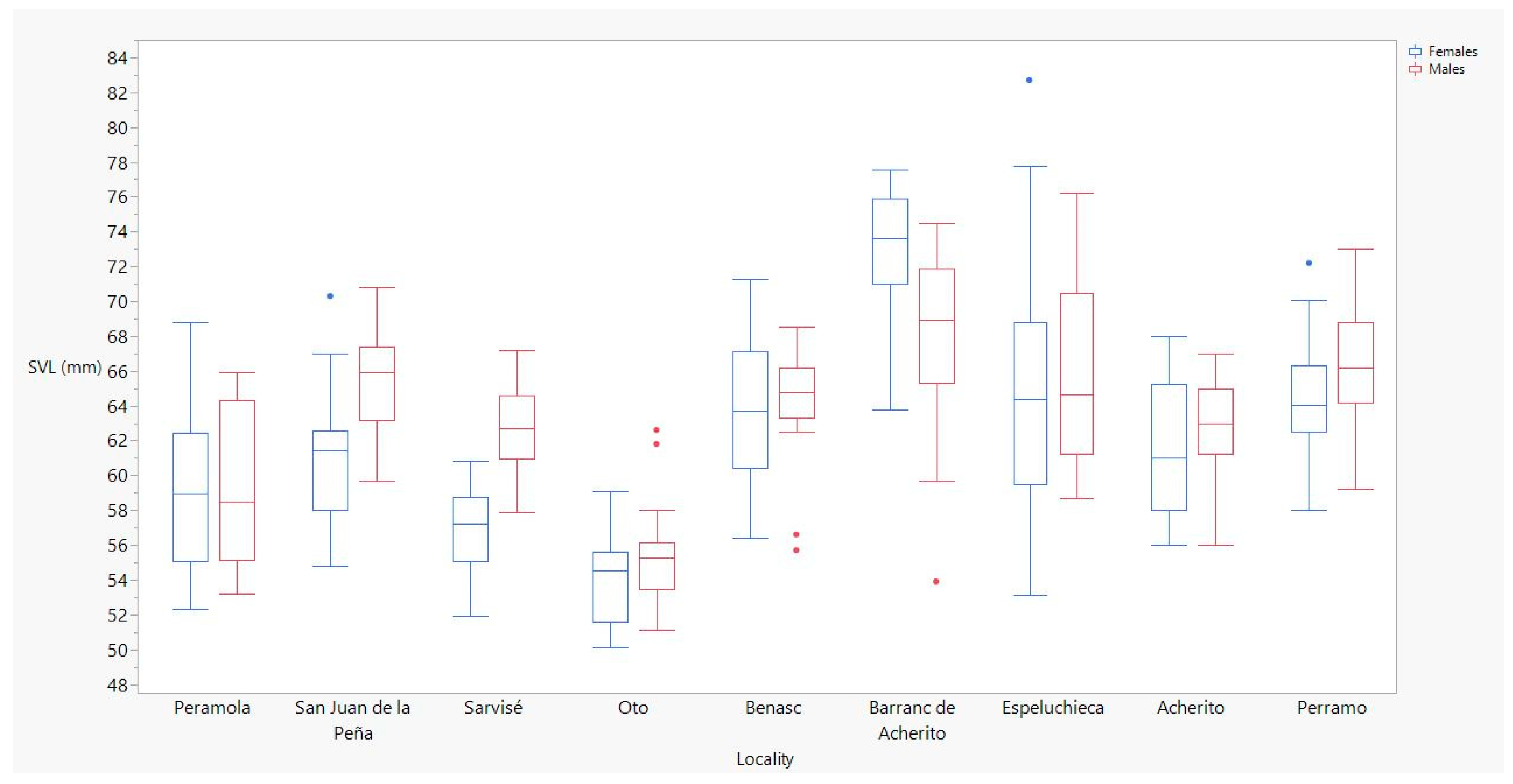
| Stream | Peramola | Females | 16 | 6 | 18 | 10.0 | 52.3 | 68.8 | 59.0 ± 1.1 |
| Males | 12 | 7 | 27 | 12.0 | 53.2 | 65.9 | 58.5 ± 1.3 | ||
| San Juan de la Peña | Females | 19 | 8 | 31 | 15.0 | 54.8 | 70.3 | 61.4 ± 0.9 | |
| Males | 19 | 8 | 22 | 15.0 | 59.7 | 70.8 | 65.9 ± 0.6 | ||
| Sarvisé | Females | 17 | 8 | 22 | 17.0 | 51.9 | 58.6 | 57.2 ± 0.6 | |
| Males | 21 | 9 | 31 | 16.0 | 57.9 | 67.2 | 62.7 ± 0.5 | ||
| Oto | Females | 23 | 7 | 19 | 12.0 | 50.1 | 59.1 | 54.5 ± 0.6 | |
| Males | 21 | 7 | 28 | 17.0 | 51.1 | 62.6 | 55.3 ± 0.6 | ||
| Benasc | Females | 19 | 7 | 26 | 14.0 | 56.4 | 71.3 | 63.7 ± 0.9 | |
| Males | 18 | 9 | 23 | 14.5 | 55.7 | 68.5 | 64.8 ± 0.8 | ||
| Barranc de Acherito | Females | 20 | 4 | 8 | 5.0 | 63.8 | 77.6 | 73.6 ± 0.8 | |
| Males | 27 | 4 | 7 | 5.0 | 53.9 | 74.5 | 68.9 ± 0.9 | ||
| Lake | Espeluchieca | Females | 26 | 8 | 34 | 14.5 | 53.1 | 82.7 | 64.4 ± 1.4 |
| Males | 26 | 6 | 35 | 15.5 | 58.7 | 76.2 | 64.7 ± 1.0 | ||
| Acherito | Females | 12 | 4 | 9 | 6.5 | 56.0 | 68.0 | 61.0 ± 1.1 | |
| Males | 32 | 4 | 12 | 7.0 | 56.0 | 67.0 | 63.0 ± 0.5 | ||
| Perramó | Females | 34 | 4 | 14 | 7.0 | 58.0 | 72.2 | 64.1 ± 0.6 | |
| Males | 37 | 6 | 19 | 10.0 | 59.2 | 73.0 | 66.2 ± 0.5 | ||
| Age Structure by Sex and Population | |||
|---|---|---|---|
| Factor | Degrees of freedom | F | Probability > F |
| Sex | 1 | 8.6671 | 0.0034 * |
| Locality | 8 | 74.9023 | <0.0001 * |
| Locality × sex | 8 | 2.4706 | 0.0128 * |
| SVL by sex and population | |||
| Factor | Degrees of freedom | F | Probability > F |
| Locality | 8 | 62.6399 | <0.0001 * |
| Sex | 1 | 10.2254 | 0.0015 * |
| Locality × sex | 8 | 6.3897 | <0.0001* |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camarasa, S.; Oromi, N.; Sanuy, D.; Amat, F. Demographic Traits Variation in a Pyrenean Newt (Calotriton asper) among Lacustrine and Stream Populations. Diversity 2020, 12, 471. https://doi.org/10.3390/d12120471
Camarasa S, Oromi N, Sanuy D, Amat F. Demographic Traits Variation in a Pyrenean Newt (Calotriton asper) among Lacustrine and Stream Populations. Diversity. 2020; 12(12):471. https://doi.org/10.3390/d12120471
Chicago/Turabian StyleCamarasa, Sebastià, Neus Oromi, Delfí Sanuy, and Fèlix Amat. 2020. "Demographic Traits Variation in a Pyrenean Newt (Calotriton asper) among Lacustrine and Stream Populations" Diversity 12, no. 12: 471. https://doi.org/10.3390/d12120471