
Effects of Chlortetracycline on Lignin Biosynthesis in Arabidopsis thaliana

Abstract
1. Introduction
2. Results and Discussion
2.1. Toxicity Testing
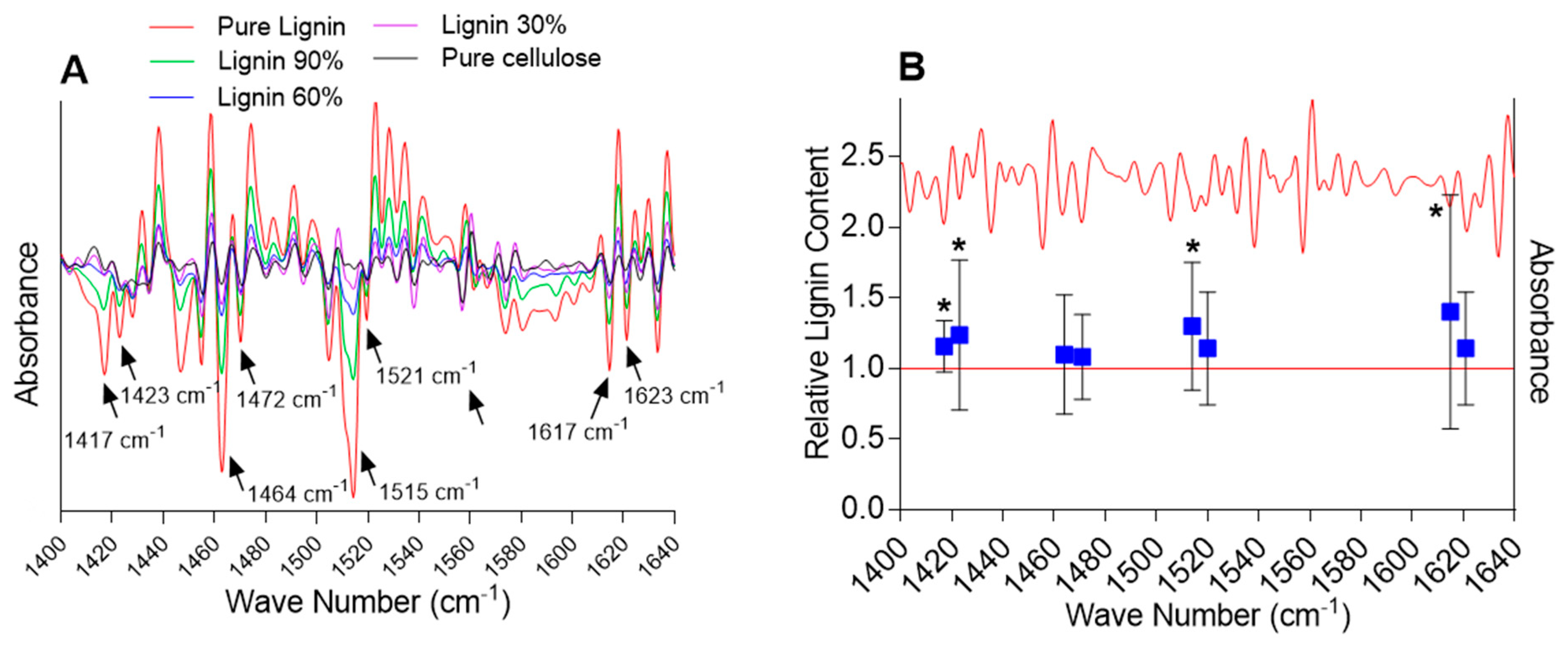
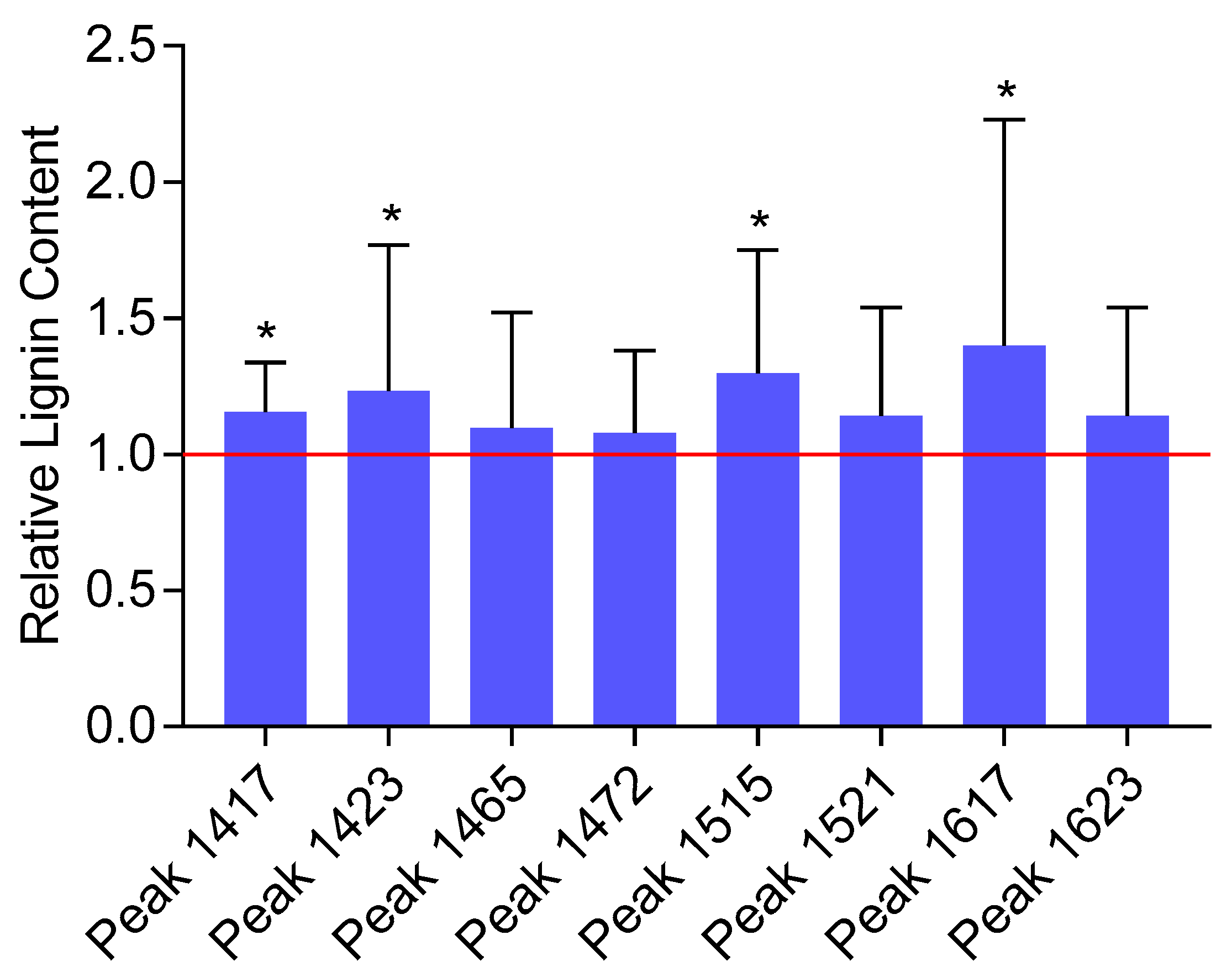
2.2. Analysis of the Lignin Content by FTIR Spectroscopy
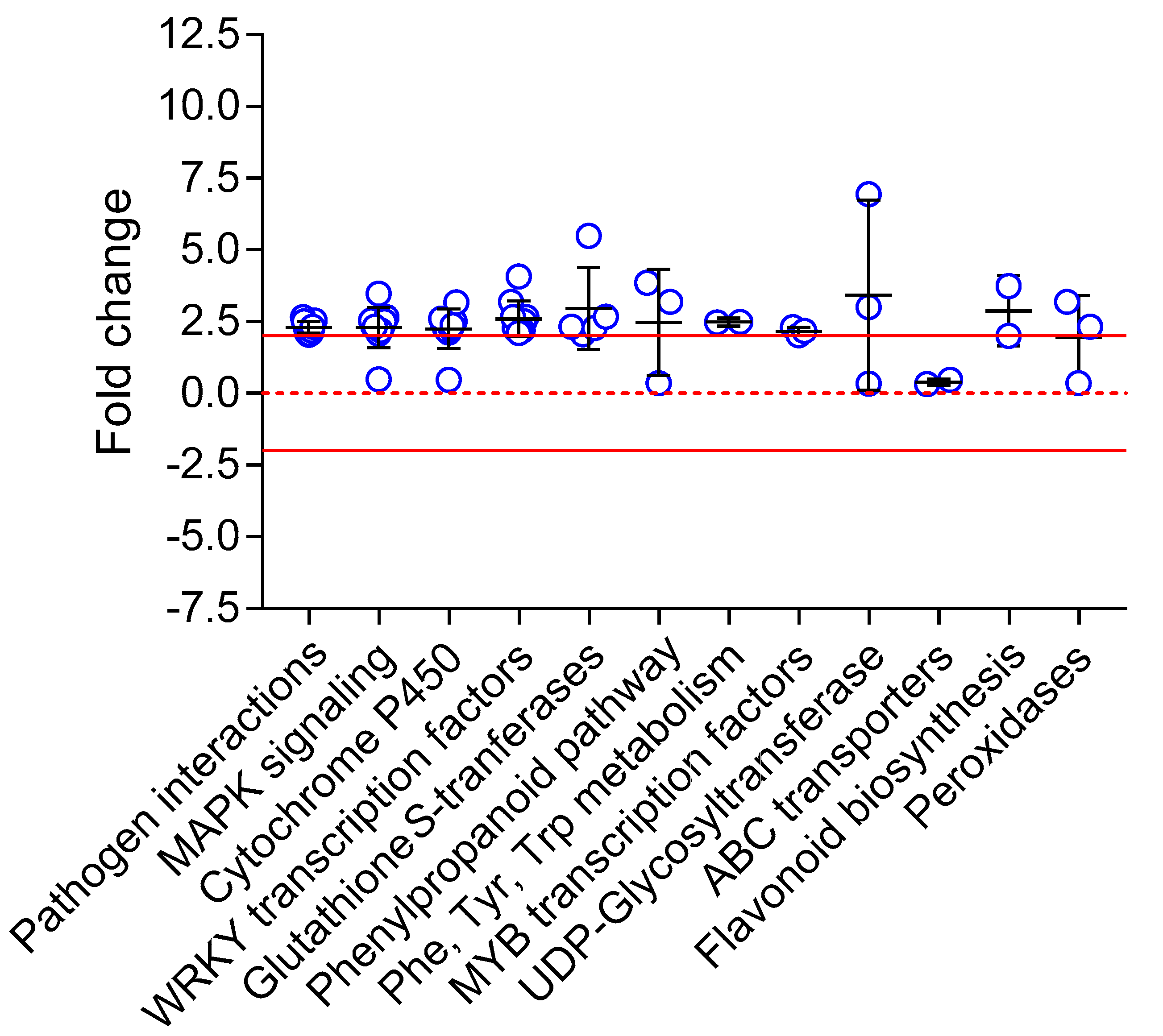
2.3. Gene Expression Analysis
3. Experimental Section
3.1. Chemicals
3.2. Toxicity Tests
3.3. FTIR Analysis
3.4. RNA Sequencing
3.5. Sequencing Data Analysis
3.6. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| FTIR | Fourier-transform infrared spectroscopy |
| ATR | Attenuated total reflection |
| PCB | Polychlorinated biphenyl |
| UVN | Unit vector normalization |
| EMSC | Extended multiplicative scatter correction |
| FDR | False discovery rate |
| DEG | Differentially expressed gene |
| TAIR | The Arabidopsis Information Resource |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
References
- Xie, M.; Zhang, J.; Tschaplinski, T.J.; Tuskan, G.A.; Chen, J.-G.; Muchero, W. Regulation of Lignin Biosynthesis and Its Role in Growth-Defense Tradeoffs. Front. Plant Sci. 2018, 9, 1427. [Google Scholar] [CrossRef]
- Abramson, M.; Shoseyov, O.; Shani, Z. Plant cell wall reconstruction toward improved lignocellulosic production and processability. Plant Sci. 2010, 178, 61–72. [Google Scholar] [CrossRef]
- Schnoor, J.L.; Light, L.A.; McCutcheon, S.C.; Wolfe, N.L.; Carreia, L.H. Phytoremediation of Organic and Nutrient Contaminants. Environ. Sci. Technol. 1995, 29, 318A–323A. [Google Scholar] [CrossRef] [PubMed]
- Salt, D.E.; Smith, R.D.; Raskin, I. Phytoremediation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 643–668. [Google Scholar] [CrossRef]
- Fässler, E.; Robinson, B.H.; Stauffer, W.; Gupta, S.K.; Papritz, A.; Schulin, R. Phytomanagement of Metal-Contaminated Agricultural Land Using Sunflower, Maize and Tobacco. Agric. Ecosyst. Environ. 2010, 136, 49–58. [Google Scholar] [CrossRef]
- Gomes, H.I. Phytoremediation for Bioenergy: Challenges and Opportunities. Environ. Technol. Rev. 2012, 1, 59–66. [Google Scholar] [CrossRef]
- Van Aken, B.; Correa, P.A.; Schnoor, J.L. Phytoremediation of Polychlorinated Biphenyls: New Trends and Promises. Environ. Sci. Technol. 2010, 44, 2767–2776. [Google Scholar] [CrossRef]
- Jing, R.; Fusi, S.; Kjellerup, B.V. Remediation of Polychlorinated Biphenyls (PCBs) in Contaminated Soils and Sediment: State of Knowledge and Perspectives. Front. Environ. Sci. 2018, 6, 79. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q. Cadmium Tolerance and Accumulation in Eight Potential Energy Crops. Biotechnol. Adv. 2009, 27, 555–561. [Google Scholar] [CrossRef]
- Freitas, E.; Salgado, J.; Alnoch, R.; Contato, A.; Habermann, E.; Michelin, M.; Martínez, C.; Polizeli, M. Challenges of Biomass Utilization for Bioenergy in a Climate Change Scenario. Biology 2021, 10, 1277. [Google Scholar] [CrossRef]
- Jing, Y.; Guo, Y.; Xia, Q.; Liu, X.; Wang, Y. Catalytic Production of Value-Added Chemicals and Liquid Fuels from Lignocellulosic Biomass | Elsevier Enhanced Reader. Chem 2019, 5, 22520–22546. [Google Scholar] [CrossRef]
- Liu, C.-J.; Eudes, A. Lignin Synthesis and Bioengineering Approaches toward Lignin Modification. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Nageswara-Rao, M.; Soneji, J.R.; Kwit, C.; Stewart, C.N. Advances in Biotechnology and Genomics of Switchgrass. Biotechnol. Biofuels 2013, 6, 77. [Google Scholar] [CrossRef]
- Schneider, W.D.H.; Fontana, R.C.; Baudel, H.M.; de Siqueira, F.G.; Rencoret, J.; Gutiérrez, A.; de Eugenio, L.I.; Prieto, A.; Martínez, M.J.; Martínez, Á.T.; et al. Lignin Degradation and Detoxification of Eucalyptus Wastes by On-Site Manufacturing Fungal Enzymes to Enhance Second-Generation Ethanol Yield. Appl. Energy 2020, 262, 114493. [Google Scholar] [CrossRef]
- Herbette, S.; Taconnat, L.; Hugouvieux, V.; Piette, L.; Magniette, M.-L.M.; Cuine, S.; Auroy, P.; Richaud, P.; Forestier, C.; Bourguignon, J.; et al. Genome-Wide Transcriptome Profiling of the Early Cadmium Response of Arabidopsis Roots and Shoots. Biochimie 2006, 88, 1751–1765. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Ali, M.B.; Singh, N.; Shohael, A.M.; Hahn, E.J.; Paek, K.-Y. Phenolics Metabolism and Lignin Synthesis in Root Suspension Cultures of Panax Ginseng in Response to Copper Stress. Plant Sci. 2006, 171, 147–154. [Google Scholar] [CrossRef]
- Chandran, D.; Sharopova, N.; Ivashuta, S.; Gantt, J.S.; Vandenbosch, K.A.; Samac, D.A. Transcriptome Profiling Identified Novel Genes Associated with Aluminum Toxicity, Resistance and Tolerance in Medicago Truncatula. Planta 2008, 228, 151–166. [Google Scholar] [CrossRef]
- Zamcho, F.; Newborn, A.; Karamat, A.; Tehrani, R.; Pleshko, N.; Aken, B.V. Effects of Polychlorinated Biphenyls on Lignin Biosynthesis in Arabidopsis thaliana. ACS Agric. Sci. Technol. 2021, 1, 202–210. [Google Scholar] [CrossRef]
- Han, X.; Zhao, Y.; Chen, Y.; Xu, J.; Jiang, C.; Wang, X.; Zhuo, R.; Lu, M.-Z.; Zhang, J. Lignin Biosynthesis and Accumulation in Response to Abiotic Stresses in Woody Plants. For. Res. 2022, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.S.; Gupta, R.K.; Kim, I.-W.; Lee, J.-K. Coriolus Versicolor Laccase-Based Inorganic Protein Hybrid Synthesis for Application in Biomass Saccharification to Enhance Biological Production of Hydrogen and Ethanol. Enzym. Microb. Technol. 2023, 170, 110301. [Google Scholar] [CrossRef] [PubMed]
- Jukes, T.H. Some Historical Notes on Chlortetracycline. Rev. Infect. Dis. 1985, 7, 702–707. [Google Scholar] [CrossRef]
- Lee, C.; Jeong, S.; Ju, M.; Kim, J.Y. Fate of Chlortetracycline Antibiotics during Anaerobic Degradation of Cattle Manure. J. Hazard. Mater. 2020, 386, 121894. [Google Scholar] [CrossRef]
- Tian, Y.; Wei, L.; Yin, Z.; Feng, L.; Zhang, L.; Liu, Y.; Zhang, L. Photosensitization Mechanism of Algogenic Extracellular Organic Matters (EOMs) in the Photo-Transformation of Chlortetracycline: Role of Chemical Constituents and Structure. Water Res. 2019, 164, 114940. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xue, Y.; Wang, H.; Chen, Y. Effects of Chlortetracycline on the Growth of Eggplant and Associated Rhizosphere Bacterial Communities. Sustainability 2023, 15, 14593. [Google Scholar] [CrossRef]
- Kumar, K.; Gupta, S.C.; Baidoo, S.K.; Chander, Y.; Rosen, C.J. Antibiotic Uptake by Plants from Soil Fertilized with Animal Manure. J. Environ. Qual. 2005, 34, 2082–2085. [Google Scholar] [CrossRef]
- Liu, F.; Ying, G.-G.; Tao, R.; Zhao, J.-L.; Yang, J.-F.; Zhao, L.-F. Effects of Six Selected Antibiotics on Plant Growth and Soil Microbial and Enzymatic Activities. Environ. Pollut. 2009, 157, 1636–1642. [Google Scholar] [CrossRef]
- Sikorski, Ł.; Bęś, A.; Karetko-Sikorska, E.; Truszkowski, W.; Tomaszewska, K. Ion-Exchange Chromatography in the Assessment of Environmental Pollution with Chlortetracycline. Aquat. Toxicol. 2024, 277, 107142. [Google Scholar] [CrossRef]
- Batchelder, A.R. Chlortetracycline and Oxytetracycline Effects on Plant Growth and Development in Liquid Cultures. J. Environ. Qual. 1981, 10, 515–518. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, J.; Gao, L.; Kong, F.; Shen, G.; Wang, R.; Gao, J.; Zhang, J. The Effects of Tetracycline Residues on the Microbial Community Structure of Tobacco Soil in Pot Experiment. Sci. Rep. 2020, 10, 8804. [Google Scholar] [CrossRef]
- Ahmed, M.B.M.; Rajapaksha, A.U.; Lim, J.E.; Vu, N.T.; Kim, I.S.; Kang, H.M.; Lee, S.S.; Ok, Y.S. Distribution and Accumulative Pattern of Tetracyclines and Sulfonamides in Edible Vegetables of Cucumber, Tomato, and Lettuce. J. Agric. Food Chem. 2015, 63, 398–405. [Google Scholar] [CrossRef]
- Albero, B.; Tadeo, J.L.; Miguel, E.; Pérez, R.A. Rapid Determination of Antibiotic Residues in Cereals by Liquid Chromatography Triple Mass Spectrometry. Anal. Bioanal. Chem. 2019, 411, 6129–6139. [Google Scholar] [CrossRef] [PubMed]
- Bowman, S.M.; Drzewiecki, K.E.; Mojica, E.-R.E.; Zielinski, A.M.; Siegel, A.; Aga, D.S.; Berry, J.O. Toxicity and Reductions in Intracellular Calcium Levels Following Uptake of a Tetracycline Antibiotic in Arabidopsis. Environ. Sci. Technol. 2011, 45, 8958–8964. [Google Scholar] [CrossRef]
- Hillis, D.G.; Fletcher, J.; Solomon, K.R.; Sibley, P.K. Effects of Ten Antibiotics on Seed Germination and Root Elongation in Three Plant Species. Arch. Environ. Contam. Toxicol. 2011, 60, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Minden, V.; Deloy, A.; Volkert, A.M.; Leonhardt, S.D.; Pufal, G. Antibiotics Impact Plant Traits, Even at Small Concentrations. AoB PLANTS 2017, 9, plx010. [Google Scholar] [CrossRef]
- Cheong, M.S.; Yoon, Y.-E.; Kim, J.W.; Hong, Y.K.; Kim, S.C.; Lee, Y.B. Chlortetracycline Inhibits Seed Germination and Seedling Growth in Brassica Campestris by Disrupting H2O2 Signaling. Appl. Biol. Chem. 2020, 63, 1. [Google Scholar] [CrossRef]
- Lupoi, J.S.; Singh, S.; Simmons, B.A.; Henry, R.J. Assessment of Lignocellulosic Biomass Using Analytical Spectroscopy: An Evolution to High-Throughput Techniques. Bioenergy Res. 2014, 7, 1–23. [Google Scholar] [CrossRef]
- Lee, B.-J.; Zhou, Y.; Lee, J.S.; Shin, B.K.; Seo, J.-A.; Lee, D.; Kim, Y.-S.; Choi, H.-K. Discrimination and Prediction of the Origin of Chinese and Korean Soybeans Using Fourier Transform Infrared Spectrometry (FT-IR) with Multivariate Statistical Analysis. PLoS ONE 2018, 13, e0196315. [Google Scholar] [CrossRef]
- Derkacheva, O.; Sukhov, D. Investigation of Lignins by FTIR Spectroscopy. Macromol. Symp. 2008, 265, 61–68. [Google Scholar] [CrossRef]
- Boeriu, C.G.; Bravo, D.; Gosselink, R.J.A.; Van Dam, J.E.G. Characterisation of Structure-Dependent Functional Properties of Lignin with Infrared Spectroscopy. Ind. Crops Prod. 2004, 20, 205–218. [Google Scholar] [CrossRef]
- Zhou, G.; Taylor, G.; Polle, A. FTIR-ATR-Based Prediction and Modelling of Lignin and Energy Contents Reveals Independent Intra-Specific Variation of These Traits in Bioenergy Poplars. Plant Methods 2011, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Javier-Astete, R.; Jimenez-Davalos, J.; Zolla, G. Determination of Hemicellulose, Cellulose, Holocellulose and Lignin Content Using FTIR in Calycophyllum spruceanum (Benth.) K. Schum. and Guazuma crinita Lam. PLoS ONE 2021, 16, e0256559. [Google Scholar] [CrossRef]
- Dokken, K.; Davis, L.; Erickson, L.; Castro Diaz, S. Fourier Transform Infrared Spectroscopy as a Tool to Monitor Changes in Plant Structure in Response to Soil Contaminants. Proc. Waste Res. Technol. 2011. [Google Scholar]
- Rieppo, L.; Saarakkala, S.; Närhi, T.; Helminen, H.J.; Jurvelin, J.S.; Rieppo, J. Application of Second Derivative Spectroscopy for Increasing Molecular Specificity of Fourier Transform Infrared Spectroscopic Imaging of Articular Cartilage. Osteoarthr. Cartil. 2012, 20, 451–459. [Google Scholar] [CrossRef]
- Sharma, S.; Uttam, K.N. Nondestructive and Rapid Probing of Biochemical Response of Arsenic Stress on the Leaves of Wheat Seedlings Using Attenuated Total Reflectance Fourier Transform Infrared Spectroscopy. Anal. Lett. 2019, 52, 268–287. [Google Scholar] [CrossRef]
- Yu, S.; Sheng, L.; Zhang, C.; Deng, H. Physiological Response of Arundo Donax to Cadmium Stress by Fourier Transform Infrared Spectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 198, 88–91. [Google Scholar] [CrossRef]
- Karmanov, A.P.; Derkacheva, O.Y. Application of Fourier Transform Infrared Spectroscopy for the Study of Lignins of Herbaceous Plants. Russ. J. Bioorg. Chem. 2013, 39, 677–685. [Google Scholar] [CrossRef]
- Shi, Z.; Xu, G.; Deng, J.; Dong, M.; Murugadoss, V.; Liu, C.; Shao, Q.; Wu, S.; Guo, Z. Structural Characterization of Lignin from D. sinicus by FTIR and NMR Techniques. Green Chem. Lett. Rev. 2019, 12, 235–243. [Google Scholar] [CrossRef]
- Xu, F.; Yu, J.; Tesso, T.; Dowell, F.; Wang, D. Qualitative and Quantitative Analysis of Lignocellulosic Biomass Using Infrared Techniques: A Mini-Review. Appl. Energy 2013, 104, 801–809. [Google Scholar] [CrossRef]
- Jesus, E.; Franca, T.; Calvani, C.; Lacerda, M.; Gonçalves, D.; Oliveira, S.L.; Marangoni, B.; Cena, C. Making Wood Inspection Easier: FTIR Spectroscopy and Machine Learning for Brazilian Native Commercial Wood Species Identification. RSC Adv. 2024, 14, 7283–7289. [Google Scholar] [CrossRef] [PubMed]
- Elobeid, M.; Göbel, C.; Feussner, I.; Polle, A. Cadmium Interferes with Auxin Physiology and Lignification in Poplar. J. Exp. Bot. 2012, 63, 1413–1421. [Google Scholar] [CrossRef]
- Broda, M.; Yelle, D.J.; Serwańska, K. Bioethanol Production from Lignocellulosic Biomass—Challenges and Solutions. Molecules 2022, 27, 8717. [Google Scholar] [CrossRef] [PubMed]
- Eswaran, S.C.; Subramaniam, S.; Sanyal, U.; Rallo, R.; Zhang, X. Molecular Structural Dataset of Lignin Macromolecule Elucidating Experimental Structural Compositions. Sci. Data 2022, 9, 647. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.; Engle, N.; Tschaplinski, T.; Pu, Y.; Ragauskas, A.J. Variable Lignin Structure Revealed in Populus Leaves. RSC Adv. 2023, 13, 20187–20197. [Google Scholar] [CrossRef]
- Hori, C.; Yu, X.; Mortimer, J.C.; Sano, R.; Matsumoto, T.; Kikuchi, J.; Demura, T.; Ohtani, M. Impact of Abiotic Stress on the Regulation of Cell Wall Biosynthesis in Populus trichocarpa. Plant Biotechnol. 2020, 37, 273–283. [Google Scholar] [CrossRef]
- Lima, R.B.; Dos Santos, T.B.; Vieira, L.G.E.; Ferrarese, M.D.L.L.; Ferrarese-Filho, O.; Donatti, L.; Boeger, M.R.T.; Petkowicz, C.L.D.O. Heat Stress Causes Alterations in the Cell-Wall Polymers and Anatomy of Coffee Leaves (Coffea arabica L.). Carbohydr. Polym. 2013, 93, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Daniel, B.; Pavkov-Keller, T.; Steiner, B.; Dordic, A.; Gutmann, A.; Nidetzky, B.; Sensen, C.W.; van der Graaff, E.; Wallner, S.; Gruber, K.; et al. Oxidation of Monolignols by Members of the Berberine Bridge Enzyme Family Suggests a Role in Plant Cell Wall Metabolism. J. Biol. Chem. 2015, 290, 18770–18781. [Google Scholar] [CrossRef]
- Jiang, D.; Yan, S. Effects of Cd, Zn, or Pb Stress in Populus Alba Berolinensis on the Antioxidant, Detoxifying, and Digestive Enzymes of Lymantria Dispar. Environ. Entomol. 2018, 47, 1323–1328. [Google Scholar] [CrossRef]
- Sharma, Y.K.; Davis, K.R. Ozone-Induced Expression of Stress-Related Genes in Arabidopsis thaliana. Plant Physiol. 1994, 105, 1089–1096. [Google Scholar] [CrossRef]
- Eckey-Kaltenbach, H.; Ernst, D.; Heller, W.; Jr, H.S. Biochemical Plant Responses to Ozone (IV. Cross-Induction of Defensive Pathways in Parsley (Petroselinum crispum L.) Plants). Plant Physiol. 1994, 104, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Foley, R.C.; Oñate-Sánchez, L. Transcription Factors in Plant Defense and Stress Responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.H.Y.; Tyler, B.M. Mechanisms and Evolution of Virulence in Oomycetes. Annu. Rev. Phytopathol. 2012, 50, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.J.; Urwin, P.E. The Interaction of Plant Biotic and Abiotic Stresses: From Genes to the Field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef]
- Ramel, F.; Sulmon, C.; Serra, A.-A.; Gouesbet, G.; Couée, I. Xenobiotic Sensing and Signalling in Higher Plants. J. Exp. Bot. 2012, 63, 3999–4014. [Google Scholar] [CrossRef]
- Nguyen, D.; Rieu, I.; Mariani, C.; van Dam, N.M. How Plants Handle Multiple Stresses: Hormonal Interactions Underlying Responses to Abiotic Stress and Insect Herbivory. Plant Mol. Biol. 2016, 91, 727–740. [Google Scholar] [CrossRef]
- Berens, M.L.; Wolinska, K.W.; Spaepen, S.; Ziegler, J.; Nobori, T.; Nair, A.; Krüler, V.; Winkelmüller, T.M.; Wang, Y.; Mine, A.; et al. Balancing Trade-Offs between Biotic and Abiotic Stress Responses through Leaf Age-Dependent Variation in Stress Hormone Cross-Talk. Proc. Natl. Acad. Sci. USA 2019, 116, 2364–2373. [Google Scholar] [CrossRef]
- Swindell, W.R. The Association Among Gene Expression Responses to Nine Abiotic Stress Treatments in Arabidopsis thaliana. Genetics 2006, 174, 1811–1824. [Google Scholar] [CrossRef]
- Sandermann, H. Higher Plant Metabolism of Xenobiotics: The “green Liver” Concept. Pharmacogenetics 1994, 4, 225–241. [Google Scholar] [CrossRef]
- Hernández Estévez, I.; Rodríguez Hernández, M. “Plant Glutathione S-Transferases: An Overview”. Plant Gene 2020, 23, 100233. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF Transcription Factor Regulatory Networks in Hormone and Abiotic Stress Responses in Arabidopsis. Front. Plant Sci. 2019, 10, 228. [Google Scholar] [CrossRef]
- Liu, J.-X.; Howell, S.H. bZIP28 and NF-Y Transcription Factors Are Activated by ER Stress and Assemble into a Transcriptional Complex to Regulate Stress Response Genes in Arabidopsis. Plant Cell 2010, 22, 782–796. [Google Scholar] [CrossRef]
- Javed, T.; Gao, S.-J. WRKY Transcription Factors in Plant Defense. Trends Genet. 2023, 39, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Ambawat, S.; Sharma, P.; Yadav, N.R.; Yadav, R.C. MYB Transcription Factor Genes as Regulators for Plant Responses: An Overview. Physiol. Mol. Biol. Plants 2013, 19, 307–321. [Google Scholar] [CrossRef]
- Subramanian, S.; Schnoor, J.L.; Van Aken, B. Effects of Polychlorinated Biphenyls (PCBs) and Their Hydroxylated Metabolites (OH-PCBs) on Arabidopsis thaliana. Environ. Sci. Technol. 2017, 51, 7263–7270. [Google Scholar] [CrossRef] [PubMed]
- Rivero, L.; Scholl, R.; Holomuzki, N.; Crist, D.; Grotewold, E.; Brkljacic, J. Handling Arabidopsis Plants: Growth, Preservation of Seeds, Transformation, and Genetic Crosses. In Arabidopsis Protocols; Sanchez-Serrano, J.J., Salinas, J., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2014; Volume 1062, pp. 3–25. ISBN 978-1-62703-579-8. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics Enrichment Tools: Paths toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Evangelou, M.W.H.; Conesa, H.M.; Robinson, B.H.; Schulin, R. Biomass Production on Trace Element–Contaminated Land: A Review. Environ. Eng. Sci. 2012, 29, 823–839. [Google Scholar] [CrossRef]
- Fatma, S.; Hameed, A.; Noman, M.; Ahmed, T.; Shahid, M.; Tariq, M.; Sohail, I.; Tabassum, R. Lignocellulosic Biomass: A Sustainable Bioenergy Source for the Future. Protein Pept. Lett. 2018, 25, 148–163. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Biological Process Term | Total Genes | # DEGs | # DEGs Expected | Fold Enrichment | Bonferroni p-Value |
|---|---|---|---|---|---|
| Cellular response to sulfur starvation (GO:0010438) | 7 | 4 | 0.13 | 29.68 | 1.25 × 10−2 |
| Cellular response to iron ion starvation (GO:0010106) | 8 | 4 | 0.15 | 25.97 | 2.46 × 10−2 |
| Priming of cellular response to stress (GO:0080136) | 13 | 6 | 0.25 | 23.97 | 2.08 × 10−4 |
| Systemic acquired resistance (GO:0009627) | 67 | 13 | 1.29 | 10.08 | 1.17× 10−6 |
| Cellular response to decreased oxygen levels (GO:0036294) | 239 | 28 | 4.6 | 6.08 | 7.25 × 10−11 |
| Innate immune response (GO:0045087) | 183 | 16 | 3.52 | 4.54 | 1.57 × 10−3 |
| Response to reactive oxygen species (GO:0000302) | 166 | 14 | 3.2 | 4.38 | 1.21 × 10−2 |
| Defense response to bacterium (GO:0042742) | 389 | 31 | 7.49 | 4.14 | 8.96 × 10−8 |
| Regulation of response to stress (GO:0080134) | 474 | 31 | 9.13 | 3.4 | 1.13 × 10−5 |
| Response to external stimulus (GO:0009605) | 1410 | 83 | 27.15 | 3.06 | 2.07 × 10−16 |
| Response to osmotic stress (GO:0006970) | 579 | 34 | 11.15 | 3.05 | 3.01 × 10−5 |
| Response to salt stress (GO:0009651) | 482 | 28 | 9.28 | 3.02 | 7.52 × 10−4 |
| Cellular response to chemical stimulus (GO:0070887) | 1133 | 62 | 21.81 | 2.84 | 4.19 × 10−10 |
| Response to chemical (GO:0042221) | 2743 | 134 | 52.81 | 2.54 | 4.21 × 10−21 |
| Response to abiotic stimulus (GO:0009628) | 2257 | 93 | 43.46 | 2.14 | 5.86 × 10−9 |
| Macromolecule biosynthetic process (GO:0009059) | 2351 | 13 | 45.27 | 0.29 | 2.69 × 10−5 |
| Gene expression (GO:0010467) | 1924 | 8 | 37.04 | 0.22 | 2.01 × 10−5 |
| Nucleic acid metabolic process (GO:0090304) | 1870 | 7 | 36 | 0.19 | 7.49 × 10−6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Newborn, A.; Karamat, A.; Van Aken, B. Effects of Chlortetracycline on Lignin Biosynthesis in Arabidopsis thaliana. Int. J. Mol. Sci. 2025, 26, 2288. https://doi.org/10.3390/ijms26052288
Newborn A, Karamat A, Van Aken B. Effects of Chlortetracycline on Lignin Biosynthesis in Arabidopsis thaliana. International Journal of Molecular Sciences. 2025; 26(5):2288. https://doi.org/10.3390/ijms26052288
Chicago/Turabian StyleNewborn, Aaron, Ayesha Karamat, and Benoit Van Aken. 2025. "Effects of Chlortetracycline on Lignin Biosynthesis in Arabidopsis thaliana" International Journal of Molecular Sciences 26, no. 5: 2288. https://doi.org/10.3390/ijms26052288
APA StyleNewborn, A., Karamat, A., & Van Aken, B. (2025). Effects of Chlortetracycline on Lignin Biosynthesis in Arabidopsis thaliana. International Journal of Molecular Sciences, 26(5), 2288. https://doi.org/10.3390/ijms26052288