
Hydroxyapatite Scaffold and Bioactive Factor Combination as a Tool to Improve Osteogenesis, In Vitro and In Vivo Experiments Using Phage Display Technology

 ,
,  , , , , ,
, , , , , 


Abstract
1. Introduction
2. Results
2.1. Peptide Alignment Characteristics
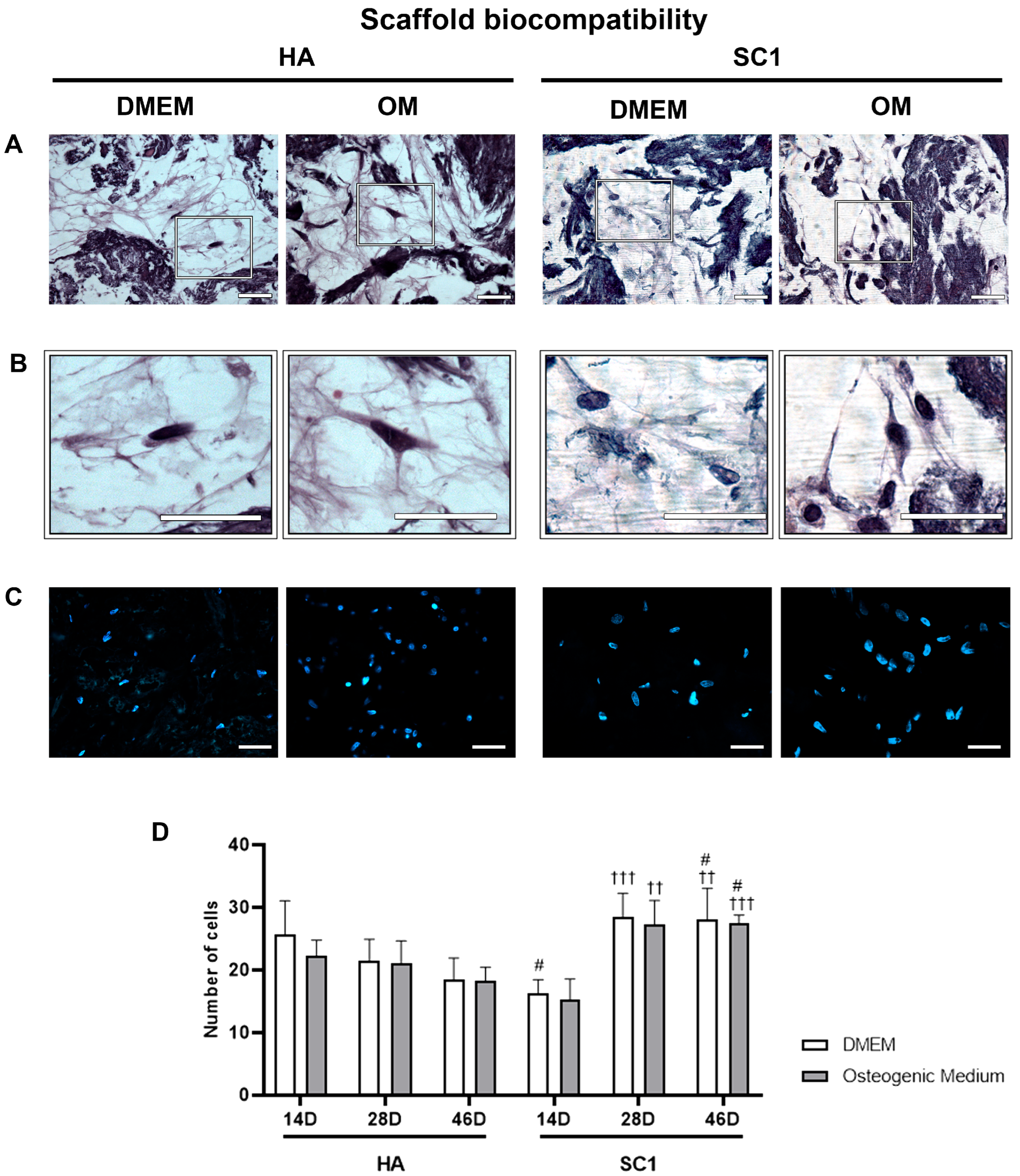
2.2. Scaffold Biocompatibility In Vitro
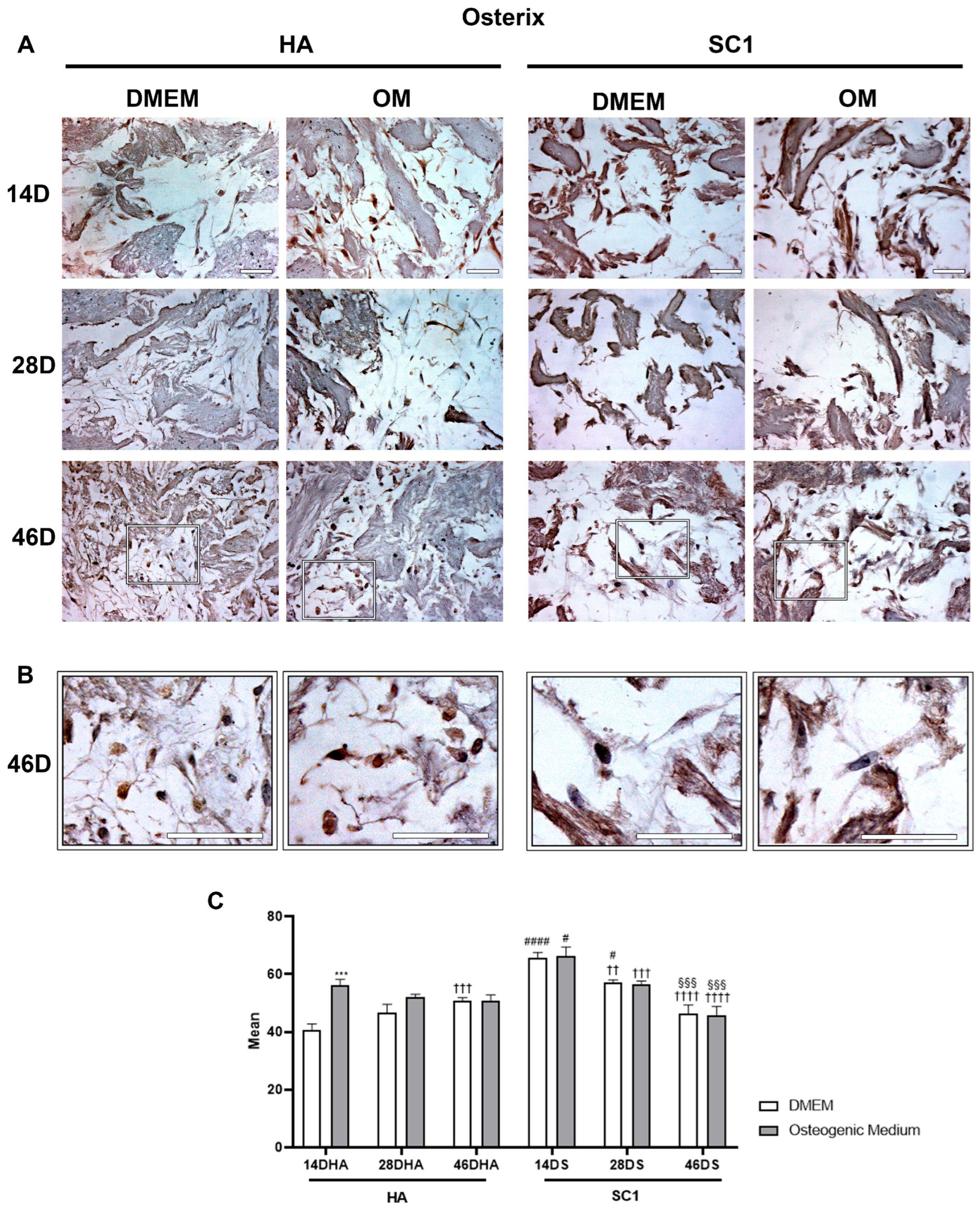
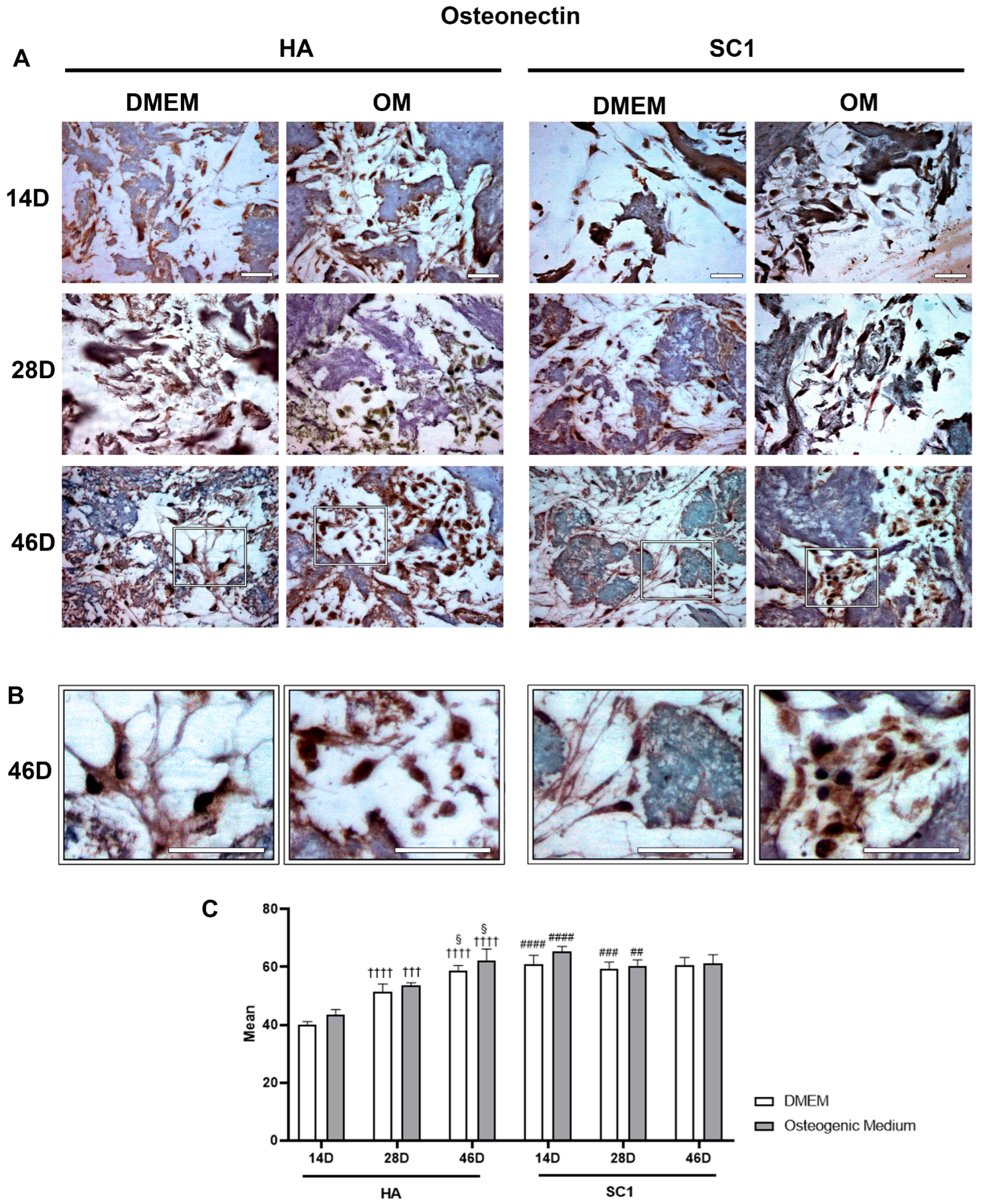
2.3. Scaffold Osteo-Inductive Effects on hASCs In Vitro
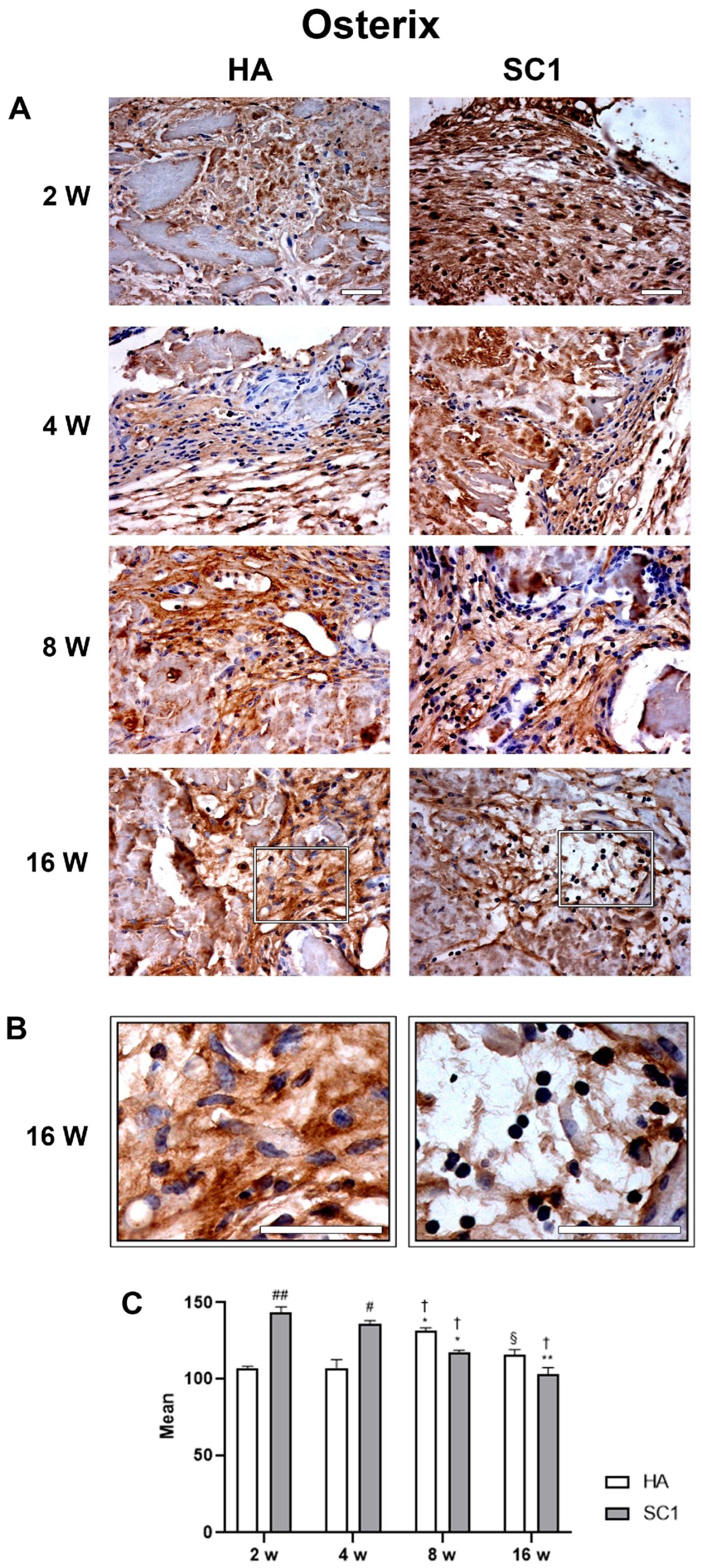
2.4. Osteogenic Differentiation In Vivo
3. Discussion
4. Materials and Methods
4.1. Scaffold Structure and Preparation
4.2. Phage Selection, Identification, and Propagation
4.3. hASC Cultures
4.4. In Vivo Experiments
4.5. Experimental Design
4.5.1. Scaffold-Supported hASC Osteogenic Differentiation In Vitro
4.5.2. Scaffold-Supported Osteogenic Differentiation In Vivo
4.6. Histological Procedures
4.7. Immunohistochemical Procedures
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BDDGE | 1,4-butanediol diglycidyl ether |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FBS | fetal bovine serum |
| HA | hydroxyapatite |
| hASC | human adipose-derived mesenchymal stem cell |
| MgHA | Mg-substituted HA |
| MSC | mesenchymal stem cell |
| MSCGS | MSC growth supplement |
| OM | osteogenic medium |
| PBS | phosphate-buffered saline |
| SC1 | scaffold functionalized with phage clones |
| TGFβ | transforming growth factor beta |
| TRIP-1 | TGFβ receptor-interacting protein 1 |
References
- Alonzo, M.; Primo, F.A.; Kumar, S.A.; Mudloff, J.A.; Dominguez, E.; Fregoso, G.; Ortiz, N.; Weiss, W.M.; Joddar, B. Bone tissue engineering techniques, advances and scaffolds for treatment of bone defects. Curr. Opin. Biomed. Eng. 2021, 17, 100248. [Google Scholar] [CrossRef] [PubMed]
- Xue, N.; Ding, X.; Huang, R.; Jiang, R.; Huang, H.; Pan, X.; Min, W.; Chen, J.; Duan, J.A.; Liu, P.; et al. Bone Tissue Engineering in the Treatment of Bone Defects. Pharmaceuticals 2022, 15, 879. [Google Scholar] [CrossRef] [PubMed]
- Banimohamad-Shotorbani, B.; Rahbarghazi, R.; Jarolmasjed, S.; Mehdipour, A.; Shafaei, H. Combination of mesenchymal stem cell sheet with poly-caprolactone nanofibrous mat and Gelfoam increased osteogenesis capacity in rat calvarial defect. Bioimpacts 2025, 15, 30006. [Google Scholar] [CrossRef] [PubMed]
- Ci, H.; Jie, J.; Zhang, G.; Wu, L.; Wang, Z.; Sun, J. Injectable Polyhydroxyalkanoate-Nano-Clay Microcarriers Loaded with r-BMSCs Enhance the Repair of Cranial Defects in Rats. Int. J. Nanomed. 2024, 19, 13839–13855. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Giuffrida, R.; Fabbi, C.; Figallo, E.; Lo Furno, D.; Gulino, R.; Colarossi, C.; Fullone, F.; Parenti, R.; Memeo, L.; et al. Collagen-Hydroxyapatite Scaffolds Induce Human Adipose Derived Stem Cells Osteogenic Differentiation In Vitro. PLoS ONE 2016, 11, e0151181. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yu, X.; Hao, M.; Wang, Y.; Fan, G.; Qian, H.; Jiang, P.; Cai, Z.; He, Z.; Zhou, G. Biomimetic Fabrication and Osteogenic Effects of E7BMP-2 Peptide Coassembly Microspheres Based α-Tricalcium Phosphate with Silk Fibroin. ACS Appl. Bio Mater. 2025, 8, 2078–2089. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Russo, C.; Maugeri, G.; Musumeci, G.; Vicario, N.; Tibullo, D.; Giuffrida, R.; Parenti, R.; Lo Furno, D. Adult stem cell niches for tissue homeostasis. J. Cell Physiol. 2022, 237, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Lo Furno, D.; Mannino, G.; Pellitteri, R.; Zappalà, A.; Parenti, R.; Gili, E.; Vancheri, C.; Giuffrida, R. Conditioned Media From Glial Cells Promote a Neural-Like Connexin Expression in Human Adipose-Derived Mesenchymal Stem Cells. Front. Physiol. 2018, 9, 1742. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Petrakova, K.V.; Kurolesova, A.I.; Frolova, G.P. Heterotopic of bone marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation 1968, 6, 230–247. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; McKee, C.; Bakshi, S.; Walker, K.; Hakman, E.; Halassy, S.; Svinarich, D.; Dodds, R.; Govind, C.K.; Chaudhry, G.R. Mesenchymal stem cells: Cell therapy and regeneration potential. J. Tissue Eng. Regen. Med. 2019, 13, 1738–1755. [Google Scholar] [CrossRef] [PubMed]
- Kokai, L.E.; Marra, K.; Rubin, J.P. Adipose stem cells: Biology and clinical applications for tissue repair and regeneration. Transl. Res. 2014, 163, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Shingyochi, Y.; Orbay, H.; Mizuno, H. Adipose-derived stem cells for wound repair and regeneration. Expert. Opin. Biol. Ther. 2015, 15, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Lo Furno, D.; Mannino, G.; Cardile, V.; Parenti, R.; Giuffrida, R. Potential Therapeutic Applications of Adipose-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2016, 25, 1615–1628. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Zhang, Y.; Zhao, R.; Lin, H.; Yang, X.; Zhu, X.; Zhang, K.; Mikos, A.G.; Zhang, X. A unique biomimetic modification endows polyetherketoneketone scaffold with osteoinductivity by activating cAMP/PKA signaling pathway. Sci. Adv. 2022, 8, eabq7116. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.R.; D’Angeli, F.; Vicario, N.; Russo, C.; Genovese, C.; Lo Furno, D.; Mannino, G.; Tamburino, S.; Parenti, R.; Giuffrida, R. Adipose-Derived Mesenchymal Stromal Cells: A Tool for Bone and Cartilage Repair. Biomedicines 2023, 11, 1781. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, P.; Alves, B.; Inocêncio, S.; Nunes, P.; Richardson, S.M.; Gloria, A.; Serra, A.; Fonseca, A.C.; Domingos, M. Synthesis and characterization of poly(ester amide)-based materials for 3D printing of tissue engineering scaffolds. J. Mater. Chem. B 2025, 13, 3049–3066. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, F.; Tanaka, Y.; Minari, Y.; Tokui, N. Designing scaffolds of peptides for phage display libraries. J. Biosci. Bioeng. 2005, 99, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Zambrano-Mila, M.S.; Blacio, K.E.S.; Vispo, N.S. Peptide Phage Display: Molecular Principles and Biomedical Applications. Ther. Innov. Regul. Sci. 2020, 54, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Metz-Estrella, D.; Jonason, J.H.; Sheu, T.J.; Mroczek-Johnston, R.M.; Puzas, J.E. TRIP-1: A regulator of osteoblast function. J. Bone Miner. Res. 2012, 27, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhong, W.; Cao, W.; Zhang, Q.; Wu, G. Chondroinductive/chondroconductive peptides and their-functionalized biomaterials for cartilage tissue engineering. Bioact. Mater. 2022, 9, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.G.; Palermo, N.; D’Amora, U.; Oddo, S.; Guglielmino, S.P.P.; Conoci, S.; Szychlinska, M.A.; Calabrese, G. Multipotential Role of Growth Factor Mimetic Peptides for Osteochondral Tissue Engineering. Int. J. Mol. Sci. 2022, 23, 7388. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.A.; Gunn, W.G.; Peister, A.; Prockop, D.J. An Alizarin red-based assay of mineralization by adherent cells in culture: Comparison with cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.D.; Brett, E.A.; Moore, A.L.; Wan, D.C.; Longaker, M.T. In Vitro and In Vivo Osteogenic Differentiation of Human Adipose-Derived Stromal Cells. Methods Mol. Biol. 2019, 1891, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Cho, D.H.; Chung, D.J.; Lee, K.Y. Osterix plays a critical role in BMP4-induced promoter activity of connexin43. Biochem. Biophys. Res. Commun. 2016, 478, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Tampieri, A.; Sprio, S.; Sandri, M.; Valentini, F. Mimicking natural bio-mineralization processes: A new tool for osteochondral scaffold development. Trends Biotechnol. 2011, 29, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Erisken, C.; Kalyon, D.M.; Wang, H.; Ornek-Ballanco, C.; Xu, J. Osteochondral tissue formation through adipose-derived stromal cell differentiation on biomimetic polycaprolactone nanofibrous scaffolds with graded insulin and Beta-glycerophosphate concentrations. Tissue Eng. Part A 2011, 17, 1239–1252. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Li, J.; Li, Y.; Khadka, A.; Zuo, Y.; Wang, H.; Liu, Y.; Cheng, L. Ectopic osteochondral formation of biomimetic porous PVA-n-HA/PA6 bilayered scaffold and BMSCs construct in rabbit. J. Biomed. Mater. Res. B Appl. Biomater. 2011, 96, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Alidadi, S.; Moshiri, A.; Maffulli, N. Bone regenerative medicine: Classic options, novel strategies, and future directions. J. Orthop. Surg. Res. 2014, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Forte, S.; Gulino, R.; Cefalì, F.; Figallo, E.; Salvatorelli, L.; Maniscalchi, E.T.; Angelico, G.; Parenti, R.; Gulisano, M.; et al. Combination of Collagen-Based Scaffold and Bioactive Factors Induces Adipose-Derived Mesenchymal Stem Cells Chondrogenic Differentiation. Front. Physiol. 2017, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Giudici, C.; Raynal, N.; Wiedemann, H.; Cabral, W.A.; Marini, J.C.; Timpl, R.; Bächinger, H.P.; Farndale, R.W.; Sasaki, T.; Tenni, R. Mapping of SPARC/BM-40/osteonectin-binding sites on fibrillar collagens. J. Biol. Chem. 2008, 283, 19551–19560. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, M.; Wang, S.; Xiao, Z.; Xiong, Y.; Wang, G. Recent Advances of Osterix Transcription Factor in Osteoblast Differentiation and Bone Formation. Front. Cell Dev. Biol. 2020, 8, 601224. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Friedman, M.S.; Luo, W.; Woolf, P.; Hankenson, K.D. The transcription factor osterix (SP7) regulates BMP6-induced human osteoblast differentiation. J. Cell Physiol. 2012, 227, 2677–2685. [Google Scholar] [CrossRef] [PubMed]
- Rosset, E.M.; Bradshaw, A.D. SPARC/osteonectin in mineralized tissue. Matrix Biol. 2016, 52–54, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Mo, T.T.; Jiang, C.; Zhang, J.N. Osteonectin bidirectionally regulates osteoblast mineralization. J. Orthop. Surg. Res. 2023, 18, 761. [Google Scholar] [CrossRef] [PubMed]
- Delany, A.M.; Kalajzic, I.; Bradshaw, A.D.; Sage, E.H.; Canalis, E. Osteonectin-null mutation compromises osteoblast formation, maturation, and survival. Endocrinology 2003, 144, 2588–2596. [Google Scholar] [CrossRef] [PubMed]
- Delany, A.M.; Hankenson, K.D. Thrombospondin-2 and SPARC/osteonectin are critical regulators of bone remodeling. J. Cell Commun. Signal 2009, 3, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Gu, Y.; Jiang, C.; Chen, L. Osteonectin regulates the extracellular matrix mineralization of osteoblasts through P38 signaling pathway. J. Cell Physiol. 2020, 235, 2220–2231. [Google Scholar] [CrossRef] [PubMed]
- Motamed, K. SPARC (osteonectin/BM-40). Int. J. Biochem. Cell Biol. 1999, 31, 1363–1366. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, P.T.; Jansen, M.S.; Ganesan, S.; Benson, R.E.; Hyde-Deruyscher, R.; Beyer, W.F.; Gile, J.C.; Nair, S.A.; Hodges, J.A.; Grøn, H. Improved bone morphogenetic protein-2 retention in an injectable collagen matrix using bifunctional peptides. PLoS ONE 2013, 8, e70715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Tang, Y.; Chen, Q.; Liu, Y. The Screening of Therapeutic Peptides for Anti-Inflammation through Phage Display Technology. Int. J. Mol. Sci. 2022, 23, 8554. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.G.; De Plano, L.M.; Palermo, N.; Franco, D.; Nicolò, M.; Sciuto, E.L.; Calabrese, G.; Oddo, S.; Conoci, S.; Guglielmino, S.P.P. A Novel Serum-Based Diagnosis of Alzheimer’s Disease Using an Advanced Phage-Based Biochip. Adv. Sci. (Weinh.) 2023, 10, e2301650. [Google Scholar] [CrossRef] [PubMed]
- Felici, F.; Luzzago, A.; Folgori, A.; Cortese, R. Mimicking of discontinuous epitopes by phage-displayed peptides, II. Selection of clones recognized by a protective monoclonal antibody against the Bordetella pertussis toxin from phage peptide libraries. Gene 1993, 128, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Lentini, G.; Fazio, E.; Calabrese, F.; De Plano, L.M.; Puliafico, M.; Franco, D.; Nicolò, M.S.; Carnazza, S.; Trusso, S.; Allegra, A.; et al. Phage-AgNPs complex as SERS probe for U937 cell identification. Biosens. Bioelectron. 2015, 74, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Aiyar, A. The use of CLUSTAL W and CLUSTAL X for multiple sequence alignment. Methods Mol. Biol. 2000, 132, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.R.; D’Angeli, F.; Gili, E.; Fruciano, M.; Lombardo, G.A.G.; Mannino, G.; Vicario, N.; Russo, C.; Parenti, R.; Vancheri, C.; et al. Melatonin Enhances Neural Differentiation of Adipose-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2024, 25, 4891. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Gulino, R.; Giuffrida, R.; Forte, S.; Figallo, E.; Fabbi, C.; Salvatorelli, L.; Memeo, L.; Gulisano, M.; Parenti, R. Evaluation of Biocompatibility and Chondrogenic Potential of a Cell-Free Collagen-Based Scaffold. Front. Physiol. 2017, 8, 984. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Groups | Time Points | Number of Mice |
|---|---|---|
| HA | 2, 4, 8, 16 weeks | n. 28 (7 × each time point) |
| SC1 | 2, 4, 8, 16 weeks | n. 28 (7 × each time point) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Furno, D.; Romano, I.R.; Russo, V.; Rizzo, M.G.; Mannino, G.; Calabrese, G.; Giuffrida, R.; D’Aprile, S.; Salvatorelli, L.; Magro, G.; et al. Hydroxyapatite Scaffold and Bioactive Factor Combination as a Tool to Improve Osteogenesis, In Vitro and In Vivo Experiments Using Phage Display Technology. Int. J. Mol. Sci. 2025, 26, 7040. https://doi.org/10.3390/ijms26157040
Lo Furno D, Romano IR, Russo V, Rizzo MG, Mannino G, Calabrese G, Giuffrida R, D’Aprile S, Salvatorelli L, Magro G, et al. Hydroxyapatite Scaffold and Bioactive Factor Combination as a Tool to Improve Osteogenesis, In Vitro and In Vivo Experiments Using Phage Display Technology. International Journal of Molecular Sciences. 2025; 26(15):7040. https://doi.org/10.3390/ijms26157040
Chicago/Turabian StyleLo Furno, Debora, Ivana R. Romano, Vincenzo Russo, Maria Giovanna Rizzo, Giuliana Mannino, Giovanna Calabrese, Rosario Giuffrida, Simona D’Aprile, Lucia Salvatorelli, Gaetano Magro, and et al. 2025. "Hydroxyapatite Scaffold and Bioactive Factor Combination as a Tool to Improve Osteogenesis, In Vitro and In Vivo Experiments Using Phage Display Technology" International Journal of Molecular Sciences 26, no. 15: 7040. https://doi.org/10.3390/ijms26157040
APA StyleLo Furno, D., Romano, I. R., Russo, V., Rizzo, M. G., Mannino, G., Calabrese, G., Giuffrida, R., D’Aprile, S., Salvatorelli, L., Magro, G., Bendoni, R., Dolcini, L., Zappalà, A., Guglielmino, S. P. P., Conoci, S., & Parenti, R. (2025). Hydroxyapatite Scaffold and Bioactive Factor Combination as a Tool to Improve Osteogenesis, In Vitro and In Vivo Experiments Using Phage Display Technology. International Journal of Molecular Sciences, 26(15), 7040. https://doi.org/10.3390/ijms26157040