
Multi-Omics Analysis Revealed the Molecular Mechanisms Affecting Average Daily Gain in Cattle
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
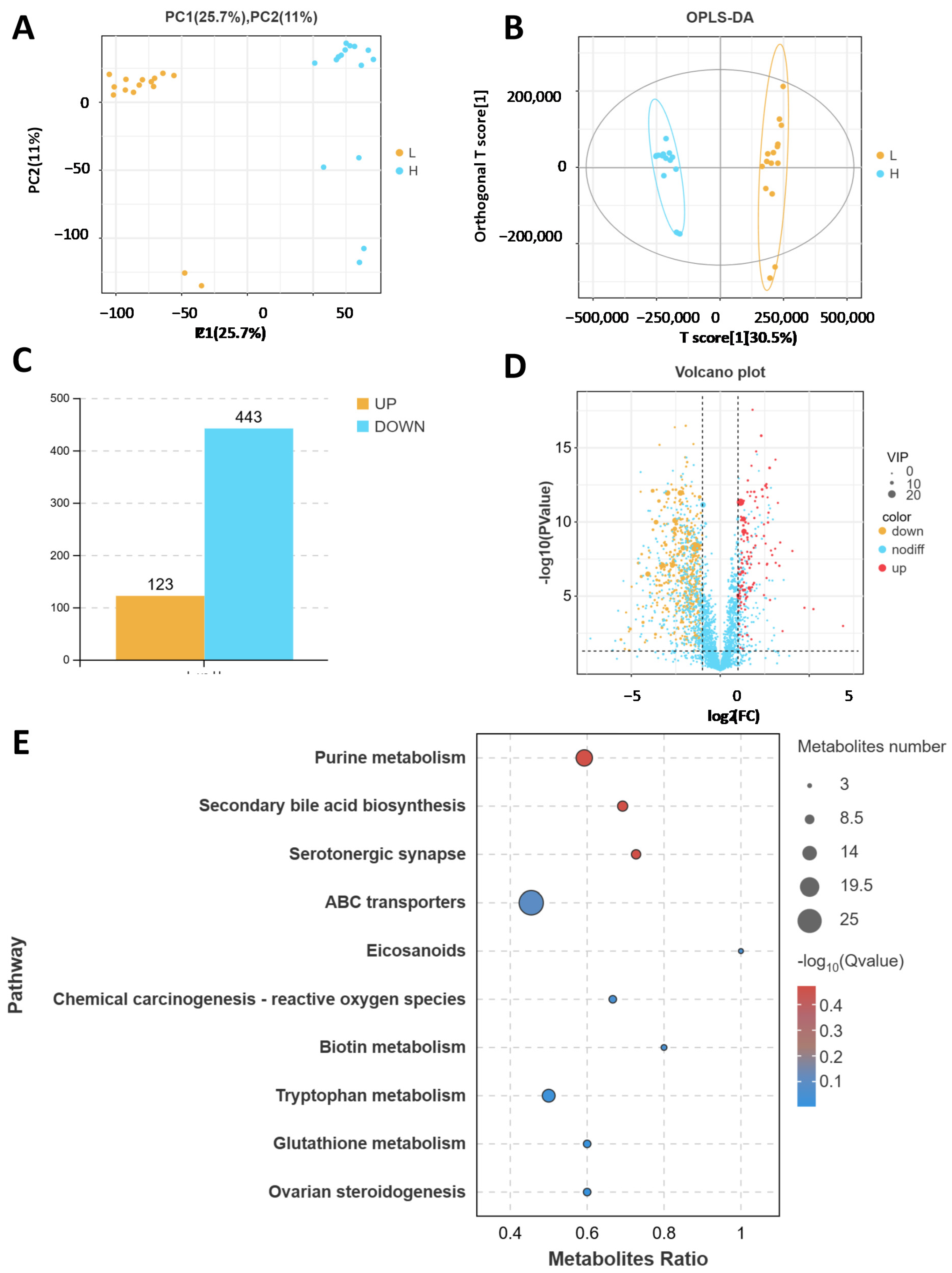
2.1. Comparison of the Fecal Microbiotas of High- and Low-Daily-Gain Performance Groups
2.2. Comparison of the Fecal Metabolites of High- and Low-Daily-Gain Performance Groups
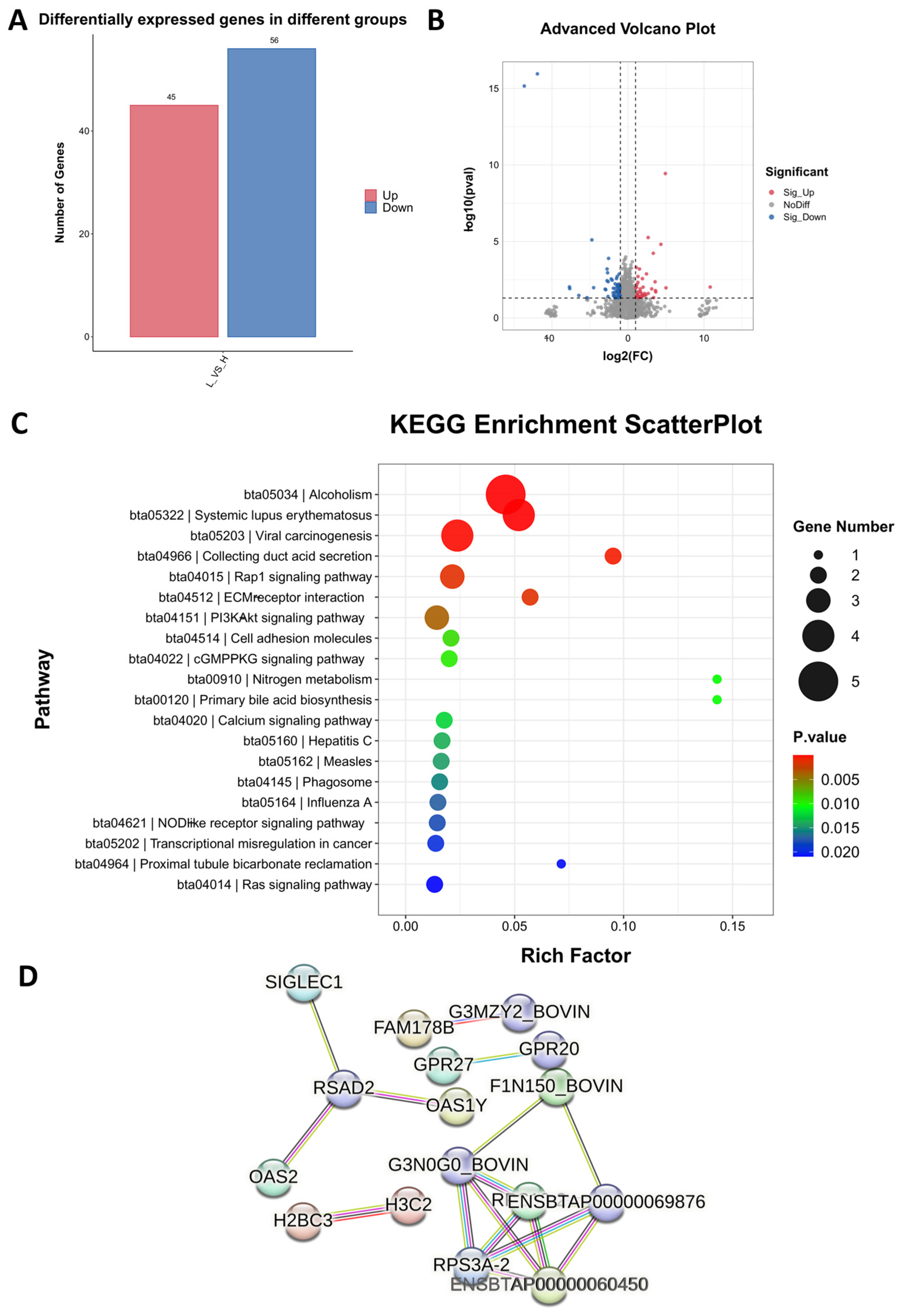
2.3. Comparison of the Blood Transcriptome of High- and Low-Daily-Gain Performance Groups
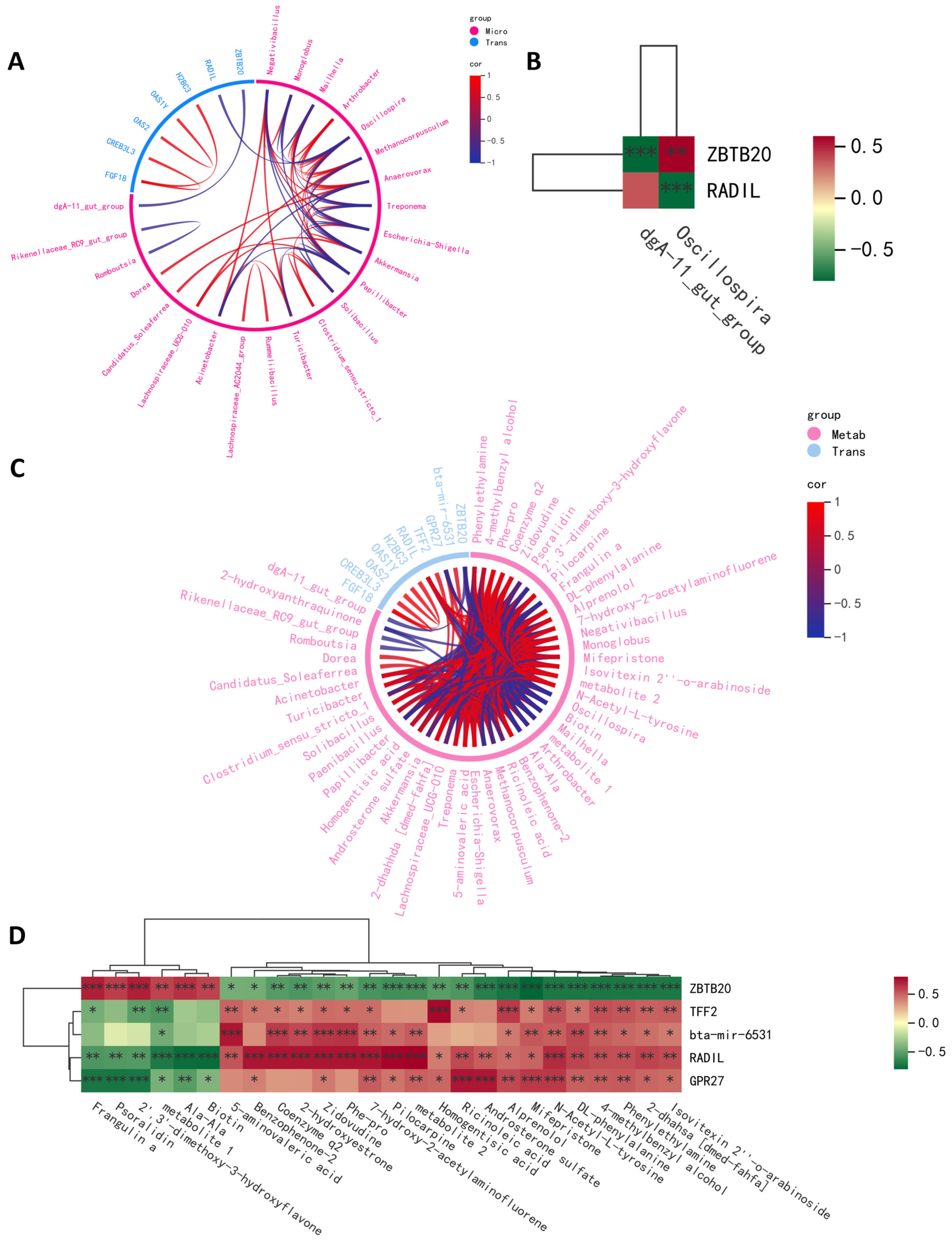
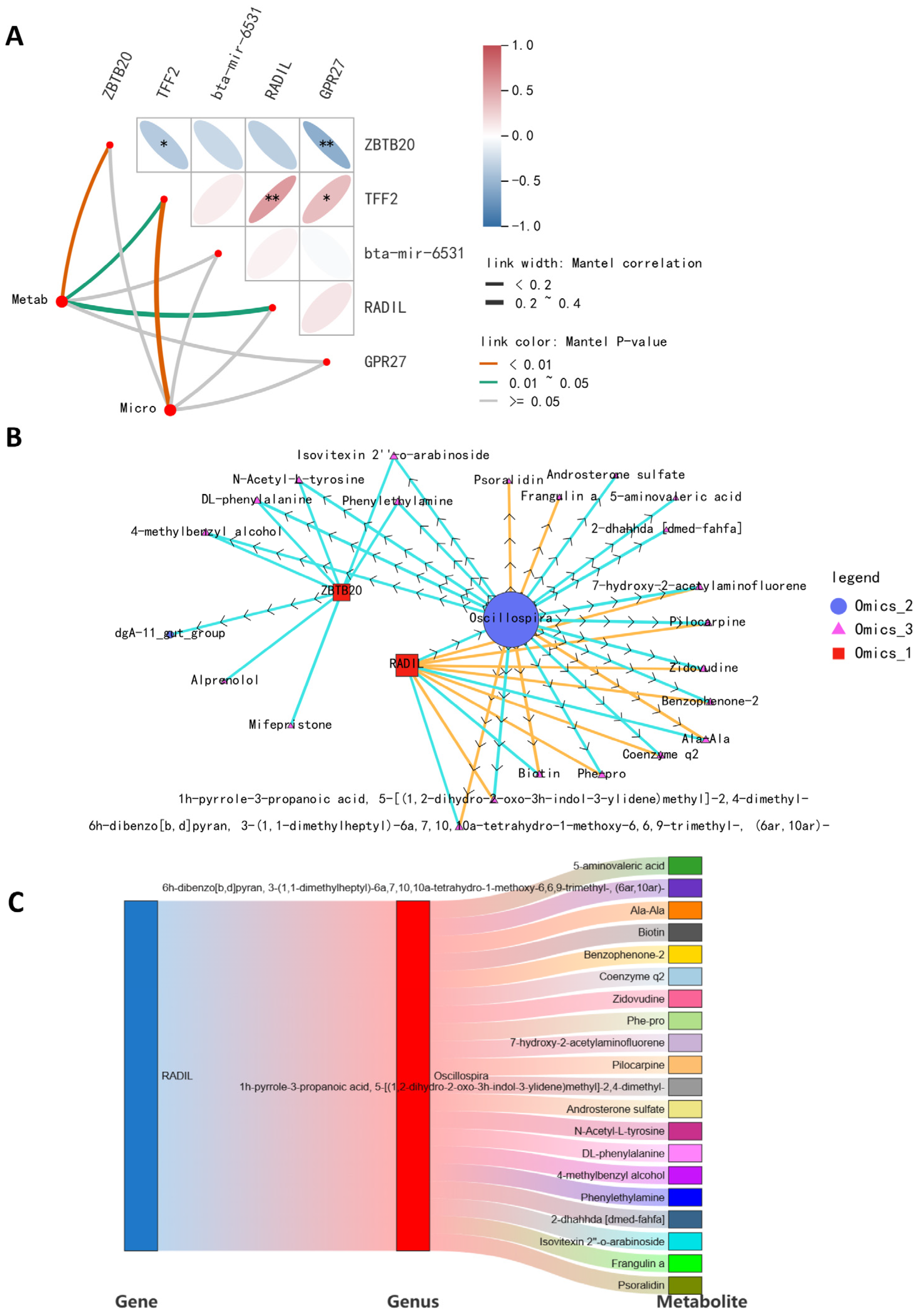
2.4. Integrative Crosstalk of Multi-Omics
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Collection
4.2. Amplicon Sequencing 16S rRNA and Subsequent Analysis
4.3. Untargeted Metabolomics Study and Analysis
4.4. RNA-Seq and Transcriptome Data Analysis
4.5. Correlation Analysis of Microbiome, Metabolome, and Transcriptom
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magalhaes, D.R.; Çakmakçı, C.; Campo, M.D.M.; Çakmakçı, Y.; Makishi, F.; Silva, V.; Trindade, M.A. Changes in the Current Patterns of Beef Consumption and Consumer Behavior Trends-Cross-Cultural Study Brazil-Spain-Turkey. Foods 2023, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Place, N.T.; Heinrichs, A.J.; Erb, H.N. The effects of disease, management, and nutrition on average daily gain of dairy heifers from birth to four months. J. Dairy Sci. 1998, 81, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Ding, L.; Wang, X.; Yan, Q.; Jiang, C.; Hu, C.; Wang, G.; Zhou, Y.; Henkin, Z.; Degen, A.A. Astragalus root extract improved average daily gain, immunity, antioxidant status and ruminal microbiota of early weaned yak calves. J. Sci. Food Agric. 2021, 101, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Voisinet, B.D.; Grandin, T.; Tatum, J.D.; O’Connor, S.F.; Struthers, J.J. Feedlot cattle with calm temperaments have higher average daily gains than cattle with excitable temperaments. J. Anim. Sci. 1997, 75, 892–896. [Google Scholar] [CrossRef]
- Foote, A.P.; Keel, B.N.; Zarek, C.M.; Lindholm-Perry, A.K. Beef steers with average dry matter intake and divergent average daily gain have altered gene expression in the jejunum. J. Anim. Sci. 2017, 95, 4430–4439. [Google Scholar] [CrossRef]
- Sheet, S.; Jang, S.S.; Kim, J.H.; Park, W.; Kim, D. A transcriptomic analysis of skeletal muscle tissues reveals promising candidate genes and pathways accountable for different daily weight gain in Hanwoo cattle. Sci. Rep. 2024, 14, 315. [Google Scholar] [CrossRef]
- Yin, X.; Duan, C.; Ji, S.; Tian, P.; Ju, S.; Yan, H.; Zhang, Y.; Liu, Y. Average Daily Gain in Lambs Weaned at 60 Days of Age Is Correlated with Rumen and Rectum Microbiota. Microorganisms 2023, 11, 348. [Google Scholar] [CrossRef]
- Weng, Y.; Xu, T.; Wang, C.; Jin, Y. Oral Exposure to Epoxiconazole Disturbed the Gut Micro-Environment and Metabolic Profiling in Male Mice. Metabolites 2023, 13, 522. [Google Scholar] [CrossRef]
- Zierer, J.; Jackson, M.A.; Kastenmüller, G.; Mangino, M.; Long, T.; Telenti, A.; Mohney, R.P.; Small, K.S.; Bell, J.T.; Steves, C.J.; et al. The fecal metabolome as a functional readout of the gut microbiome. Nat. Genet. 2018, 50, 790–795. [Google Scholar] [CrossRef]
- Song, F.; Qian, Y.; Peng, X.; Li, X.; Xing, P.; Ye, D.; Lei, H. The frontline of immune response in peripheral blood. PLoS ONE 2017, 12, e0182294. [Google Scholar] [CrossRef]
- Miao, Z.; Humphreys, B.D.; McMahon, A.P.; Kim, J. Multi-omics integration in the age of million single-cell data. Nat. Rev. Nephrol. 2021, 17, 710–724. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; McCoun, M. Average daily gain divergence in beef steers is associated with altered plasma metabolome and whole blood immune-related gene expression. Transl. Anim. Sci. 2020, 4, txaa074. [Google Scholar] [CrossRef]
- Du, L.; Li, K.; Chang, T.; An, B.; Liang, M.; Deng, T.; Cao, S.; Du, Y.; Cai, W.; Gao, X.; et al. Integrating genomics and transcriptomics to identify candidate genes for subcutaneous fat deposition in beef cattle. Genomics 2022, 114, 110406. [Google Scholar] [CrossRef]
- Yu, H.; Wang, J.; Zhang, K.; Cheng, G.; Mei, C.; Zan, L. Integrated multi-omics analysis reveals variation in intramuscular fat among muscle locations of Qinchuan cattle. BMC Genom. 2023, 24, 367. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Perlman, D.; Martínez-Álvaro, M.; Moraïs, S.; Altshuler, I.; Hagen, L.H.; Jami, E.; Roehe, R.; Pope, P.B.; Mizrahi, I. Concepts and Consequences of a Core Gut Microbiota for Animal Growth and Development. Annu. Rev. Anim. Biosci. 2022, 10, 177–201. [Google Scholar] [CrossRef]
- Lourenco, J.M.; Welch, C.B.; Krause, T.R.; Wieczorek, M.A.; Fluharty, F.L.; Rothrock, M.J.; Pringle, T.D.; Callaway, T.R. Fecal Microbiome Differences in Angus Steers with Differing Feed Efficiencies during the Feedlot-Finishing Phase. Microorganisms 2022, 10, 1128. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Zhang, M.L.; Zhang, R.Y.; Zhu, W.Y.; Mao, S.Y. Comparative studies of the composition of bacterial microbiota associated with the ruminal content, ruminal epithelium and in the faeces of lactating dairy cows. Microb. Biotechnol. 2016, 9, 257–268. [Google Scholar] [CrossRef]
- Zhou, G.; Liang, X.; He, X.; Li, J.; Tian, G.; Liu, Y.; Wang, X.; Chen, Y.; Yang, Y. Compound enzyme preparation supplementation improves the production performance of goats by regulating rumen microbiota. Appl. Microbiol. Biotechnol. 2023, 107, 7287–7299. [Google Scholar] [CrossRef]
- Jiang, S.; Si, J.; Mo, J.; Zhang, S.; Chen, K.; Gao, J.; Xu, D.; Bai, L.; Lan, G.; Liang, J. Integrated Microbiome and Serum Metabolome Analysis Reveals Molecular Regulatory Mechanisms of the Average Daily Weight Gain of Yorkshire Pigs. Animals 2024, 14, 278. [Google Scholar] [CrossRef]
- Fang, S.; Chen, X.; Pan, J.; Chen, Q.; Zhou, L.; Wang, C.; Xiao, T.; Gan, Q.F. Dynamic distribution of gut microbiota in meat rabbits at different growth stages and relationship with average daily gain (ADG). BMC Microbiol. 2020, 20, 116. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Niu, Y.; Wang, H.; Qin, S.; Zhang, R.; Wu, Y.; Xiao, X.; Xu, Y.; Yang, C. Effects of three different plant-derived polysaccharides on growth performance, immunity, antioxidant function, and cecal microbiota of broilers. J. Sci. Food Agric. 2024, 104, 1020–1029. [Google Scholar] [CrossRef]
- Huang, T.; Han, J.; Liu, Y.; Fei, M.; Du, X.; He, K.; Zhao, A. Dynamic distribution of gut microbiota in posthatching chicks and its relationship with average daily gain. Poult. Sci. 2023, 102, 103008. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Carlin, F.; Federighi, M.; Filippitzi, M.E.; Fournier, M.; Fravalo, P.; Ganière, J.P.; Grisot, L.; Guillier, L.; Hilaire, D.; et al. Clostridium botulinum type C, D, C/D, and D/C: An update. Front. Microbiol. 2022, 13, 1099184. [Google Scholar] [CrossRef]
- Alonazi, M.; Ben Bacha, A.; Al Suhaibani, A.; Almnaizel, A.T.; Aloudah, H.S.; El-Ansary, A. Psychobiotics improve propionic acid-induced neuroinflammation in juvenile rats, rodent model of autism. Transl. Neurosci. 2022, 13, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Kearney, S.M.; Gibbons, S.M.; Poyet, M.; Gurry, T.; Bullock, K.; Allegretti, J.R.; Clish, C.B.; Alm, E.J. Endospores and other lysis-resistant bacteria comprise a widely shared core community within the human microbiota. ISME J. 2018, 12, 2403–2416. [Google Scholar] [CrossRef]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A.J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; et al. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat. Commun. 2020, 11, 4457. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; La, A.T.Z.; Evans, A.; Gao, S.; Yu, Z.; Bu, D.; Ma, L. Supplementation with sodium butyrate improves growth and antioxidant function in dairy calves before weaning. J. Anim. Sci. Biotechnol. 2021, 12, 2. [Google Scholar] [CrossRef]
- Yu, H.; Yu, S.; Guo, J.; Wang, J.; Mei, C.; Abbas Raza, S.H.; Cheng, G.; Zan, L. Comprehensive Analysis of Transcriptome and Metabolome Reveals Regulatory Mechanism of Intramuscular Fat Content in Beef Cattle. J. Agric. Food Chem. 2024, 72, 2911–2924. [Google Scholar] [CrossRef]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef]
- Xue, M.Y.; Sun, H.Z.; Wu, X.H.; Liu, J.X.; Guan, L.L. Multi-omics reveals that the rumen microbiome and its metabolome together with the host metabolome contribute to individualized dairy cow performance. Microbiome 2020, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Ding, H.; Wang, J.; Xu, W.; Liu, Y.; Kenéz, Á. Metabolite Profile of Sheep Serum With High or Low Average Daily Gain. Front. Vet. Sci. 2021, 8, 662536. [Google Scholar] [CrossRef]
- Ogunade, I.; Oyebade, A.; Osa-Andrews, B.; Peters, S. Plasma Carboxyl-Metabolome Is Associated with Average Daily Gain Divergence in Beef Steers. Animals 2021, 11, 67. [Google Scholar] [CrossRef]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the human gut microbiome and host metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef] [PubMed]
- Goedert, J.J.; Sampson, J.N.; Moore, S.C.; Xiao, Q.; Xiong, X.; Hayes, R.B.; Ahn, J.; Shi, J.; Sinha, R. Fecal metabolomics: Assay performance and association with colorectal cancer. Carcinogenesis 2014, 35, 2089–2096. [Google Scholar] [CrossRef]
- Xu, S.Y.; Feng, X.R.; Zhao, W.; Bi, Y.L.; Diao, Q.Y.; Tu, Y. Rumen and hindgut microbiome regulate average daily gain of preweaning Holstein heifer calves in different ways. Microbiome 2024, 12, 131. [Google Scholar] [CrossRef] [PubMed]
- Mehrban, H.; Naserkheil, M.; Lee, D.H.; Ibáñez-Escriche, N. Genetic parameters and correlations of related feed efficiency, growth, and carcass traits in Hanwoo beef cattle. Anim. Biosci. 2021, 34, 824–832. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Guo, B.; Greenwood, P.L.; Cafe, L.M.; Zhou, G.; Zhang, W.; Dalrymple, B.P. Transcriptome analysis of cattle muscle identifies potential markers for skeletal muscle growth rate and major cell types. BMC Genom. 2015, 16, 177. [Google Scholar] [CrossRef]
- Sheet, S.; Jang, S.S.; Lim, J.A.; Park, W.; Kim, D. Molecular signatures diversity unveiled through a comparative transcriptome analysis of longissimus dorsi and psoas major muscles in Hanwoo cattle. Anim. Biotechnol. 2024, 35, 2379883. [Google Scholar] [CrossRef]
- Garner, J.B.; Chamberlain, A.J.; Vander Jagt, C.; Nguyen, T.T.T.; Mason, B.A.; Marett, L.C.; Leury, B.J.; Wales, W.J.; Hayes, B.J. Gene expression of the heat stress response in bovine peripheral white blood cells and milk somatic cells in vivo. Sci. Rep. 2020, 10, 19181. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Gong, S.; Han, M.; Wang, J.; Shi, H.; Jiang, X.; Guo, L.; Duan, Y.; Guo, Q.; Chen, Q.; et al. Leucine regulates lipid metabolism in adipose tissue through adipokine-mTOR-SIRT1 signaling pathway and bile acid-microbe axis in a finishing pig model. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2024, 16, 158–173. [Google Scholar] [CrossRef]
- Russell, D.W. The enzymes, regulation, and genetics of bile acid synthesis. Annu. Rev. Biochem. 2003, 72, 137–174. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Yu, Y.; Wang, H.; Yang, H.; Tao, F.; Yang, S.; Liu, J.; Li, Z.; Yang, C. Dietary Clostridium butyricum and 25-Hydroxyvitamin D(3) modulate bone metabolism of broilers through the gut-brain axis. Poult. Sci. 2024, 103, 103966. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shi, Y.; Li, J.; Shao, C.; Ma, S.; Shen, C.; Zhao, R. Dietary bile acids improve breast muscle growth in chickens through FXR/IGF2 pathway. Poult. Sci. 2024, 103, 103346. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Kou, Z.; Colon, T.M.; Chen, C.H.; Chen, Y.; Dai, W. Identification of Radil as a Ras binding partner and putative activator. J. Biol. Chem. 2021, 296, 100314. [Google Scholar] [CrossRef]
- Yanagita, K.; Manome, A.; Meng, X.Y.; Hanada, S.; Kanagawa, T.; Tsuchida, T.; Mackie, R.I.; Kamagata, Y. Flow cytometric sorting, phylogenetic analysis and in situ detection of Oscillospira guillermondii, a large, morphologically conspicuous but uncultured ruminal bacterium. Int. J. Syst. Evol. Microbiol. 2003, 53, 1609–1614. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D. Thermal processing of food reduces gut microbiota diversity of the host and triggers adaptation of the microbiota: Evidence from two vertebrates. Microbiome 2018, 6, 99. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, M.; Jiang, H.; Ma, F.; Li, S.; Guo, Y.; Zhu, L.; Shi, C.; Na, R.; Wang, Y.; Zhang, W. Multi-Omics Analysis Revealed the Molecular Mechanisms Affecting Average Daily Gain in Cattle. Int. J. Mol. Sci. 2025, 26, 2343. https://doi.org/10.3390/ijms26052343
Gu M, Jiang H, Ma F, Li S, Guo Y, Zhu L, Shi C, Na R, Wang Y, Zhang W. Multi-Omics Analysis Revealed the Molecular Mechanisms Affecting Average Daily Gain in Cattle. International Journal of Molecular Sciences. 2025; 26(5):2343. https://doi.org/10.3390/ijms26052343
Chicago/Turabian StyleGu, Mingjuan, Hongyu Jiang, Fengying Ma, Shuai Li, Yaqiang Guo, Lin Zhu, Caixia Shi, Risu Na, Yu Wang, and Wenguang Zhang. 2025. "Multi-Omics Analysis Revealed the Molecular Mechanisms Affecting Average Daily Gain in Cattle" International Journal of Molecular Sciences 26, no. 5: 2343. https://doi.org/10.3390/ijms26052343
APA StyleGu, M., Jiang, H., Ma, F., Li, S., Guo, Y., Zhu, L., Shi, C., Na, R., Wang, Y., & Zhang, W. (2025). Multi-Omics Analysis Revealed the Molecular Mechanisms Affecting Average Daily Gain in Cattle. International Journal of Molecular Sciences, 26(5), 2343. https://doi.org/10.3390/ijms26052343