
Feasibility of Manufacturing and Antitumor Activity of TIL for Advanced Endometrial Cancers
Abstract
1. Introduction
2. Results
2.1. Successful Manufacturing of Endometrial TIL
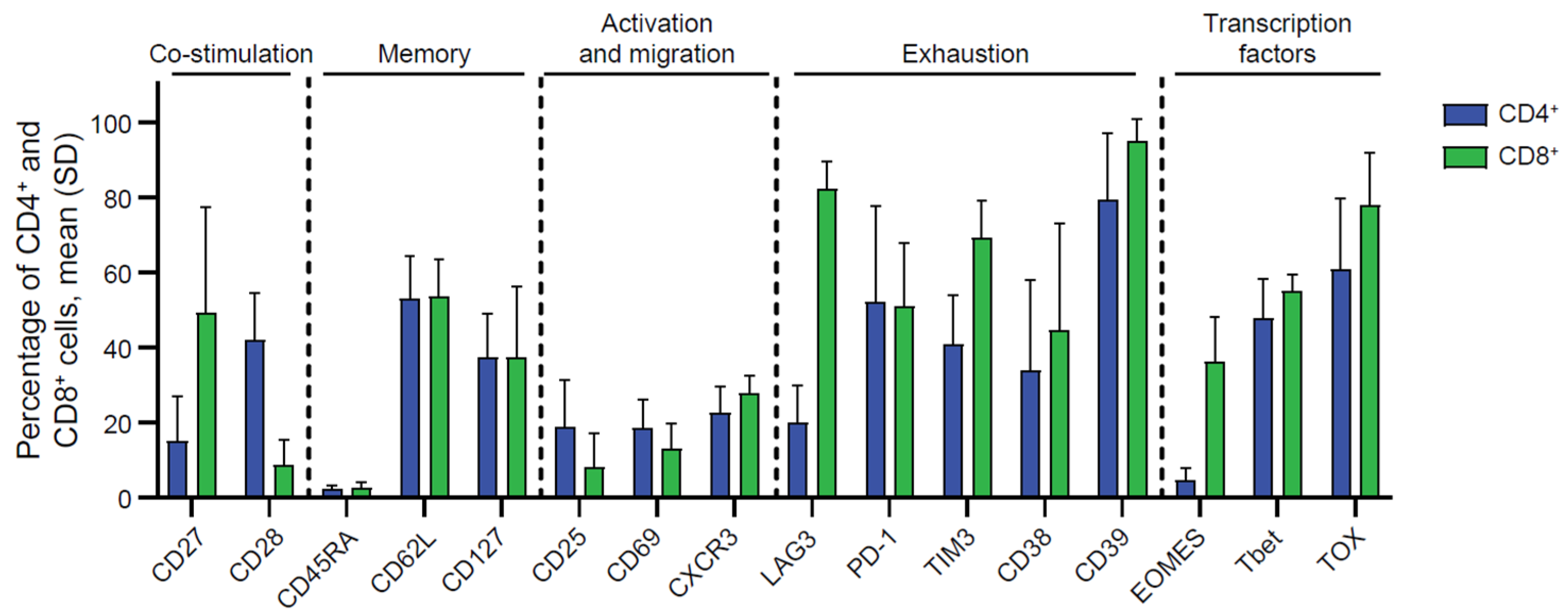
2.2. Phenotypic Characterization of Endometrial TILs
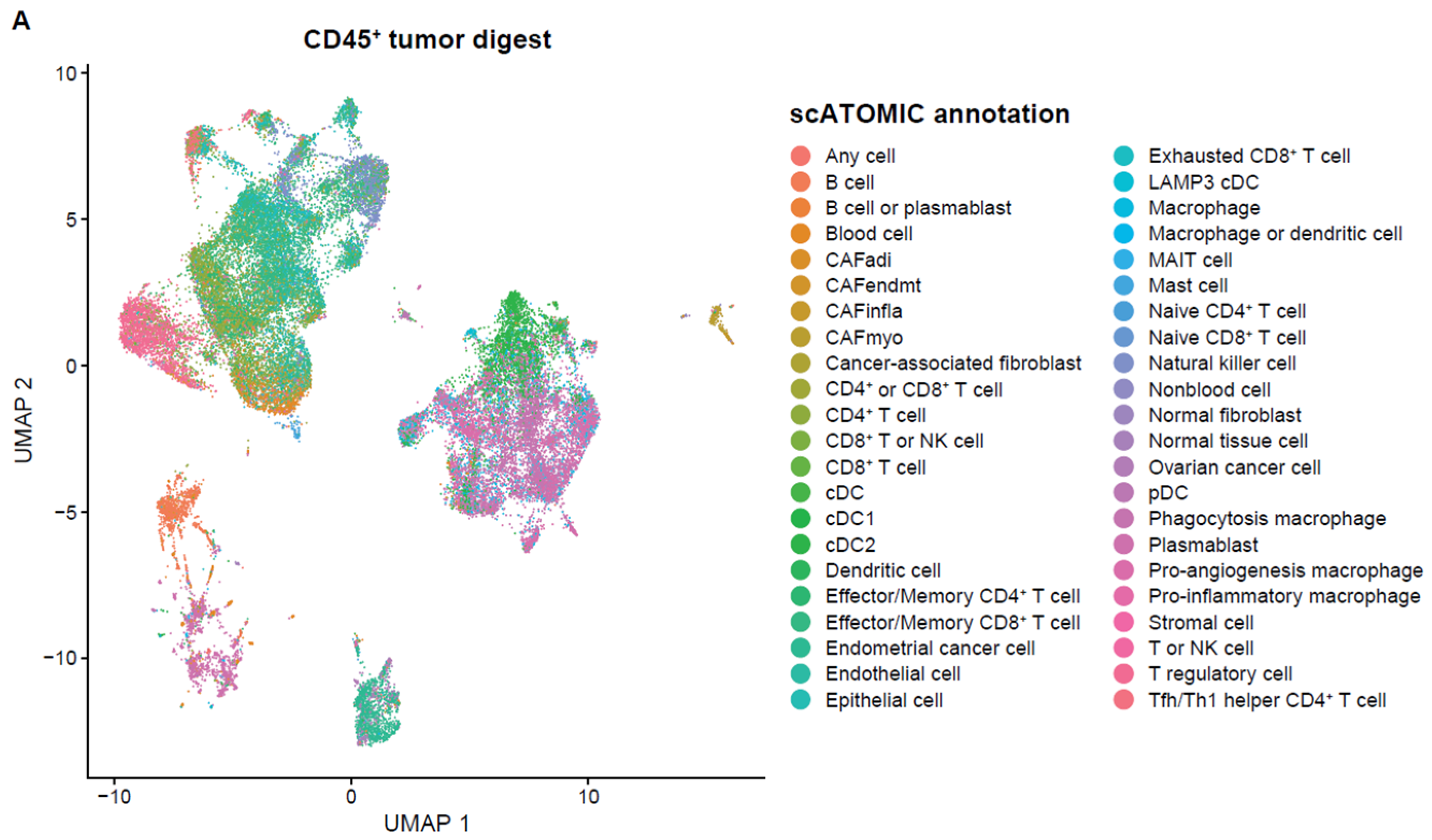
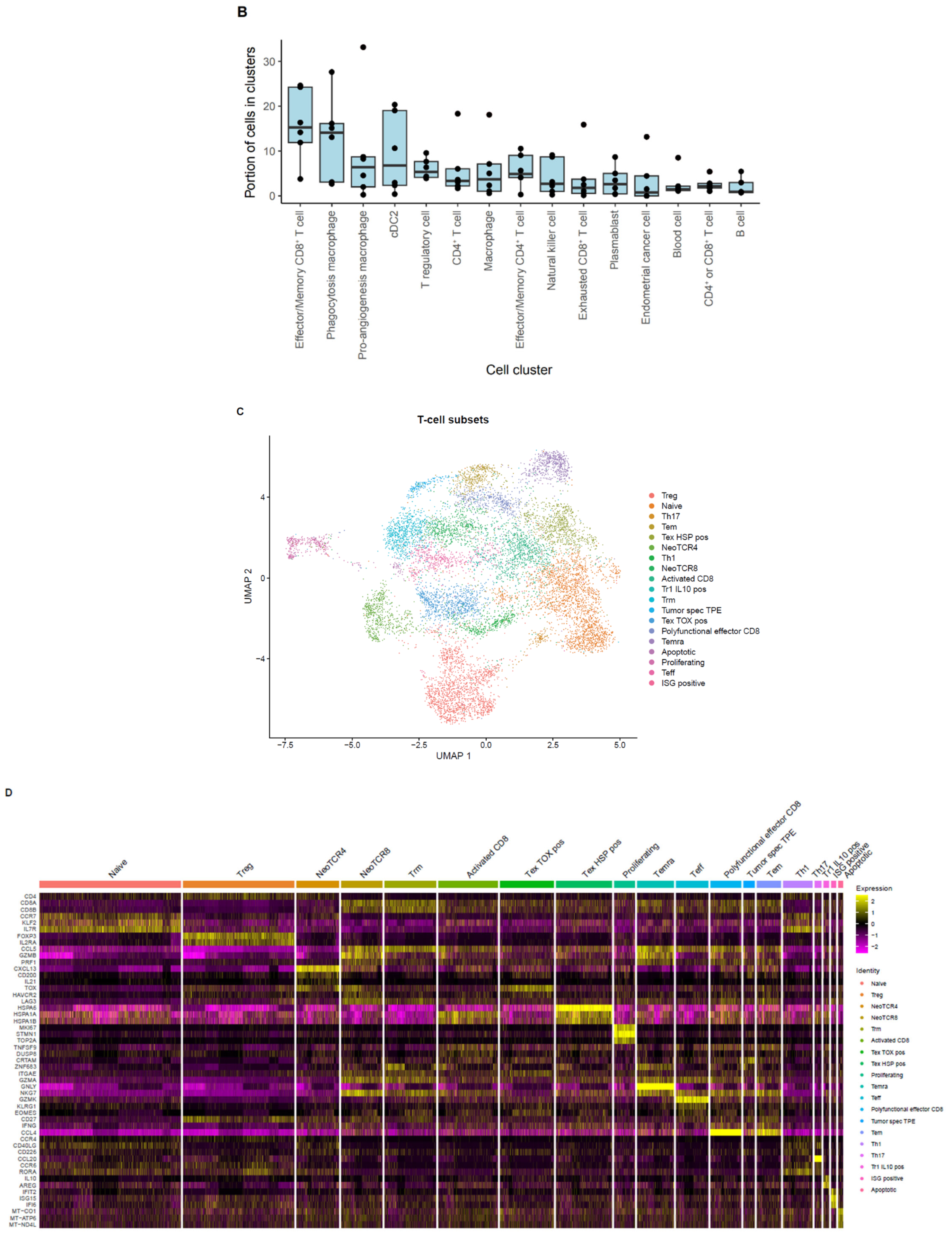
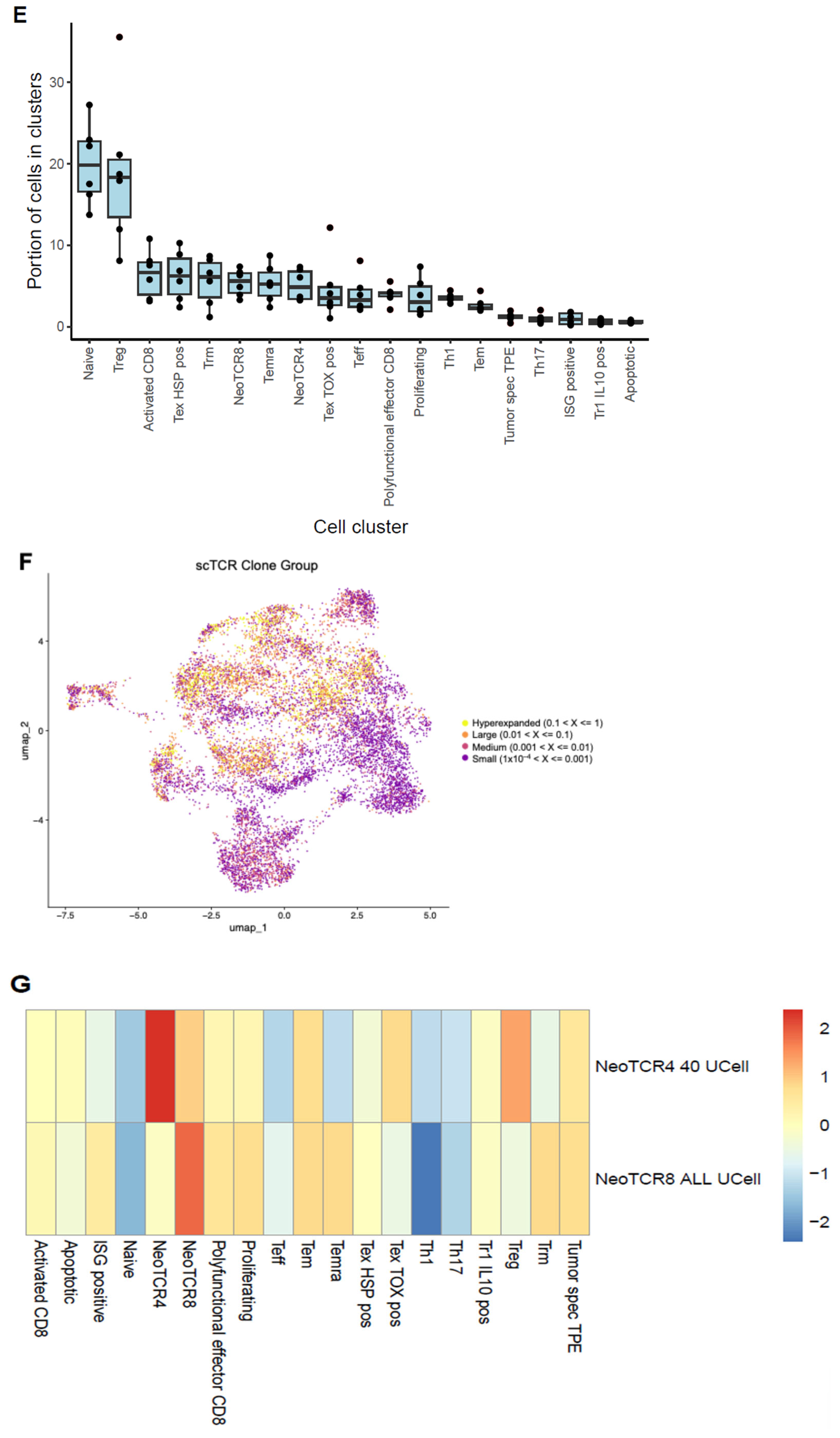
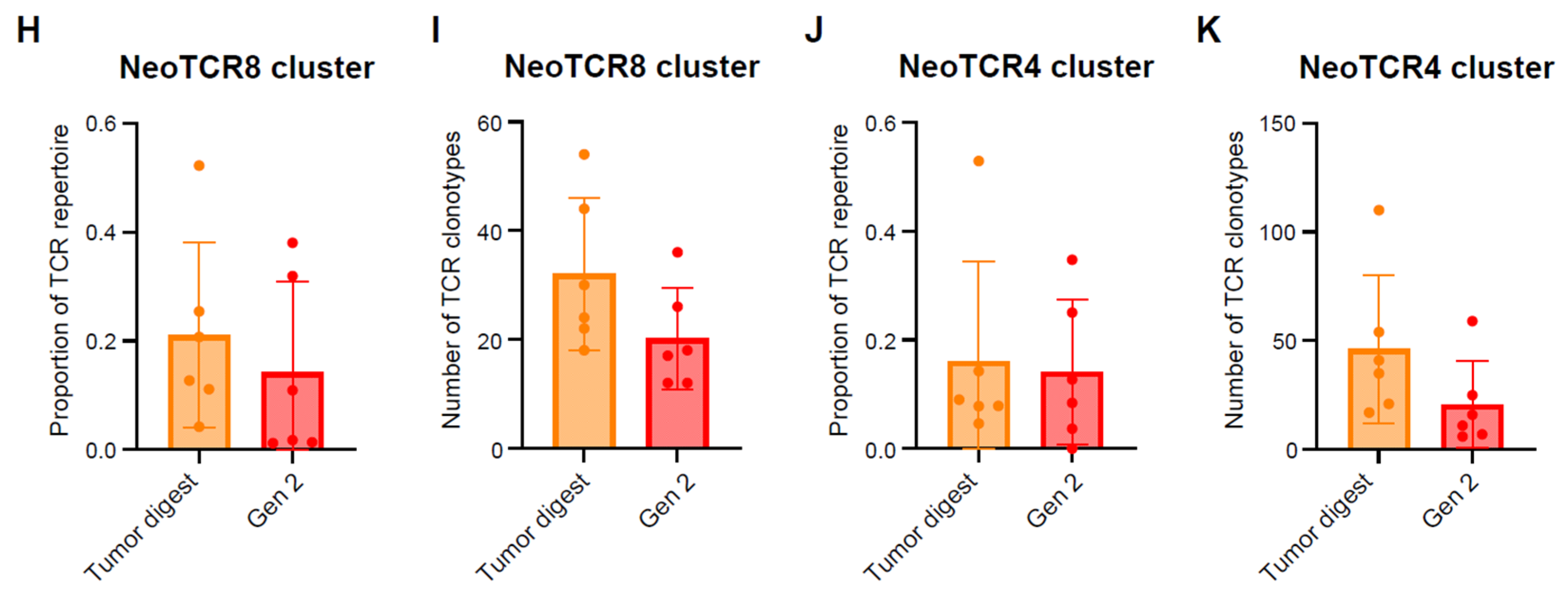
2.3. Single-Cell Analysis of Endometrial TILs
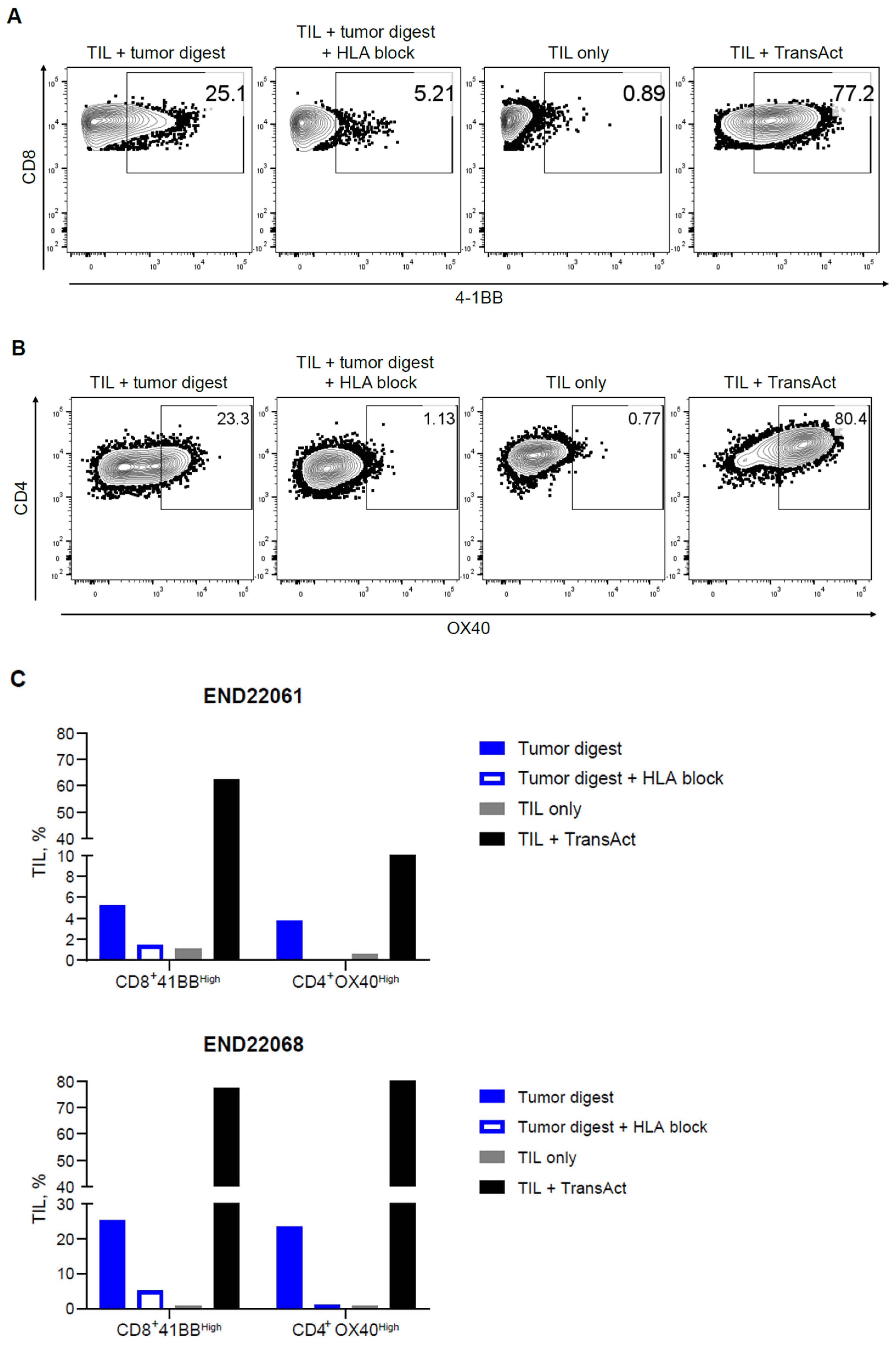
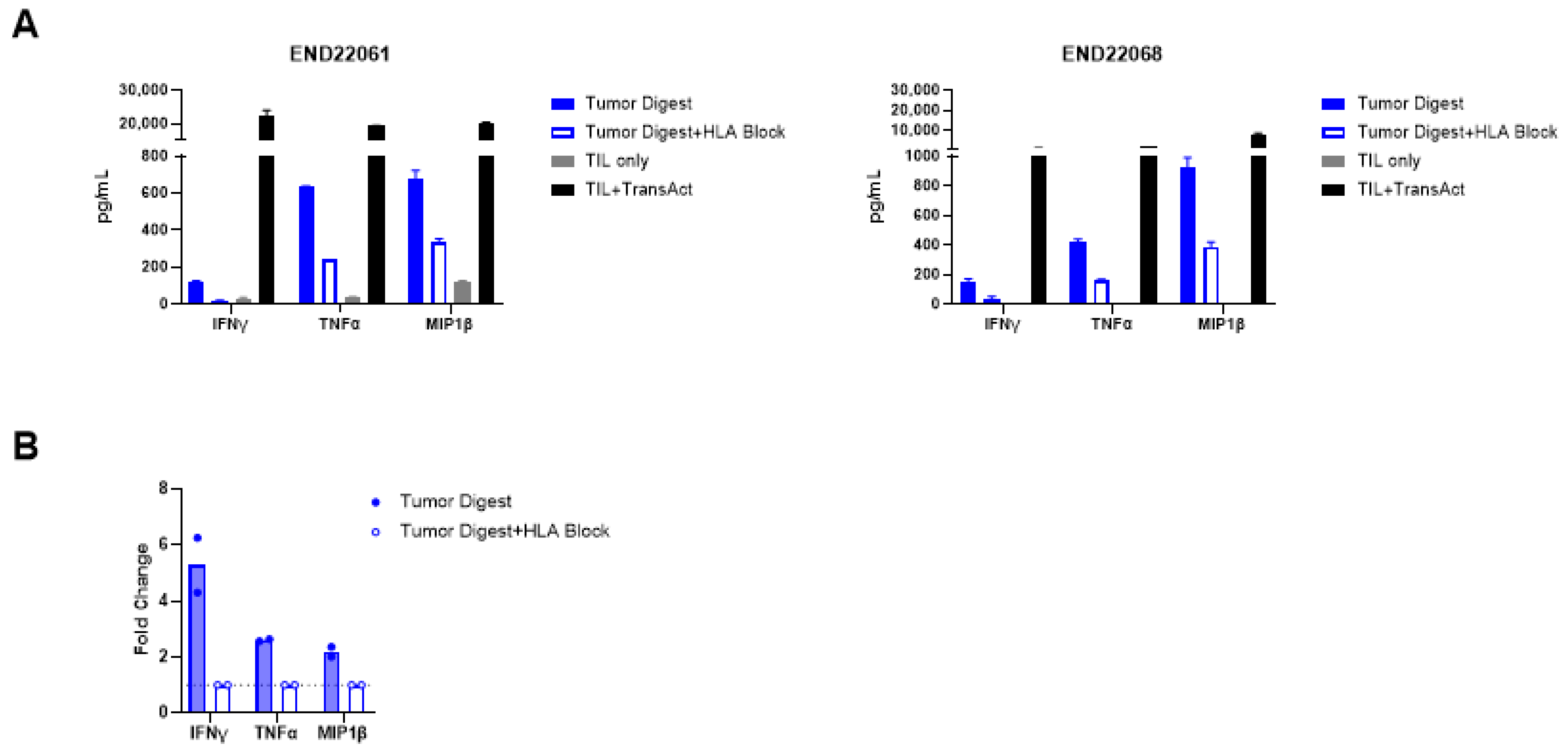
2.4. Functional Evaluation of Endometrial TIL Reactivity
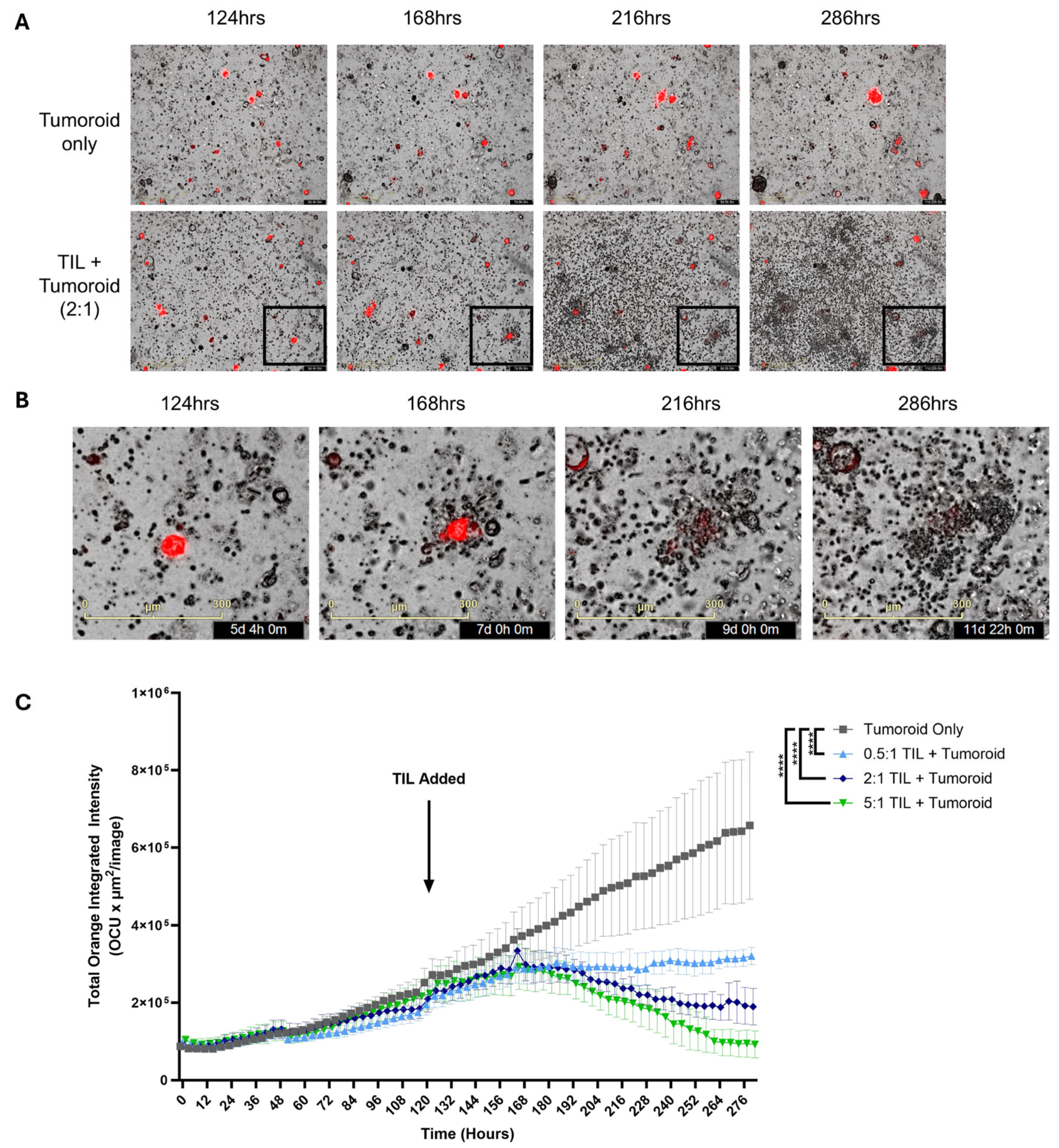
2.5. In Vitro Antitumor Activity of Endometrial TIL
2.6. Full-Scale Manufacturing of the Endometrial TIL Drug Products
3. Discussion
4. Materials and Methods
4.1. Tumor Processing for Expansion of TILs and Immunophenotyping
4.2. TIL Reactivity Assay
4.3. Single-Cell RNA and Bulk TCR Sequencing
4.4. Tumor Organoid Establishment and TIL Co-Culture Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACT | Adoptive cell therapy |
| ANOVA | Analysis of variance |
| BSA | Bovine serum albumin |
| CDR3 | Complementarity determining region 3 |
| cDC | Conventional dendritic cell |
| cDC1 | Conventional dendritic cell type 1 |
| cDC2 | Conventional dendritic cell type 2 |
| dMMR | Mismatch repair-deficient |
| DP | Drug product |
| EC | Endometrial cancer |
| ET | Effector-to-target |
| FDA | US Food and Drug Administration |
| FIGO | International Federation of Gynecology and Obstetrics |
| Gen2 | Generation 2 |
| HLA | Human leukocyte antigen |
| ICI | Immune checkpoint inhibitor |
| ICOS | Inducible T-cell costimulator |
| IFN-γ | Interferon-gamma |
| IL-2 | Interleukin-2 |
| ISG | Interferon-stimulated gene |
| IU | International units |
| LAG3 | Lymphocyte activation gene-3 |
| LAMP3 | Lysosome-associated membrane protein 3 |
| MHC | Major histocompatibility complex |
| MIP-1β | Macrophage inflammatory protein beta |
| MMR | Mismatch repair |
| MSI-H | Microsatellite instability-high |
| N/A | Not applicable |
| NeoTCR | Neoantigen-reactive T cells |
| NIR | Near-infrared spectroscopy |
| NK | Natural killer |
| NSCLC | Non-small cell lung cancer |
| PBMC | Peripheral blood mononuclear cells |
| PBS | Phosphate-buffered saline |
| pDC | Plasmacytoid dendritic cell |
| PD-1 | Programmed cell death protein-1 |
| pMMR | Mismatch repair-proficient |
| pNx | No lymph node was collected |
| pos | Positive |
| QC | Quality control |
| REP | Rapid expansion protocol |
| scTCR | Single cell T-cell receptor |
| SD | Standard deviation |
| SEM | Standard error of the mean |
| TCR | T-cell receptor |
| Tem | Effector memory T cells |
| Tfh | Follicular helper T cells |
| Th1 | T helper 1 cells |
| TIL | Tumor-infiltrating lymphocytes |
| TIM3 | T-cell immunoglobulin and mucin domain-containing protein 3 |
| TME | Tumor microenvironment |
| TNF-α | Tumor necrosis factor-alpha |
| TNM | Tumor, node, and metastasis |
| Treg | Regulatory T cells |
| TRM | Tissue-resident memory T cells |
| TVC | Total viable cells |
| UMAP | Uniform Manifold Approximation and Projection |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor ID | Tumor Site | Age | Race/ Ethnicity | Pathology | FIGO Grade | TNM Classification | MMR | MLH1 Promoter Methylation |
|---|---|---|---|---|---|---|---|---|
| END22061 | Uterus | 61 | White | Endometrioid adenocarcinoma | FIGO grade II | pT3a, pN0 | pMMR | |
| END22062 | Uterus | 68 | White | Endometrioid adenocarcinoma | FIGO grade II | pT1b, pN0 | dMMR | 81% |
| END22063 | Uterus | 67 | White | Endometrioid adenocarcinoma | FIGO grade II | pT1b, pN0 | dMMR | 97% |
| END22091 | Uterus | 83 | White | Endometrioid carcinoma | FIGO grade II | pT1a, pN0 | pMMR | |
| END22093 | Uterus | 63 | White | Endometrioid carcinoma | FIGO grade II | pT1b, pN0 | pMMR | |
| END22094 | Uterus | 66 | White | Endometrioid adenocarcinoma | FIGO grade I | pT1b, pN0 | pMMR | |
| END22095 | Uterus | 81 | White | Serous carcinoma | High grade (grade III) | pT3a, pN0 | pMMR | |
| END22096 | Uterus | 45 | White | Endometrioid carcinoma | FIGO grade II | pT3b, pNx | dMMR | Positive |
| END22098 | Uterus | 67 | Hispanic | Endometrioid carcinoma | FIGO grade II | pT1a, pN0 | dMMR | Positive |
| END22100 | Uterus | 69 | White | Endometrioid carcinoma | FIGO grade II | pT1a, pN0 | dMMR | MLH1 intact |
| END22102 | Uterus | 84 | White | Endometrioid carcinoma | FIGO grade III | pT1a, pNx | pMMR | |
| END22111 | Uterus | 69 | Not available | Endometrioid | FIGO grade I | pT1a, pN0 | pMMR | |
| END22133 | Uterus | 49 | Not available | Endometrioid carcinoma | FIGO grade II | pT1a, pN0 | dMMR | MLH1 intact |
| END22134 | Uterus | 35 | Not available | Endometrioid adenocarcinoma | FIGO grade II | pT1a, pN0 | pMMR | |
| END22135 | Uterus | 49 | White | Adenocarcinoma | Poorly differentiated | T1bN1M0 | Not available |
| Sample | Cell Count Before Filter | Cell Count After Filter | After T-Cell Subset |
|---|---|---|---|
| END22094 | 6106 | 5408 | 1830 |
| END22095 | 6413 | 5878 | 1655 |
| END22096 | 7722 | 6612 | 1705 |
| END22098 | 12,495 | 10,544 | 4761 |
| END22100 | 5498 | 4798 | 732 |
| END22102 | 4376 | 3814 | 667 |
| Media Component | Working Concentration | Manufacturer | Catalog # |
|---|---|---|---|
| 17B-Estradiol-Water Soluble | 10 mg/mL | Sigma Aldrich (St. Louis, MO, USA) | E4389 |
| A83-01 | 5 mM | Sigma Aldrich | SML0788-5MG |
| B27 (50X) | 1X | Gibco (Waltham, MA, USA) | 17504-044 |
| Chemically defined lipid concentrate | 1% | Life technologies (Carlsbad, CA, USA) | 11905031 |
| EGF | 50 ng/mL | Peprotech (Cranbury, NJ, USA) | AF-100-15 |
| GlutaMAX™ (100X) | 1X | Life technologies | 35050061 |
| HGF | 20 ng/mL | Peprotech | 100-39 |
| IGF-1 | 40 ng/mL | Peprotech | 100-11 |
| N2 (100X) | 1X | Life technologies | 17502001 |
| N-acetyl-cysteine | 1.25 mM | Sigma Aldrich | A7250-50G |
| Nicotinamide | 5 mM | Sigma Aldrich | 73240-100G |
| Noggin | 10 ng/mL | Peprotech | 120-10C |
| RH R-Spondin 3 | 25 ng/mL | R&D Systems | 3500-RS-025/CF |
| SB202190 | 100 nM | Sigma Aldrich | S7067-5MG |
| Y-27632 | 10 µM | Selleckchem (Houston, TX, USA) | S1049 |
| Anti-Anti (100X) | 1X | Gibco | 15-240-062 |
| Advance DMEM/F12 | n/a | Invitrogen | 12634028 |
References
- Crosbie, E.J.; Kitson, S.J.; McAlpine, J.N.; Mukhopadhyay, A.; Powell, M.E.; Singh, N. Endometrial cancer. Lancet 2022, 399, 1412–1428. [Google Scholar] [CrossRef] [PubMed]
- Eskander, R.N.; Sill, M.W.; Beffa, L.; Moore, R.G.; Hope, J.M.; Musa, F.B.; Mannel, R.; Shahin, M.S.; Cantuaria, G.H.; Girda, E.; et al. Pembrolizumab plus Chemotherapy in Advanced Endometrial Cancer. N. Engl. J. Med. 2023, 388, 2159–2170. [Google Scholar] [CrossRef] [PubMed]
- Mirza, M.R.; Chase, D.M.; Slomovitz, B.M.; de Pont Christensen, R.; Novák, Z.; Black, D.; Gilbert, L.; Sharma, S.; Valabrega, G.; Landrum, L.M.; et al. Dostarlimab for Primary Advanced or Recurrent Endometrial Cancer. N. Engl. J. Med. 2023, 388, 2145–2158. [Google Scholar] [CrossRef]
- Westin, S.N.; Moore, K.; Chon, H.S.; Lee, J.Y.; Thomes Pepin, J.; Sundborg, M.; Shai, A.; de la Garza, J.; Nishio, S.; Gold, M.A.; et al. Durvalumab plus carboplatin/paclitaxel followed by maintenance durvalumab with or without olaparib as first-line treatment for advanced endometrial cancer: The phase III DUO-E trial. J. Clin. Oncol. 2024, 42, 283–299. [Google Scholar] [CrossRef]
- Sarnaik, A.A.; Hamid, O.; Khushalani, N.I.; Lewis, K.D.; Medina, T.; Kluger, H.M.; Thomas, S.S.; Domingo-Musibay, E.; Pavlick, A.C.; Whitman, E.D.; et al. Lifileucel, a tumor-infiltrating lymphocyte therapy, in metastatic melanoma. J. Clin. Oncol. 2021, 39, 2656–2666. [Google Scholar] [CrossRef]
- Schoenfeld, A.J.; Lee, S.M.; Doger de Spéville, B.; Gettinger, S.N.; Häfliger, S.; Sukari, A.; Papa, S.; Rodríguez-Moreno, J.F.; Graf Finckenstein, F.; Fiaz, R.; et al. Lifileucel, an Autologous Tumor-Infiltrating Lymphocyte Monotherapy, in Patients with Advanced Non-Small Cell Lung Cancer Resistant to Immune Checkpoint Inhibitors. Cancer Discov. 2024, 14, 1389–1402. [Google Scholar] [CrossRef]
- Muul, L.M.; Spiess, P.J.; Director, E.P.; Rosenberg, S.A. Identification of specific cytolytic immune responses against autologous tumor in humans bearing malignant melanoma. J. Immunol. 1987, 138, 989–995. [Google Scholar] [CrossRef]
- Pockaj, B.A.; Sherry, R.M.; Wei, J.P.; Yannelli, J.R.; Carter, C.S.; Leitman, S.F.; Carasquillo, J.A.; Steinberg, S.M.; Rosenberg, S.A.; Yang, J.C. Localization of 111indium-labeled tumor infiltrating lymphocytes to tumor in patients receiving adoptive immunotherapy. Augmentation with cyclophosphamide and correlation with response. Cancer 1994, 73, 1731–1737. [Google Scholar] [CrossRef]
- Dudley, M.E.; Wunderlich, J.R.; Robbins, P.F.; Yang, J.C.; Hwu, P.; Schwartzentruber, D.J.; Topalian, S.L.; Sherry, R.; Restifo, N.P.; Hubicki, A.M.; et al. Cancer regression and autoimmunity in patients after clonal repopulation with antitumor lymphocytes. Science 2002, 298, 850–854. [Google Scholar] [CrossRef]
- Available online: https://www.fda.gov/drugs/resources-information-approved-drugs/fda-grants-accelerated-approval-lifileucel-unresectable-or-metastatic-melanoma (accessed on 20 June 2025).
- Wardell, S.; Frank, I.; Karyampudi, L.; Veerapathran, A.; Simpson-Abelson, M.; Lienlaf Moreno, M.; Blaskovich, M.; Wypych, J.; Fardis, M. A cryopreserved tumor infiltrating lymphocyte (TIL) product for LN-144, generated with an abbreviated method suitable for high throughput commercial manufacturing exhibits favorable quality attributes for adoptive cell transfer [poster]. In Proceedings of the Annual Meeting of the Society for Immunotherapy of Cancer, National Harbor, MD, USA, 8–12 November 2017. [Google Scholar]
- Wardell, S.; Lienlaf-Moreno, M.; Blaskovich, M.; Frank, I.; Van Grod, V.; Fardis, M.; Wypych, J. Iovance gen 2 TIL manufacturing process produces drug products that exhibit favorable quality attributes for adoptive cell transfer across 5 solid tumor indications [poster]. In Proceedings of the Annual Meeting of the Society for Immunotherapy of Cancer, National Harbor, MD, USA, 6–10 November 2019. [Google Scholar]
- de Jong, R.A.; Leffers, N.; Boezen, H.M.; ten Hoor, K.A.; van der Zee, A.G.; Hollema, H.; Nijman, H.W. Presence of tumor-infiltrating lymphocytes is an independent prognostic factor in type I and II endometrial cancer. Gynecol. Oncol. 2009, 114, 105–110. [Google Scholar] [CrossRef]
- Son, J.; George, G.C.; Nardo, M.; Krause, K.J.; Jazaeri, A.A.; Biter, A.B.; Hong, D.S. Adoptive cell therapy in gynecologic cancers: A systematic review and meta-analysis. Gynecol. Oncol. 2022, 165, 664–670. [Google Scholar] [CrossRef]
- Cubas, R.; Yuhas, A.; Machin, M.; Zhang, Y. Decitabine treatment of tumor-infiltrating lymphocytes (TIL) during ex vivo expansion induces a more memory-like phenotype, reduces inhibitory receptor expression, and increases functionality [poster 270]. In Proceedings of the Annual Transplantation and Cellular Therapy Meetings of ASTCT and CIBMTR, Salt Lake City, UT, USA, 23–26 April 2022. [Google Scholar]
- Beltra, J.C.; Manne, S.; Abdel-Hakeem, M.S.; Kurachi, M.; Giles, J.R.; Chen, Z.; Casella, V.; Ngiow, S.F.; Khan, O.; Huang, Y.J.; et al. Developmental Relationships of Four Exhausted CD8(+) T Cell Subsets Reveals Underlying Transcriptional and Epigenetic Landscape Control Mechanisms. Immunity 2020, 52, 825–841.e828. [Google Scholar] [CrossRef]
- Chiffelle, J.; Barras, D.; Pétremand, R.; Orcurto, A.; Bobisse, S.; Arnaud, M.; Auger, A.; Rodrigo, B.N.; Ghisoni, E.; Sauvage, C.; et al. Tumor-reactive T cell clonotype dynamics underlying clinical response to TIL therapy in melanoma. Immunity 2024, 57, 2466–2482.e2412. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Liu, X.; Zhang, J.; Cao, Y.; Wei, B. Immune disorder in endometrial cancer: Immunosuppressive microenvironment, mechanisms of immune evasion and immunotherapy. Oncol. Lett. 2020, 20, 2075–2090. [Google Scholar] [CrossRef] [PubMed]
- Lowery, F.J.; Krishna, S.; Yossef, R.; Parikh, N.B.; Chatani, P.D.; Zacharakis, N.; Parkhurst, M.R.; Levin, N.; Sindiri, S.; Sachs, A.; et al. Molecular signatures of antitumor neoantigen-reactive T cells from metastatic human cancers. Science 2022, 375, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.; Donia, M.; Ellebaek, E.; Borch, T.H.; Kongsted, P.; Iversen, T.Z.; Hölmich, L.R.; Hendel, H.W.; Met, Ö.; Andersen, M.H.; et al. Long-Lasting Complete Responses in Patients with Metastatic Melanoma after Adoptive Cell Therapy with Tumor-Infiltrating Lymphocytes and an Attenuated IL2 Regimen. Clin. Cancer Res. 2016, 22, 3734–3745. [Google Scholar] [CrossRef]
- Kristensen, N.P.; Heeke, C.; Tvingsholm, S.A.; Borch, A.; Draghi, A.; Crowther, M.D.; Carri, I.; Munk, K.K.; Holm, J.S.; Bjerregaard, A.M.; et al. Neoantigen-reactive CD8+ T cells affect clinical outcome of adoptive cell therapy with tumor-infiltrating lymphocytes in melanoma. J. Clin. Investig. 2022, 132, e150535. [Google Scholar] [CrossRef]
- Ye, Q.; Song, D.G.; Poussin, M.; Yamamoto, T.; Best, A.; Li, C.; Coukos, G.; Powell, D.J., Jr. CD137 accurately identifies and enriches for naturally occurring tumor-reactive T cells in tumor. Clin. Cancer Res. 2014, 20, 44–55. [Google Scholar] [CrossRef]
- Vetto, J.T.; Lum, S.; Morris, A.; Sicotte, M.; Davis, J.; Lemon, M.; Weinberg, A. Presence of the T-cell activation marker OX-40 on tumor infiltrating lymphocytes and draining lymph node cells from patients with melanoma and head and neck cancers. Am. J. Surg. 1997, 174, 258–265. [Google Scholar] [CrossRef]
- Marín-Jiménez, J.A.; García-Mulero, S.; Matías-Guiu, X.; Piulats, J.M. Facts and Hopes in Immunotherapy of Endometrial Cancer. Clin. Cancer Res. 2022, 28, 4849–4860. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M., 3rd; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated analysis of multimodal single-cell data. Cell 2021, 184, 3573–3587.e3529. [Google Scholar] [CrossRef] [PubMed]
- Borcherding, N.; Bormann, N.L.; Kraus, G. scRepertoire: An R-based toolkit for single-cell immune receptor analysis. F1000Research 2020, 9, 47. [Google Scholar] [CrossRef]
- Hafemeister, C.; Satija, R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 2019, 20, 296. [Google Scholar] [CrossRef]
- Korsunsky, I.; Millard, N.; Fan, J.; Slowikowski, K.; Zhang, F.; Wei, K.; Baglaenko, Y.; Brenner, M.; Loh, P.R.; Raychaudhuri, S. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 2019, 16, 1289–1296. [Google Scholar] [CrossRef]
- Nofech-Mozes, I.; Soave, D.; Awadalla, P.; Abelson, S. Pan-cancer classification of single cells in the tumour microenvironment. Nat. Commun. 2023, 14, 1615. [Google Scholar] [CrossRef]
- Jones, M.; Detomaso, D.; Ashuach, T.; Rosen, Y. VISION: Functional Interpretation of Single Cell RNA-seq Latent Manifolds. Available online: https://yoseflab.github.io/VISION/authors.html#citation (accessed on 21 May 2025).
- Borcherding, N.; Andrews, J. Package ‘Escape’. Easy Single Cell Analysis Platform for Enrichment. Available online: https://git.bioconductor.org/packages/escape (accessed on 21 May 2025).
- Andreatta, M.; Carmona, S.J. UCell: Robust and scalable single-cell gene signature scoring. Comput. Struct. Biotechnol. J. 2021, 19, 3796–3798. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. Available online: https://raivokolde.r-universe.dev/pheatmap (accessed on 21 May 2025).
- Wickham, H.; Navarro, D.; Pedersen, T.L. ggplot2: Elegant Graphics for Data Analysis; Springer Nature: New York, NY, USA, 2016. [Google Scholar]
- Gerritsen, B.; Pandit, A.; Andeweg, A.C.; de Boer, R.J. RTCR: A pipeline for complete and accurate recovery of T cell repertoires from high throughput sequencing data. Bioinformatics 2016, 32, 3098–3106. [Google Scholar] [CrossRef]
- Popov, A.; Samokhina, M.; Balashov, I.; immunarch.bot; Rumynskiy, E.; Nazarov, V.I.; gracecodeadventures; tsvvas; Zarodniuk, M. Immunomind/Immunarch: Immunarch 0.9.1 (0.9.1). Available online: https://doi.org/10.5281/zenodo.10840553 (accessed on 21 May 2025).
| Tumor ID | Tumor Size (mg) | Total # of Fragments | TVC (Extrapolation) | Viability (NC200), % | CD45+CD3+% | CD4+CD3+% | CD8+CD3+% |
|---|---|---|---|---|---|---|---|
| END22061 | 1083 | 63 | 2.71 × 1010 | 90.1 | 96.6 | 75.0 | 23.0 |
| END22062 | 1100 | 48 | 1.24 × 1010 | 82.7 | 98.1 | 86.2 | 12.9 |
| END22063 | 1032 | 52 | N/A | N/A | N/A | N/A | N/A |
| END22091 | 1013 | 50 | 6.64 × 109 | 72.4 | 98.3 | 76.2 | 22.9 |
| END22093 | 1010 | 30 | 8.60 × 109 | 77.1 | 97.3 | 91.7 | 6.4 |
| END22094 | 1050 | 53 | 1.94 × 1010 | 87.9 | 90.2 | 67.0 | 31.7 |
| END22095 | 1131 | 48 | 8.10 × 109 | 82.8 | 97.8 | 81.6 | 17.1 |
| END22096 | 1400 | 59 | 9.77 × 109 | 84.9 | 93.0 | 78.8 | 19.9 |
| END22098 | 1400 | 80 | 3.53 × 1010 | 86.6 | 97.3 | 34.4 | 63.1 |
| END22100 | 1327 | 50 | 3.80 × 109 | 69.3 | 95.6 | 49.4 | 48.5 |
| END22102 | 1150 | 29 | 1.16 × 1010 | 72.3 | 94.6 | 51.3 | 46.5 |
| Total, median | 1.07 × 1010 | 82.8 | 97.0 | 75.6 | 23.0 |
| Final Product Attributes | Run 1 | Run 2 | Run 3 | Run 4 | Expected Results |
|---|---|---|---|---|---|
| END22111 | END22133 | END22134 | END22135 | ||
| Purity (cell viability, %) | 88.5 | 95.5 | 89.1 | 89.5 | ≥70% |
| Dose (TVC) | 5.9 × 109 | 77.1 × 109 | 2.7 × 109 | 68.3 × 109 | 1 × 109–150 × 109 cells |
| Identity (% CD45+/CD3+) | 96.2 | 99.4 | 96.1 | 99.2 | ≥90% |
| Cellular impurity (% TROP2+ or EPCAM+ tumor cells) | 0.06 | 0.04 | 0.06 | 0.08 | N/A |
| Potency (Dynabead-based IFN-γ release, pg/mL) | 9410 | 3940 | 12,620 | 6475 | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Moore, K.N.; Jazaeri, A.A.; Fang, J.; Patel, I.; Yuhas, A.; Innamarato, P.; Gilbert, N.; Dean, J.W.; Damirchi, B.; et al. Feasibility of Manufacturing and Antitumor Activity of TIL for Advanced Endometrial Cancers. Int. J. Mol. Sci. 2025, 26, 7151. https://doi.org/10.3390/ijms26157151
Zhang Y, Moore KN, Jazaeri AA, Fang J, Patel I, Yuhas A, Innamarato P, Gilbert N, Dean JW, Damirchi B, et al. Feasibility of Manufacturing and Antitumor Activity of TIL for Advanced Endometrial Cancers. International Journal of Molecular Sciences. 2025; 26(15):7151. https://doi.org/10.3390/ijms26157151
Chicago/Turabian StyleZhang, Yongliang, Kathleen N. Moore, Amir A. Jazaeri, Judy Fang, Ilabahen Patel, Andrew Yuhas, Patrick Innamarato, Nathan Gilbert, Joseph W. Dean, Behzad Damirchi, and et al. 2025. "Feasibility of Manufacturing and Antitumor Activity of TIL for Advanced Endometrial Cancers" International Journal of Molecular Sciences 26, no. 15: 7151. https://doi.org/10.3390/ijms26157151
APA StyleZhang, Y., Moore, K. N., Jazaeri, A. A., Fang, J., Patel, I., Yuhas, A., Innamarato, P., Gilbert, N., Dean, J. W., Damirchi, B., Yglesias, J., Qi, R., Simpson-Abelson, M. R., Cammaart, E., Hall, S. R. R., & Yin, H. (2025). Feasibility of Manufacturing and Antitumor Activity of TIL for Advanced Endometrial Cancers. International Journal of Molecular Sciences, 26(15), 7151. https://doi.org/10.3390/ijms26157151