
Functional Divergence of NOTCH1 and NOTCH2 in Human Cerebral Organoids Reveals Receptor-Specific Roles in Early Corticogenesis

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Depletion of NOTCH1, but Not NOTCH2, Alters External Morphology of COs
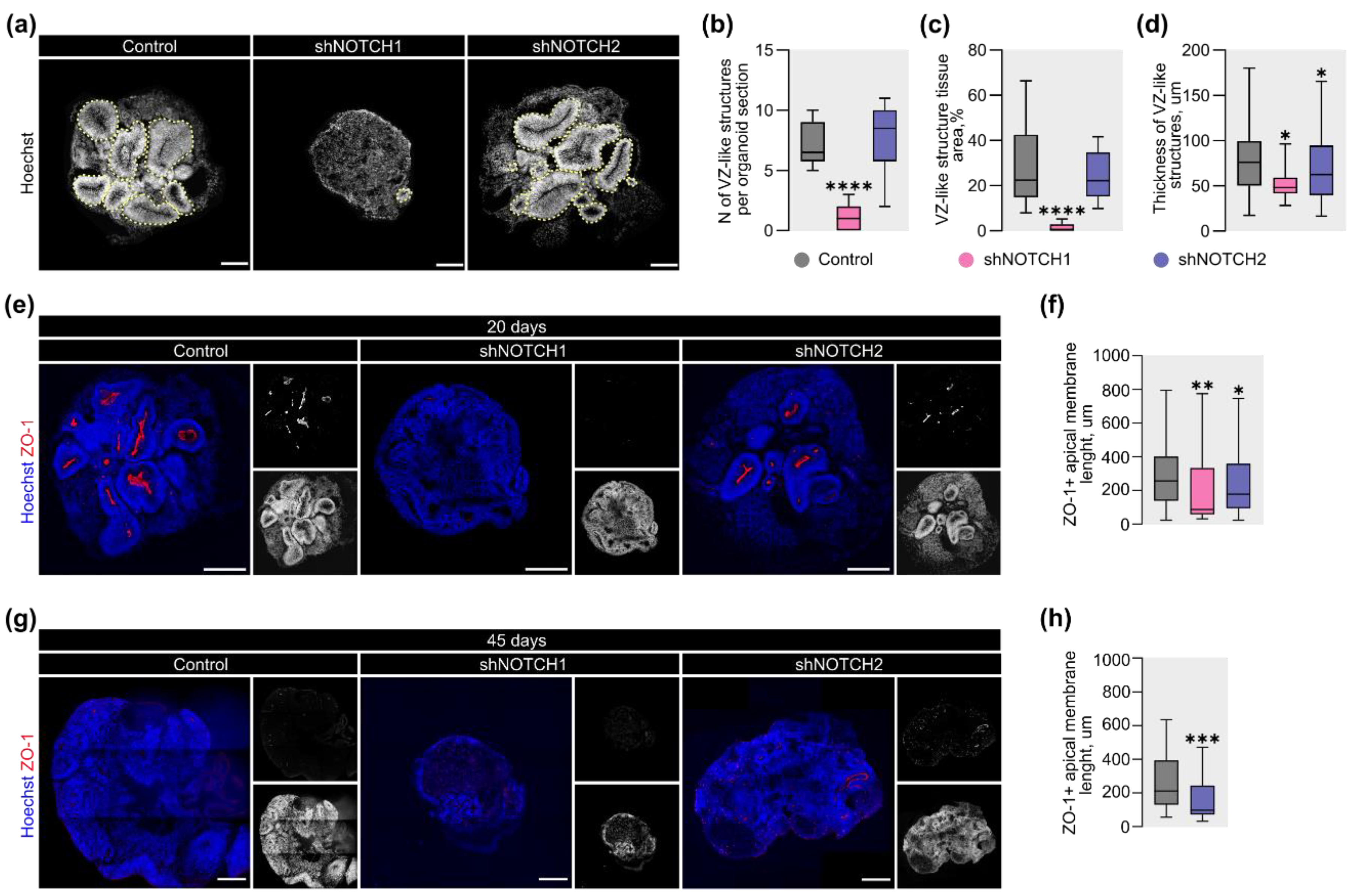
2.2. Differential Effects of NOTCH1 and NOTCH2 on Apical Polarity and Ventricle-Like Structures
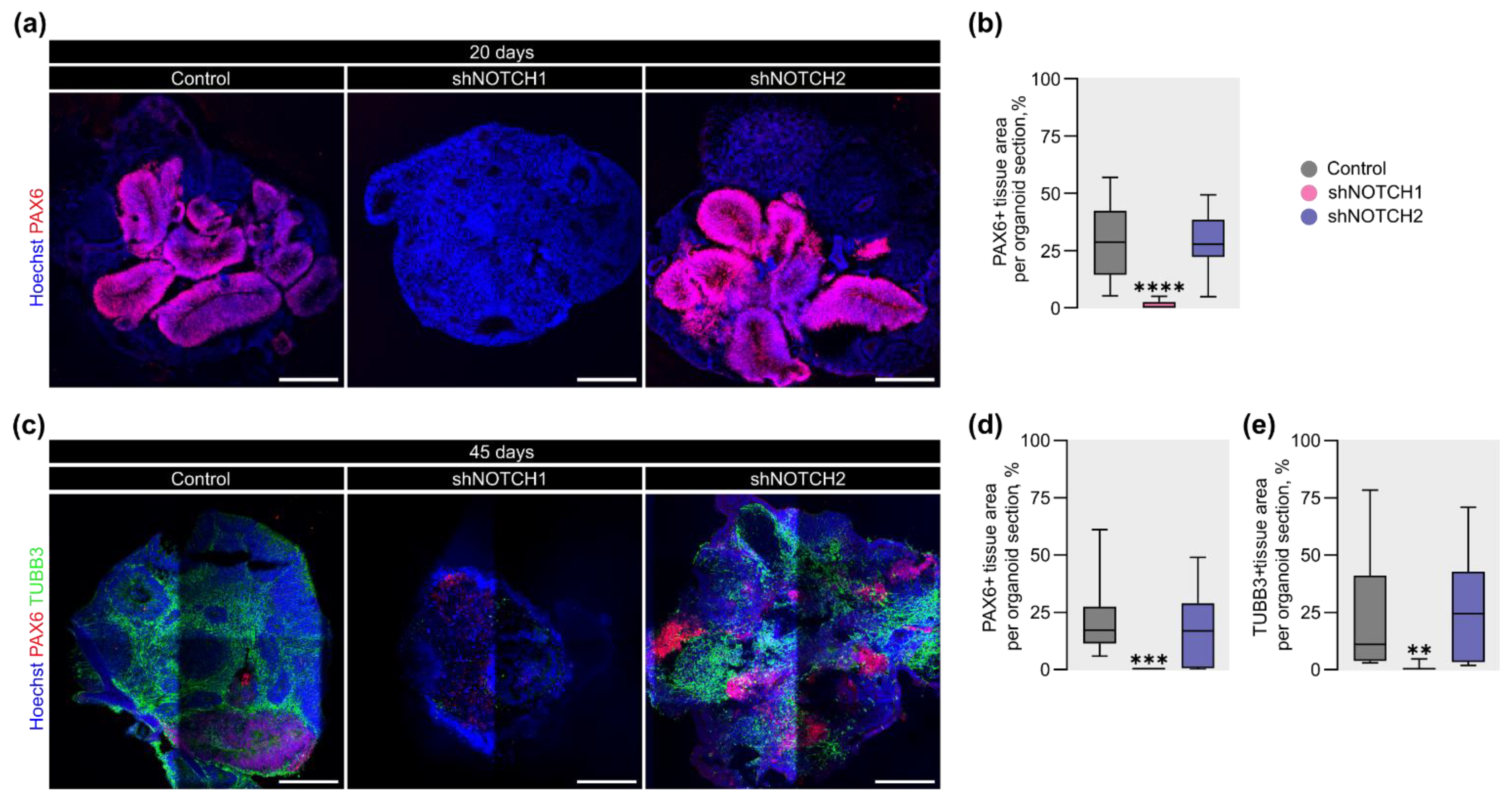
2.3. NOTCH1 Is Required for RGC Fate Specification and Maintenance of Cortical Progenitor Identity
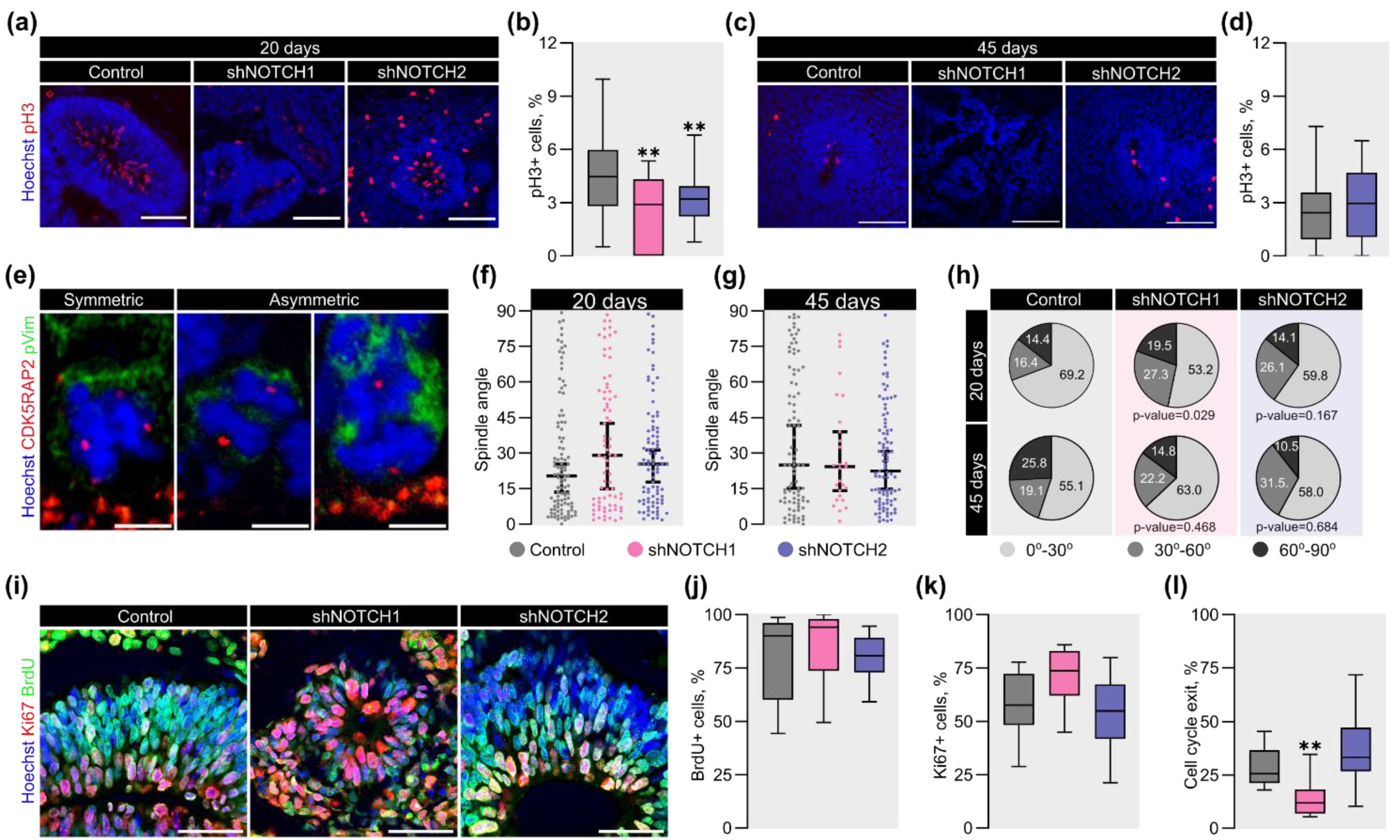
2.4. NOTCH1 Regulates the Proliferative Capacity and Division Mode of RGC in COs
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Production of Second-Generation Lentiviruses and Transduction
4.3. Generation of COs
4.4. Fixation and Cryosectioning
4.5. Immunohistochemistry and Imaging
4.6. Image Processing and Analysis
4.7. Western Blotting
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| bFGF | basic Fibroblast Growth Factor |
| BrdU | Directory of open access journals |
| CO | Cerebral organoid |
| EB | Embryoid body |
| hiPSC | Human-induced pluripotent stem cell |
| shRNA | Short-hairpin RNA |
| VLS | Ventricle-like structure |
References
- Louvi, A.; Artavanis-Tsakonas, S. Notch Signalling in Vertebrate Neural Development. Nat. Rev. Neurosci. 2006, 7, 93–102. [Google Scholar] [CrossRef]
- Pierfelice, T.; Alberi, L.; Gaiano, N. Notch in the Vertebrate Nervous System: An Old Dog with New Tricks. Neuron 2011, 69, 840–855. [Google Scholar] [CrossRef]
- Yoon, K.; Gaiano, N. Notch Signaling in the Mammalian Central Nervous System: Insights from Mouse Mutants. Nat. Neurosci. 2005, 8, 709–715. [Google Scholar] [CrossRef]
- Ishibashi, M.; Ang, S.L.; Shiota, K.; Nakanishi, S.; Kageyama, R.; Guillemot, F. Targeted Disruption of Mammalian Hairy and Enhancer of Split Homolog-1 (HES-1) Leads to up-Regulation of Neural Helix-Loop-Helix Factors, Premature Neurogenesis, and Severe Neural Tube Defects. Genes Dev. 1995, 9, 3136–3148. [Google Scholar] [CrossRef]
- Handler, M.; Yang, X.; Shent, J. Presenilin-1 Regulates Neuronal Differentiation during Neurogenesis. Development 2000, 127, 2593–2606. [Google Scholar] [CrossRef]
- Hitoshi, S.; Alexson, T.; Tropepe, V.; Donoviel, D.; Elia, A.J.; Nye, J.S.; Conlon, R.A.; Mak, T.W.; Bernstein, A.; van der Kooy, D. Notch Pathway Molecules Are Essential for the Maintenance, but Not the Generation, of Mammalian Neural Stem Cells. Genes Dev. 2002, 16, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Lütolf, S.; Radtke, F.; Aguet, M.; Suter, U.; Taylor, V. Notch1 Is Required for Neuronal and Glial Differentiation in the Cerebellum. Development 2002, 129, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Grandbarbe, L.; Bouissac, J.; Rand, M.; Hrabé de Angelis, M.; Artavanis-Tsakonas, S.; Mohier, E. Delta-Notch Signaling Controls the Generation of Neurons/Glia from Neural Stem Cells in a Stepwise Process. Development 2003, 130, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.-Y.; Hu, Q.-D.; Tekaya, M.; Shimoda, Y.; Ang, B.-T.; Nie, D.-Y.; Sun, L.; Hu, W.-P.; Karsak, M.; Duka, T.; et al. NB-3/Notch1 Pathway via Deltex1 Promotes Neural Progenitor Cell Differentiation into Oligodendrocytes. J. Biol. Chem. 2004, 279, 25858–25865. [Google Scholar] [CrossRef]
- Hatakeyama, J.; Bessho, Y.; Katoh, K.; Ookawara, S.; Fujioka, M.; Guillemot, F.; Kageyama, R. Hes Genes Regulate Size, Shape and Histogenesis of the Nervous System by Control of the Timing of Neural Stem Cell Differentiation. Development 2004, 131, 5539–5550. [Google Scholar] [CrossRef]
- Koo, B.-K.; Lim, H.-S.; Song, R.; Yoon, M.-J.; Yoon, K.-J.; Moon, J.-S.; Kim, Y.-W.; Kwon, M.; Yoo, K.-W.; Kong, M.-P.; et al. Mind Bomb 1 Is Essential for Generating Functional Notch Ligands to Activate Notch. Development 2005, 132, 3459–3470. [Google Scholar] [CrossRef]
- Hamada, Y.; Kadokawa, Y.; Okabe, M.; Ikawa, M.; Coleman, J.R.; Tsujimoto, Y. Mutation in Ankyrin Repeats of the Mouse Notch2 Gene Induces Early Embryonic Lethality. Development 1999, 126, 3415–3424. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Klein, R.; Tian, X.; Cheng, H.-T.; Kopan, R.; Shen, J. Notch Activation Induces Apoptosis in Neural Progenitor Cells through a P53-Dependent Pathway. Dev. Biol. 2004, 269, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Mason, H.A.; Rakowiecki, S.M.; Gridley, T.; Fishell, G. Loss of Notch Activity in the Developing Central Nervous System Leads to Increased Cell Death. Dev. Neurosci. 2006, 28, 49–57. [Google Scholar] [CrossRef]
- Berezovska, O.; McLean, P.; Knowles, R.; Frosh, M.; Lu, F.M.; Lux, S.E.; Hyman, B.T. Notch1 Inhibits Neurite Outgrowth in Postmitotic Primary Neurons. Neuroscience 1999, 93, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.L.; Berechid, B.E.; Cutting, F.B.; Presente, A.; Chambers, C.B.; Foltz, D.R.; Ferreira, A.; Nye, J.S. Autonomous and Non-Autonomous Regulation of Mammalian Neurite Development by Notch1 and Delta1. Curr. Biol. 1999, 9, 1448–1457. [Google Scholar] [CrossRef]
- Sestan, N.; Artavanis-Tsakonas, S.; Rakic, P. Contact-Dependent Inhibition of Cortical Neurite Growth Mediated by Notch Signaling. Science 1999, 286, 741–746. [Google Scholar] [CrossRef]
- Huang, E.J.; Li, H.; Tang, A.A.; Wiggins, A.K.; Neve, R.L.; Zhong, W.; Jan, L.Y.; Jan, Y.N. Targeted Deletion of Numb and Numblike in Sensory Neurons Reveals Their Essential Functions in Axon Arborization. Genes Dev. 2005, 19, 138–151. [Google Scholar] [CrossRef]
- Redmond, L.; Oh, S.-R.; Hicks, C.; Weinmaster, G.; Ghosh, A. Nuclear Notch1 Signaling and the Regulation of Dendritic Development. Nat. Neurosci. 2000, 3, 30–40. [Google Scholar] [CrossRef]
- Cotter, D.; Honavar, M.; Lovestone, S.; Raymond, L.; Kerwin, R.; Anderton, B.; Everall, I. Disturbance of Notch-1 and Wnt Signalling Proteins in Neuroglial Balloon Cells and Abnormal Large Neurons in Focal Cortical Dysplasia in Human Cortex. Acta Neuropathol. 1999, 98, 465–472. [Google Scholar] [CrossRef]
- Fiddes, I.T.; Lodewijk, G.A.; Mooring, M.; Bosworth, C.M.; Ewing, A.D.; Mantalas, G.L.; Novak, A.M.; van den Bout, A.; Bishara, A.; Rosenkrantz, J.L.; et al. Human-Specific NOTCH2NL Genes Affect Notch Signaling and Cortical Neurogenesis. Cell 2018, 173, 1356–1369.e22. [Google Scholar] [CrossRef]
- Fischer-Zirnsak, B.; Segebrecht, L.; Schubach, M.; Charles, P.; Alderman, E.; Brown, K.; Cadieux-Dion, M.; Cartwright, T.; Chen, Y.; Costin, C.; et al. Haploinsufficiency of the Notch Ligand DLL1 Causes Variable Neurodevelopmental Disorders. Am. J. Hum. Genet. 2019, 105, 631–639. [Google Scholar] [CrossRef]
- Zhang, Y.-H.; Wang, T.; Li, Y.-F.; Deng, Y.-N.; Shen, F.-G. Roles of the Notch Signaling Pathway and Microglia in Autism. Behav. Brain Res. 2023, 437, 114131. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, T.C.; de Oliveira, J.R.M. Reconsidering the Association between the Major Histocompatibility Complex and Bipolar Disorder. J. Mol. Neurosci. 2012, 47, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wei, J.; Zhang, X.; Guo, Y.; Xu, Q.; Liu, S.; Shi, J.; Yu, Y.; Ju, G.; Li, Y.; et al. A Review and Re-Evaluation of an Association between the NOTCH4 Locus and Schizophrenia. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2006, 141B, 902–906. [Google Scholar] [CrossRef] [PubMed]
- Hoseth, E.Z.; Krull, F.; Dieset, I.; Mørch, R.H.; Hope, S.; Gardsjord, E.S.; Steen, N.E.; Melle, I.; Brattbakk, H.-R.; Steen, V.M.; et al. Attenuated Notch Signaling in Schizophrenia and Bipolar Disorder. Sci. Rep. 2018, 8, 5349. [Google Scholar] [CrossRef]
- Gazave, E.; Lapébie, P.; Richards, G.S.; Brunet, F.; Ereskovsky, A.V.; Degnan, B.M.; Borchiellini, C.; Vervoort, M.; Renard, E. Origin and Evolution of the Notch Signalling Pathway: An Overview from Eukaryotic Genomes. BMC Evol. Biol. 2009, 9, 249. [Google Scholar] [CrossRef]
- Swiatek, P.J.; Lindsell, C.E.; del Amo, F.F.; Weinmaster, G.; Gridley, T. Notch1 Is Essential for Postimplantation Development in Mice. Genes Dev. 1994, 8, 707–719. [Google Scholar] [CrossRef]
- Higuchi, M.; Kiyama, H.; Hayakawa, T.; Hamada, Y.; Tsujimoto, Y. Differential Expression of Notch1 and Notch2 in Developing and Adult Mouse Brain. Brain Res. Mol. Brain Res. 1995, 29, 263–272. [Google Scholar] [CrossRef]
- Lindsell, C.E.; Boulter, J.; diSibio, G.; Gossler, A.; Weinmaster, G. Expression Patterns of Jagged, Delta1, Notch1, Notch2, and Notch3 Genes Identify Ligand–Receptor Pairs That May Function in Neural Development. Mol. Cell. Neurosci. 1996, 8, 14–27. [Google Scholar] [CrossRef]
- Irvin, D.K.; Zurcher, S.D.; Nguyen, T.; Weinmaster, G.; Kornblum, H.I. Expression Patterns of Notch1, Notch2, and Notch3 Suggest Multiple Functional Roles for the Notch-DSL Signaling System during Brain Development. J. Comp. Neurol. 2001, 2, 167–181. [Google Scholar] [CrossRef]
- Conlon, R.A.; Reaume, A.G.; Rossant, J. Notch1 Is Required for the Coordinate Segmentation of Somites. Development 1995, 121, 1533–1545. [Google Scholar] [CrossRef]
- de la Pompa, J.L.; Wakeham, A.; Correia, K.M.; Samper, E.; Brown, S.; Aguilera, R.J.; Nakano, T.; Honjo, T.; Mak, T.W.; Rossant, J.; et al. Conservation of the Notch Signalling Pathway in Mammalian Neurogenesis. Development 1997, 124, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Gaiano, N.; Nye, J.S.; Fishell, G. Radial Glial Identity Is Promoted by Notch1 Signaling in the Murine Forebrain. Neuron 2000, 26, 395–404. [Google Scholar] [CrossRef] [PubMed]
- McCright, B.; Gao, X.; Shen, L.; Lozier, J.; Lan, Y.; Maguire, M.; Herzlinger, D.; Weinmaster, G.; Jiang, R.; Gridley, T. Defects in Development of the Kidney, Heart and Eye Vasculature in Mice Homozygous for a Hypomorphic Notch2 Mutation. Development 2001, 128, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Krebs, L.T.; Xue, Y.; Norton, C.R.; Shutter, J.R.; Maguire, M.; Sundberg, J.P.; Gallahan, D.; Closson, V.; Kitajewski, J.; Callahan, R.; et al. Notch Signaling Is Essential for Vascular Morphogenesis in Mice. Genes Dev. 2000, 14, 1343–1352. [Google Scholar] [CrossRef]
- Chambers, C.B.; Peng, Y.; Nguyen, H.; Gaiano, N.; Fishell, G.; Nye, J.S. Spatiotemporal Selectivity of Response to Notch1 Signals in Mammalian Forebrain Precursors. Development 2001, 128, 689–702. [Google Scholar] [CrossRef]
- Mizutani, K.; Yoon, K.; Dang, L.; Tokunaga, A.; Gaiano, N. Differential Notch Signalling Distinguishes Neural Stem Cells from Intermediate Progenitors. Nature 2007, 449, 351–355. [Google Scholar] [CrossRef]
- Nandagopal, N.; Santat, L.A.; LeBon, L.; Sprinzak, D.; Bronner, M.E.; Elowitz, M.B. Dynamic Ligand Discrimination in the Notch Signaling Pathway. Cell 2018, 172, 869–880.e19. [Google Scholar] [CrossRef]
- Kakuda, S.; LoPilato, R.K.; Ito, A.; Haltiwanger, R.S. Canonical Notch Ligands and Fringes Have Distinct Effects on NOTCH1 and NOTCH2. J. Biol. Chem. 2020, 295, 14710–14722. [Google Scholar] [CrossRef]
- Kuintzle, R.; Santat, L.A.; Elowitz, M.B. Diversity in Notch Ligand-Receptor Signaling Interactions. eLife 2025, 12, RP91422. [Google Scholar] [CrossRef]
- Shimizu, K.; Chiba, S.; Saito, T.; Kumano, K.; Hamada, Y.; Hirai, H. Functional Diversity among Notch1, Notch2, and Notch3 Receptors. Biochem. Biophys. Res. Commun. 2002, 291, 775–779. [Google Scholar] [CrossRef]
- Patten, B.A.; Peyrin, J.M.; Weinmaster, G.; Corfas, G. Sequential Signaling through Notch1 and erbB Receptors Mediates Radial Glia Differentiation. J. Neurosci. 2003, 23, 6132–6140. [Google Scholar] [CrossRef] [PubMed]
- Tchorz, J.S.; Tome, M.; Cloëtta, D.; Sivasankaran, B.; Grzmil, M.; Huber, R.M.; Rutz-Schatzmann, F.; Kirchhoff, F.; Schaeren-Wiemers, N.; Gassmann, M.; et al. Constitutive Notch2 Signaling in Neural Stem Cells Promotes Tumorigenic Features and Astroglial Lineage Entry. Cell Death Dis. 2012, 3, e325. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.-J.; Koo, B.-K.; Im, S.-K.; Jeong, H.-W.; Ghim, J.; Kwon, M.; Moon, J.-S.; Miyata, T.; Kong, Y.-Y. Mind Bomb 1-Expressing Intermediate Progenitors Generate Notch Signaling to Maintain Radial Glial Cells. Neuron 2008, 58, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Mase, S.; Shitamukai, A.; Wu, Q.; Morimoto, M.; Gridley, T.; Matsuzaki, F. Notch1 and Notch2 Collaboratively Maintain Radial Glial Cells in Mouse Neurogenesis. Neurosci. Res. 2021, 170, 122–132. [Google Scholar] [CrossRef]
- Eşiyok, N.; Liutikaite, N.; Haffner, C.; Peters, J.; Heide, S.; Oegema, C.E.; Huttner, W.B.; Heide, M. A Dyad of Human-Specific NBPF14 and NOTCH2NLB Orchestrates Cortical Progenitor Abundance Crucial for Human Neocortex Expansion. Sci. Adv. 2025, 11, eads7543. [Google Scholar] [CrossRef]
- Lodewijk, G.A.; Fernandes, D.P.; Vretzakis, I.; Savage, J.E.; Jacobs, F.M.J. Evolution of Human Brain Size-Associated NOTCH2NL Genes Proceeds toward Reduced Protein Levels. Mol. Biol. Evol. 2020, 37, 2531–2548. [Google Scholar] [CrossRef]
- Suzuki, I.K.; Gacquer, D.; Van Heurck, R.; Kumar, D.; Wojno, M.; Bilheu, A.; Herpoel, A.; Lambert, N.; Cheron, J.; Polleux, F.; et al. Human-Specific NOTCH2NL Genes Expand Cortical Neurogenesis through Delta/Notch Regulation. Cell 2018, 173, 1370–1384.e16. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Chiaradia, I.; Imaz-Rosshandler, I.; Nilges, B.S.; Boulanger, J.; Pellegrini, L.; Das, R.; Kashikar, N.D.; Lancaster, M.A. Tissue Morphology Influences the Temporal Program of Human Brain Organoid Development. Cell Stem Cell 2023, 30, 1351–1367.e10. [Google Scholar] [CrossRef]
- Siebel, C.; Lendahl, U. Notch Signaling in Development, Tissue Homeostasis, and Disease. Physiol. Rev. 2017, 97, 1235–1294. [Google Scholar] [CrossRef] [PubMed]
- Main, H.; Radenkovic, J.; Jin, S.; Lendahl, U.; Andersson, E.R. Notch Signaling Maintains Neural Rosette Polarity. PLoS ONE 2013, 8, e62959. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.J.; Heisenberg, C.-P.; Hiiragi, T. Coordination of Morphogenesis and Cell-Fate Specification in Development. Curr. Biol. 2017, 27, R1024–R1035. [Google Scholar] [CrossRef] [PubMed]
- Han, J.S.; Fishman-Williams, E.; Decker, S.C.; Hino, K.; Reyes, R.V.; Brown, N.L.; Simó, S.; Torre, A.L. Notch Directs Telencephalic Development and Controls Neocortical Neuron Fate Determination by Regulating microRNA Levels. Development 2023, 150, dev201408. [Google Scholar] [CrossRef]
- Solecki, D.J.; Liu, X.; Tomoda, T.; Fang, Y.; Hatten, M.E. Activated Notch2 Signaling Inhibits Differentiation of Cerebellar Granule Neuron Precursors by Maintaining Proliferation. Neuron 2001, 31, 557–568. [Google Scholar] [CrossRef]
- Yoon, K.; Nery, S.; Rutlin, M.L.; Radtke, F.; Fishell, G.; Gaiano, N. Fibroblast Growth Factor Receptor Signaling Promotes Radial Glial Identity and Interacts with Notch1 Signaling in Telencephalic Progenitors. J. Neurosci. 2004, 24, 9497–9506. [Google Scholar] [CrossRef]
- Wang, X.; Mao, X.; Xie, L.; Greenberg, D.A.; Jin, K. Involvement of Notch1 Signaling in Neurogenesis in the Subventricular Zone of Normal and Ischemic Rat Brain in Vivo. J. Cereb. Blood Flow. Metab. 2009, 29, 1644–1654. [Google Scholar] [CrossRef]
- Kawai, H.; Kawaguchi, D.; Kuebrich, B.D.; Kitamoto, T.; Yamaguchi, M.; Gotoh, Y.; Furutachi, S. Area-Specific Regulation of Quiescent Neural Stem Cells by Notch3 in the Adult Mouse Subependymal Zone. J. Neurosci. 2017, 37, 11867–11880. [Google Scholar] [CrossRef]
- Bultje, R.S.; Castaneda-Castellanos, D.R.; Jan, L.Y.; Jan, Y.-N.; Kriegstein, A.R.; Shi, S.-H. Mammalian Par3 Regulates Progenitor Cell Asymmetric Division via Notch Signaling in the Developing Neocortex. Neuron 2009, 63, 189–202. [Google Scholar] [CrossRef]
- Bohl, B.; Jabali, A.; Ladewig, J.; Koch, P. Asymmetric Notch Activity by Differential Inheritance of Lysosomes in Human Neural Stem Cells. Sci. Adv. 2022, 8, eabl5792. [Google Scholar] [CrossRef]
- Perez-Mockus, G.; Schweisguth, F. Cell Polarity and Notch Signaling: Linked by the E3 Ubiquitin Ligase Neuralized? Bioessays 2017, 39. [Google Scholar] [CrossRef]
- White, M.J.; Jacobs, K.A.; Singh, T.; Mayo, L.N.; Lin, A.; Chen, C.S.; Jun, Y.; Kutys, M.L. Notch1 Cortical Signaling Regulates Epithelial Architecture and Cell–Cell Adhesion. J. Cell Biol. 2023, 222, e202303013. [Google Scholar] [CrossRef]
- Kim, S.; Lehtinen, M.K.; Sessa, A.; Zappaterra, M.; Cho, S.-H.; Gonzalez, D.; Boggan, B.; Austin, C.A.; Wijnholds, J.; Gambello, M.J.; et al. The Apical Complex Couples Cell Fate and Cell Survival to Cerebral Cortical Development. Neuron 2010, 66, 69. [Google Scholar] [CrossRef]
- Knoblich, J.A. Mechanisms of Asymmetric Stem Cell Division. Cell 2008, 132, 583–597. [Google Scholar] [CrossRef]
- Siller, K.H.; Doe, C.Q. Spindle Orientation during Asymmetric Cell Division. Nat. Cell Biol. 2009, 11, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, F.; Shitamukai, A. Cell Division Modes and Cleavage Planes of Neural Progenitors during Mammalian Cortical Development. Cold Spring Harb. Perspect. Biol. 2015, 7, a015719. [Google Scholar] [CrossRef] [PubMed]
- Chenn, A.; McConnell, S.K. Cleavage Orientation and the Asymmetric Inheritance of Notch1 Immunoreactivity in Mammalian Neurogenesis. Cell 1995, 82, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhong, W.; Jan, Y.N.; Temple, S. Asymmetric Numb Distribution Is Critical for Asymmetric Cell Division of Mouse Cerebral Cortical Stem Cells and Neuroblasts. Development 2002, 129, 4843–4853. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Yang, N.; Yeo, S.-Y.; Chitnis, A.; Guo, S. Intralineage Directional Notch Signaling Regulates Self-Renewal and Differentiation of Asymmetrically Dividing Radial Glia. Neuron 2012, 74, 65–78. [Google Scholar] [CrossRef]
- Kechad, A.; Jolicoeur, C.; Tufford, A.; Mattar, P.; Chow, R.W.Y.; Harris, W.A.; Cayouette, M. Numb Is Required for the Production of Terminal Asymmetric Cell Divisions in the Developing Mouse Retina. J. Neurosci. 2012, 32, 17197–17210. [Google Scholar] [CrossRef]
- Gridina, M.M.; Matveeva, N.M.; Fishman, V.S.; Menzorov, A.G.; Kizilova, H.A.; Beregovoy, N.A.; Kovrigin, I.I.; Pristyazhnyuk, I.E.; Oscorbin, I.P.; Filipenko, M.L.; et al. Allele-Specific Biased Expression of the CNTN6 Gene in iPS Cell-Derived Neurons from a Patient with Intellectual Disability and 3p26.3 Microduplication Involving the CNTN6 Gene. Mol. Neurobiol. 2018, 55, 6533–6546. [Google Scholar] [CrossRef]
- Perepletchikova, D.; Kuchur, P.; Basovich, L.; Khvorova, I.; Lobov, A.; Azarkina, K.; Aksenov, N.; Bozhkova, S.; Karelkin, V.; Malashicheva, A. Endothelial-Mesenchymal Crosstalk Drives Osteogenic Differentiation of Human Osteoblasts through Notch Signaling. Cell Commun. Signal. 2025, 23, 100. [Google Scholar] [CrossRef] [PubMed]
- Moffat, J.; Grueneberg, D.A.; Yang, X.; Kim, S.Y.; Kloepfer, A.M.; Hinkle, G.; Piqani, B.; Eisenhaure, T.M.; Luo, B.; Grenier, J.K.; et al. A Lentiviral RNAi Library for Human and Mouse Genes Applied to an Arrayed Viral High-Content Screen. Cell 2006, 124, 1283–1298. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Iefremova, V.; Manikakis, G.; Krefft, O.; Jabali, A.; Weynans, K.; Wilkens, R.; Marsoner, F.; Brändl, B.; Müller, F.-J.; Koch, P.; et al. An Organoid-Based Model of Cortical Development Identifies Non-Cell-Autonomous Defects in Wnt Signaling Contributing to Miller-Dieker Syndrome. Cell Rep. 2017, 19, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Yeung, Y.-G.; Stanley, E.R. A Solution for Stripping Antibodies from Polyvinylidene Fluoride Immunoblots for Multiple Reprobing. Anal. Biochem. 2009, 389, 89–91. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yakovleva, S.; Knyazeva, A.; Yunusova, A.; Allayarova, E.; Lanshakov, D.; Malashicheva, A.; Shnaider, T. Functional Divergence of NOTCH1 and NOTCH2 in Human Cerebral Organoids Reveals Receptor-Specific Roles in Early Corticogenesis. Int. J. Mol. Sci. 2025, 26, 7309. https://doi.org/10.3390/ijms26157309
Yakovleva S, Knyazeva A, Yunusova A, Allayarova E, Lanshakov D, Malashicheva A, Shnaider T. Functional Divergence of NOTCH1 and NOTCH2 in Human Cerebral Organoids Reveals Receptor-Specific Roles in Early Corticogenesis. International Journal of Molecular Sciences. 2025; 26(15):7309. https://doi.org/10.3390/ijms26157309
Chicago/Turabian StyleYakovleva, Sophia, Anastasia Knyazeva, Anastasia Yunusova, Elina Allayarova, Dmitriy Lanshakov, Anna Malashicheva, and Tatiana Shnaider. 2025. "Functional Divergence of NOTCH1 and NOTCH2 in Human Cerebral Organoids Reveals Receptor-Specific Roles in Early Corticogenesis" International Journal of Molecular Sciences 26, no. 15: 7309. https://doi.org/10.3390/ijms26157309
APA StyleYakovleva, S., Knyazeva, A., Yunusova, A., Allayarova, E., Lanshakov, D., Malashicheva, A., & Shnaider, T. (2025). Functional Divergence of NOTCH1 and NOTCH2 in Human Cerebral Organoids Reveals Receptor-Specific Roles in Early Corticogenesis. International Journal of Molecular Sciences, 26(15), 7309. https://doi.org/10.3390/ijms26157309