

Arabidopsis Antiporter Genes as Targets of NO Signalling: Phylogenetic, Structural, and Expression Analysis
 , , , and
, , , and
Abstract
1. Introduction
2. Results
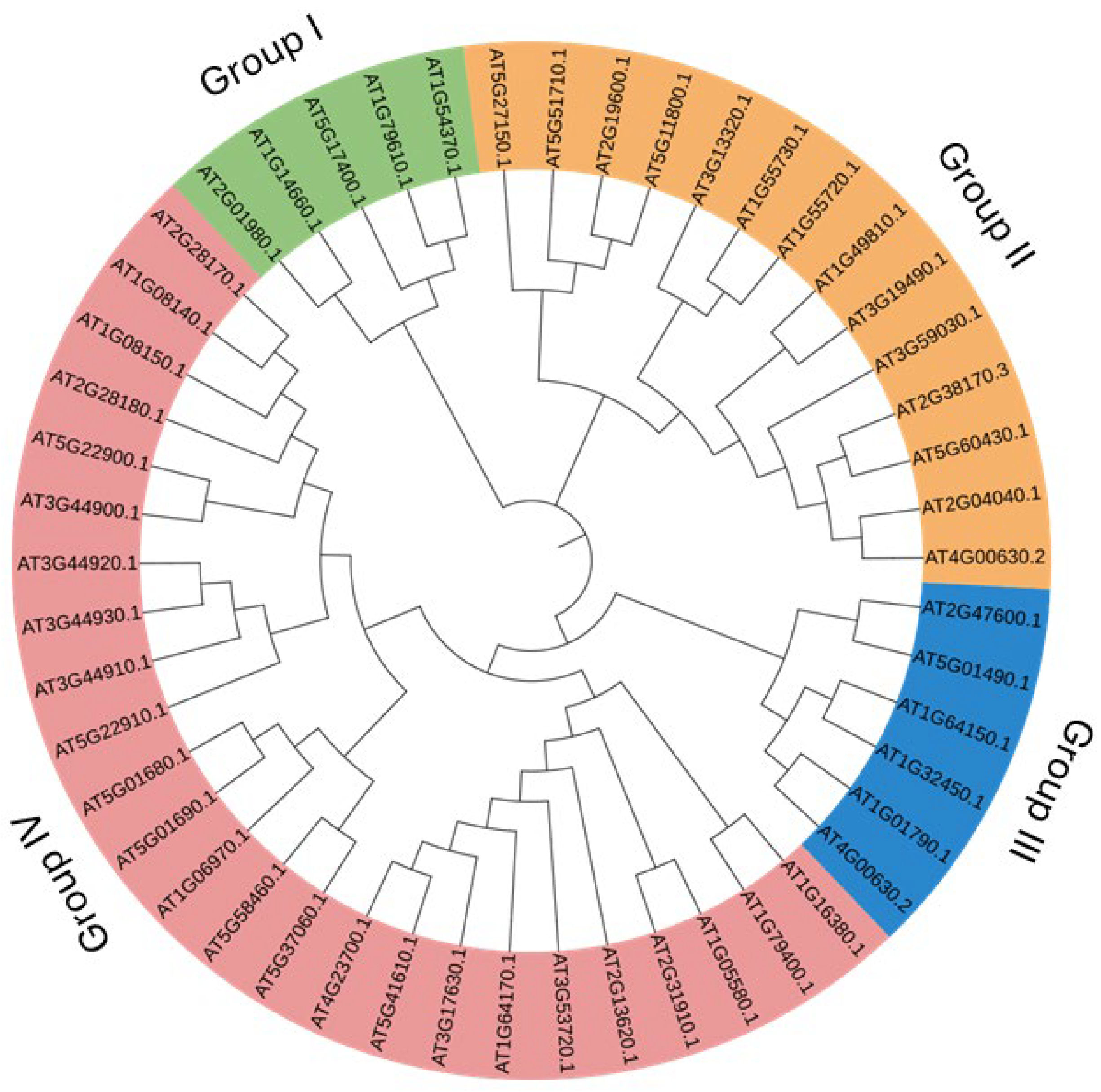
2.1. Identification of Genes and Phylogenetic Analysis
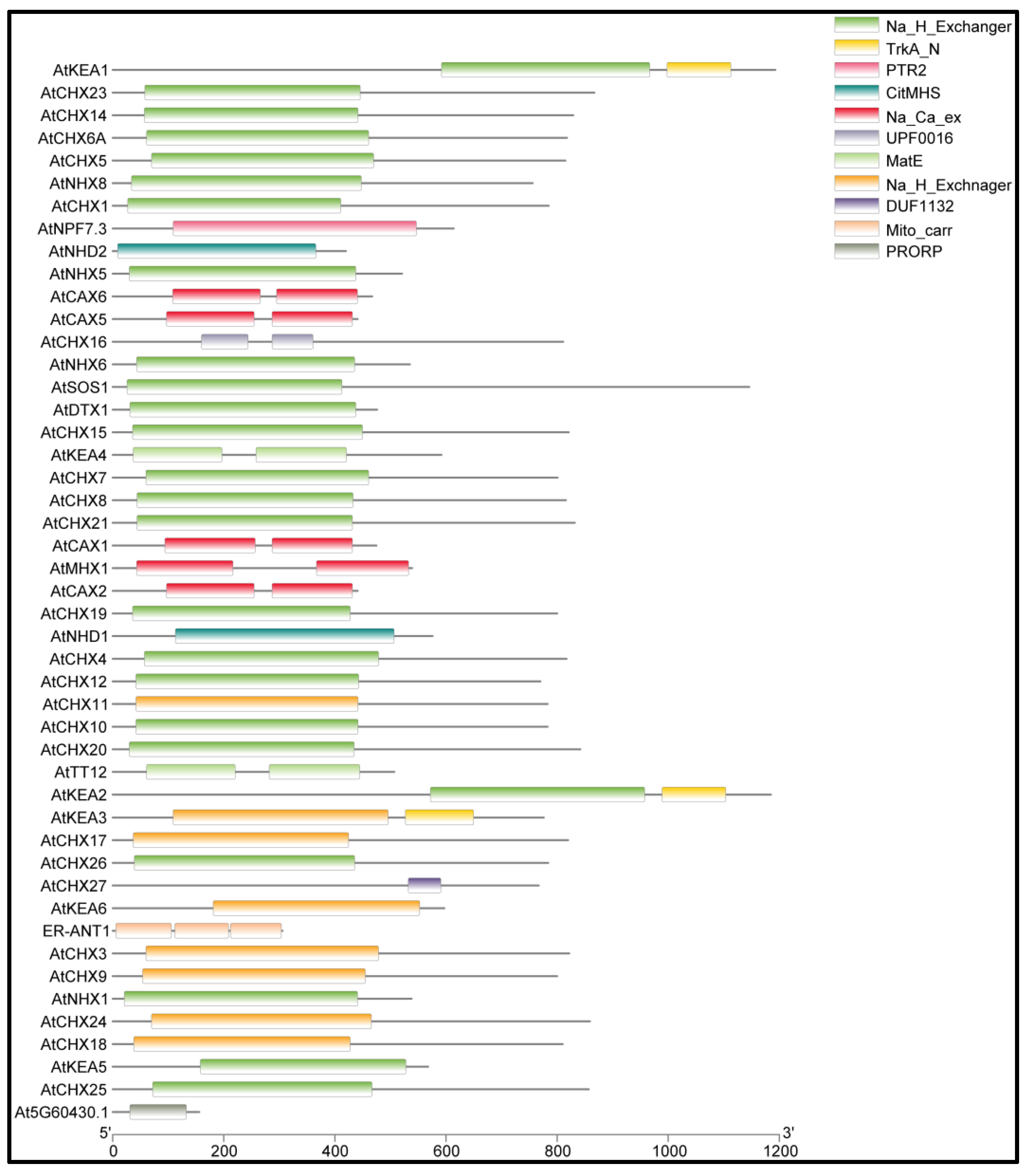
2.2. Domain Analysis
2.3. Chromosomal Distribution of Genes
2.4. Physiochemical Attributes of Antiporter Genes
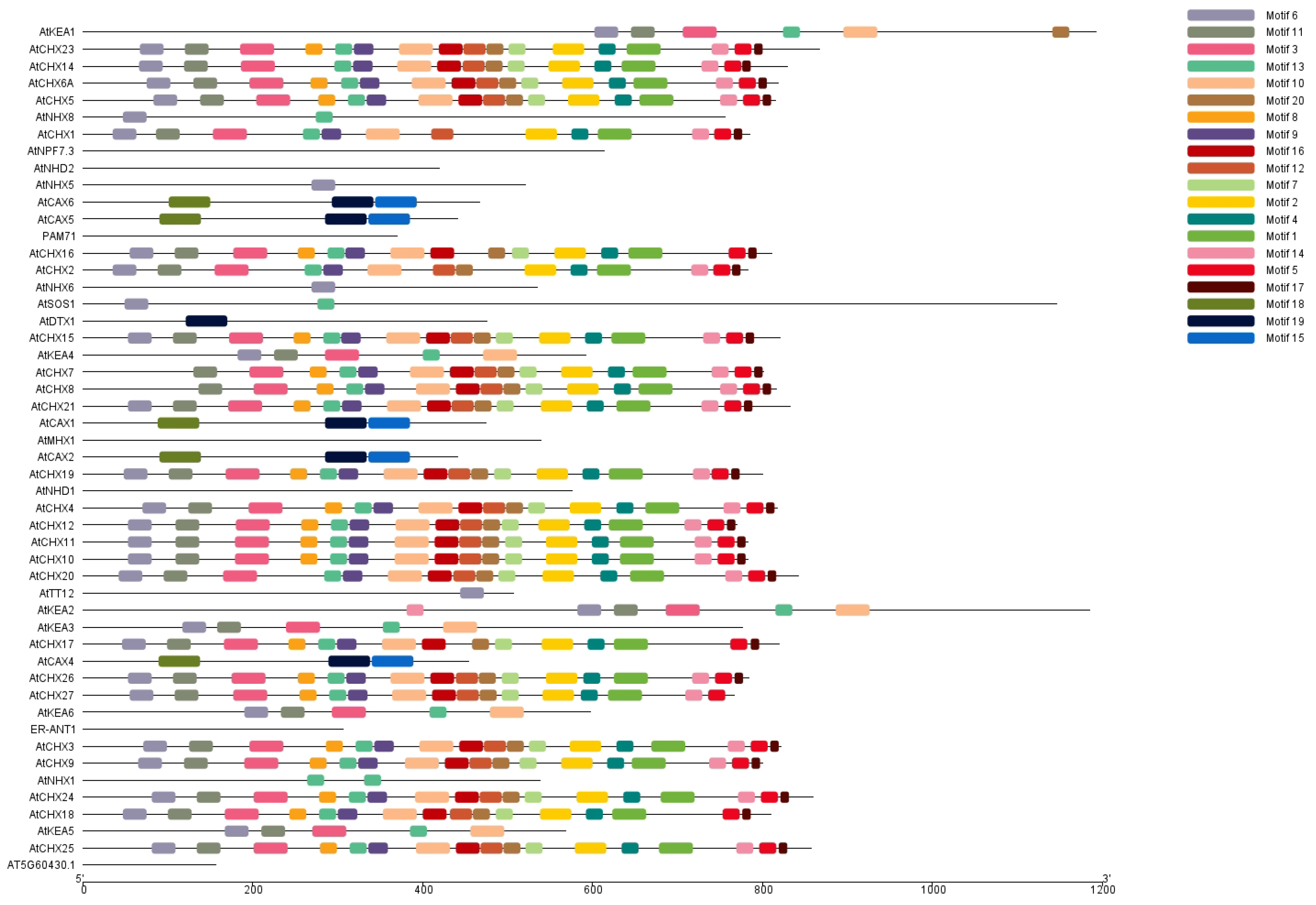
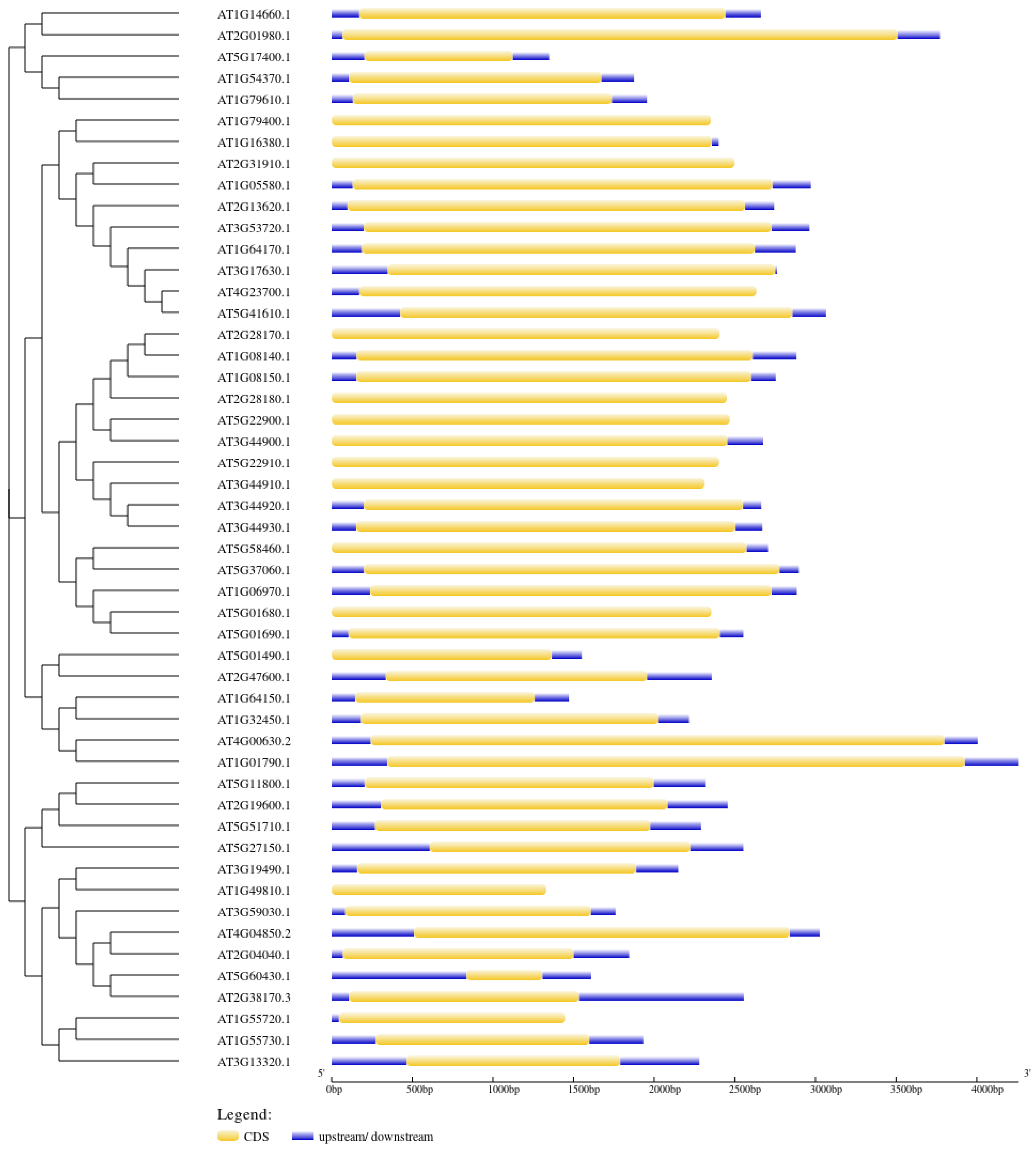
2.5. Motif Composition and Gene Structure Analysis
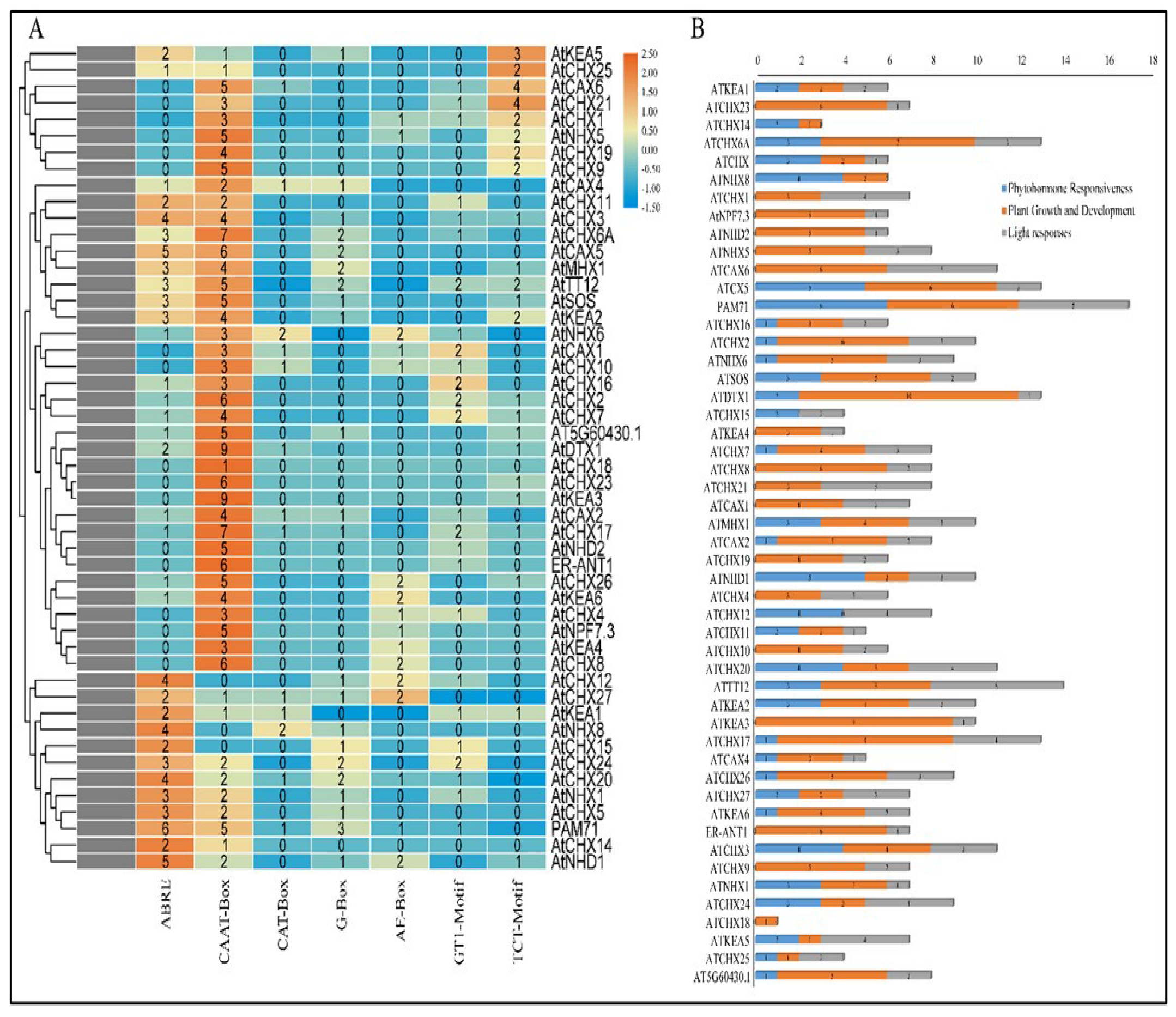
2.6. Cis-Elements Regulatory Analysis
2.7. Homology Modelling
2.8. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Identification of Antiporter Gene in Arabidopsis
4.2. Phylogenetic Analysis and Characterisation of Antiporter Proteins
4.3. Conserved Motif Analysis, Physiochemical Characterisation of Antiporter Genes
4.4. Chromosomal Distribution of Antiporter Genes
4.5. Cis-Element Regulatory Analysis and Homology Modelling
4.6. Plant Growth and CysNO Treatment
Real-Time PCR Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bassil, E.; Blumwald, E. The ins and outs of intracellular ion homeostasis: NHX-type cation/H+ transporters. Curr. Opin. Plant Biol. 2014, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.-Y.; Yang, X.; Xia, X.; Yin, W. Comparative analysis of cation/proton antiporter superfamily in plants. Gene 2013, 521, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Chanroj, S.; Wang, G.; Venema, K.; Zhang, M.W.; Delwiche, C.F.; Sze, H. Conserved and diversified gene families of monovalent cation/H+ antiporters from algae to flowering plants. Front. Plant Sci. 2012, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Fliegel, L. Structure and function of yeast and fungal Na+/H+ antiporters. IUBMB Life 2018, 70, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.; Saie, M.S., Jr. A novel family of ubiquitous heavy metal ion transport proteins. J. Membr. Biol. 1997, 156, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Chopin, F.; Orsel, M.; Dorbe, M.F.; Chardon, F.; Truong, H.N.; Miller, A.J.; Krapp, A.; Daniel-Vedele, F. The Arabidopsis ATNRT2. 7 nitrate transporter controls nitrate content in seeds. Plant Cell 2007, 19, 1590–1602. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.-S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Dechorgnat, J.; Nguyen, C.T.; Armengaud, P.; Jossier, M.; Diatloff, E.; Filleur, S.; Daniel-Vedele, F. From the soil to the seeds: The long journey of nitrate in plants. J. Exp. Bot. 2011, 62, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Little, D.Y.; Rao, H.; Oliva, S.; Daniel-Vedele, F.; Krapp, A.; Malamy, J.E. The putative high-affinity nitrate transporter NRT2. 1 represses lateral root initiation in response to nutritional cues. Proc. Natl. Acad. Sci. USA 2005, 102, 13693–13698. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, G. A nitrate transporter for both roots and shoots. Plant Cell 2012, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.D. Breathing new life into nitric oxide signaling: A brief overview of the interplay between oxygen and nitric oxide. Redox Biol. 2015, 5, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Castillo, M.-C.; Coego, A.; Costa-Broseta, Á.; León, J. Nitric oxide responses in Arabidopsis hypocotyls are mediated by diverse phytohormone pathways. J. Exp. Bot. 2018, 69, 5265–5278. [Google Scholar] [CrossRef] [PubMed]
- Arc, E.; Galland, M.; Godin, B.; Cueff, G.; Rajjou, L. Nitric oxide implication in the control of seed dormancy and germination. Front. Plant Sci. 2013, 4, 346. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Juste, J.; León, J. Nitric oxide regulates DELLA content and PIF expression to promote photomorphogenesis in Arabidopsis. Plant Physiol. 2011, 156, 1410–1423. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-C.; Delk, N.A.; Chowdhury, N.I.; Braam, J. Arabidopsis potential calcium sensors regulate nitric oxide levels and the transition to flowering. Plant Signal. Behav. 2007, 2, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, G.; Lokesh, V.; Neelwarne, B. Nitric oxide in fruit ripening: Trends and opportunities. Biotechnol. Adv. 2010, 28, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, M.; Kong, D.; Wang, L.; Lv, Q.; Wang, J.; Bao, F.; Gong, Q.; Xia, J.; He, Y. Nitric oxide induces cotyledon senescence involving co-operation of the NES1/MAD1 and EIN2-associated ORE1 signalling pathways in Arabidopsis. J. Exp. Bot. 2014, 65, 4051–4063. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Role of nitric oxide in tolerance of plants to abiotic stress. Protoplasma 2011, 248, 447–455. [Google Scholar] [CrossRef] [PubMed]
- da-Silva, C.J.; Modolo, L.V. Hydrogen sulfide: A new endogenous player in an old mechanism of plant tolerance to high salinity. Acta Bot. Bras. 2017, 32, 150–160. [Google Scholar] [CrossRef]
- Ruan, H.-H.; Shen, W.-B.; Xu, L.-L. Nitric oxide modulates the activities of plasma membrane H+-ATPase and PPase in wheat seedling roots and promotes the salt tolerance against salt stress. J. Integr. Plant Biol. 2004, 46, 415–422. [Google Scholar]
- Zhang, Y.; Wang, L.; Liu, Y.; Zhang, Q.; Wei, Q.; Zhang, W. Nitric oxide enhances salt tolerance in maize seedlings through increasing activities of proton-pump and Na+/H+ antiport in the tonoplast. Planta 2006, 224, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, W.-H.; Wu, F.-H.; He, E.-M.; Liu, X.; Shangguan, Z.-P.; Zheng, H.-L. Hydrogen sulfide enhances salt tol- erance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci. Rep. 2015, 5, 12516. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xiong, D.-Y.; Wang, W.-H.; Hu, W.-J.; Simon, M.; Xiao, Q.; Liu, T.-W.; Liu, X.; Zheng, H.-L. Nitric oxide mediates root K+/Na+ balance in a mangrove plant, Kandelia obovata, by enhancing the expression of AKT1-type K+ channel and Na+/H+ antiporter under high salinity. PLoS ONE 2013, 8, e71543. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Sottosanto, J.B.; Blumwald, E. Vacuolar cation/H+ exchange; ion homeostasis, and leaf development are altered in a T-DNA insertional mutant of AtNHX1, the Arabidopsis vacuolar Na+/H+ antiporter. Plant J. 2003, 36, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, M.; Fukada-Tanaka, S.; Hoshino, A.; Takada, J.; Inagaki, Y.; Iida, S. Characterization of a novel Na+/H+ antiporter gene InNHX2 and comparison of InNHX2 with InNHX1, which is responsible for blue flower coloration by increasing the vacuolar pH in the Japanese morning glory. Plant Cell Physiol. 2005, 46, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Leidi, E.O.; Pardo, J.M. How do vacuolar NHX exchangers function in plant salt tolerance? Plant Signal. Behav. 2010, 5, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.S. Plant and yeast NHX antiporters: Roles in membrane trafficking F. J. Integr. Plant Biol. 2012, 54, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, P.; Motes, C.M.; Park, S.; Hirschi, K.D. CHX 14 is a plasma membrane K-efflux transporter that regulates K+ redistribution in A rabidopsis thaliana. Plant Cell Environ. 2015, 38, 2223–2238. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.; Evans, A.; Newbury, H.; Pritchard, J. Functional analysis of CHX21: A putative sodium transporter in Arabidopsis. J. Exp. Bot. 2006, 57, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Chanroj, S.; Zulkifli, L.; Johnson, M.A.; Uozumi, N.; Cheung, A.; Sze, H. Pollen tubes lacking a pair of K+ trans-porters fail to target ovules in Arabidopsis. Plant Cell 2011, 23, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Qi, K.; Liu, X.; Yin, H.; Wang, P.; Chen, J.; Wu, J.; Zhang, S. Genome-wide identification and comparative analysis of the cation proton antiporters family in pear and four other Rosaceae species. Mol. Genet. Genom. 2016, 291, 1727–1742. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.M.; Nawaz, M.A.; Shah, Z.H.; Daur, I.; Khatoon, S.; Yang, S.H.; Chung, G. In-depth genomic and transcriptomic analysis of five K+ transporter gene families in soybean confirm their differential expression for nodulation. Front. Plant Sci. 2017, 8, 804. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Mun, B.-G.; Imran, Q.M.; Lee, S.-U.; Adamu, T.A.; Shahid, M.; Kim, K.-M.; Yun, B.-W. Nitric oxide mediatedtranscriptome profiling reveals activation of multiple regulatory pathways in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 975. [Google Scholar] [CrossRef] [PubMed]
- Feechan, A.; Kwon, E.; Yun, B.W.; Wang, Y.Q.; Pallas, J.A.; Loake, G.J. A central role for S-nitrosothiols in plant disease resistance. Proc. Natl. Acad. Sci. USA 2005, 102, 8054–8059. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.; Feechan, A.; Yun, B.W.; Hwang, B.H.; Pallas, J.A.; Kang, J.G.; Loake, G.J. AtGSNOR1 function is required formultiple developmental programs in Arabidopsis. Planta 2012, 236, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Leshem, Y.; Wills, R. Harnessing senescence delaying gases nitric oxide and nitrous oxide: A novel approach to post-harvest control of fresh horticultural produce. Biol. Plant. 1998, 41, 1–10. [Google Scholar] [CrossRef]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [PubMed]
- Ahern, G.P.; Klyachko, V.A.; Jackson, M.B. cGMP and S-nitrosylation: Two routes for modulation of neuronal excitability by NO. Trends Neurosci. 2002, 25, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Lamotte, O.; Courtois, C.; Dobrowolska, G.; Besson, A.; Pugin, A.; Wendehenne, D. Mechanisms of nitric-oxide-induced increase of free cytosolic Ca2+ concentration in Nicotiana plumbaginifolia cells. Free Radic. Biol. Med. 2006, 40, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33 (Suppl. S2), W116–W120. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.; Johnson, J.; Grant, C. Noble. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35 (Suppl. S2), W585–W587. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Gene ID | Length | MW | pI | GRAVY | Aliphatic Index | TM | Subcellular Localisation |
|---|---|---|---|---|---|---|---|---|
| 1 | AT1G01790.1 | 1193 | 128,033.50 | 5.22 | 0.080 | 105.80 | 10 | plas |
| 2 | AT1G05580.1 | 867 | 95,867.43 | 6.02 | 0.246 | 99.64 | 12 | plas |
| 3 | AT1G06970.1 | 829 | 92,159.60 | 6.56 | 0.335 | 109.5 | 12 | plas |
| 4 | AT1G08140.1 | 818 | 93,395.50 | 7.27 | 0.146 | 97.21 | 10 | plas |
| 5 | AT1G08150.1 | 815 | 91,606.19 | 6.56 | 0.328 | 106.50 | 10 | plas |
| 6 | AT1G14660.1 | 756 | 83,475.75 | 6.58 | 0.273 | 108.94 | 12 | plas |
| 7 | AT1G16380.1 | 785 | 88,908.78 | 6.31 | 0.295 | 107.62 | 12 | plas |
| 8 | AT1G32450.1 | 614 | 68,699.10 | 5.50 | 0.076 | 88.93 | 11 | plas |
| 9 | AT1G49810.1 | 420 | 45,446.85 | 6.19 | 0.823 | 124.19 | 9 | vacu |
| 10 | AT1G54370.1 | 521 | 57,335.14 | 4.98 | 0.444 | 96.89 | 10 | plas |
| 11 | AT1G55720.1 | 467 | 51,837.90 | 5.44 | 0.530 | 113.58 | 10 | plas |
| 12 | AT1G55730.1 | 441 | 48,096.18 | 4.86 | 0.563 | 114.54 | 11 | Plas/vacu |
| 13 | AT1G64150.1 | 370 | 39,073.58 | 4.88 | 0.462 | 104.19 | 7 | Plas/chlo |
| 14 | AT1G64170.1 | 811 | 88,050.60 | 8.82 | 0.404 | 111.36 | 10 | plas |
| 15 | AT1G79400.1 | 783 | 88,204.04 | 6.39 | 0.290 | 108.15 | 12 | plas |
| 16 | AT1G79610.1 | 535 | 59,314.39 | 5.68 | 0.377 | 97.48 | 9 | plas |
| 17 | AT2G01980.1 | 1146 | 127,188.31 | 7.62 | 0.098 | 105.30 | 9 | Plas |
| 18 | AT2G04040.1 | 476 | 51,844.92 | 7.88 | 0.703 | 119.62 | 12 | Vacu/plas |
| 19 | AT2G13620.1 | 821 | 89,859.54 | 5.71 | 0.363 | 109.93 | 12 | plas |
| 20 | AT2G19600.1 | 592 | 64,249.42 | 5.91 | 0.589 | 118.72 | 12 | plas |
| 21 | AT2G28170.1 | 801 | 90,989.71 | 7.29 | 0.261 | 104.84 | 10 | plas |
| 22 | AT2G28180.1 | 816 | 90,956.09 | 7.22 | 0.384 | 116.81 | 12 | plas |
| 23 | AT2G31910.1 | 832 | 91,982.03 | 5.38 | 0.256 | 102.43 | 12 | plas |
| 24 | AT2G38170.3 | 475 | 51,636.65 | 5.92 | 0.604 | 124.42 | 8 | Plas/vacu |
| 25 | AT2G47600.1 | 539 | 59,792.38 | 5.84 | 0.413 | 111.41 | 10 | plas |
| 26 | AT3G13320.1 | 441 | 48,215.38 | 4.72 | 0.573 | 114.10 | 10 | plas/vacu |
| 27 | AT3G17630.1 | 800 | 86,915.25 | 8.76 | 0.370 | 112 | 12 | plas |
| 28 | AT3G19490.1 | 576 | 61,368.48 | 5.49 | 0.528 | 112.26 | 8 | Plas/chlo |
| 29 | AT3G44900.1 | 817 | 92,007.93 | 8.51 | 0.337 | 107.85 | 10 | nucl |
| 30 | AT3G44910.1 | 770 | 85,819.50 | 6.02 | 0.381 | 115.16 | 13 | plas |
| 31 | AT3G44920.1 | 783 | 88,432.71 | 6.16 | 0.406 | 118.35 | 11 | plas |
| 32 | AT3G44930.1 | 783 | 88,143.29 | 6.09 | 0.397 | 118.10 | 11 | plas |
| 33 | AT3G53720.1 | 842 | 91,553.09 | 8.93 | 0.276 | 110.20 | 10 | plas |
| 34 | AT3G59030.1 | 507 | 55,147.11 | 8.36 | 0.745 | 120.18 | 12 | plas |
| 35 | AT4G00630.2 | 1185 | 127,605.10 | 5.11 | 0.086 | 105.22 | 10 | plas |
| 36 | AT4G04850.2 | 776 | 83,790.69 | 5.53 | 0.369 | 111.69 | - | plas |
| 37 | AT4G23700.1 | 820 | 89,165.51 | 8.06 | 0.372 | 112.80 | 12 | plas |
| 38 | AT5G01490.1 | 454 | 49,609.17 | 6.08 | 0.519 | 117.91 | 10 | plas |
| 39 | AT5G01680.1 | 784 | 87,038.08 | 7.02 | 0.256 | 106.98 | 10 | plas |
| 40 | AT5G01690.1 | 767 | 86,990.99 | 8.74 | 0.396 | 113.82 | 11 | plas |
| 41 | AT5G11800.1 | 597 | 64,391.65 | 7.10 | 0.599 | 125.53 | 12 | plas |
| 42 | AT5G17400.1 | 306 | 33,788.18 | 9.82 | 0.041 | 88.99 | 4 | cyto/cyto_E.R |
| 43 | AT5G22900.1 | 822 | 92,452.77 | 7.50 | 0.277 | 101.50 | 11 | plas |
| 44 | AT5G22910.1 | 800 | 89,059.65 | 7.90 | 0.359 | 118.41 | 11 | plas |
| 45 | AT5G27150.1 | 538 | 59,513.42 | 6.73 | 0.458 | 106.71 | 12 | vacu |
| 46 | AT5G37060.1 | 859 | 96,680.97 | 6.34 | 0.115 | 95.17 | 10 | plas |
| 47 | AT5G41610.1 | 810 | 87,383.34 | 8.71 | 0.398 | 113.38 | 12 | plas |
| 48 | AT5G51710.1 | 568 | 61,598.40 | 5.84 | 0.613 | 123.86 | 11 | plas |
| 49 | AT5G58460.1 | 857 | 95,833.29 | 8.32 | 0.118 | 94.39 | 11 | plas |
| 50 | AT5G60430.1 | 156 | 17,439.83 | 5.76 | −0.114 | 78.65 | - | chlo |
| Sr. No. | Gene | Primer Name | Sequence | Amplicon Size (bp) |
|---|---|---|---|---|
| 1 | AT1G16380 | AtCHX1_L1 | ATTGTCGGAGACGCGGTGAC | 177 |
| AtCHX1_R1 | CAGCGTCTGCATTCCGTTGC | |||
| 2 | AT1G79400 | AtCHX2_L1 | CGCGGTGCGATCTCTCTTGT | 145 |
| AtCHX2_R1 | TGCCTCCATCTTCGTCGTGC | |||
| 3 | AT2G38170 | ATCAX1_L1 | ACGGCGAAAGGATCGAGCAG | 233 |
| ATCAX1_R1 | CAAGGCTGACTGACGCCACA | |||
| 4 | AT2G47600 | ATMHX1_L1 | GTCAGCCATCACTGCACGGT | 218 |
| ATMHX1_R1 | ACCGCTCCCCCATATTCCGT | |||
| 5 | AT3G13320 | AtCAX2_L1 | CAGCTGAACATGCAGGGGCT | 115 |
| AtCAX2_R1 | ACGCAGAATGGGACCGCAAA | |||
| 6 | AT4G23700 | ATCHX17_L1 | ATTTGCCAAAGCGCGGAACG | 267 |
| ATCHX17_R1 | CGCAAACGCTAACGCCTCAC | |||
| 7 | AT5G11800 | ATKEA6_L1 | TCGCCTTTGCTTGTGCTGGA | 280 |
| ATKEA6_R1 | TCCGAACGTTTACCACCGCA | |||
| 8 | AT5G17400 | ER-ANT1_L1 | CCTCTGCAGGCGTCATTGCT | 192 |
| ER-ANT1_R1 | CAAGCACTCCTGCTCCTGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amir, R.; Qayyum, Z.; Hussain, S.; Yun, B.-W.; Hussain, A.; Mun, B.-G. Arabidopsis Antiporter Genes as Targets of NO Signalling: Phylogenetic, Structural, and Expression Analysis. Int. J. Mol. Sci. 2025, 26, 7195. https://doi.org/10.3390/ijms26157195
Amir R, Qayyum Z, Hussain S, Yun B-W, Hussain A, Mun B-G. Arabidopsis Antiporter Genes as Targets of NO Signalling: Phylogenetic, Structural, and Expression Analysis. International Journal of Molecular Sciences. 2025; 26(15):7195. https://doi.org/10.3390/ijms26157195
Chicago/Turabian StyleAmir, Rabia, Zuhra Qayyum, Sajeel Hussain, Byung-Wook Yun, Adil Hussain, and Bong-Gyu Mun. 2025. "Arabidopsis Antiporter Genes as Targets of NO Signalling: Phylogenetic, Structural, and Expression Analysis" International Journal of Molecular Sciences 26, no. 15: 7195. https://doi.org/10.3390/ijms26157195
APA StyleAmir, R., Qayyum, Z., Hussain, S., Yun, B.-W., Hussain, A., & Mun, B.-G. (2025). Arabidopsis Antiporter Genes as Targets of NO Signalling: Phylogenetic, Structural, and Expression Analysis. International Journal of Molecular Sciences, 26(15), 7195. https://doi.org/10.3390/ijms26157195