Factors Influencing Epiphytic Lichen Species Distribution in a Managed Mediterranean Pinus nigra Arnold Forest

 ,
,
Abstract
1. Introduction
2. Materials and Methods
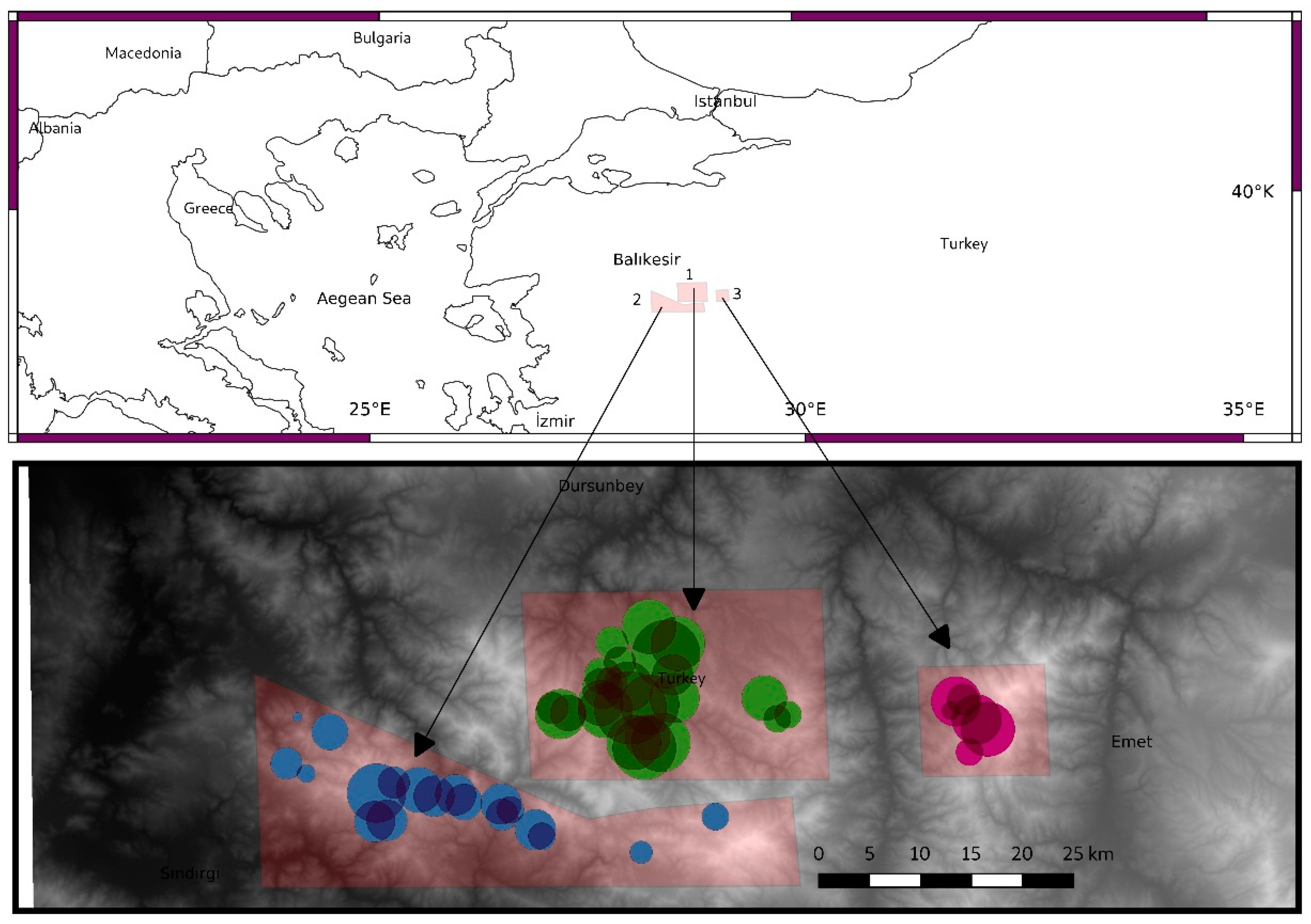
2.1. Study Area
2.2. Data
2.2.1. Species Data
2.2.2. Environment Data
2.3. Statistical Analyses
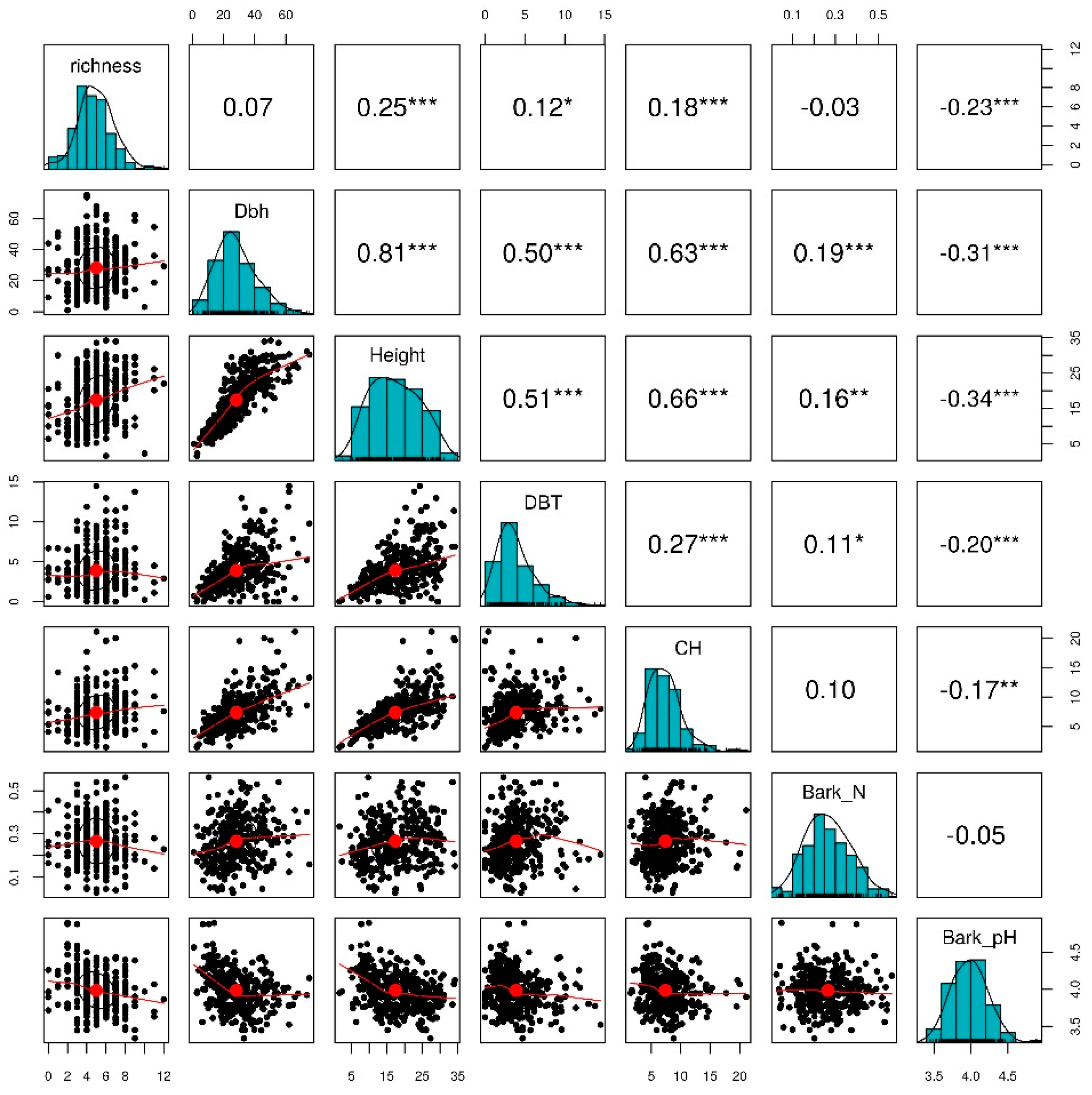
3. Results
3.1. Epiphytic Lichens in Black Pine Forests
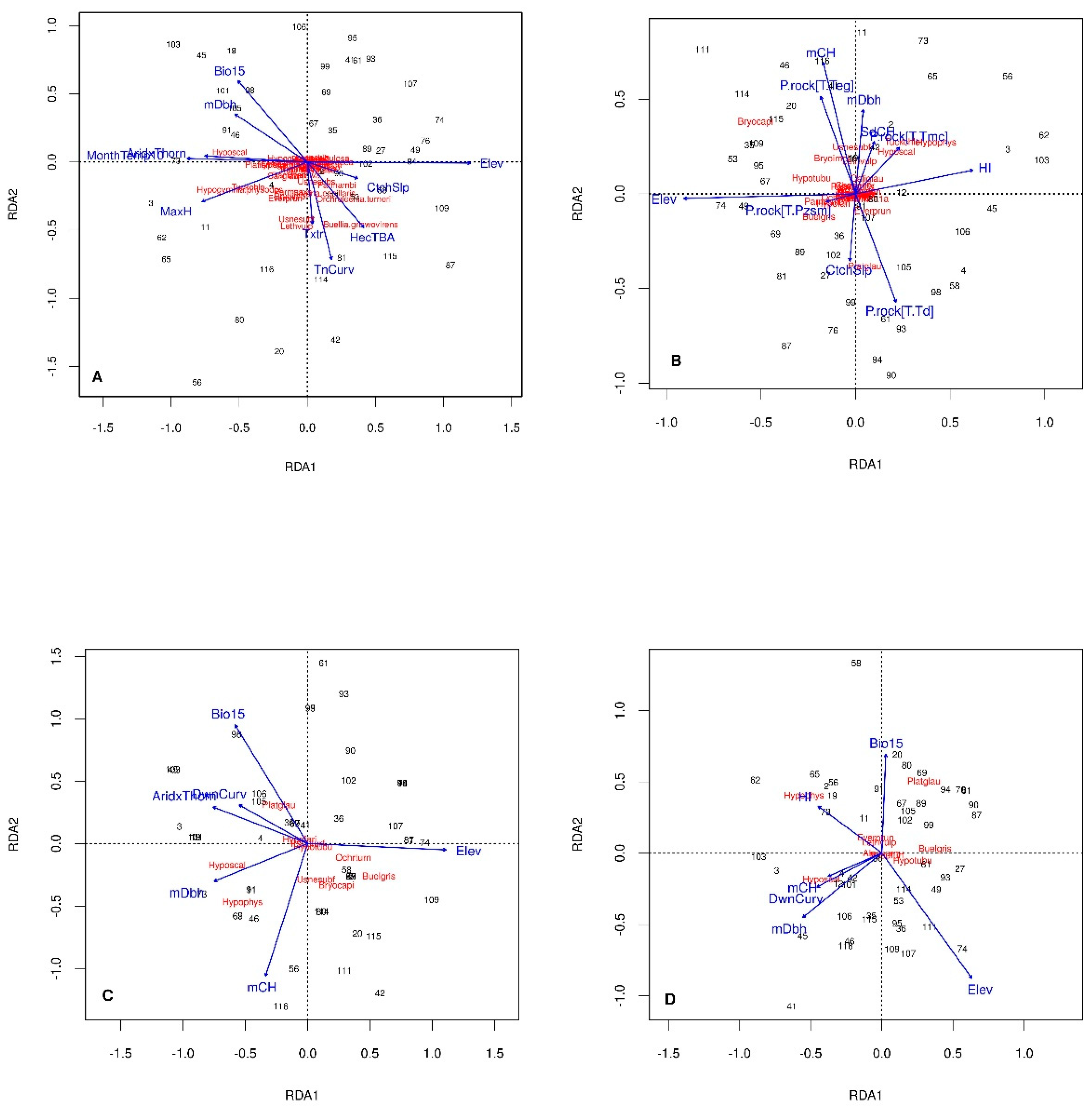
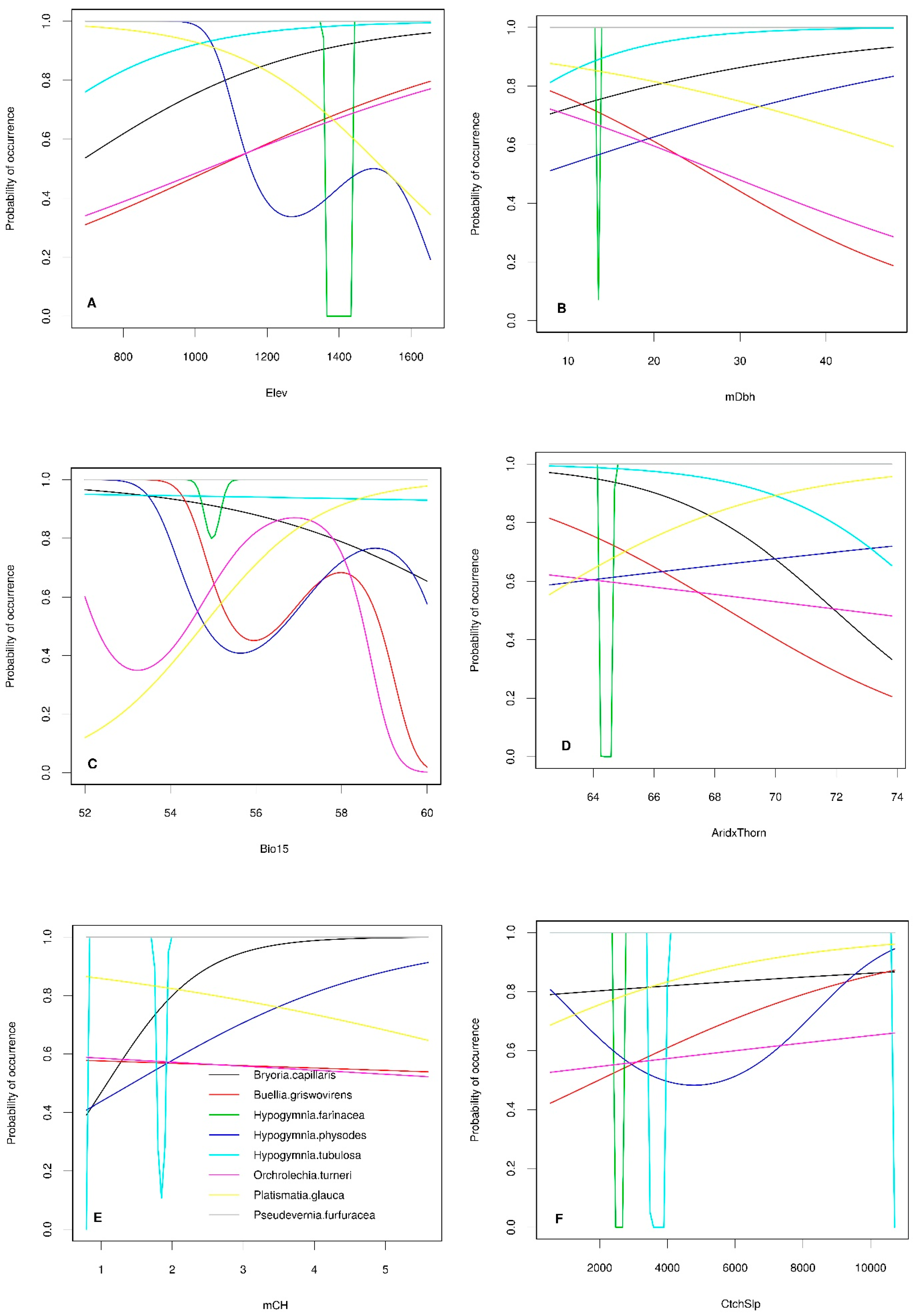
3.2. Stand-Level Lichen Species Composition
3.3. Tree-Level Lichen Species Richness
4. Discussion
4.1. Epiphytic Lichen Species in Black Pine Forests
4.2. Stand-Level Lichen Species Composition
4.3. Tree-Level Lichen Richness
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Used Variables in Varpart after Multicorelation
| Variable Group | Variables | Description |
| Bioclimatic (n = 6) | Bio4 (Temperature seasonality (SD x 100)), Bio15 (Precipitation seasonality, coefficient of variation), AridxThorn (index of the degree of water deficit below water need), MonthTemp10 (count of the number of months with mean temperature greater than 10 °C), TopoWet (SAGA-GIS topographic wetness index), Terrough (terrain roughness index), | [64,65] |
| Topographic (n = 20) | Elev (Elevation), East (Eastness), Nrth (Northness), MBI (Mass balance index), PrI (Protection index), MRRTF (Multiresolution index of the ridge top flatness), slp (Slope), GnCurv (General curvature), TnCurv (Tangential curvature), MnCurv (Minimal curvature), ToCurv (Total curvature), FLCurv (Flow line curvature), Txtr (Texture), Cnvx (Convexity), TPI (Topographic position index), DwnCurv (Downslope curvature), ValDp (Valley depth), HI (Hill index), HSI (Hillslope index), VTR (Vector terrain ruggedness), ValDp1 (Valley depth 1), CtchSlp (Catchment slope), SlpLen (Slope length), | Derived from DEM with SAGA GIS terrain analyses |
| Stand structure | mAge (mean value of increment cores collected from 4–7 trees in stand), minAge (minimum stand age), sdAge (Standart Deviation of stand age), CaCl (Canopy closure degree of trees in the stand) hecTBA (Total basal area of black pine ha−2), mDbh (Mean diameter breast height of black pine trees in the stand), sdDbh (Standard deviation of black pine breast height diameter values), maxH (Maximum height in stand), mCH (Mean crown height of black pine trees in the stand), sdCH (Standard deviation of crown height of black pine trees in the stand), mDBT (Mean dry branch thickness of black pine trees in the stand), | Calculated from study field data |
| Tree | Dbh (Diameter at breast height of tree), Bark pH (Tree bark pH), BarkN (Tree bark Nitrogen), Tree height, CH (Crown height of black pine tree), DBT (Dry branch thickness of black pine tree), |
Appendix B. Lichen Species According to Region and Type in Black Pine Forests
| Species | Region | Type | ||
| Dursunbey (n = 26) | Simav (n = 6) | Sindirgi (n = 16) | ||
| Bryoria capillaris (Ach.) Brodo & D.Hawksw. | 22 | 6 | 11 | fruticose |
| Bryoria fuscescens var. fuscescens (Gyeln.) Brodo & D.Hawksw. | 1 | 0 | 1 | fruticose |
| Bryoria implexa (Hoffm.) Brodo & D.Hawksw. | 6 | 3 | 0 | fruticose |
| Buellia erubescens Arnold | 0 | 0 | 1 | crustose |
| Buellia griseovirens (Turner & Borrer ex Sm.) Almb. | 13 | 6 | 8 | crustose |
| Calicium glaucellum Ach. | 3 | 0 | 0 | crustose |
| Chaenotheca chrysocephala (Turner ex Ach.) Th.Fr. | 1 | 1 | 0 | crustose |
| Cladonia chlorophaea (Flörke ex Sommerf.) Spreng. | 2 | 0 | 1 | crustose |
| Evernia divaricata (L.) Ach. | 3 | 0 | 1 | fruticose |
| Evernia prunastri (L.) Ach. | 8 | 1 | 6 | fruticose |
| Hypocenomyyce scalaris (Ach. ex Lilj.) M.Choisy | 14 | 1 | 3 | squamulose |
| Hypogymnia farinacea Zopf | 26 | 5 | 16 | foliose |
| Hypogymnia physodes (L.) Nyl. | 20 | 4 | 7 | foliose |
| Hypogymnia tubulosa (Schaer.) Hav. | 24 | 6 | 15 | foliose |
| Lecanora chlarotera Nyl. | 1 | 0 | 0 | crustose |
| Lecidella elaeochroma (Ach.) M. Choisy f. elaeochroma | 0 | 0 | 1 | crustose |
| Letharia vulpina (L.) Hue | 7 | 3 | 0 | fruticose |
| Ochrolechia turneri (Sm.) Hasselrot | 19 | 2 | 6 | fruticose |
| Parmelia saxatilis (L.) Ach. | 10 | 1 | 4 | foliose |
| Parmeliopsis ambigua (Wulfen) Nyl. | 2 | 2 | 1 | foliose |
| Pertusaria albescens (Huds.) M. Choisy & Werner var. albescens | 4 | 0 | 0 | crustose |
| Pertusaria hemisphaerica (Flörke) Erichsen | 1 | 1 | 0 | crustose |
| Platismatia glauca (L.) W.L. Culb. & C.F. Culb. | 23 | 1 | 14 | foliose |
| Pseudevernia furfuracea var. furfuracea (L.) Zopf | 26 | 6 | 16 | fruticose |
| Ramalina farinacea (L.) Ach. | 2 | 0 | 0 | fruticose |
| Trapeliopsis flexuosa (Fr.) Coppins & P.James | 1 | 0 | 0 | crustose |
| Tuckermanopsis chlorophylla (Willd.) Hale | 12 | 0 | 3 | foliose |
| Usnea florida (L.) Weber ex F.H. Wigg. | 0 | 1 | 0 | fruticose |
| Usnea glabrescens (Nyl. ex Vain.) Vain. | 1 | 0 | 3 | fruticose |
| Usnea hirta (L.) Weber ex F.H. Wigg. | 1 | 0 | 0 | fruticose |
| Usnea scabrata Nyl. | 7 | 1 | 3 | fruticose |
| Usnea subfloridana Stirt. | 10 | 4 | 3 | fruticose |
| Usnea subscabrosa Nyl. ex Motyka | 2 | 1 | 2 | fruticose |
References
- Numa, C.; Valderrábano, M.; Alcázar, E.; Barrios, V.; Bourlon, S.; Bugalho, M.; Buse, J.; Vecchia, I.D.; Garnier, E.; Kabouya-Loucif, I.; et al. Biodiversity conservation and protected areas in the Mediterranean region. In State of Mediterranean Forests 2018; Food and Agriculture Organization of the United Nations: Rome, Italy; Marseille, France, 2018; pp. 147–162. ISBN 978-92-5-131047-2. [Google Scholar]
- Barbero, M.; Loisel, R.; Quezel, P.; Richardson, D.M.; Romane, F. Pines or the Mediterranean Basin. In Ecology and Biogeography of Pinus, 1st ed.; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 153–170. ISBN 0521551765. [Google Scholar]
- Bussotti, F. Pinus nigra Arnold. In Pines of Silvicultural Importance; CABI Publishing: Wallingford, UK, 2002; pp. 266–286. [Google Scholar]
- Akkemik, U.; Yılmaz, H.; Oral, D.; Kaya, A. Pinus. In Türkiye’nin Doğal Gymnospermleri (Açık Tohumlular), 1st ed.; Yaltırık, F., Akkemik, U., Eds.; Turkish Ministry of Enviroment and Forestry Press: Ankara, Turkey, 2011. [Google Scholar]
- Isajev, V.; Fady, B.; Semerci, H.; Andonovski, V. EUFORGEN Technical Guidelines for Genetic Conservation and Use for European Black Pine (Pinus nigra); International Plant Genetic Resources Institute: Rome, Italy, 2004; 6p. [Google Scholar]
- Mayer, H.; Aksoy, H. Türkiye Ormanları; Orman Bakanlığı Batı Karadeniz Ormancılık Araştırma Enstitüsü Müdürlüğü Yayınları: Bolu, Turkey, 1998; ISBN 975-7829-56-0.
- Kalıpsız, A. Türkiye’de Karaçam Meşcerelerinin Tabii Büyümesi ve Verim Kudreti Üzerine Araştırmalar; OGM Publications Sequence Nu: İstanbul, Turkey, 1963. [Google Scholar]
- Kavgacı, A.; Sevgi, O.; Tecimen, H.B.; Yılmaz, O.Y.; Serdar Carus, S.; Türker Dündar, T. Classification and Ordination of Pinus nigra Dominated Forests at Alaçam Mountains (Nw Anatolia-Turkey). Eurasian J. For. Sci. 2013, 1, 38–50. [Google Scholar]
- Gerson, U.; Seaward, M.R.D. Lichen–Invertebrate Associations. In Lichen Ecology; Seaward, M.R.D., Ed.; Academic Press: London, UK, 1977; pp. 69–120. ISBN 0-12-634350-0. [Google Scholar]
- Peck, J.E.; McCune, B. Remmant Trees and Canopy Lichen Communities in Western Oregon: A Retrospective Approach. Ecol. Appl. 1997, 7, 1181–1187. [Google Scholar] [CrossRef]
- Huneck, S. The Significance of Lichens and Their Metabolites. Naturwissenschaften 1999, 86, 559–570. [Google Scholar] [CrossRef]
- Sillett, S.C.; McCune, B.; Peck, J.E.; Rambo, T.R.; Ruchty, A. Dispersal Limitations of Epiphytic Lichens Result in Species Dependent on Old-growth Forests. Ecol. Appl. 2000, 10, 789–799. [Google Scholar] [CrossRef]
- Price, K.; Hochachka, G. Epiphytic Lichen Abundance: Effects of Stand Age and Composition in Cosatal British Columbia. Ecol. Appl. 2001, 11, 904–913. [Google Scholar] [CrossRef]
- Nash, T.H. Nitrogen, its metabolism and potential contribution to ecosystems. In Lichen Biology, 1st ed.; Nash, T.H., Ed.; Cambridge University Press: New York, NY, USA, 1996. [Google Scholar]
- Belinchón, R.; Martínez, I.; Escudero, A.; Aragón, G.; Valladares, F. Edge Effects on Epiphytic Communities in a Mediterranean Quercus pyrenaica Forest. J. Veg. Sci. 2007, 18, 81–90. [Google Scholar] [CrossRef]
- Nöske, N.M.; Hilt, N.; Werner, F.A.; Brehm, G.; Fiedler, K.; Sipman, H.J.M.; Gradstein, S.R. Disturbance effects on diversity of epiphytes and moths in a montane forest in Ecuador. Basic Appl. Ecol. 2008, 9, 4–12. [Google Scholar] [CrossRef]
- Aragón, G.; Martínez, I.; Izquierdo, P.; Relinchón, R.; Escudero, A. Effects of forest management on epiphytic lichen diversity in Mediterranean forests. Appl. Veg. Sci. 2010, 13, 183–194. [Google Scholar] [CrossRef]
- Pinho, P.; Correia, O.; Lecoq, M.; Munzi, S.; Vasconcelos, S.; Gonçalves, P.; Rebelo, R.; Antunes, C.; Silva, P.; Freitas, C.; et al. Evaluating green infrastructure in urban environments using a multi-taxa and functional diversity approach. Environ. Res. 2016, 147, 601–610. [Google Scholar] [CrossRef]
- Nascimbene, J.; Benesperi, R.; Bruniaiti, G.; Catalano, I.; Vedove, M.D.; Grillo, M.; Isocrono, D.; Matteucci, E.; Potenza, G.; Puntillo, D.; et al. Patterns and drivers of ß-diversity and similarity of Lobaria pulmonaria communities in Italian. J. Ecol. 2013, 101, 493–505. [Google Scholar] [CrossRef]
- Cardós, J.L.H.; Martínez, I.; Calvo, V.; Aragón, G. Epiphyte communities in Mediterranean fragmented forests: Importance of the fragment size and the surrounding matrix. Landsc. Ecol. 2016, 31, 1975–1995. [Google Scholar] [CrossRef]
- Pinho, P.; Dias, T.; Cruz, C.; Sim Tang, Y.; Sutton, M.A.; Martins-Loução, M.A.; Branquinho, C. Using lichen functional diversity to assess the effects of atmospheric ammonia in Mediterranean woodlands. J. Appl. Ecol. 2011, 48, 1107–1116. [Google Scholar] [CrossRef]
- Scott, M.G.; Hutchinson, T.C. Experiments and Observations on Epiphytic Lichens as Early Warning Sentinels of Forest Decline. In Biologic Markers of Air Pollution Stress and Damage in Forests; The National Academies Press: Washington, DC, USA, 1989; pp. 205–215. [Google Scholar]
- Jackson, L.; Ford, J.; Schwatzman, D. Collection and Chemical Analysis of Lichens for Biomonitoring. In Lichens as Bioindicators of Air Quality; USDA Forest Service General Technical Report RM-224; Huckaby, L.S., Ed.; USDA Forest Service: Fort Collins, CO, USA, 1993; pp. 96–115. [Google Scholar]
- Aamlid, D.; Torseth, K.; Venn, K.; Stuanes, A.O.; Solberg, S.; Hylen, G.; Christophersen, N.; Framstad, E. Changes of Forest Health in Norwegian Boreal Forest During 15 Years. For. Ecol. Manag. 2000, 127, 103–118. [Google Scholar] [CrossRef]
- McCune, B.; Rogers, P.; Ruchty, A.; Ryan, B. Lichen Communities for Forest Health Monitoring in Colorado, USA; A Report to the USDA Forest Service; Forest Health Monitoring National Office, Southern Research Station, Research Triangle Park: Washington, DC, USA, 1998; 29p.
- McCune, B. Lichen Communities as Indicators of Forest Health. Bryologist 2000, 103, 353–356. [Google Scholar] [CrossRef]
- Garty, J. Biomonitoring Atmospheric Heavy Metals with Lichens: Theory and Application. CRC Crit. Rev. Plant Sci. 2001, 20, 309–371. [Google Scholar] [CrossRef]
- Asta, J.; Erhardt, W.; Ferretti, M.; Fornasier, F.; Kirschbaum, U.; Nimis, P.L.; Purvis, O.W.; Pirintsos, S.; Scheidegger, C.; Haluwyn, C.V.; et al. Mapping Lichen Diversity as in Indicator of Environmental Quality. In Monitoring with Lichens-Monitoring Lichen; Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; NATO Science Series (Series IV: Earth and Environmental Sciences); Springer: Dordrecht, The Netherlands, 2002; Volume 7, pp. 273–279. [Google Scholar]
- Barkmann, J.J. Phytosociology and Ecology of Cryptogamic Epiphytes; Van Gorcum: Assen, The Netherlands, 1958. [Google Scholar]
- Kuusinen, M. Epiphyte Flora and Diversity on Basal Trunks of Six Old-Growth Forest Tree Species in Southern and Middle Boreal Finland. Lichenologist 1996, 28, 443–463. [Google Scholar] [CrossRef]
- Dietrch, M.; Scheidegger, C. Frequency, Diversity and Ecological Strategies of Epiphytic Lichens in the Swiss Central Plateau and Pre-alps. Lichenologist 1997, 29, 237–258. [Google Scholar] [CrossRef]
- Neitlich, P.N.; McCune, B. Hostpots of Epiphytic Lichen Diversity in Two Young Managed Forests. Conserv. Biol. 1997, 11, 172–182. [Google Scholar] [CrossRef]
- Giordani, P. Variables influencing the distribution of epiphytic lichens in heterogeneous areas: A case study for Liguria, NW Italy. J. Veg. Sci. 2006, 17, 195–206. [Google Scholar] [CrossRef]
- Fritz, Ö.; Niklasson, M.; Churski, M. Tree age is a factor for the conservation of epiphytic lichens and bryophytes in beech forests. Appl. Veg. Sci. 2008, 12, 93–106. [Google Scholar] [CrossRef]
- Lie, M.H.; Arup, U.; Grytnes, J.A.; Ohlson, M. The importance of host tree age, size and growth rate as determinants of epiphytic lichen diversity in boreal spruce forests. Biodivers. Conserv. 2009, 18, 3579–3596. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Motta, R.; Nimis, P.L. Influence of tree age, tree size and crown structure on lichen communities in mature Alpine spruce forests. Biodivers. Conserv. 2009, 18, 1509–1522. [Google Scholar] [CrossRef]
- Ellis, C.J. Lichen epiphyte diversity: A species, community and trait-based review. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 131–152. [Google Scholar] [CrossRef]
- Ódor, P.; Király, I.; Tinya, F.; Bortignon, F.; Nascimbene, J. Patterns and drivers of species composition of epiphytic bryophytes and lichens in managed temperate forests. For. Ecol. Manag. 2013, 306, 256–265. [Google Scholar] [CrossRef]
- Bäcklund, S.; Jönsson, M.; Strengbom, J.; Frisch, A.; Thor, G. A Pine is a Pine and a Spruce is a Spruce—The Effect of Tree Species and Stand Age on Epiphytic Lichen Communities. PLoS ONE 2016, 11, e0147004. [Google Scholar] [CrossRef]
- Hörsch, B. Modelling the spatial distribution of montane and subalpie forests in the central Alps using digital elevation models. Ecol. Model. 2003, 168, 267–282. [Google Scholar] [CrossRef]
- Hoersch, B.; Braun, G.; Schmidt, U. Relation between landform and vegetation in alpine regions of Wallis, Switzerland. A multiscale remote sensing and GIS approach. Comput. Environ. Urban 2002, 26, 113–139. [Google Scholar] [CrossRef]
- Saco, P.M.; Willgoose, G.R.; Hancock, G.R. Eco-geomorphology of banded vegetation patterns in arid and semi-arid regions. Hydrol. Earth Syst. Sci. 2007, 11, 1717–1730. [Google Scholar] [CrossRef]
- Engelhardt, B.M.; Weisberg, P.J.; Chambers, J.C. Influences of watershed geomorphology on extent and composition of riparian vegetation. J. Veg. Sci. 2012, 23, 127–139. [Google Scholar] [CrossRef]
- Jimenez-Alfaro, B.; Marceno, C.; Bueno, A.; Gavilán, R.; Obeso, J.R. Biogeographic deconstruction of alpine plant communities along altitudinal and topographic gradients. J. Veg. Sci. 2014, 25, 160–171. [Google Scholar] [CrossRef]
- Ardelean, I.V.; Keller, C.; Scheidegger, C. Effects of Management on Lichen Species Richness, Ecological Traits and Community Structure in the Rodnei Mountains National Park (Romania). PLoS ONE 2015, 10, e0145808. [Google Scholar] [CrossRef]
- Burgaz, A.R.; Fuertes, E.; Escudero, A. Ecology of cryptogamic epiphytes and their communities in deciduous forests in mediterranean Spain. Vegetatio 1994, 112, 73–86. [Google Scholar] [CrossRef]
- Fuertes, E.; Burgaz, A.R.; Escudero, A. Pre-climax epiphyte communities of bryophytes and lichens in Mediterranean forests from the Central Plateau (Spain). Vegetatio 1996, 123, 139–151. [Google Scholar] [CrossRef]
- Aragón, G.; Belinchón, R.; Martínez, I.; Prieto, M. Estimating epiphytic lichen richness by single families in Mediterranean forests. For. Ecol. Manag. 2013, 310, 187–193. [Google Scholar] [CrossRef]
- Christensen, S.N. The epiphytic lichen flora of Platanus orientalis stands in Greece. Willdenowia 2014, 44, 209–227. [Google Scholar] [CrossRef]
- Pirintsos, S.A.; Diamantopoulos, J.; Stamou, G.P. Analysis of the Distribution of Epiphytic Lichens within Homogeneous Fagus sylvatica Stands along an Altitudinal Gradient (Mount Olympos, Greece). Vegetatio 1995, 116, 33–40. [Google Scholar]
- Belinchón, R.; Martinez, I.; Otalora, M.A.G.; Aragón, G.; Dimas, J.; Escudero, A. Fragment quality and matrix affect epiphytic performance in a mediterranean forest landscape. Am. J. Bot. 2009, 96, 1974–1982. [Google Scholar] [CrossRef] [PubMed]
- Pirintsos, S.A.; Diamantopoulos, J.; Stamou, G.P. Analysis of the Vertical Distribution of Epiphytic Lichens on Pinus nigra (MountOlympos, Greece) along an Altitudinal Gradient. Vegetatio 1993, 109, 63–70. [Google Scholar] [CrossRef]
- Christensen, S.N. Lichen associated with Pinus nigra on Mt. Trapezitsa, Epirus, NW Greece. Acta Bot. Fenn. 1994, 150, 11–20. [Google Scholar]
- Guvenc, S.; Oran, S.; Ozturk, Ş. The Epiphytic Lichens on Anatolian Black Pine [Pinus nigra Arnd. subsp. pallasiana (Lamb.) Holmboe in Mt. Uludag (Bursa–Turkey). JABS 2009, 3, 157–161. [Google Scholar]
- Sevgi, O.; Yılmaz, O.; Carus, S.; Dündar, T.; Kavgacı, A.; Tecimen, H.B. Alaçam Dağlarındaki Karaçam Ormanlarının Yükseltiye Göre Beslenme-Büyüme Modelleri ve Odununun Teknolojik Özellikleri; TOVAG 104 O 551 Project Report; TÜBİTAK: Ankara, Turkey, 2010; p. 478.
- Pamay, B. Yaş Sınıfları Amenajman Metodunun Türkiye Ormanlarına Uygulanması İmkanları ve Karşılaşılan Güçlükler. İstanbul Üniv. Orman Fakültesi Derg. 1968, 18, 23–41. [Google Scholar]
- Atay, İ. Silvikültür II; İstanbul Üniversitesi: İstanbul, Turkey, 1990; 242p, ISBN 975-404-174-1. [Google Scholar]
- Purvis, O.W.; Coppins, B.J.; Hawksworth, D.L.; James, P.W.; Moore, D.M. The Lichen Flora of Great Britain and Ireland; Natural History Museum Publications: London, UK, 1992. [Google Scholar]
- Wirth, V. Die Flechten Baden-Württem Bergs 2 Vols; Eugen Ulmer: Stuttgart, Germany, 1995. [Google Scholar]
- Brodo, I.M.; Sharnoff, S.D.; Sharnoff, S. Lichens of North America; Yale University Press: New Haven/London, UK, 2001. [Google Scholar]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: Functions for Spatial Thinning of Species Occurrence Records for Use in Ecological Models. R Package Version 0.1.0. 2014. Available online: https://CRAN.R-project.org/package=spThin (accessed on 25 December 2018).
- Moning, C.; Werth, S.; Dziock, F.; Bässler, C.; Bradtka, J.; Hothorn, T.; Müller, J. Lichen diversity in temperate montane forests is influenced by forest structure more than climate. For. Ecol. Manag. 2009, 258, 745–751. [Google Scholar] [CrossRef]
- Goral, F.; Schellenberg, J. Function for Community Data and Ordinations. R Package Version 0.3.3. 2018. Available online: https://cran.r-project.org/web/packages/goeveg/index.html (accessed on 25 December 2018).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for automated geoscientific analyses (SAGA) v. 2.1.4. 475. Geosci. Model Dev. 2015, 8, 1991–2007. [Google Scholar] [CrossRef]
- Kudrnovsky, H. Geographic Resources Analysis Support System (GRASS) Software. 2018. Available online: https://grass.osgeo.org/grass7/manuals/addons/i.segment.stats.html (accessed on 25 December 2018).
- Gauslaa, Y. The Lobarion and Epiphytic Community of Ancient Forests Threatened by Acid Rain. Lichenologist 1995, 27, 59–76. [Google Scholar]
- Poikolainen, J. Mosses, Epiphytic Lichens and Tree Barks as Biomonitors for Air Pollutants—Specifically for Heavy Metals in Regional Surveys; The Finnish Forest Research Institute, Oulu University Press: Oulu, Finland, 2004.
- Bremmer, J.M.; Mulvaney, C.S. Nitrogen–Total. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Soil Science Society of America Inc.: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-4. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 25 December 2019).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 119, 1674–1684. [Google Scholar] [CrossRef]
- Revelle, W. psych: Procedures for Personality and Psychological Research; Northwestern University: Evanston, IL, USA, 2018. [Google Scholar]
- R Development Core Team. A language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Development Core Team: Vienna, Australia, 2018; Available online: http://www.R-project.org (accessed on 10 June 2018).
- Nascimbene, J.; Marini, L.; Nimis, P.L. Influence of tree species on epiphytic macrolichens in temperate mixed forests of northern Italy. Can. J. For. Res. 2009, 39, 785–791. [Google Scholar] [CrossRef]
- Çobanoğlu, G.; Sevgi, E.; Sevgi, O. Lichen Mycota along Uludağ Fir (Abies bornmuelleriana MATTF); University of Craiova Seria: Craiova, Romania, 2008; Volume 13, pp. 15–19. [Google Scholar]
- Nascimbene, J.; Marini, L.; Ódor, P. Drivers of lichen species richness at multiple spatial scales in temperate forests. Plant Ecol. Div. 2012, 5, 355–363. [Google Scholar] [CrossRef]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers. Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef]
- Kreylinga, J.; Wiesenbergb, G.L.B.; Thiela, D.; Wohlfarta, C.; Huberc, G.; Walterd, J.; Jentschd, A.; Konnertc, M.; Beierkuhnleina, C. Cold hardiness of Pinus nigra Arnold as influenced by geographic origin, warming, and extreme summer drought. Environ. Exp. Bot. 2012, 78, 99–108. [Google Scholar] [CrossRef]
- Enescu, C.M.; de Rigo, D.; Caudullo, G.; Mauri, A.; Houston Durrant, T. Pinus nigra in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; European Commission: Luxembourg, 2016; Volume 6, pp. 126–127. [Google Scholar]
- Cobanoglu, G.; Sevgi, O. Analysis of the Distribution of Epiphytic Lichens on Cedrus libani in Elmali Research Forest (Antalya, Turkey). J. Environ. Biol. 2009, 30, 205–212. [Google Scholar]
- Sevgi, O.; Tecimen, H.B.; Carus, S.; Akburak, S.; Çaksir, E. Some bark characteristics of Black Pine (Arnold.) and their variation throughout the tree height Pinus nigra. J. Environ. Biol. 2016, 37, 1347–1354. [Google Scholar]
- Marmor, L.; Randlane, T.E. Effects of road traffic on bark pH and epiphytic lichens in Tallinn. Folia Cryptog. Est. 2007, 43, 23–37. [Google Scholar]
- Selva, S.B. Lichen diversity and stand continuity in the northern hardwoods and spruce-fir forests in northern New England and western New Brunswick. Bryologist 1994, 97, 424–429. [Google Scholar] [CrossRef]
- Svoboda, D.; Peksa, O.; Vesela, J. Epiphytic lichen diversity in central European oak forests: Assessment of the effects of natural environmental factors and human influences. Environ. Pollut. 2010, 158, 812–819. [Google Scholar] [CrossRef]
- Rubio-Salcedo, M.; Psormas, A.; Prieto, M.; Zimmermann, N.E.; Martinez, I. Case study of the implications of climate change for lichen diversity and distribution. Biodivers. Conserv. 2017, 26, 1121–1141. [Google Scholar] [CrossRef]
- Giorani, P.; Incerti, G. The influence of climate on the distribution of lichens: A case study in a borderline area (Liguria, NW Italy). Plant Ecol. 2008, 195, 257–272. [Google Scholar] [CrossRef]
- Jovan, S.; McCune, B. Regional Variation in Epiphytic Macrolichen Communities in Northern and Central California Forests. Bryologist 2004, 107, 328–339. [Google Scholar] [CrossRef]
- Hauck, M. Site factors controlling epiphytic lichen abundance in northern coniferous forests. Flora 2011, 206, 81–90. [Google Scholar] [CrossRef]
- Dymytrova, L.; Nadyeina, O.; Hobi, M.L.; Scheidegger, C. Topographic and forest-stand variables determining epiphytic lichen diversity in the primeval beech forest in the Ukrainian Carpathians. Biodivers. Conserv. 2014, 23, 1367–1394. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L. Epiphytic lichen diversity along elevational gradients: Biological traits reveal a complex response to water and energy. J. Biogeogr. 2015, 42, 1222–1232. [Google Scholar] [CrossRef]
- Aussenac, G. Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Ann. For. Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]
- Dettki, H.; Klintberg, P.; Esseen, P.A. Are epiphytic lichens in young forests limited by local dispersal? Écoscience 2000, 7, 317–325. [Google Scholar] [CrossRef]
- Aude, E.; Poulsen, R.S. Influence of management on the species composition of epiphytic cryptogams in Danish Fagus forests. Appl. Veg. Sci. 2000, 3, 81–88. [Google Scholar] [CrossRef]
- Mikhailova, I.; Trubina, M.; Vorobeichik, E.; Scheidegger, C. Influence of environmental factors on the local-scale distribution of cyanobacterial lichens: Case study in the North Urals, Russia. Folia Cryptogam. Est. 2005, 41, 45–54. [Google Scholar]
- Löbel, S.; Snäll, T.; Rydin, H. Species richness patterns and metapopulation processes-evidence from epiphyte communities in boreo-nemoral forests. Ecography 2006, 29, 169–182. [Google Scholar] [CrossRef]
- Fritz, Ö.; Gustafsson, L.; Larsson, K. Does forest continuity matter in conservation? A study of epiphytic lichens and bryophytes in beech forests of southern Sweden. Biol. Conserv. 2008, 141, 655–668. [Google Scholar] [CrossRef]
- Mežaka, A.; Brūmelis, G.; Piterāns, A. Tree and stand-scale factors affecting richness and composition of epiphytic bryophytes and lichens in deciduous woodland key habitats. Biodivers. Conserv. 2012, 21, 3221–3241. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explanatory Variable | Number of Variables | Source of the Data |
|---|---|---|
| Bioclimatic | 19 bioclimatic and 18 ENVIREM variables, 37 in total | [64,65] |
| Topographic | 40 | Derived from Aster digital elevation model (DEM) using Saga GIS software [66] |
| Stand structure | 10 | Field survey |
| Parent rock | 1 categorical variable | 1:25,000 parent rock maps |
| Tree properties | 6 | Field survey and laboratory work |
| Group | Indicator Species | A# | B## | IndVal | p-Value |
|---|---|---|---|---|---|
| Geographic Region (LPA data) | |||||
| Simav | Bryoria implexa | 0.6842 | 0.5000 | 0.585 | 0.022 * |
| Letharia vulpina | 0.6500 | 0.5000 | 0.570 | 0.042 * | |
| Dursunbey and Sındırgı | Platismatia glauca | 0.9135 | 0.8810 | 0.897 | 0.002 ** |
| Geographic Region (LFQ data) | |||||
| Simav | Usnea subfloridana | 0.7301 | 0.6667 | 0.698 | 0.008 ** |
| Dursunbey and Sındırgı | Platismatia glauca | 0.9693 | 0.8810 | 0.924 | 0.001 *** |
| Age Class (LPA data) | |||||
| Old-growth | Hypocenomyce scalaris | 0.6652 | 0.8333 | 0.745 | 0.003 ** |
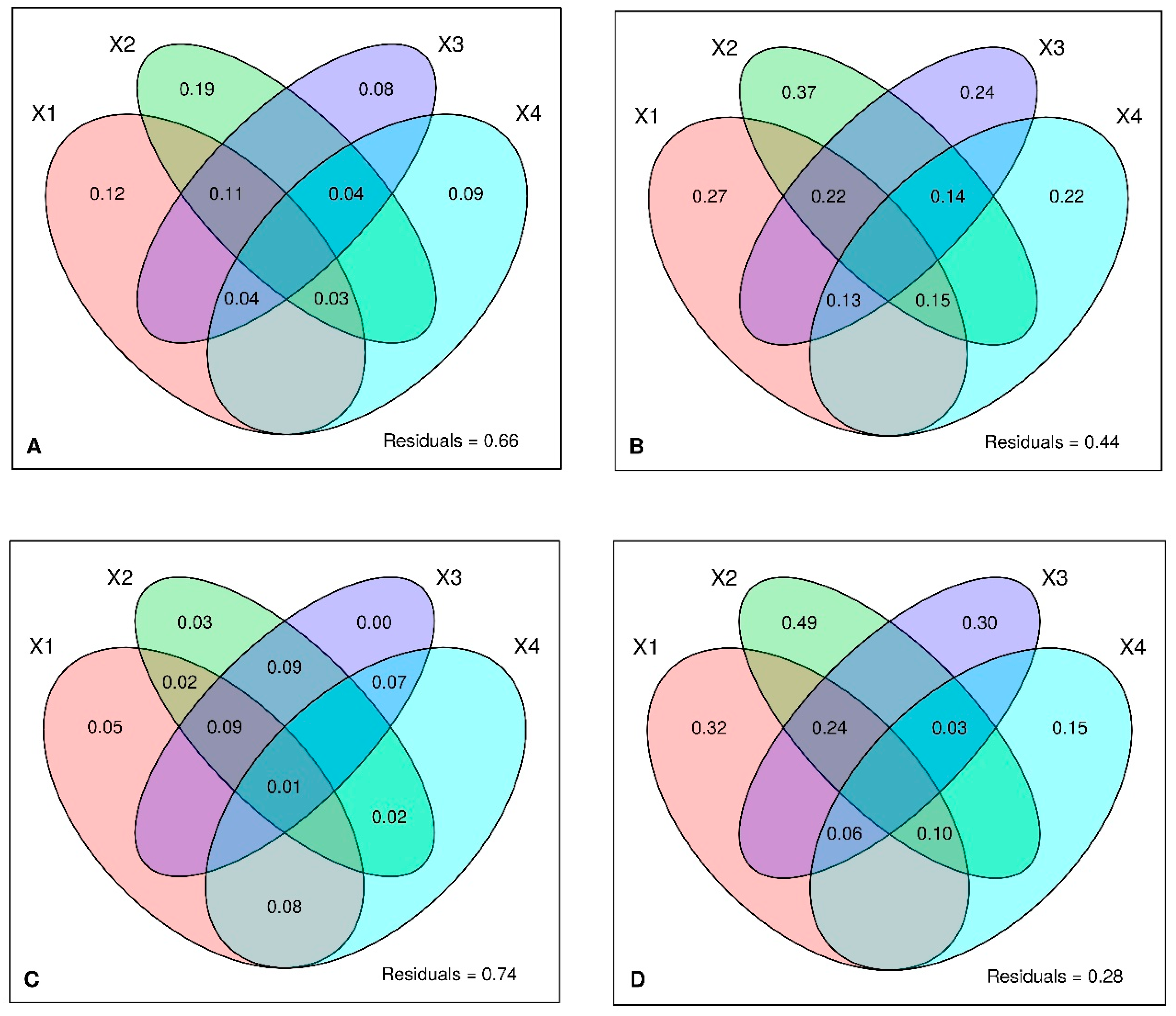
| Variable | Variance | F | p-Value |
|---|---|---|---|
| LPA (R2 = 0.365, adjusted R2 = 0.254) | |||
| Elev | 0.2645 | 4.4409 | 0.001 |
| maxH | 0.1866 | 3.1317 | 0.002 |
| Bio15 | 0.1434 | 2.4076 | 0.003 |
| mDbh | 0.1206 | 2.0252 | 0.016 |
| TnCurv | 0.1304 | 2.1893 | 0.006 |
| CtchSlp | 0.1206 | 2.0245 | 0.009 |
| Txtr | 0.1271 | 2.1342 | 0.011 |
| hecTBA | 0.1183 | 1.9865 | 0.012 |
| AridxThorn | 0.0962 | 1.6156 | 0.054 |
| MonthTemp10 | 0.0971 | 1.6306 | 0.049 |
| GLPA (R2 = 0.389 adjusted R2 = 0.306) | |||
| Elev | 0.1523 | 5.9435 | 0.001 |
| mDbh | 0.1105 | 4.3146 | 0.002 |
| mCH | 0.0860 | 3.3587 | 0.001 |
| MonthTemp10 | 0.0621 | 2.4253 | 0.015 |
| Bio15 | 0.0604 | 2.3577 | 0.026 |
| DwnCurv | 0.0587 | 2.2918 | 0.032 |
| AridxThorn | 0.0592 | 2.3092 | 0.026 |
| LFQ (R2 = 0.397, adjusted R2 = 0.234) | |||
| Elev | 0.0993 | 5.9836 | 0.001 |
| mCH | 0.0631 | 3.7976 | 0.001 |
| mDbh | 0.0476 | 2.8651 | 0.002 |
| Parent rock | 0.1061 | 1.5978 | 0.008 |
| HI | 0.0305 | 1.8374 | 0.050 |
| sdCH | 0.0298 | 1.7967 | 0.045 |
| CtchSlp | 0.0286 | 1.7233 | 0.068 |
| GLFQ (R2 = 0.342, adjusted R2 = 0.245) | |||
| Elev | 0.0561 | 6.9371 | 0.001 |
| mDbh | 0.0340 | 4.1971 | 0.002 |
| Bio15 | 0.0291 | 3.5964 | 0.006 |
| DwnCurv | 0.0183 | 2.2653 | 0.030 |
| HI | 0.0173 | 2.1435 | 0.049 |
| mCH | 0.0172 | 2.1230 | 0.053 |
| Model | Adjusted R2 | F-Statistic | p-Value |
|---|---|---|---|
| Richness = 4.687085 + 0.010691 × Dbh | 0.0028 | 2.0624 | 0.152 |
| Richness = 3.80421 + 0.06814 × Height | 0.0577 | 23.7665 | 0.0001 *** |
| Richness = 5.1316 − 0.5181 × BarkN | −0.0021 | 0.2865 | 0.593 |
| Richness = 12.0355 − 1.7572 × Bark_pH | 0.0515 | 20.8721 | 0.0001 *** |
| Richness = 4.63093 + 0.09175 × DBT | 0.0116 | 5.36 | 0.0211* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sevgi, E.; Yılmaz, O.Y.; Çobanoğlu Özyiğitoğlu, G.; Tecimen, H.B.; Sevgi, O. Factors Influencing Epiphytic Lichen Species Distribution in a Managed Mediterranean Pinus nigra Arnold Forest. Diversity 2019, 11, 59. https://doi.org/10.3390/d11040059
Sevgi E, Yılmaz OY, Çobanoğlu Özyiğitoğlu G, Tecimen HB, Sevgi O. Factors Influencing Epiphytic Lichen Species Distribution in a Managed Mediterranean Pinus nigra Arnold Forest. Diversity. 2019; 11(4):59. https://doi.org/10.3390/d11040059
Chicago/Turabian StyleSevgi, Ece, Osman Yalçın Yılmaz, Gülşah Çobanoğlu Özyiğitoğlu, Hüseyin Barış Tecimen, and Orhan Sevgi. 2019. "Factors Influencing Epiphytic Lichen Species Distribution in a Managed Mediterranean Pinus nigra Arnold Forest" Diversity 11, no. 4: 59. https://doi.org/10.3390/d11040059
APA StyleSevgi, E., Yılmaz, O. Y., Çobanoğlu Özyiğitoğlu, G., Tecimen, H. B., & Sevgi, O. (2019). Factors Influencing Epiphytic Lichen Species Distribution in a Managed Mediterranean Pinus nigra Arnold Forest. Diversity, 11(4), 59. https://doi.org/10.3390/d11040059