
Integrated Metabolomic and Transcriptomic Analysis Reveals the Regulatory Effects of Curcumin on Bovine Ovarian Granulosa Cells
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
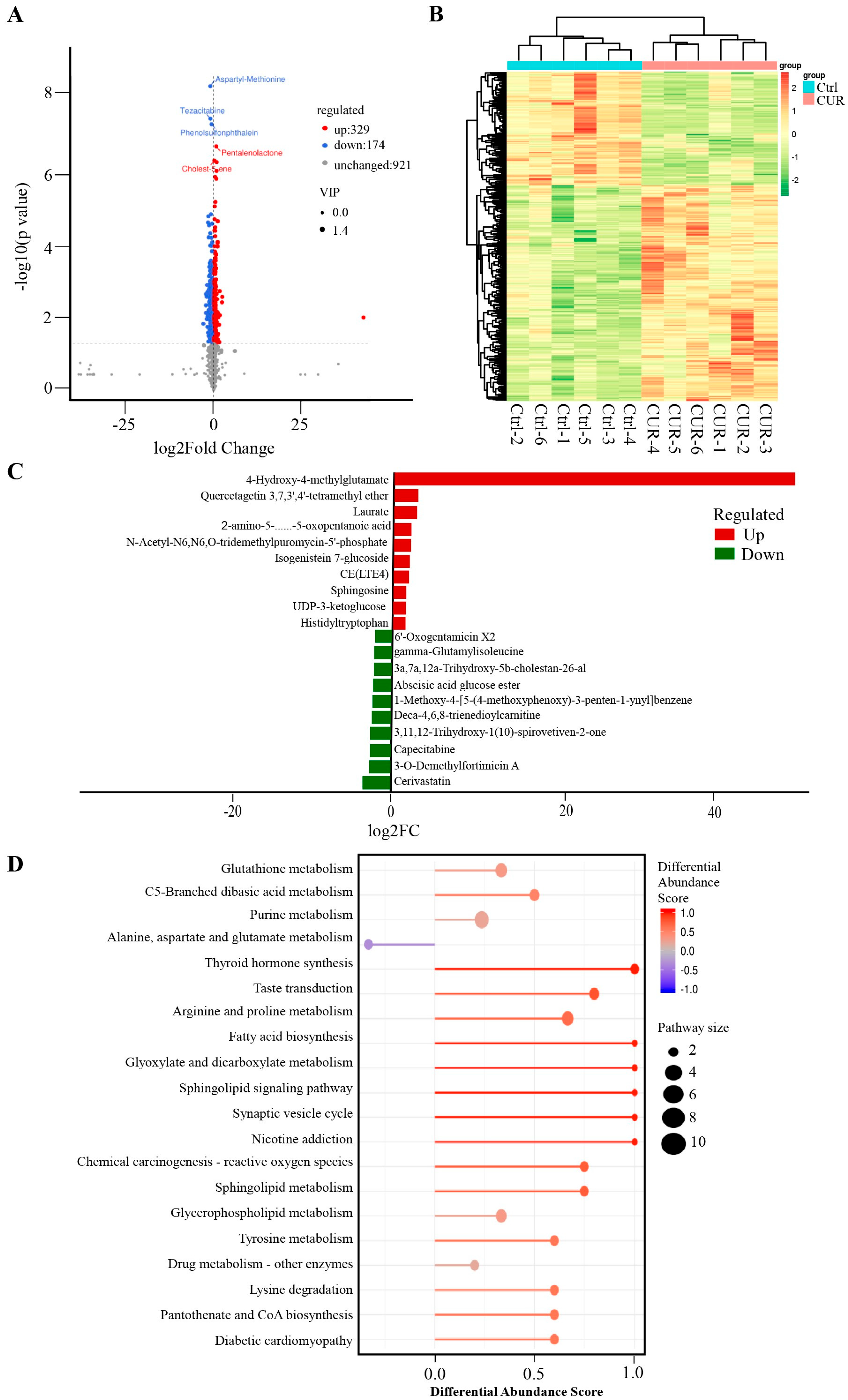
2.1. Effect of Curcumin Treatment on the Metabolomics of GCs in Positive Mode
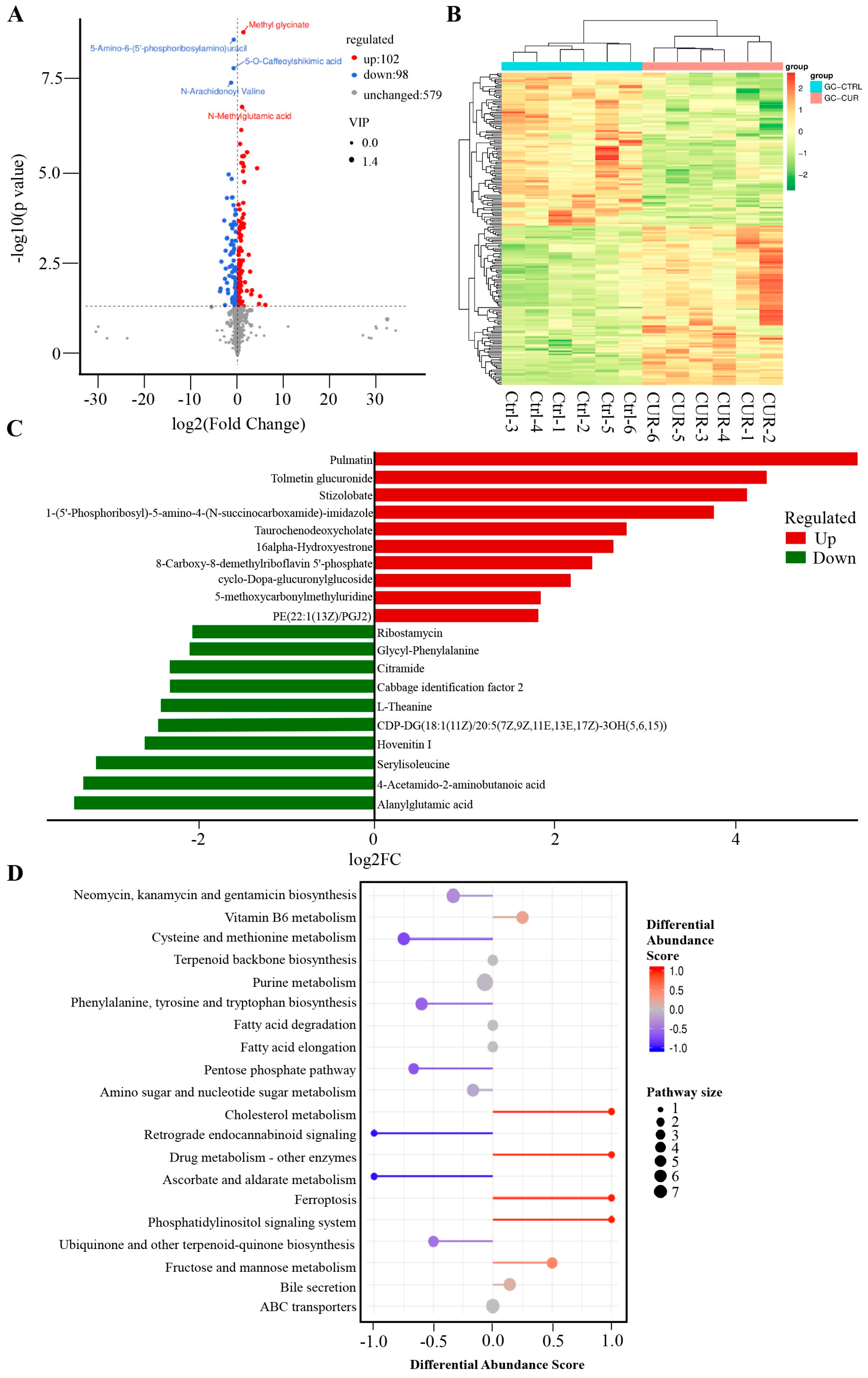
2.2. Effect of Curcumin Treatment on the Metabolomics of GCs in Negative Mode
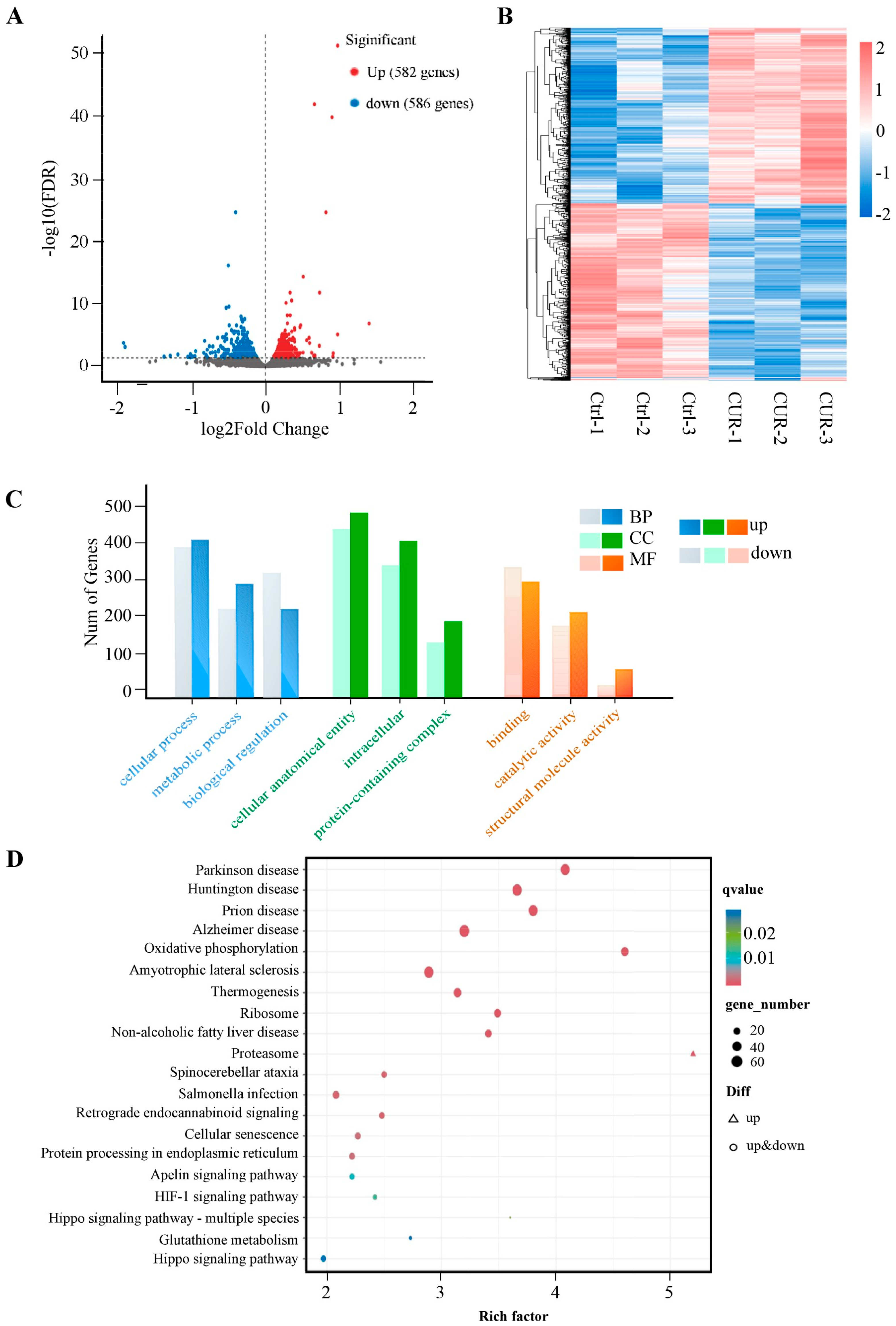
2.3. Transcriptomic Analysis of Curcumin-Treated GCs
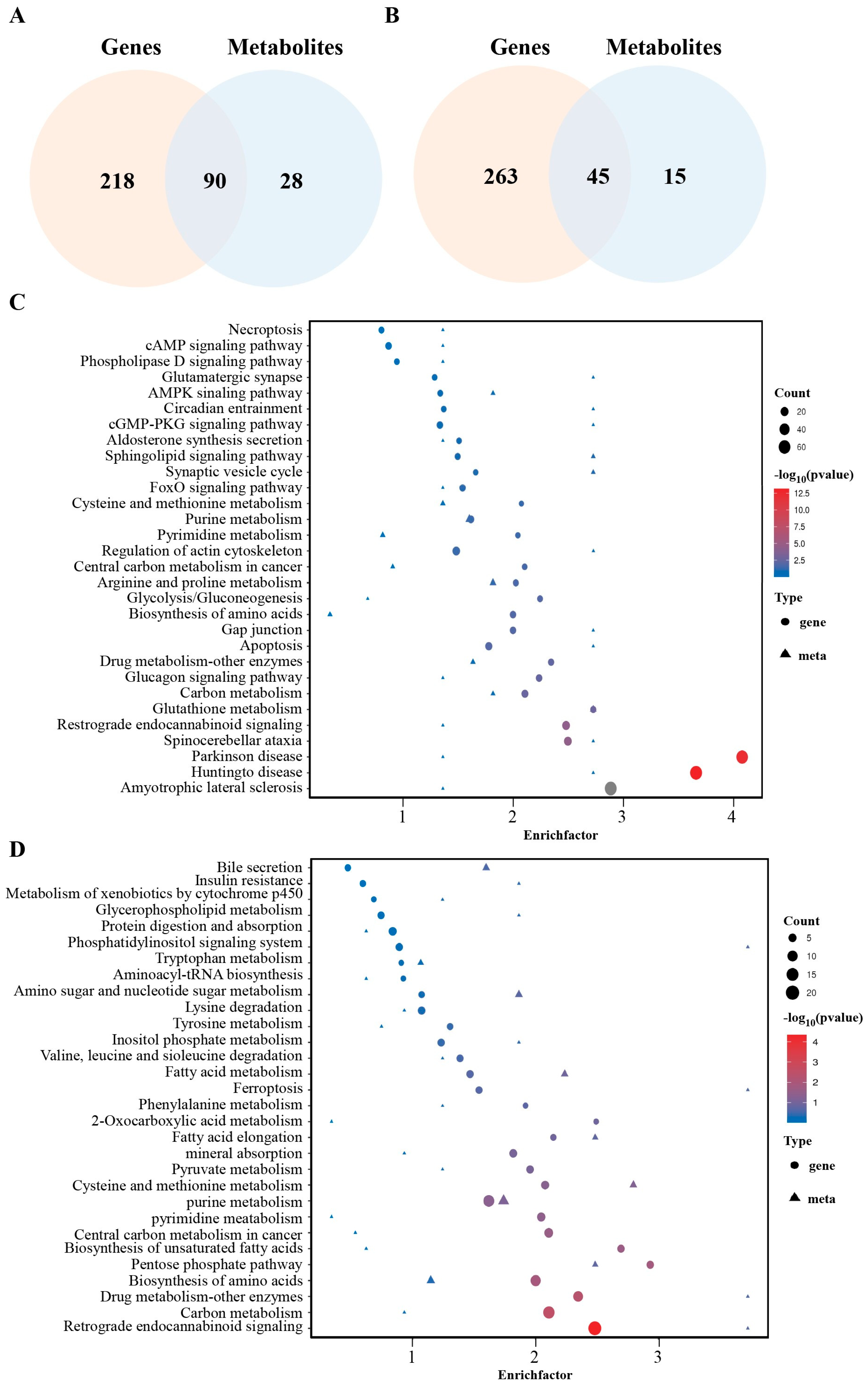
2.4. Integrated Transcriptomic and Metabolomic Analysis of Curcumin-Treated GCs
3. Discussion
4. Materials and Methods
4.1. Bovine Granulosa Cell Culture
4.2. Library Preparation and Transcriptome Analysis
4.3. Metabolite Extraction
4.4. LC-MS/MS Analysis
4.5. Data Processing and Annotation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richards, J.S.; Pangas, S.A. The ovary: Basic biology and clinical implications. J. Clin. Investig. 2010, 120, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Da Broi, M.G.; Giorgi, V.S.I.; Wang, F.; Keefe, D.L.; Albertini, D.; Navarro, P.A. Influence of follicular fluid and cumulus cells on oocyte quality: Clinical implications. J. Assist. Reprod. Genet. 2018, 35, 735–751. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, G.S.; Carvalho, K.C.; Ferreira, C.D.S.; Alvarez, P.A.C.; Monteleone, P.A.A.; Baracat, E.C.; Soares Júnior, J.M. Granulosa cells and follicular development: A brief review. Rev. Assoc. Med. Bras. 2023, 69, e20230175. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.G.; Wu, H.C.; Lian, F.; Zhang, X.X.; Pang, C.H.; Guo, Y.; Song, J.Y.; Wang, A.J.; Shi, L.; Han, L.T. Human Follicular Fluid Metabolomics Study of Follicular Development and Oocyte Quality. Chromatographia 2017, 80, 901–909. [Google Scholar] [CrossRef]
- Ji, R.; Jia, F.; Chen, X.; Gao, Y.; Yang, J. Carnosol inhibits KGN cells oxidative stress and apoptosis and attenuates polycystic ovary syndrome phenotypes in mice through Keap1-mediated Nrf2/HO-1 activation. Phytother. Res. 2023, 37, 1405–1421. [Google Scholar] [CrossRef]
- Khan, M.Z.; Khan, A.; Chen, W.; Chai, W.; Wang, C. Advancements in Genetic Biomarkers and Exogenous Antioxidant Supplementation for Safeguarding Mammalian Cells against Heat-Induced Oxidative Stress and Apoptosis. Antioxidants 2024, 13, 258. [Google Scholar] [CrossRef]
- Alam, M.H.; Miyano, T. Interaction between growing oocytes and granulosa cells in vitro. Reprod. Med. Biol. 2020, 19, 13–23. [Google Scholar] [CrossRef]
- Regan, S.L.P.; Knight, P.G.; Yovich, J.L.; Leung, Y.; Arfuso, F.; Dharmarajan, A. Granulosa Cell Apoptosis in the Ovarian Follicle-A Changing View. Front. Endocrinol. 2018, 9, 61. [Google Scholar] [CrossRef]
- Hao, X.; Anastácio, A.; Liu, K.; Rodriguez-Wallberg, K.A. Ovarian Follicle Depletion Induced by Chemotherapy and the Investigational Stages of Potential Fertility-Protective Treatments—A Review. Int. J. Mol. Sci. 2019, 20, 4720. [Google Scholar] [CrossRef]
- Fu, Y.S.; Chen, T.H.; Weng, L.; Huang, L.; Lai, D.; Weng, C.F. Pharmacological properties and underlying mechanisms of curcumin and prospects in medicinal potential. Biomed. Pharmacother. 2021, 141, 111888. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, H.; Bu, L.; Chen, C.; Ye, X. Review of the Effects and Mechanism of Curcumin in the Treatment of Inflammatory Bowel Disease. Front. Pharmacol. 2022, 13, 908077. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, X.; Pi, C.; Yang, H.; Zheng, X.; Zhao, L.; Wei, Y. Review of Curcumin Physicochemical Targeting Delivery System. Int. J. Nanomed. 2020, 15, 9799–9821. [Google Scholar] [CrossRef] [PubMed]
- Abadi, A.J.; Mirzaei, S.; Mahabady, M.K.; Hashemi, F.; Zabolian, A.; Hashemi, F.; Raee, P.; Aghamiri, S.; Ashrafizadeh, M.; Aref, A.R.; et al. Curcumin and its derivatives in cancer therapy: Potentiating antitumor activity of cisplatin and reducing side effects. Phytother. Res. 2022, 36, 189–213. [Google Scholar] [CrossRef] [PubMed]
- Aditi, K.; Singh, A.; Shakarad, M.N.; Agrawal, N. Management of altered metabolic activity in Drosophila model of Huntington’s disease by curcumin. Exp. Biol. Med. 2022, 247, 152–164. [Google Scholar] [CrossRef]
- Voznesens’ka, T.I.; Bryzhina, T.M.; Sukhina, V.S.; Makohon, N.V.; Aleksieieva, I.M. Effect of NF-kappaB activation inhibitor curcumin on the oogenesis and follicular cell death in immune ovarian failure in mice. Fiziolohichnyi Zhurnal. 2010, 56, 96–101. [Google Scholar] [CrossRef]
- Caesar, J.; Widjiati, W.; Herupradoto, E.B.A.; Sukmanadi, M.; Madyawati, S.P.; Plumeriastuti, H.; Luqman, E.M. Effect of curcumin nanoparticles on the number of preantral and antral follicles of white rats (Rattus norvegicus) exposed to carbon black. Open Vet. J. 2024, 14, 3309–3316. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Liu, D.; Feng, H.; Wang, X.; Su, J.; Yao, Y.; Ng, E.H.Y.; Yeung, W.S.B.; Li, R.H.W.; et al. Identification of curcumin as a novel potential drug for promoting the development of small ovarian follicles for infertility treatment. PNAS Nexus 2022, 1, 108. [Google Scholar] [CrossRef]
- Nakanishi, T.; Tonai, S.; Ichikawa, H.; Mori, S.; Ishihara, S.; Chang, Y.J.; Yamashita, Y. Curcumin Suppresses ROS Production and Increases Mitochondrial Activity in Cumulus Cells and Oocytes of COCs Derived from Non-Vascularized Follicles in Pigs. Anim. Sci. J. 2025, 96, e70032. [Google Scholar] [CrossRef]
- Li, W.J.; Xu, J.B.; Wang, J.Y.; Zhang, J.X.; Deng, D.S. Regulation of Curcumin on Follicle Initiation Rate in Diminished Ovarian Reserve. Comb. Chem. High Throughput Screen. 2024; online ahead of print. [Google Scholar] [CrossRef]
- Li, W.; Xu, J.; Shi, D.; Wang, J.; Liu, T.; Yang, J.; Deng, D. Curcumin Regulating Primordial Follicle Initiation by Restoring the Oxidative-antioxidant Balance. Comb. Chem. High Throughput Screen. 2025. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, P.F.; Dai, W.C.; Zheng, Z.Q.; Wang, H.L. Curcumin Alleviates Hyperandrogenism and Promotes Follicular Proliferation in Polycystic Ovary Syndrome Rats: Insights on IRS1/PI3K/GLUT4 and PTEN Modulations. Chin. J. Integr. Med. 2022, 28, 1088–1095. [Google Scholar] [CrossRef]
- Yan, Z.; Dai, Y.; Fu, H.; Zheng, Y.; Bao, D.; Yin, Y.; Chen, Q.; Nie, X.; Hao, Q.; Hou, D.; et al. Curcumin exerts a protective effect against premature ovarian failure in mice. J. Mol. Endocrinol. 2018, 60, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Wang, H.; Li, S.; Wei, Z.; Han, S.; Chen, B. Effects of dietary supplementation with quercetagetin on nutrient digestibility, intestinal morphology, immunity, and antioxidant capacity of broilers. Front. Vet. Sci. 2022, 9, 1060140. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Su, Z.; Huo, M.; Zhong, C.; Wang, F.; Zhang, Y.; Song, Y.; Shi, Y. Effect of Supplementation of Quercetagetin on the Antioxidant Function, Liver Mitochondrial Function and Gut Microbiota of Broilers at High Stocking Density. Animals 2025, 15, 398. [Google Scholar] [CrossRef] [PubMed]
- Dubo, A.; Dawud, F.; Umar, I.; Alex, E.; Baiyekusi, S.; Farra’u, U. Lauric acid alleviates inflammation and structural changes in the lungs of type II diabetic male Wistar rats. J. Afr. Assoc. Physiol. Sci. 2019, 7, 88–96. [Google Scholar]
- Wang, L.; Huang, Q.; Gu, Z.; Liu, C.; Ren, X.; Dai, Y.; Ye, T.; Yan, J.; Meng, J.; Zheng, Y.; et al. The effects of dietary curcumin on growth, antioxidant, transcriptome, and metabolomics in Macrobrachium rosenbergii fed a high plant-protein diet. Aquac. Rep. 2025, 42, 102782. [Google Scholar] [CrossRef]
- Chen, D.; Yu, Q.; Sheng, S.; Cai, L.; Zheng, J.; Zhang, Y. Transcriptomic analysis of the effects exerted by curcumin on dihydrotestosterone-induced ovarian granulosa cells. Front. Endocrinol. 2025, 16, 1522269. [Google Scholar] [CrossRef]
- Grosbois, J.; Demeestere, I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum. Reprod. 2018, 33, 1705–1714. [Google Scholar] [CrossRef]
- Hsueh, A.J.W.; Kawamura, K. Hippo signaling disruption and ovarian follicle activation in infertile patients. Fertil. Steril. 2020, 114, 458–464. [Google Scholar] [CrossRef]
- Duan, H.; Yang, S.; Yang, S.; Zeng, J.; Yan, Z.; Zhang, L.; Ma, X.; Dong, W.; Zhang, Y.; Zhao, X. The mechanism of curcumin to protect mouse ovaries from oxidative damage by regulating AMPK/mTOR mediated autophagy. Phytomedicine 2024, 128, 155468. [Google Scholar] [CrossRef]
- Shen, M.; Liu, Z.; Li, B.; Teng, Y.; Zhang, J.; Tang, Y.; Sun, S.C.; Liu, H. Involvement of FoxO1 in the effects of follicle-stimulating hormone on inhibition of apoptosis in mouse granulosa cells. Cell Death Dis. 2014, 5, e1475. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Gan, M.; Wu, S.; He, Y.; Feng, J.; Jing, Y.; Li, J.; Chen, Q.; Tong, J.; Kang, L.; et al. tRF-Gly-GCC in Atretic Follicles Promotes Ferroptosis in Granulosa Cells by Down-Regulating MAPK1. Int. J. Mol. Sci. 2024, 25, 9061. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ni, F.; Huang, J.; Hu, Y.; Wang, J.; Wang, X.; Du, X.; Jiang, H. PPAR-α inhibits DHEA-induced ferroptosis in granulosa cells through upregulation of FADS2. Biochem. Biophys. Res. Commun. 2024, 715, 150005. [Google Scholar] [CrossRef] [PubMed]
- Abudureyimu, G.; Wu, Y.; Chen, Y.; Wang, L.; Hao, G.; Yu, J.; Wang, J.; Lin, J.; Huang, J. MiR-134-3p targets HMOX1 to inhibit ferroptosis in granulosa cells of sheep follicles. J. Ovarian Res. 2024, 17, 3. [Google Scholar] [CrossRef]
- Manabe, N.; Goto, Y.; Matsuda-Minehata, F.; Inoue, N.; Maeda, A.; Sakamaki, K.; Miyano, T. Regulation mechanism of selective atresia in porcine follicles: Regulation of granulosa cell apoptosis during atresia. J. Reprod. Dev. 2004, 50, 493–514. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Chen, L.; Mei, L.; Jia, X.; Chen, S.; Wang, J.; Yu, H.; Lai, S.; Sun, W. Integrated Metabolomic and Transcriptomic Analysis Reveals the Regulatory Effects of Curcumin on Bovine Ovarian Granulosa Cells. Int. J. Mol. Sci. 2025, 26, 6713. https://doi.org/10.3390/ijms26146713
Zhang B, Chen L, Mei L, Jia X, Chen S, Wang J, Yu H, Lai S, Sun W. Integrated Metabolomic and Transcriptomic Analysis Reveals the Regulatory Effects of Curcumin on Bovine Ovarian Granulosa Cells. International Journal of Molecular Sciences. 2025; 26(14):6713. https://doi.org/10.3390/ijms26146713
Chicago/Turabian StyleZhang, Bingfei, Le Chen, Liping Mei, Xianbo Jia, Shiyi Chen, Jie Wang, Hengwei Yu, Songjia Lai, and Wenqiang Sun. 2025. "Integrated Metabolomic and Transcriptomic Analysis Reveals the Regulatory Effects of Curcumin on Bovine Ovarian Granulosa Cells" International Journal of Molecular Sciences 26, no. 14: 6713. https://doi.org/10.3390/ijms26146713
APA StyleZhang, B., Chen, L., Mei, L., Jia, X., Chen, S., Wang, J., Yu, H., Lai, S., & Sun, W. (2025). Integrated Metabolomic and Transcriptomic Analysis Reveals the Regulatory Effects of Curcumin on Bovine Ovarian Granulosa Cells. International Journal of Molecular Sciences, 26(14), 6713. https://doi.org/10.3390/ijms26146713