
Ganoderic Acid Ameliorates Ulcerative Colitis by Improving Intestinal Barrier Function via Gut Microbiota Modulation

 ,
,  and
and
Abstract
1. Introduction
2. Results
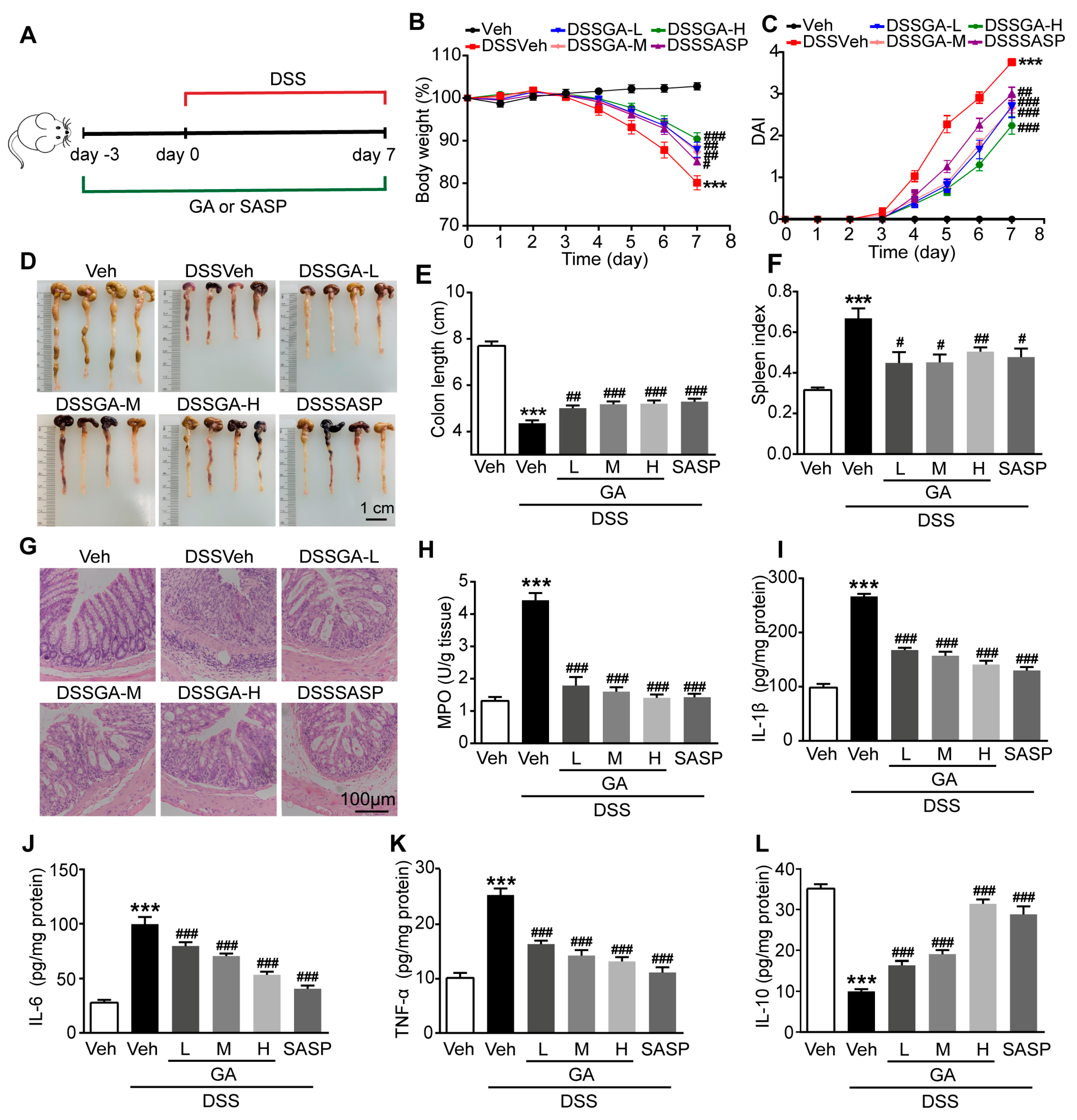
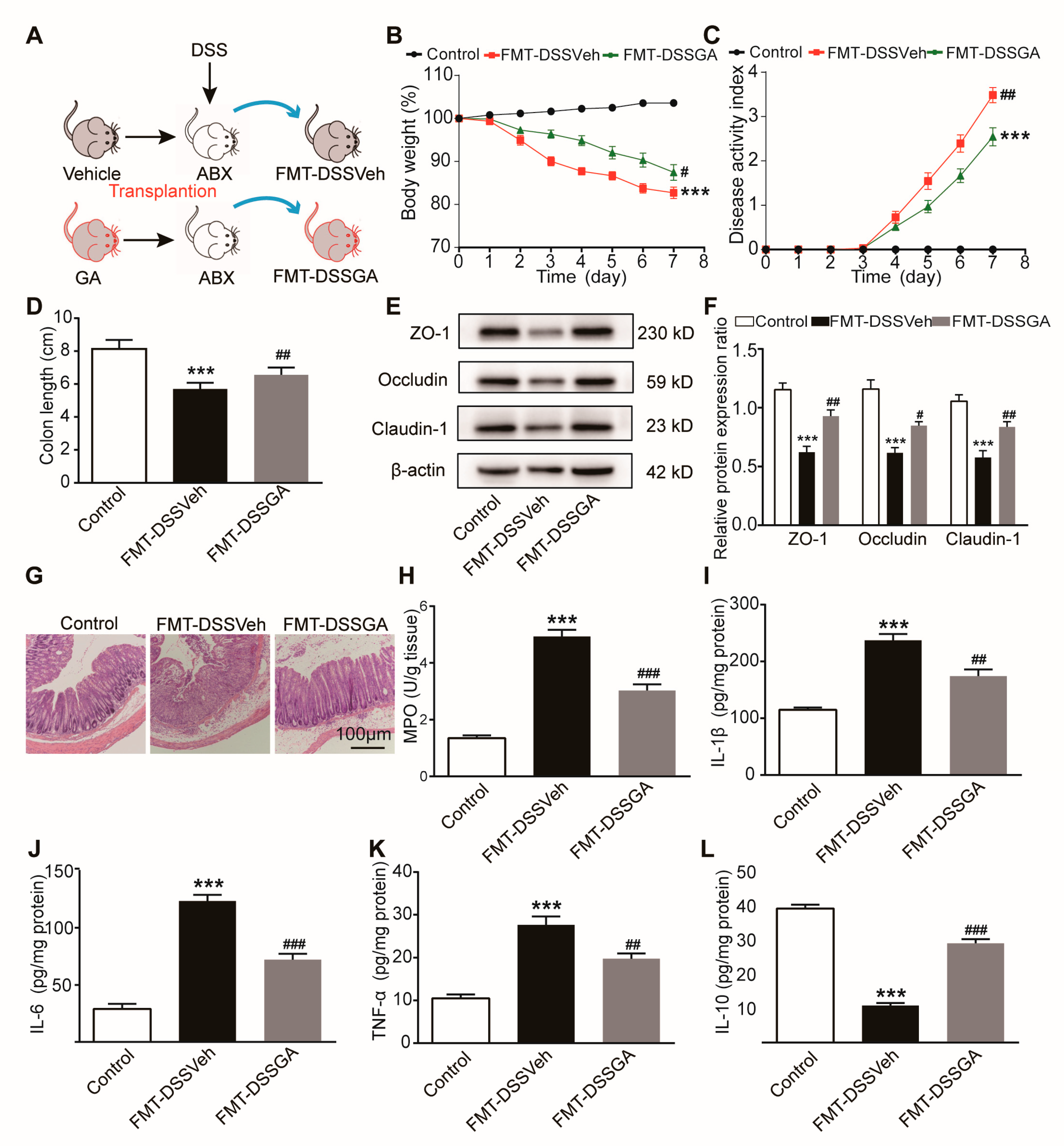
2.1. GA Ameliorated DSS-Induced UC in Mice
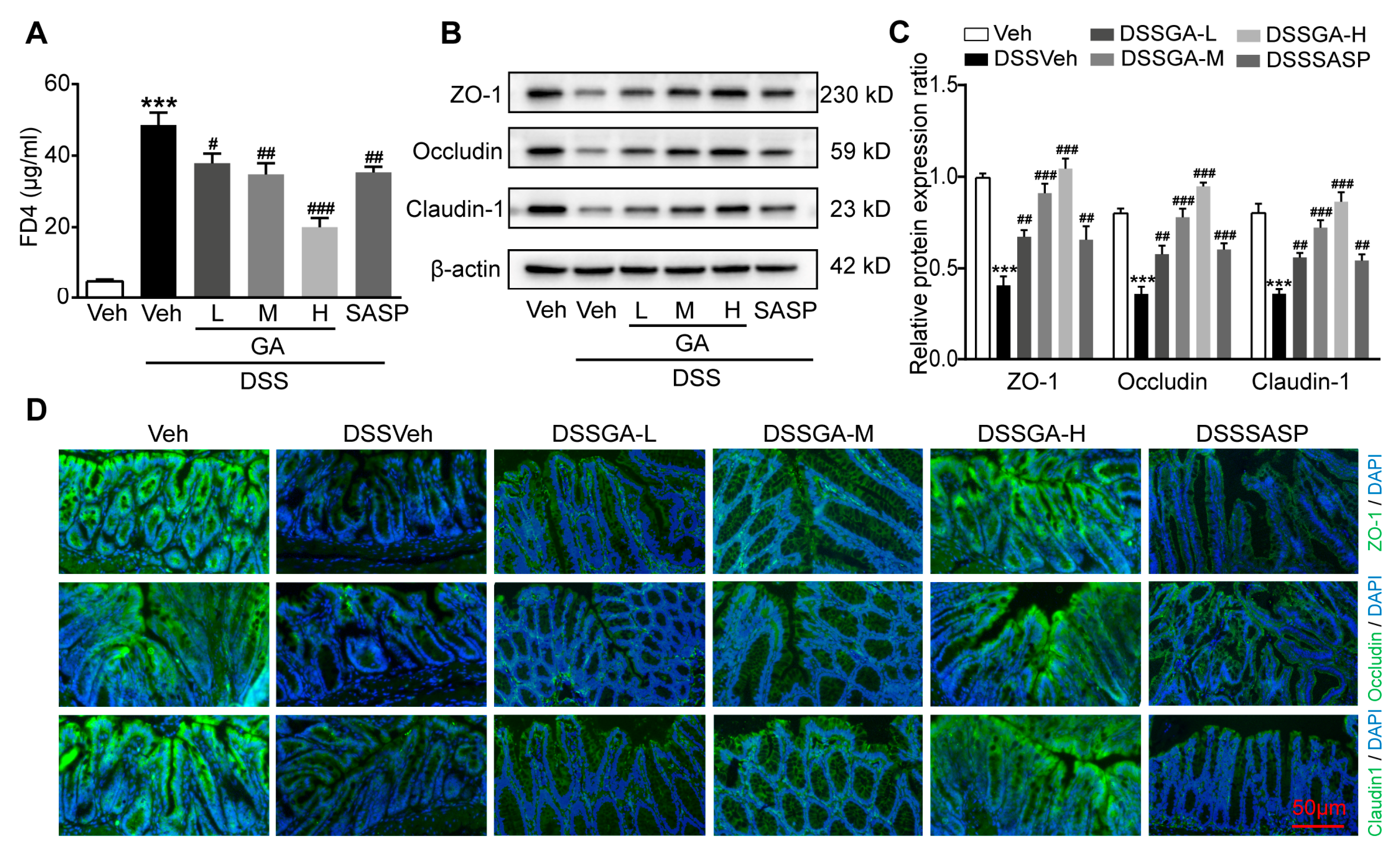
2.2. GA Improved Intestinal Barrier Function in DSS-Induced UC Mice
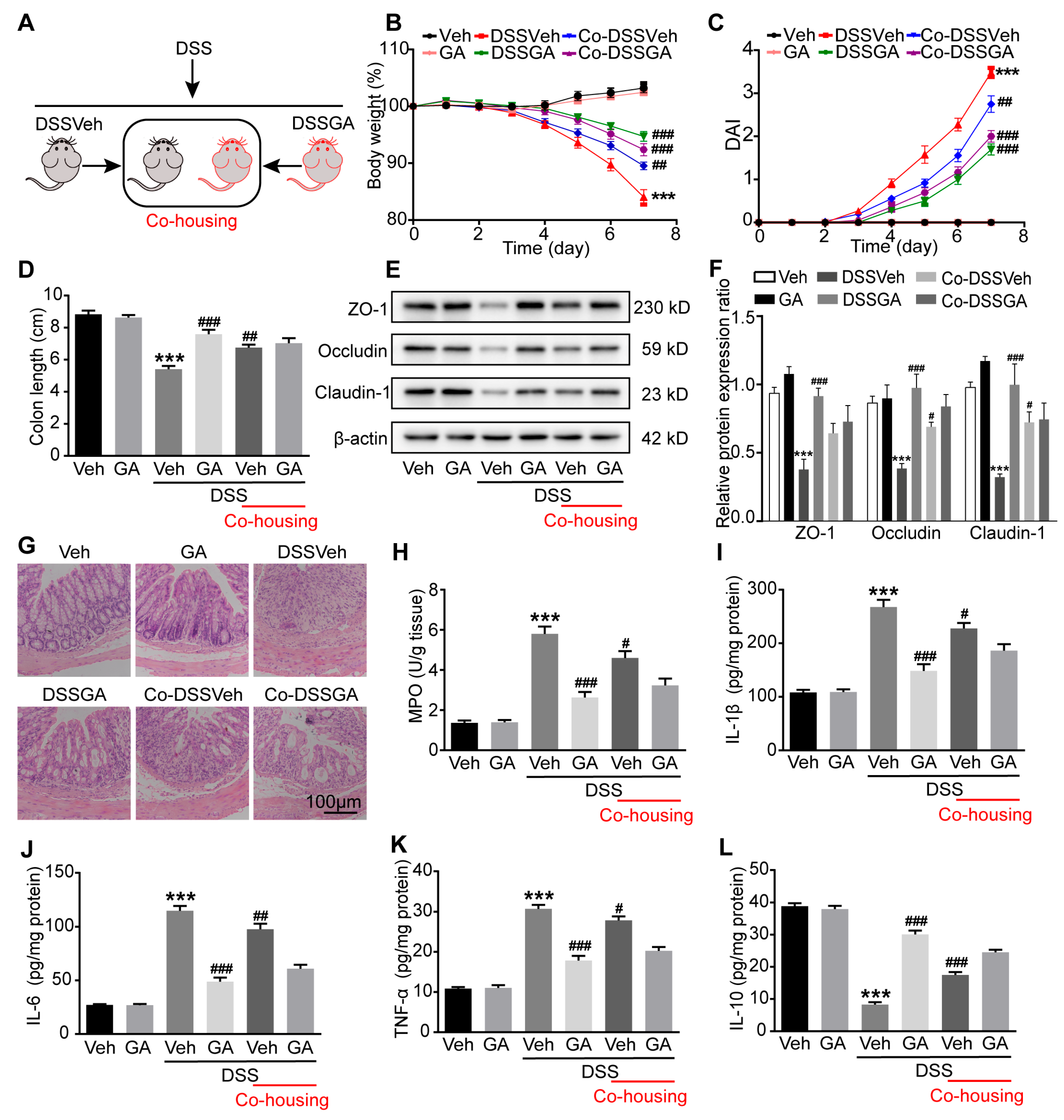
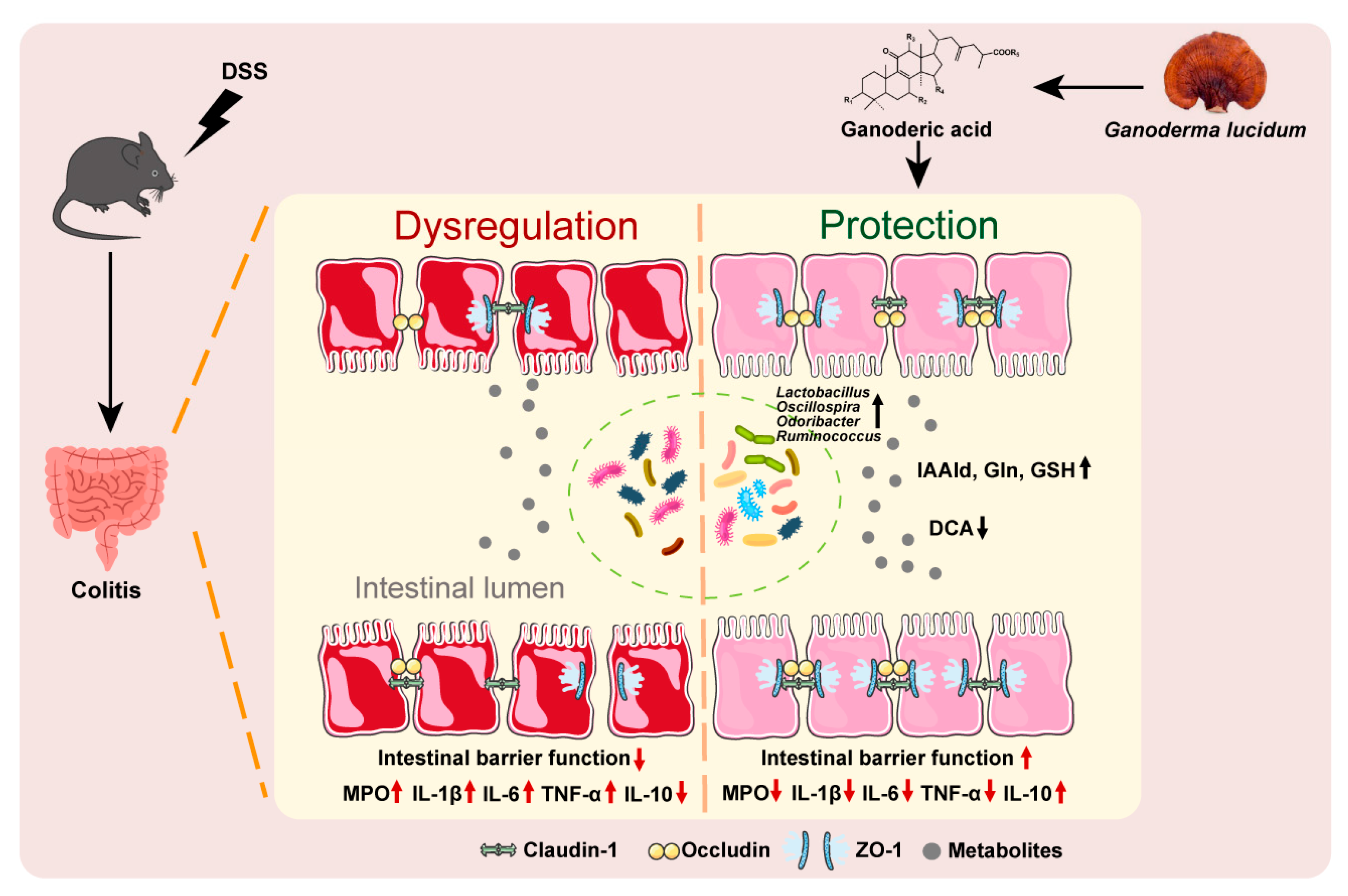
2.3. Gut Microbiota Were Involved in the Protective Effect GA on UC
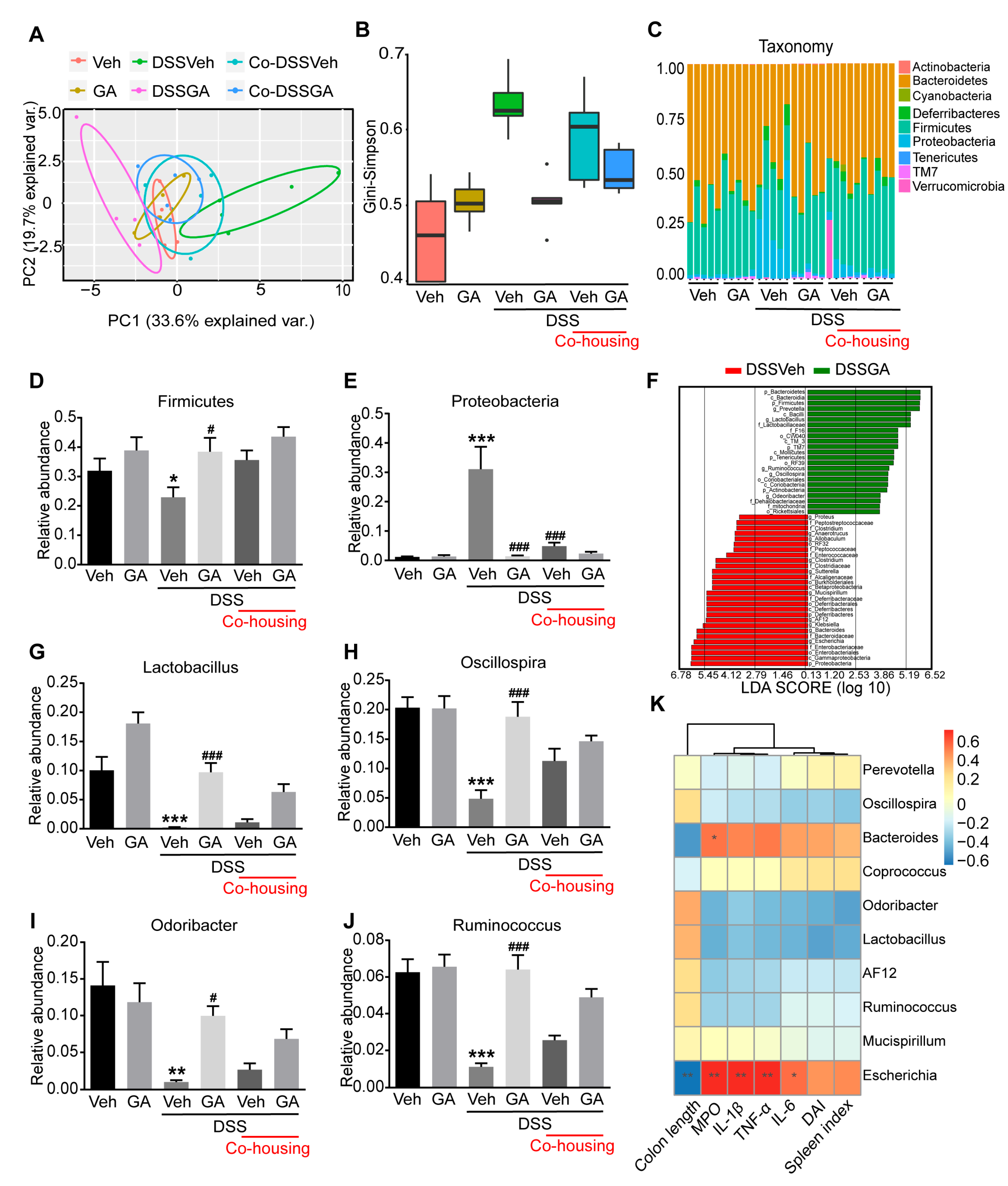
2.4. GA Alleviates Gut Microbiota Dysbiosis in DSS-Induced UC Mice
2.5. GA Ameliorated UC via Regulating the Gut Microbiota
2.6. GA Ameliorated UC by Gut Microbiota-Related Metabolites
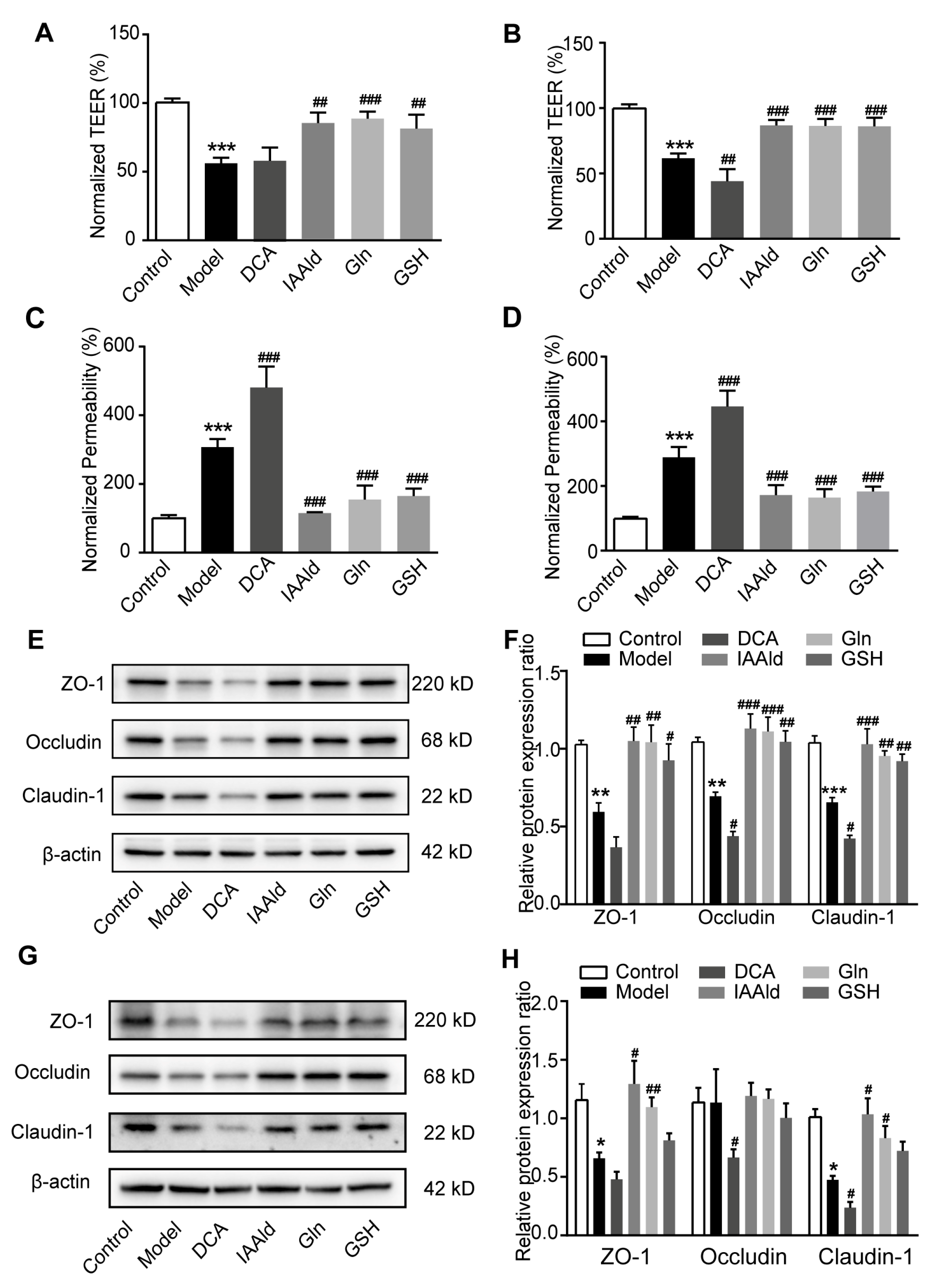
2.7. IAAld, Gln and GSH Directly Protected Intestinal Barrier Damage
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals and Reagents
4.3. Establishment and Treatment of Ulcerative Colitis
4.4. Co-Housing Experiment
4.5. Fecal Microbiota Transplantation
4.6. Assessment of the Disease Activity Index
4.7. Histological Assessment
4.8. Biochemical Analyses
4.9. Intestinal Permeability Assay
4.10. Real-Time qPCR
4.11. Western Blot Analysis
4.12. Immunofluorescence
4.13. 16S rDNA High-Throughput Sequencing and Analysis
4.14. UHPLC-MS/MS Analysis
4.15. Cell Culture and Metabolites Treatment
4.16. Measurement of Transepithelial Electrical Resistance
4.17. Paracellular Permeability Assay
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GA | Ganoderic acid |
| DSS | Dextran sulfate sodium |
| UC | Ulcerative colitis |
| SASP | Sulfasalazine |
| DAI | Disease activity index |
| FMT | Fecal microbiota transplantation |
| TNF-α | Tumor necrosis factor-α |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin-6 |
| IL-10 | Interleukin-10 |
| ZO-1 | Zonula occludens-1 |
| MPO | Myeloperoxidase |
| IAAld | Indole-3-acetaldehyde |
| Gln | Glutamine |
| GSH | Glutathione |
| DCA | Deoxycholic acid |
| H&E | Hematoxylin and eosin |
| PCoA | Principal co-ordinate analysis |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LEfSe | Linear discriminant analysis effect size |
References
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.-F. Ulcerative colitis. The Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Siegmund, B.; Le Berre, C.; Wei, S.C.; Ferrante, M.; Shen, B.; Bernstein, C.N.; Danese, S.; Peyrin-Biroulet, L.; Hibi, T. Ulcerative colitis. Nat. Rev. Dis. Primers 2020, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Gros, B.; Kaplan, G.G. Ulcerative Colitis in Adults: A Review. Jama J. Am. Med. Assoc. 2023, 330, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Bernstein, C.N. Environmental risk factors for inflammatory bowel disease. United Eur. Gastroenterol. J. 2022, 10, 1047–1053. [Google Scholar] [CrossRef]
- Qiu, P.; Ishimoto, T.; Fu, L.F.; Zhang, J.; Zhang, Z.Y.; Liu, Y. The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell. Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef]
- Peng, S.; Shen, L.; Yu, X.Y.; Zhang, L.; Xu, K.; Xia, Y.; Zha, L.L.; Wu, J.; Luo, H.S. The role of Nrf2 in the pathogenesis and treatment of ulcerative colitis. Front. Immunol. 2023, 14, 1200111. [Google Scholar] [CrossRef]
- Biedermann, L.; Rogler, G. The intestinal microbiota: Its role in health and disease. Eur. J. Pediatr. 2015, 174, 151–167. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Peng, X.X.; Shao, H.Q.; Li, X.Y.; Wu, Y.; Tan, Z.J. Gut Microbiota Comparison Between Intestinal Contents and Mucosa in Mice With Repeated Stress-Related Diarrhea Provides Novel Insight. Front. Microbiol. 2021, 12, 626691. [Google Scholar] [CrossRef]
- Imhann, F.; Vich Vila, A.; Bonder, M.J.; Fu, J.; Gevers, D.; Visschedijk, M.C.; Spekhorst, L.M.; Alberts, R.; Franke, L.; van Dullemen, H.M.; et al. Interplay of host genetics and gut microbiota underlying the onset and clinical presentation of inflammatory bowel disease. Gut 2018, 67, 108–119. [Google Scholar] [CrossRef]
- Lee, W.J.; Hase, K. Gut microbiota-generated metabolites in animal health and disease. Nat. Chem. Biol. 2014, 10, 416–424. [Google Scholar] [CrossRef]
- Caenepeel, C.; Sadat Seyed Tabib, N.; Vieira-Silva, S.; Vermeire, S. Review article: How the intestinal microbiota may reflect disease activity and influence therapeutic outcome in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2020, 52, 1453–1468. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Cao, G.Q.; Luo, C.; Tan, D.C.; Vong, C.T.; Xu, Y.Y.; Wang, S.C.; Lu, H.T.; Wang, Y.T.; Jing, W.H. Emerging pharmacotherapy for inflammatory bowel diseases. Pharmacol. Res. 2022, 178, 106146. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Dong, S.; Luo, X.; Liu, J.; Wei, B.; Du, W.; Yang, L.; Luo, H.; Wang, Y.; Wang, S.; et al. Berberine improves colitis by triggering AhR activation by microbial tryptophan catabolites. Pharmacol. Res. 2021, 164, 105358. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Zhu, M.; Wang, K.; Zhao, X.; Hu, L.; Jing, W.; Lu, H.; Wang, S. Dihydromyricetin improves DSS-induced colitis in mice via modulation of fecal-bacteria-related bile acid metabolism. Pharmacol. Res. 2021, 171, 105767. [Google Scholar] [CrossRef]
- Qi, M.; Chu, S.H.; Wang, W.X.; Fu, X.L.; Jiang, C.; Zhang, L.; Ali, M.H.; Lu, Y.T.; Jia, M.W.; Ubul, D.; et al. Safflower polysaccharide ameliorates acute ulcerative colitis by regulating STAT3/NF-κB signaling pathways and repairing intestinal barrier function. Biomed. Pharmacother. 2024, 174, 116553. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Ahmad, F.A.; Zeyaullah, M.; Alsayegh, A.A.; Mahmood, S.E.; AlShahrani, A.M.; Khan, M.S.; Shama, E.; Hamouda, A.; Elbendary, E.Y.; et al. Novel Insight into Hepatoprotective Potential with Mechanisms of Action. Nutrients 2023, 15, 1874. [Google Scholar] [CrossRef]
- Meng, M.; Wang, L.; Yao, Y.; Lin, D.; Wang, C.; Yao, J.; Sun, H.; Liu, M. Ganoderma lucidum polysaccharide peptide (GLPP) attenuates rheumatic arthritis in rats through inactivating NF-κB and MAPK signaling pathways. Phytomedicine 2023, 119, 155010. [Google Scholar] [CrossRef]
- Guo, C.; Guo, D.; Fang, L.; Sang, T.; Wu, J.; Guo, C.; Wang, Y.; Wang, Y.; Chen, C.; Chen, J.; et al. Ganoderma lucidum polysaccharide modulates gut microbiota and immune cell function to inhibit inflammation and tumorigenesis in colon. Carbohydr. Polym. 2021, 267, 118231. [Google Scholar] [CrossRef]
- Rahman, M.; Al-Ghamdi, S.A.; Alharbi, K.S.; Beg, S.; Sharma, K.; Anwar, F.; Al-Abbasi, F.A.; Kumar, V. Ganoderic acid loaded nano-lipidic carriers improvise treatment of hepatocellular carcinoma. Drug Deliv. 2019, 26, 782–793. [Google Scholar] [CrossRef]
- Abulizi, A.; Ran, J.; Ye, Y.; An, Y.; Zhang, Y.; Huang, Z.; Lin, S.; Zhou, H.; Lin, D.; Wang, L.; et al. Ganoderic acid improves 5-fluorouracil-induced cognitive dysfunction in mice. Food Funct. 2021, 12, 12325–12337. [Google Scholar] [CrossRef]
- Shao, G.; He, J.; Meng, J.; Ma, A.; Geng, X.; Zhang, S.; Qiu, Z.; Lin, D.; Li, M.; Zhou, H.; et al. Ganoderic Acids Prevent Renal Ischemia Reperfusion Injury by Inhibiting Inflammation and Apoptosis. Int. J. Mol. Sci. 2021, 22, 10229. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.L.; Guo, J.B.; Liu, B.Y.; Lu, J.Q.; Chen, M.; Liu, B.; Bai, W.D.; Rao, P.F.; Ni, L.; Lv, X.C. Ganoderic acid A from Ganoderma lucidum ameliorates lipid metabolism and alters gut microbiota composition in hyperlipidemic mice fed a high-fat diet. Food Funct. 2020, 11, 6818–6833. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Qi, H.; Tang, W.; Zhang, C.; Liu, Z.; Liu, Y.; Wei, X.; Kong, Z.; Jia, S.; et al. Probiotic fermentation of Ganoderma lucidum fruiting body extracts promoted its immunostimulatory activity in mice with dexamethasone-induced immunosuppression. Biomed. Pharmacother. 2021, 141, 111909. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.W.; Li, Z.Q.; Wang, J.L.; Jiang, S.Q.; Zhang, R.J.; He, Y.Q.; Xia, B.; Gao, J.M. Ganoderic Acid A Mitigates Inflammatory Bowel Disease through Modulation of AhR Activity by Microbial Tryptophan Metabolism. J. Agric. Food Chem. 2024, 72, 17912–17923. [Google Scholar] [CrossRef]
- Rath, T.; Atreya, R.; Bodenschatz, J.; Uter, W.; Geppert, C.E.; Vitali, F.; Fischer, S.; Waldner, M.J.; Colombel, J.F.; Hartmann, A.; et al. Intestinal Barrier Healing Is Superior to Endoscopic and Histologic Remission for Predicting Major Adverse Outcomes in Inflammatory Bowel Disease: The Prospective ERIca Trial. Gastroenterology 2023, 164, 241–255. [Google Scholar] [CrossRef]
- Vivinus-Nebot, M.; Frin-Mathy, G.; Bzioueche, H.; Dainese, R.; Bernard, G.; Anty, R.; Filippi, J.; Saint-Paul, M.C.; Tulic, M.K.; Verhasselt, V.; et al. Functional bowel symptoms in quiescent inflammatory bowel diseases: Role of epithelial barrier disruption and low-grade inflammation. Gut 2014, 63, 744–752. [Google Scholar] [CrossRef]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Wan, P.; Peng, Y.; Chen, G.; Xie, M.; Dai, Z.; Huang, K.; Dong, W.; Zeng, X.; Sun, Y. Modulation of gut microbiota by Ilex kudingcha improves dextran sulfate sodium-induced colitis. Food Res. Int. 2019, 126, 108595. [Google Scholar] [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Zuo, Y.; Xu, H.; Feng, D.; Shi, Y.; Li, J.; Wei, M.; Wang, L.; Zhang, Y.; Hu, P. Effect of fermented Rosa roxburghii Tratt fruit juice on gut microbiota in a dextran sulfate sodium-induced colitis mouse model. Food Sci. Hum. Wellness 2024, 13, 2234–2243. [Google Scholar] [CrossRef]
- Du, C.; Li, Z.; Zhang, J.; Yin, N.; Tang, L.; Li, J.; Sun, J.; Yu, X.; Chen, W.; Xiao, H.; et al. The protective effect of carnosic acid on dextran sulfate sodium-induced colitis based on metabolomics and gut microbiota analysis. Food Sci. Hum. Wellness 2023, 12, 1212–1223. [Google Scholar] [CrossRef]
- Ga, Y.; Wei, Y.; Zhao, Q.; Fan, Y.; Zhang, Y.; Zhang, Z.; Hao, S.; Wang, L.; Wang, Z.; Han, J.; et al. Puerariae Radix protects against ulcerative colitis in mice by inhibiting NLRP3 inflammasome activation. Food Sci. Hum. Wellness 2024, 13, 2266–2276. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, F.; Zhang, H.; Ye, Y.; Liu, P.; Lin, D.; Zhou, H.; Li, M.; Yang, B. Polysaccharides from Agaricus blazei Murrill ameliorate dextran sulfate sodium-induced colitis via attenuating intestinal barrier dysfunction. J. Funct. Foods 2022, 92, 105072. [Google Scholar] [CrossRef]
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef]
- Kuo, W.T.; Odenwald, M.A.; Turner, J.R.; Zuo, L. Tight junction proteins occludin and ZO-1 as regulators of epithelial proliferation and survival. Ann. N. Y. Acad. Sci. 2022, 1514, 21–33. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert. Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Landy, J.; Ronde, E.; English, N.; Clark, S.K.; Hart, A.L.; Knight, S.C.; Ciclitira, P.J.; Al-Hassi, H.O. Tight junctions in inflammatory bowel diseases and inflammatory bowel disease associated colorectal cancer. World J. Gastroenterol. 2016, 22, 3117–3126. [Google Scholar] [CrossRef]
- Chang, J.; Leong, R.W.; Wasinger, V.C.; Ip, M.; Yang, M.; Phan, T.G. Impaired Intestinal Permeability Contributes to Ongoing Bowel Symptoms in Patients With Inflammatory Bowel Disease and Mucosal Healing. Gastroenterology 2017, 153, 723–731.e1. [Google Scholar] [CrossRef]
- Kayama, H.; Okumura, R.; Takeda, K. Interaction Between the Microbiota, Epithelia, and Immune Cells in the Intestine. Annu. Rev. Immunol. 2020, 38, 23–48. [Google Scholar] [CrossRef]
- Khan, I.; Wei, J.; Li, A.; Liu, Z.; Yang, P.; Jing, Y.; Chen, X.; Zhao, T.; Bai, Y.; Zha, L.; et al. Lactobacillus plantarum strains attenuated DSS-induced colitis in mice by modulating the gut microbiota and immune response. Int. Microbiol. 2022, 25, 587–603. [Google Scholar] [CrossRef]
- Xu, D.W.; Xie, Y.C.; Cheng, J.; He, D.W.; Liu, J.X.; Fu, S.P.; Hu, G.Q. Amygdalin Alleviates DSS-Induced Colitis by Restricting Cell Death and Inflammatory Response, Maintaining the Intestinal Barrier, and Modulating Intestinal Flora. Cells 2024, 13, 444. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef] [PubMed]
- Scanu, M.; Toto, F.; Petito, V.; Masi, L.; Fidaleo, M.; Puca, P.; Baldelli, V.; Reddel, S.; Vernocchi, P.; Pani, G.; et al. An integrative multi-omic analysis defines gut microbiota, mycobiota, and metabolic fingerprints in ulcerative colitis patients. Front. Cell. Infect. Microbiol. 2024, 14, 1366192. [Google Scholar] [CrossRef]
- Chen, Y.; Jin, Y.; Stanton, C.; Paul Ross, R.; Zhao, J.; Zhang, H.; Yang, B.; Chen, W. Alleviation effects of Bifidobacterium breve on DSS-induced colitis depends on intestinal tract barrier maintenance and gut microbiota modulation. Eur. J. Nutr. 2021, 60, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.; Wang, M.; Ajibade, A.A.; Tan, P.; Fu, C.; Chen, L.; Zhu, M.; Hao, Z.Z.; Chu, J.; Yu, X.; et al. Microbiota regulate innate immune signaling and protective immunity against cancer. Cell Host Microbe 2021, 29, 959–974.e7. [Google Scholar] [CrossRef]
- Nascimento, R.P.D.; Machado, A.; Galvez, J.; Cazarin, C.B.B.; Marostica Junior, M.R. Ulcerative colitis: Gut microbiota, immunopathogenesis and application of natural products in animal models. Life Sci. 2020, 258, 118129. [Google Scholar] [CrossRef]
- Carstens, A.; Dicksved, J.; Nelson, R.; Lindqvist, M.; Andreasson, A.; Bohr, J.; Tysk, C.; Talley, N.J.; Agreus, L.; Engstrand, L.; et al. The Gut Microbiota in Collagenous Colitis Shares Characteristics With Inflammatory Bowel Disease-Associated Dysbiosis. Clin. Transl. Gastroenterol. 2019, 10, e00065. [Google Scholar] [CrossRef]
- Yu, S.; Balasubramanian, I.; Laubitz, D.; Tong, K.; Bandyopadhyay, S.; Lin, X.; Flores, J.; Singh, R.; Liu, Y.; Macazana, C.; et al. Paneth Cell-Derived Lysozyme Defines the Composition of Mucolytic Microbiota and the Inflammatory Tone of the Intestine. Immunity 2020, 53, 398–416.e8. [Google Scholar] [CrossRef]
- Henke, M.T.; Kenny, D.J.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Ruminococcus gnavus, a member of the human gut microbiome associated with Crohn’s disease, produces an inflammatory polysaccharide. Proc. Natl. Acad. Sci. USA 2019, 116, 12672–12677. [Google Scholar] [CrossRef]
- Palmela, C.; Chevarin, C.; Xu, Z.; Torres, J.; Sevrin, G.; Hirten, R.; Barnich, N.; Ng, S.C.; Colombel, J.F. Adherent-invasive Escherichia coli in inflammatory bowel disease. Gut 2018, 67, 574–587. [Google Scholar] [CrossRef]
- Cevallos, S.A.; Lee, J.Y.; Velazquez, E.M.; Foegeding, N.J.; Shelton, C.D.; Tiffany, C.R.; Parry, B.H.; Stull-Lane, A.R.; Olsan, E.E.; Savage, H.P.; et al. 5-Aminosalicylic Acid Ameliorates Colitis and Checks Dysbiotic Escherichia coli Expansion by Activating PPAR-gamma Signaling in the Intestinal Epithelium. mBio 2021, 12, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ran, L.; Yang, Y.; Gao, X.L.; Peng, M.; Liu, S.D.; Sun, L.; Wan, J.; Wang, Y.; Yang, K.; et al. Deferasirox alleviates DSS-induced ulcerative colitis in mice by inhibiting ferroptosis and improving intestinal microbiota. Life Sci. 2023, 314, 121312. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Su, S.; Zha, X.L.; Wei, Y.; Yang, G.; Huang, Q.Y.; Yang, Y.J.; Xia, L.; Fan, S.J.; Peng, X. Glutamine promotes O-GlcNAcylation of G6PD and inhibits AGR2 S-glutathionylation to maintain the intestinal mucus barrier in burned septic mice. Redox Biol. 2023, 59, 102581. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Li, Y.; Bao, X.; Yang, F.; Tang, X.; Jiang, Q.; Yang, C.; Yin, Y.; Yao, K. Glutamine boosts intestinal stem cell-mediated small intestinal epithelial development during early weaning: Involvement of WNT signaling. Stem Cell Rep. 2023, 18, 1451–1467. [Google Scholar] [CrossRef]
- Fang, Y.-x.; Lu, E.-q.; Cheng, Y.-j.; Xu, E.; Zhu, M.; Chen, X. Glutamine Promotes Porcine Intestinal Epithelial Cell Proliferation through the Wnt/β-Catenin Pathway. J. Agric. Food Chem. 2024, 72, 7155–7166. [Google Scholar] [CrossRef]
- DeMarco, V.G.; Li, N.; Thomas, J.; West, C.M.; Neu, J. Glutamine and barrier function in cultured Caco-2 epithelial cell monolayers. J. Nutr. 2003, 133, 2176–2179. [Google Scholar] [CrossRef]
- Wang, J.; Su, L.; Zhang, L.; Zeng, J.; Chen, Q.; Deng, R.; Wang, Z.; Kuang, W.; Jin, X.; Gui, S.; et al. Spirulina platensis aqueous extracts ameliorate colonic mucosal damage and modulate gut microbiota disorder in mice with ulcerative colitis by inhibiting inflammation and oxidative stress. J. Zhejiang Univ. Sci. B 2022, 23, 481–501. [Google Scholar] [CrossRef]
- Sun, Y.; Zheng, Y.; Wang, C.; Liu, Y. Glutathione depletion induces ferroptosis, autophagy, and premature cell senescence in retinal pigment epithelial cells. Cell Death Dis. 2018, 9, 753. [Google Scholar] [CrossRef]
- Bonetti, L.; Horkova, V.; Grusdat, M.; Longworth, J.; Guerra, L.; Kurniawan, H.; Franchina, D.G.; Soriano-Baguet, L.; Binsfeld, C.; Verschueren, C.; et al. A Th17 cell-intrinsic glutathione/mitochondrial-IL-22 axis protects against intestinal inflammation. Cell Metab. 2024, 36, 1726–1744.e10. [Google Scholar] [CrossRef]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ji, X.; Zhao, M.; Li, J.; Yin, H.; Jin, J.; Zhao, L. Modulation of tryptophan metabolism via AHR-IL22 pathway mediates the alleviation of DSS-induced colitis by chitooligosaccharides with different degrees of polymerization. Carbohydr. Polym. 2023, 319, 121180. [Google Scholar] [CrossRef] [PubMed]
- Azuma, Y.; Uchiyama, K.; Sugaya, T.; Yasuda, T.; Hashimoto, H.; Kajiwara-Kubota, M.; Sugino, S.; Kitae, H.; Torii, T.; Mizushima, K.; et al. Deoxycholic acid delays the wound healing of colonic epithelial cells via transmembrane G-protein-coupled receptor 5. J. Gastroenterol. Hepatol. 2022, 37, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chu, Q.; Dong, W.; Wang, X.; Zhao, W.; Dai, X.; Liu, W.; Wang, B.; Liu, T.; Zhong, W.; et al. Microbial metabolite deoxycholic acid-mediated ferroptosis exacerbates high-fat diet-induced colonic inflammation. Mol. Metab. 2024, 84, 101944. [Google Scholar] [CrossRef]
- He, J.; Sun, Y.; Jia, Y.; Geng, X.; Chen, R.; Zhou, H.; Yang, B. Ganoderma triterpenes Protect Against Hyperhomocysteinemia Induced Endothelial-Mesenchymal Transition via TGF-beta Signaling Inhibition. Front. Physiol. 2019, 10, 192. [Google Scholar] [CrossRef]
- Fan, Q.; Guan, X.; Hou, Y.; Liu, Y.; Wei, W.; Cai, X.; Zhang, Y.; Wang, G.; Zheng, X.; Hao, H. Paeoniflorin modulates gut microbial production of indole-3-lactate and epithelial autophagy to alleviate colitis in mice. Phytomedicine 2020, 79, 153345. [Google Scholar] [CrossRef]
- Murthy, S.N.; Cooper, H.S.; Shim, H.; Shah, R.S.; Ibrahim, S.A.; Sedergran, D.J. Treatment of dextran sulfate sodium-induced murine colitis by intracolonic cyclosporin. Dig. Dis. Sci. 1993, 38, 1722–1734. [Google Scholar] [CrossRef]
- Liu, J.; Hu, Z.; Liu, D.; Zheng, A.; Ma, Q. Glutathione metabolism-mediated ferroptosis reduces water-holding capacity in beef during cold storage. Food Chem. 2023, 398, 133903. [Google Scholar] [CrossRef]
- Cheng, Q.; Zhang, J.; Fang, J.; Ding, H.; Xu, Y.; Lu, X.; Zhang, W. Untargeted metabolomics reveals the role of AQP9 in nonalcoholic fatty liver disease in a mice model. Int. J. Biol. Macromol. 2022, 219, 864–875. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | FORWARD PRIMER (5′-3′) | REVERSE PRIMER (5′-3′) |
|---|---|---|
| IL-1β | TCCATGAGCTTTGTACAAGGA | AGCCCATACTTTAGGAAGACA |
| IL-6 | GTTCTCTGGGAAATCGTGGA | TGTACTCCAGGTAGCTA |
| TNF-α | AGACCCTCACACTCAGATCA | TCTTTGAGATCCATGCCGTTG |
| IL-10 | ATTTGAATTCCCTGGGTGAGAAG | CACAGGGGAGAAATCGATGACA |
| GAPDH | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Abulizi, A.; Zhang, Y.; Lu, F.; An, Y.; Ren, C.; Zhang, H.; Wang, Y.; Lin, D.; Lu, D.; et al. Ganoderic Acid Ameliorates Ulcerative Colitis by Improving Intestinal Barrier Function via Gut Microbiota Modulation. Int. J. Mol. Sci. 2025, 26, 2466. https://doi.org/10.3390/ijms26062466
Ye Y, Abulizi A, Zhang Y, Lu F, An Y, Ren C, Zhang H, Wang Y, Lin D, Lu D, et al. Ganoderic Acid Ameliorates Ulcerative Colitis by Improving Intestinal Barrier Function via Gut Microbiota Modulation. International Journal of Molecular Sciences. 2025; 26(6):2466. https://doi.org/10.3390/ijms26062466
Chicago/Turabian StyleYe, Yuwei, Abudumijiti Abulizi, Yukun Zhang, Feng Lu, Yongpan An, Chaoqun Ren, Hang Zhang, Yiming Wang, Dongmei Lin, Dan Lu, and et al. 2025. "Ganoderic Acid Ameliorates Ulcerative Colitis by Improving Intestinal Barrier Function via Gut Microbiota Modulation" International Journal of Molecular Sciences 26, no. 6: 2466. https://doi.org/10.3390/ijms26062466
APA StyleYe, Y., Abulizi, A., Zhang, Y., Lu, F., An, Y., Ren, C., Zhang, H., Wang, Y., Lin, D., Lu, D., Li, M., & Yang, B. (2025). Ganoderic Acid Ameliorates Ulcerative Colitis by Improving Intestinal Barrier Function via Gut Microbiota Modulation. International Journal of Molecular Sciences, 26(6), 2466. https://doi.org/10.3390/ijms26062466