
Re-Sensitization of Resistant Ovarian Cancer SKOV3/CDDP Cells to Cisplatin by Curcumin Pre-Treatment
 , , ,
, , ,
Abstract
1. Introduction
2. Results
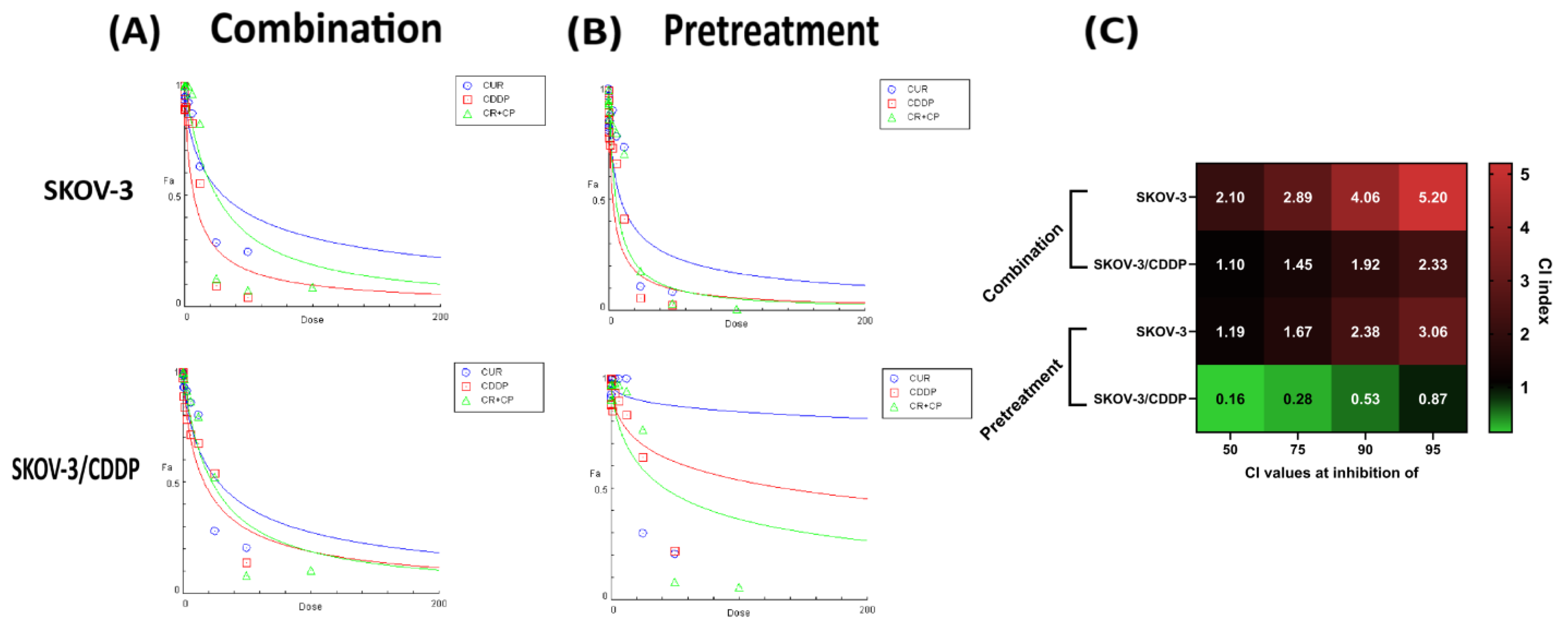
2.1. Combination CUR and CDDP Has an Antagonistic Cytotoxic Effect on SKOV-3 and SKOV-3/CDDP Cell Lines, While CUR Pre-Treatment Has a Synergistic Cytotoxic Effect Only on SKOV-3/CDDP Cells
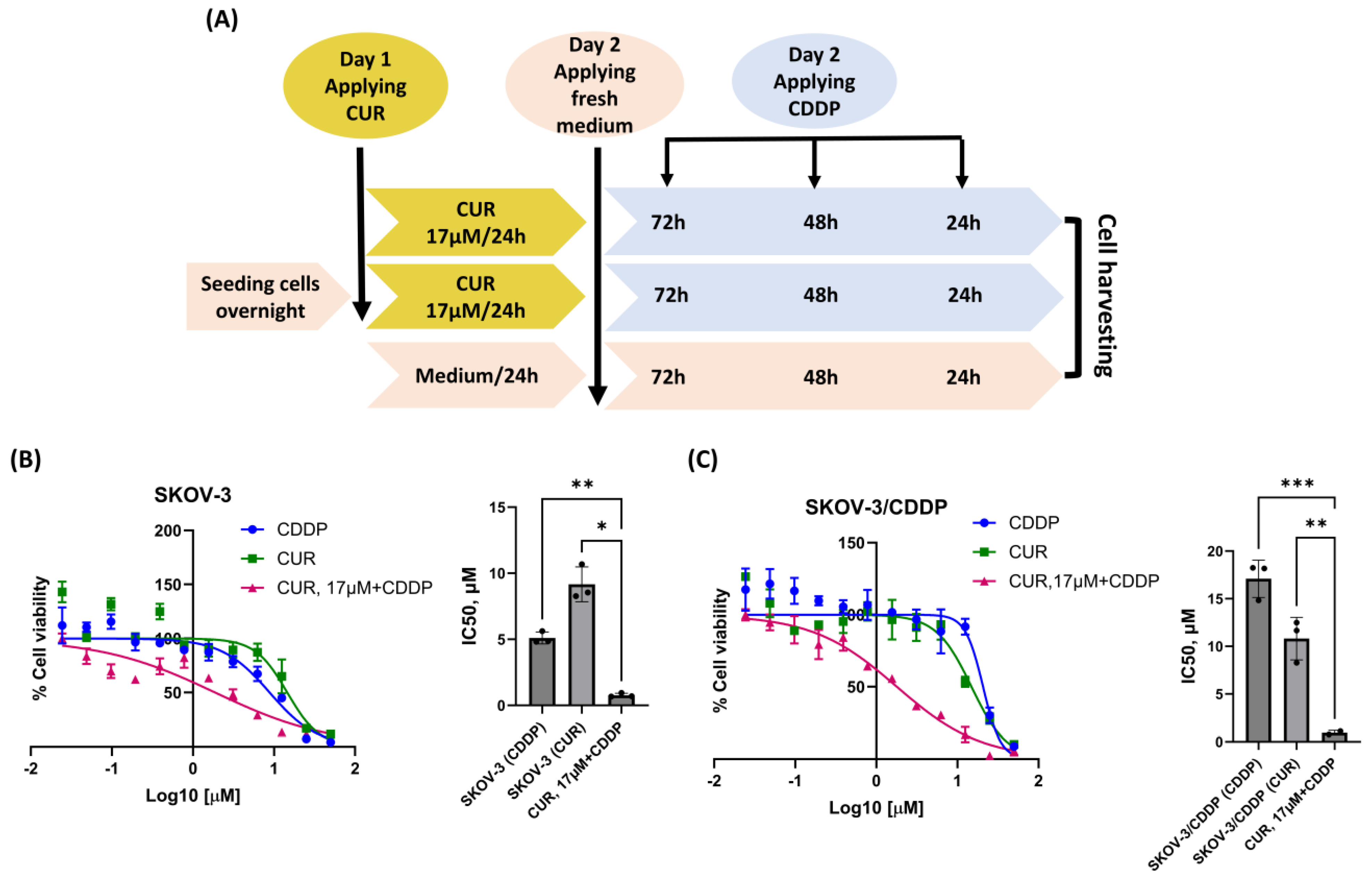
2.2. CUR Pre-Treatment Has a Synergistic Cytotoxic Effect in the SKOV-3/CDDP Cell Line
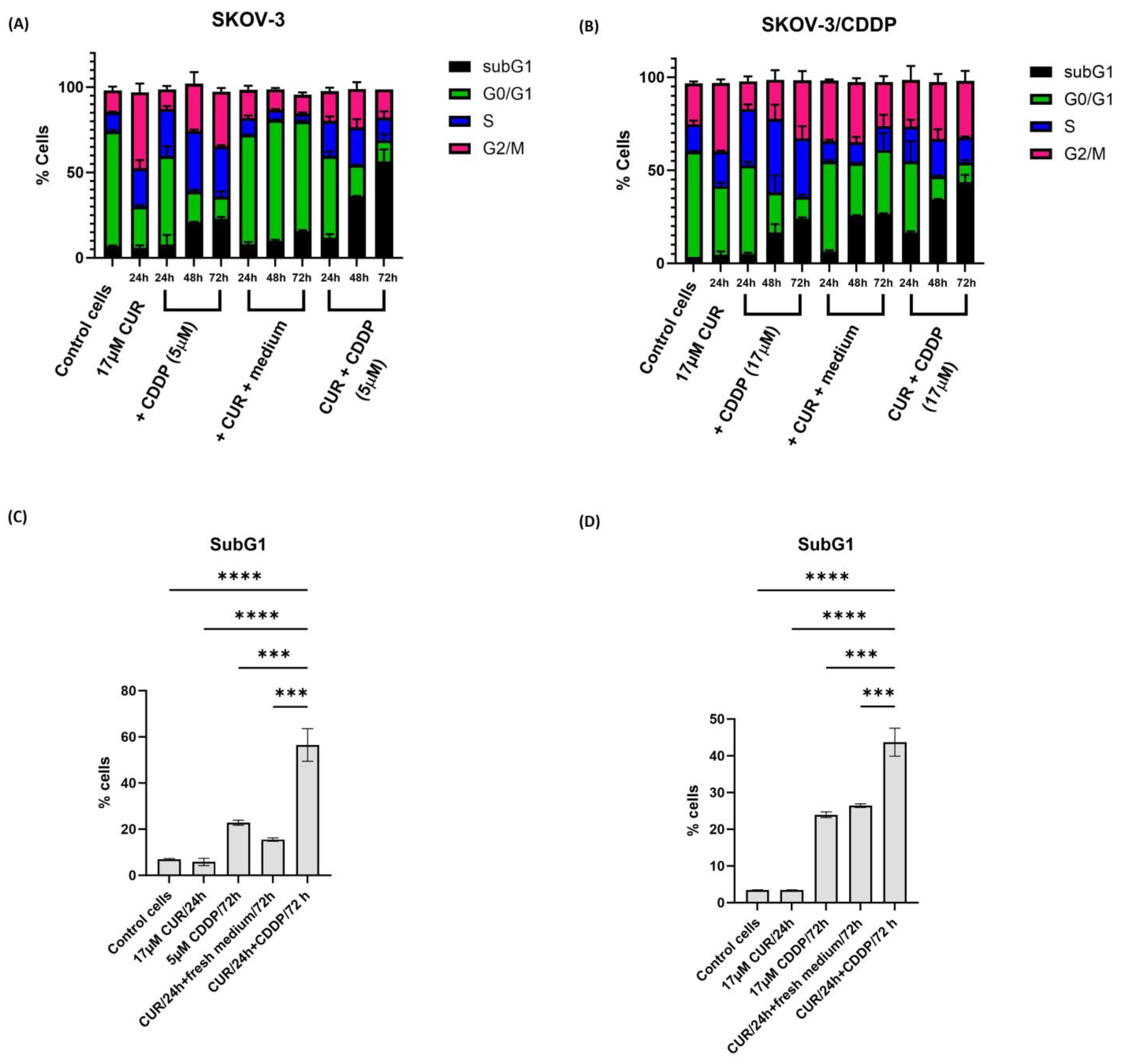
2.3. CUR Pre-Treatment Modulates Cell Cycle Distribution in SKOV-3 and SKOV-3/CDDP Cell Lines
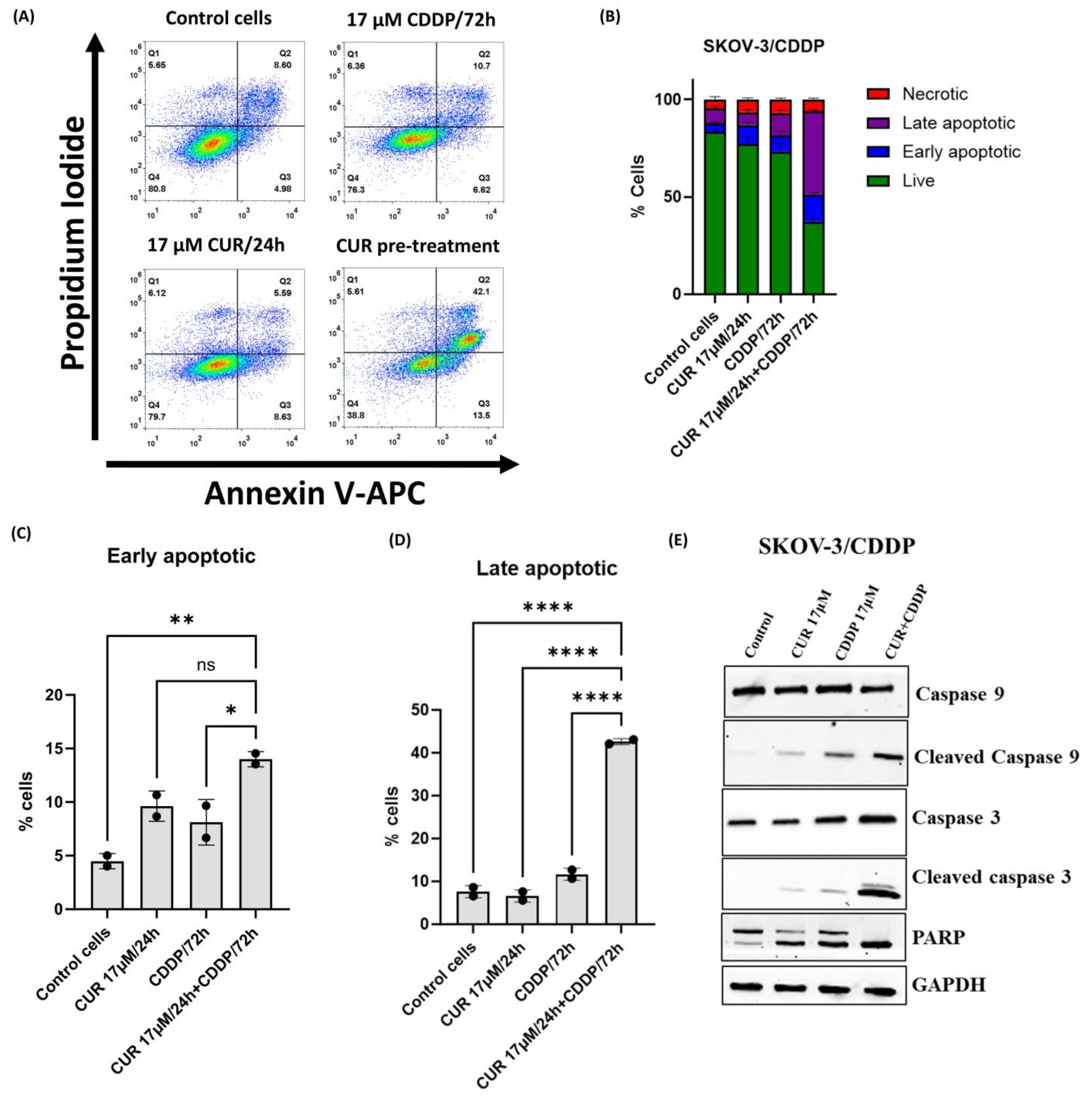
2.4. CUR Pre-Treatment Induces Apoptosis in SKOV-3/CDDP Cell Line
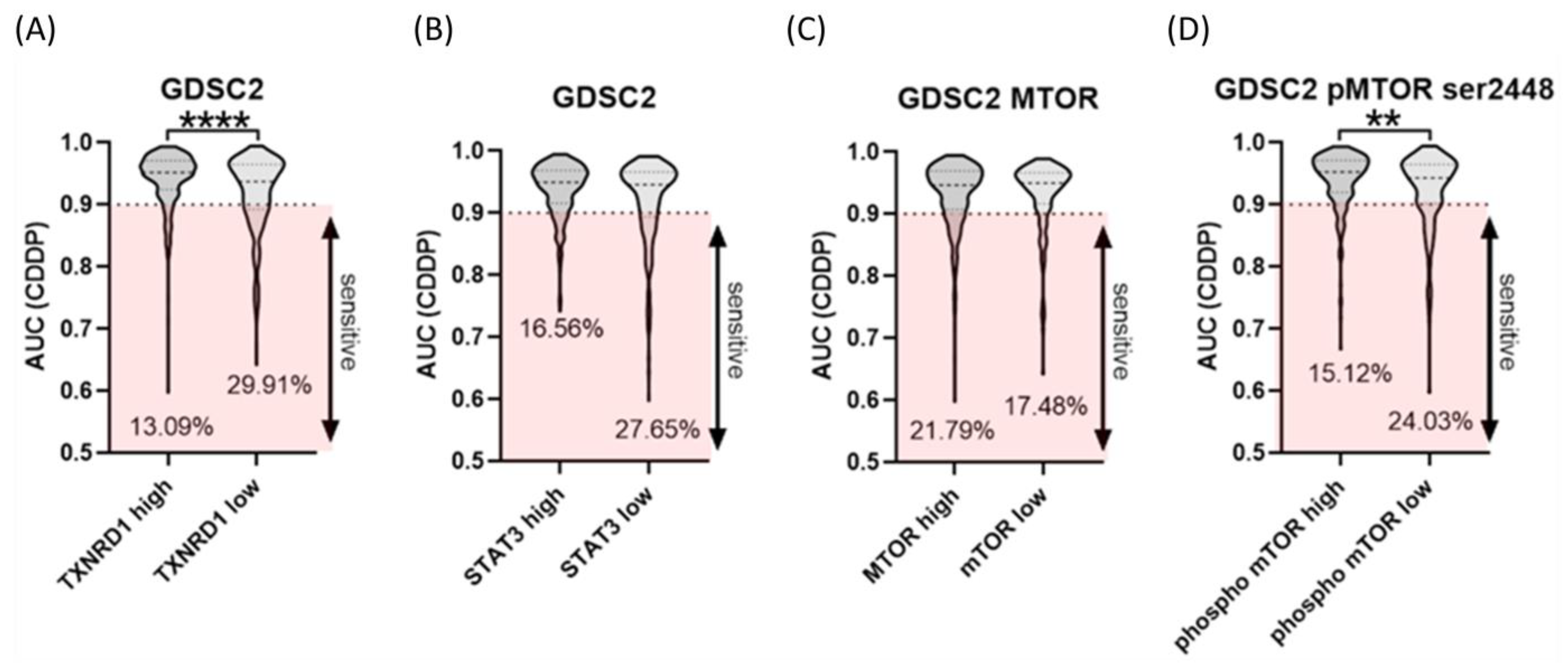
2.5. High Expression of TXNRD1 as Well Phosphorylated mTOR Level Correlate with Resistance to CDDP in Cancer Cell Lines
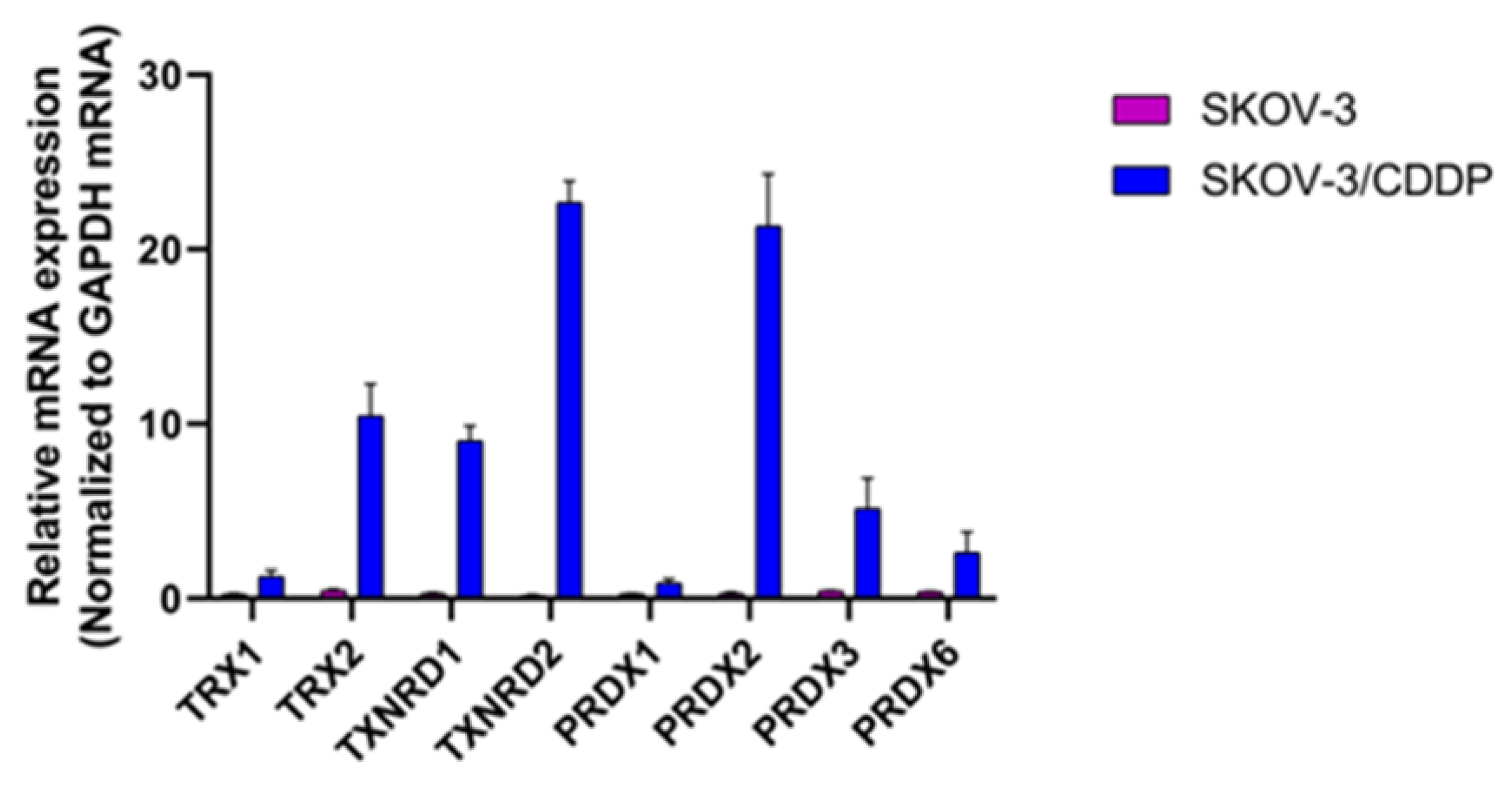
2.6. High Expression of Thioredoxin System and Peroxiredoxins Genes Could Lead to CDDP Resistance in SKOV-3/CDDP
2.7. CUR Pre-Treatment Inhibits TRXR1, mTOR, and STAT3 in the SKOV-3/CDDP Cell Line
2.8. Intracellular CUR Accumulation at Different Cell Densities
3. Discussion
4. Materials and Methods
4.1. Reagents and Drugs
4.2. Cell Lines and Culture Conditions
4.3. Evaluation of Combination and Pre-Treatment Methods
4.4. CUR Pre-Treatment Cytotoxicity Against SKOV-3 and SKOV-3/CDDP Cell Lines
4.5. Cell Cycle Assay
4.6. Apoptotic Programmed Cell Death Analysis
4.7. Data Collection and Drug (CDDP) Screening
4.8. RNA Isolation and Real-Time Quantitative RT-PCR
4.9. Western Blotting Assay
4.10. Intracellular Accumulation
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Momenimovahed, Z.; Tiznobaik, A.; Taheri, S.; Salehiniya, H. Ovarian Cancer in the World: Epidemiology and Risk Factors. Int. J. Women’s Health 2019, 11, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian Cancer Statistics, 2018. CA A Cancer J Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Cormio, G.; Rossi, C.; Cazzolla, A.; Resta, L.; Loverro, G.; Greco, P.; Selvaggi, L. Distant Metastases in Ovarian Carcinoma. Int. J. Gynecol. Cancer 2003, 13, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Elias, K.M.; Guo, J.; Bast, R.C. Early Detection of Ovarian Cancer. Hematol./Oncol. Clin. N. Am. 2018, 32, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Borkar, P.; Bhandari, P.; Yadav, S.; Prabhu, A. Cisplatin Resistance in Ovarian Cancer: Classical Outlook and Newer Perspectives. Biomed. Pharmacol. J. 2021, 14, 1993–2005. [Google Scholar] [CrossRef]
- Zoń, A.; Bednarek, I. Cisplatin in Ovarian Cancer Treatment—Known Limitations in Therapy Force New Solutions. Int. J. Mol. Sci. 2023, 24, 7585. [Google Scholar] [CrossRef]
- Morelli, A.P.; Tortelli, T.C.; Mancini, M.C.S.; Pavan, I.C.B.; Silva, L.G.S.; Severino, M.B.; Granato, D.C.; Pestana, N.F.; Ponte, L.G.S.; Peruca, G.F.; et al. STAT3 Contributes to Cisplatin Resistance, Modulating EMT Markers, and the mTOR Signaling in Lung Adenocarcinoma. Neoplasia 2021, 23, 1048–1058. [Google Scholar] [CrossRef]
- Tripathy, S.; Verma, D.K.; Thakur, M.; Patel, A.R.; Srivastav, P.P.; Singh, S.; Gupta, A.K.; Chávez-González, M.L.; Aguilar, C.N.; Chakravorty, N.; et al. Curcumin Extraction, Isolation, Quantification and Its Application in Functional Foods: A Review With a Focus on Immune Enhancement Activities and COVID-19. Front. Nutr. 2021, 8, 747956. [Google Scholar] [CrossRef]
- Xu, X.-Y.; Meng, X.; Li, S.; Gan, R.-Y.; Li, Y.; Li, H.-B. Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives. Nutrients 2018, 10, 1553. [Google Scholar] [CrossRef]
- Abd. Wahab, N.A.; H. Lajis, N.; Abas, F.; Othman, I.; Naidu, R. Mechanism of Anti-Cancer Activity of Curcumin on Androgen-Dependent and Androgen-Independent Prostate Cancer. Nutrients 2020, 12, 679. [Google Scholar] [CrossRef]
- Prasad, S.; DuBourdieu, D.; Srivastava, A.; Kumar, P.; Lall, R. Metal–Curcumin Complexes in Therapeutics: An Approach to Enhance Pharmacological Effects of Curcumin. Int. J. Mol. Sci. 2021, 22, 7094. [Google Scholar] [CrossRef] [PubMed]
- Shehzad, A.; Shahzad, R.; Lee, Y.S. Curcumin. In The Enzymes; Elsevier: Amsterdam, The Netherlands, 2014; Volume 36, pp. 149–174. ISBN 978-0-12-802215-3. [Google Scholar]
- Stanić, Z. Curcumin, a Compound from Natural Sources, a True Scientific Challenge–A Review. Plant Foods Hum. Nutr. 2017, 72, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rege, S.A.; Arya, M.; Momin, S.A. Structure Activity Relationship of Tautomers of Curcumin: A Review. Ukr. Food J. 2019, 8, 45–60. [Google Scholar] [CrossRef]
- Jakubczyk, K.; Drużga, A.; Katarzyna, J.; Skonieczna-Żydecka, K. Antioxidant Potential of Curcumin—A Meta-Analysis of Randomized Clinical Trials. Antioxidants 2020, 9, 1092. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Lu, J.; Holmgren, A. Thioredoxin Reductase Is Irreversibly Modified by Curcumin. J. Biol. Chem. 2005, 280, 25284–25290. [Google Scholar] [CrossRef]
- Gupta, A.P.; Khan, S.; Manzoor, M.M.; Yadav, A.K.; Sharma, G.; Anand, R.; Gupta, S. Anticancer Curcumin: Natural Analogues and Structure-Activity Relationship. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2017; Volume 54, pp. 355–401. ISBN 978-0-444-63929-5. [Google Scholar]
- Yang, Z.-J.; Huang, S.-Y.; Zhou, D.-D.; Xiong, R.-G.; Zhao, C.-N.; Fang, A.-P.; Zhang, Y.-J.; Li, H.-B.; Zhu, H.-L. Effects and Mechanisms of Curcumin for the Prevention and Management of Cancers: An Updated Review. Antioxidants 2022, 11, 1481. [Google Scholar] [CrossRef]
- Liu, X.; Qi, M.; Li, X.; Wang, J.; Wang, M. Curcumin: A Natural Organic Component That Plays a Multi-Faceted Role in Ovarian Cancer. J. Ovarian Res. 2023, 16, 47. [Google Scholar] [CrossRef]
- Yallapu, M.M.; Maher, D.M.; Sundram, V.; Bell, M.C.; Jaggi, M.; Chauhan, S.C. Curcumin Induces Chemo/Radio-Sensitization in Ovarian Cancer Cells and Curcumin Nanoparticles Inhibit Ovarian Cancer Cell Growth. J. Ovarian Res. 2010, 3, 11. [Google Scholar] [CrossRef]
- Selvendiran, K.; Ahmed, S.; Dayton, A.; Kuppusamy, M.L.; Rivera, B.K.; Kálai, T.; Hideg, K.; Kuppusamy, P. HO-3867, a Curcumin Analog, Sensitizes Cisplatin-Resistant Ovarian Carcinoma, Leading to Therapeutic Synergy through STAT3 Inhibition. Cancer Biol. Ther. 2011, 12, 837–845. [Google Scholar] [CrossRef]
- Kalinina, E.V.; Hasan, A.A.S.; Tatarskiy, V.V.; Volodina, Y.L.; Petrova, A.S.; Novichkova, M.D.; Zhdanov, D.D.; Nurmuradov, N.K.; Chernov, N.N.; Shtil, A.A. Suppression of PI3K/Akt/mTOR Signaling Pathway and Antioxidant System and Reversal of Cancer Cells Resistance to Cisplatin under the Effect of Curcumin. Bull. Exp. Biol. Med. 2022, 173, 371–375. [Google Scholar] [CrossRef]
- Hasan, A.A.; Kalinina, E.; Nuzhina, J.; Volodina, Y.; Shtil, A.; Tatarskiy, V. Potentiation of Cisplatin Cytotoxicity in Resistant Ovarian Cancer SKOV3/Cisplatin Cells by Quercetin Pre-Treatment. Int. J. Mol. Sci. 2023, 24, 10960. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Njiki, S.; Mbemi, A.; Yedjou, C.G.; Tchounwou, P.B. Pharmacological Effects of Cisplatin Combination with Natural Products in Cancer Chemotherapy. Int. J. Mol. Sci. 2022, 23, 1532. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shen, Z.; Zhu, T.; Lu, W.; Fu, Y. Curcumin Enhances the Anti-Cancer Efficacy of Paclitaxel in Ovarian Cancer by Regulating the miR-9-5p/BRCA1 Axis. Front. Pharmacol. 2023, 13, 1014933. [Google Scholar] [CrossRef]
- Jakobušić Brala, C.; Karković Marković, A.; Kugić, A.; Torić, J.; Barbarić, M. Combination Chemotherapy with Selected Polyphenols in Preclinical and Clinical Studies—An Update Overview. Molecules 2023, 28, 3746. [Google Scholar] [CrossRef]
- Weir, N.M.; Selvendiran, K.; Kutala, V.K.; Tong, L.; Vishwanath, S.; Rajaram, M.; Tridandapani, S.; Anant, S.; Kuppusamy, P. Curcumin Induces G2/M Arrest and Apoptosis in Cisplatin-Resistant Human Ovarian Cancer Cells by Modulating Akt and P38 mAPK. Cancer Biol. Ther. 2007, 6, 178–184. [Google Scholar] [CrossRef]
- Xiang, J.; Li, Y.; Zhang, Y.; Wang, G.; Xu, H.; Zhou, Z.; Tang, J.; Shen, Y. Polyphenol-Cisplatin Complexation Forming Core-Shell Nanoparticles with Improved Tumor Accumulation and Dual-Responsive Drug Release for Enhanced Cancer Chemotherapy. J. Control. Release 2021, 330, 992–1003. [Google Scholar] [CrossRef]
- Guerreiro, Í.; Ferreira-Pêgo, C.; Carregosa, D.; Santos, C.N.; Menezes, R.; Fernandes, A.S.; Costa, J.G. Polyphenols and Their Metabolites in Renal Diseases: An Overview. Foods 2022, 11, 1060. [Google Scholar] [CrossRef]
- Wróblewska-Łuczka, P.; Grabarska, A.; Florek-Łuszczki, M.; Plewa, Z.; Łuszczki, J.J. Synergy, Additivity, and Antagonism between Cisplatin and Selected Coumarins in Human Melanoma Cells. IJMS 2021, 22, 537. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Montanari, E.; Marzioni, D.; Goteri, G. Role of NRF2 in Ovarian Cancer. Antioxidants 2022, 11, 663. [Google Scholar] [CrossRef]
- Villa-Morales, M.; Pérez-Gómez, L.; Pérez-Gómez, E.; López-Nieva, P.; Fernández-Navarro, P.; Santos, J. Identification of NRF2 Activation as a Prognostic Biomarker in T-Cell Acute Lymphoblastic Leukaemia. Int. J. Mol. Sci. 2023, 24, 10350. [Google Scholar] [CrossRef]
- Jastrząb, A.; Skrzydlewska, E. Thioredoxin-Dependent System. Application of Inhibitors. J. Enzym. Inhib. Med. Chem. 2021, 36, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Sznarkowska, A.; Kostecka, A.; Meller, K.; Bielawski, K.P. Inhibition of Cancer Antioxidant Defense by Natural Compounds. Oncotarget 2017, 8, 15996–16016. [Google Scholar] [CrossRef] [PubMed]
- Nicolussi, A.; D’Inzeo, S.; Capalbo, C.; Giannini, G.; Coppa, A. The Role of Peroxiredoxins in Cancer. Mol. Clin. Oncol. 2017, 6, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.M.; Yoo, Y.D.; Park, J.K.; Kim, Y.T.; Kim, H.J. Increased Expression of Peroxiredoxin II Confers Resistance to Cisplatin. Anticancer. Res. 2001, 21, 1129–1133. [Google Scholar]
- Xu, J.; Su, Q.; Gao, M.; Liang, Q.; Li, J.; Chen, X. Differential Expression And Effects Of Peroxiredoxin-6 On Drug Resistance And Cancer Stem Cell-Like Properties In Non-Small Cell Lung Cancer. OTT 2019, 12, 10477–10486. [Google Scholar] [CrossRef]
- Yu, Y.; Di Trapani, G.; Tonissen, K.F. Thioredoxin and Glutathione Systems: Cancer Cells’ Defensive Weapons Against Oxidative Stress. In Handbook of Oxidative Stress in Cancer: Mechanistic Aspects; Chakraborti, S., Ray, B.K., Roychoudhury, S., Eds.; Springer Nature: Singapore, 2022; pp. 2407–2420. ISBN 978-981-15-9410-6. [Google Scholar]
- Chun, K.-S.; Jang, J.-H.; Kim, D.-H. Perspectives Regarding the Intersections between STAT3 and Oxidative Metabolism in Cancer. Cells 2020, 9, 2202. [Google Scholar] [CrossRef]
- Busker, S.; Qian, W.; Haraldsson, M.; Espinosa, B.; Johansson, L.; Attarha, S.; Kolosenko, I.; Liu, J.; Dagnell, M.; Grandér, D.; et al. Irreversible TrxR1 Inhibitors Block STAT3 Activity and Induce Cancer Cell Death. Sci. Adv. 2020, 6, eaax7945. [Google Scholar] [CrossRef]
- Dodd, K.M.; Yang, J.; Shen, M.H.; Sampson, J.R.; Tee, A.R. mTORC1 Drives HIF-1α and VEGF-A Signalling via Multiple Mechanisms Involving 4E-BP1, S6K1 and STAT3. Oncogene 2015, 34, 2239–2250. [Google Scholar] [CrossRef]
- Picco, G.; Chen, E.D.; Alonso, L.G.; Behan, F.M.; Gonçalves, E.; Bignell, G.; Matchan, A.; Fu, B.; Banerjee, R.; Anderson, E.; et al. Functional Linkage of Gene Fusions to Cancer Cell Fitness Assessed by Pharmacological and CRISPR-Cas9 Screening. Nat. Commun. 2019, 10, 2198. [Google Scholar] [CrossRef]
- DepMap, B. DepMap 24Q2 Public 2024, 25031191226 Bytes. Available online: https://plus.figshare.com/articles/dataset/DepMap_24Q2_Public/25880521/1 (accessed on 27 November 2024).
- Vasina, D.A.; Zhdanov, D.D.; Orlova, E.V.; Orlova, V.S.; Pokrovskaya, M.V.; Aleksandrova, S.S.; Sokolov, N.N. Apoptotic Endonuclease EndoG Inhibits Telomerase Activity and Induces Malignant Transformation of Human CD4+ T Cells. Biochem. Mosc. 2017, 82, 24–37. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense 5′–3′ | Antisense 5′–3′ |
|---|---|---|
| TRX1 | TGGTGAAGCAGATCGAGAGCAAGA | ACCACGTGGCTGAGAAGTCAACTA |
| TRX2 | TGGTGGCCTGACTGTAACAC | TGTTGACCACTCGGTCTTGA |
| TXNRD1 | GTGTTGTGGGCTTTCACGTA | TGGTCAGTCCACATTTGAGC |
| TXNRD2 | GCCATAGCACCTTGCATCTC | ATCCTCGATGAGGACACCTG |
| PRDX1 | ACAGCCGTTGTCAATGGAGAG | ACGTCGTGAAATTCGTTAGCTT |
| PRDX2 | CTGGCGAAGGACACCCTTGCCATC | GGCCACAGCGGTGGTTGATGGCG |
| PRDX3 | CTTGGTGTATTTATCCAGGCAAGATGGC | GGCCTGCTGCATGTGGAAGAACGA |
| PRDX6 | CAACTTTGAGGCCAATACCA | CAACTTAACATTCCTCTTGG |
| GAPDH | GAAGGTGAAGGTCGGAGTC | GAAGATGGTGATGGGATTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, A.A.; Kalinina, E.; Zhdanov, D.; Volodina, Y.; Tatarskiy, V. Re-Sensitization of Resistant Ovarian Cancer SKOV3/CDDP Cells to Cisplatin by Curcumin Pre-Treatment. Int. J. Mol. Sci. 2025, 26, 799. https://doi.org/10.3390/ijms26020799
Hasan AA, Kalinina E, Zhdanov D, Volodina Y, Tatarskiy V. Re-Sensitization of Resistant Ovarian Cancer SKOV3/CDDP Cells to Cisplatin by Curcumin Pre-Treatment. International Journal of Molecular Sciences. 2025; 26(2):799. https://doi.org/10.3390/ijms26020799
Chicago/Turabian StyleHasan, Aseel Ali, Elena Kalinina, Dmitry Zhdanov, Yulia Volodina, and Victor Tatarskiy. 2025. "Re-Sensitization of Resistant Ovarian Cancer SKOV3/CDDP Cells to Cisplatin by Curcumin Pre-Treatment" International Journal of Molecular Sciences 26, no. 2: 799. https://doi.org/10.3390/ijms26020799
APA StyleHasan, A. A., Kalinina, E., Zhdanov, D., Volodina, Y., & Tatarskiy, V. (2025). Re-Sensitization of Resistant Ovarian Cancer SKOV3/CDDP Cells to Cisplatin by Curcumin Pre-Treatment. International Journal of Molecular Sciences, 26(2), 799. https://doi.org/10.3390/ijms26020799