
Identification of Genes Linked to Meniscal Degeneration in Osteoarthritis: An In Silico Analysis


Abstract
1. Introduction
2. Results
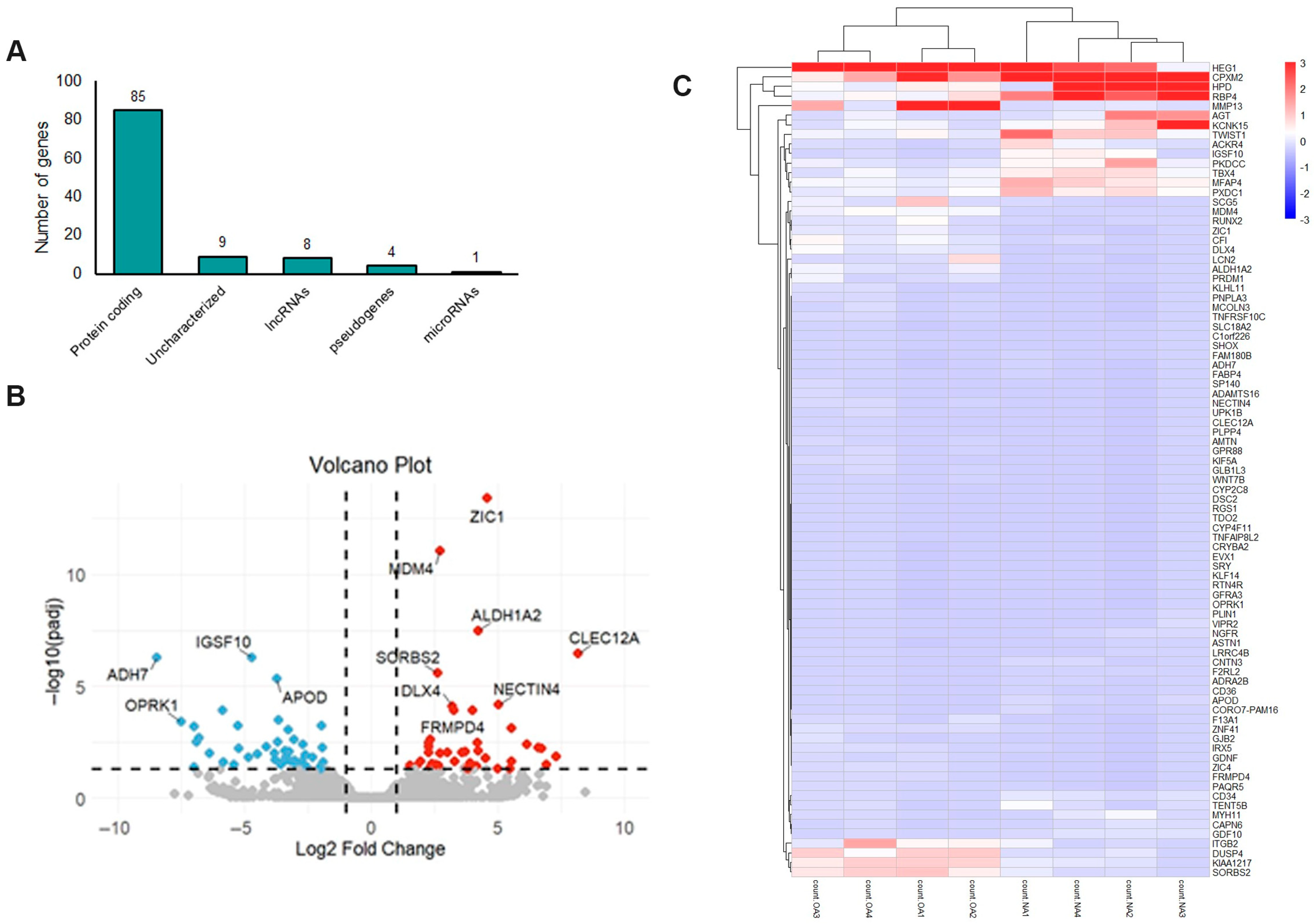
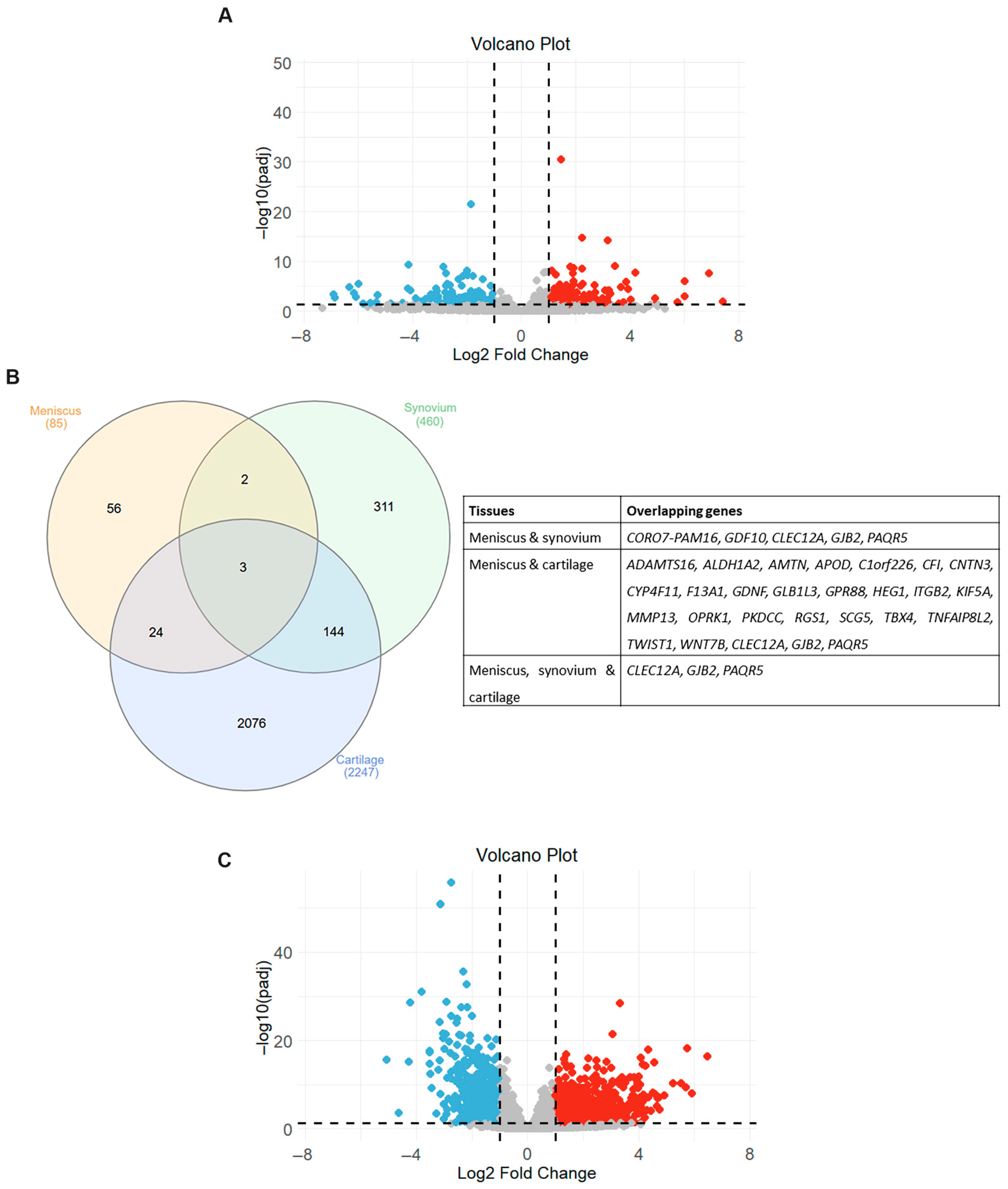
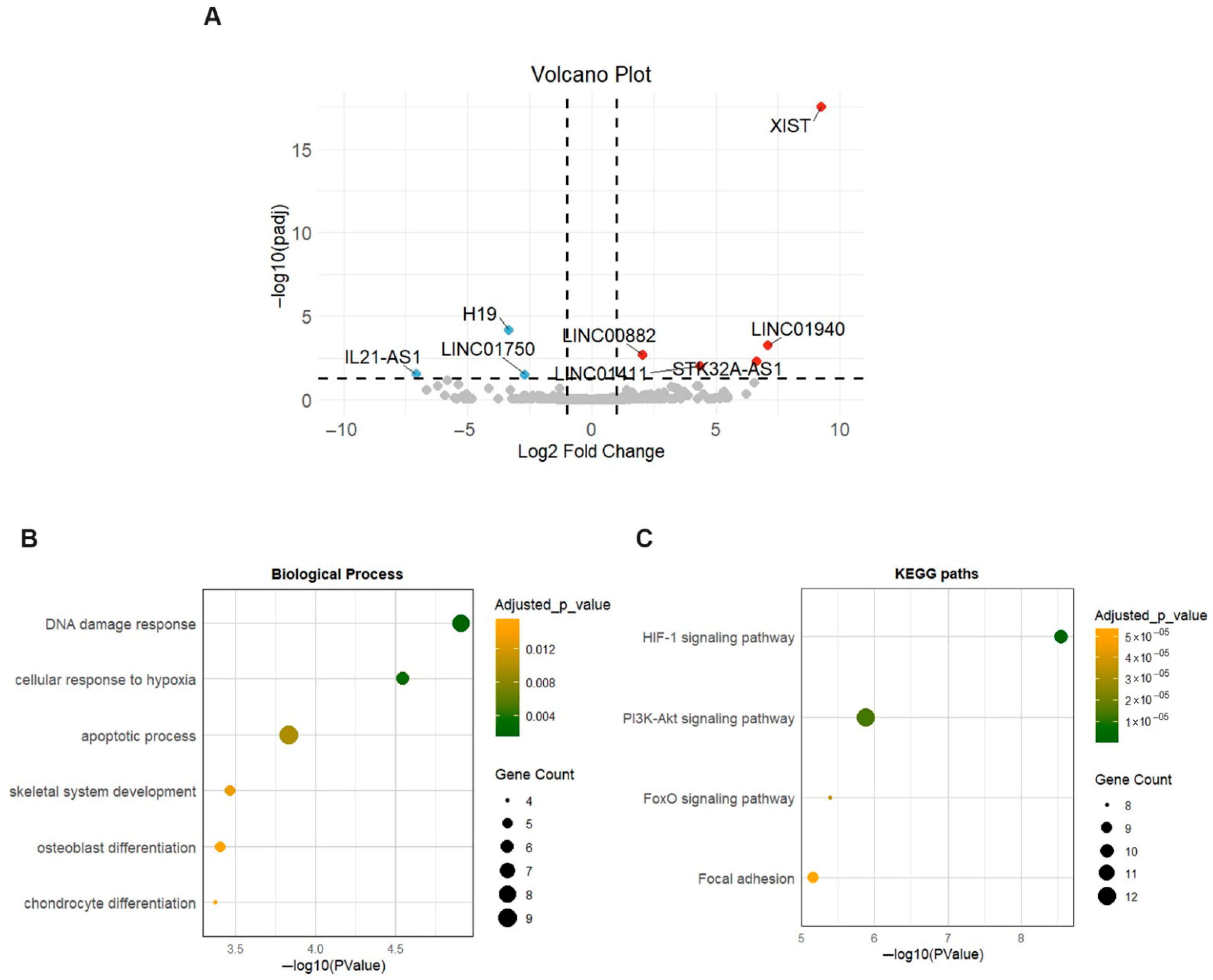
2.1. mRNA Expression Profile in OA Meniscus
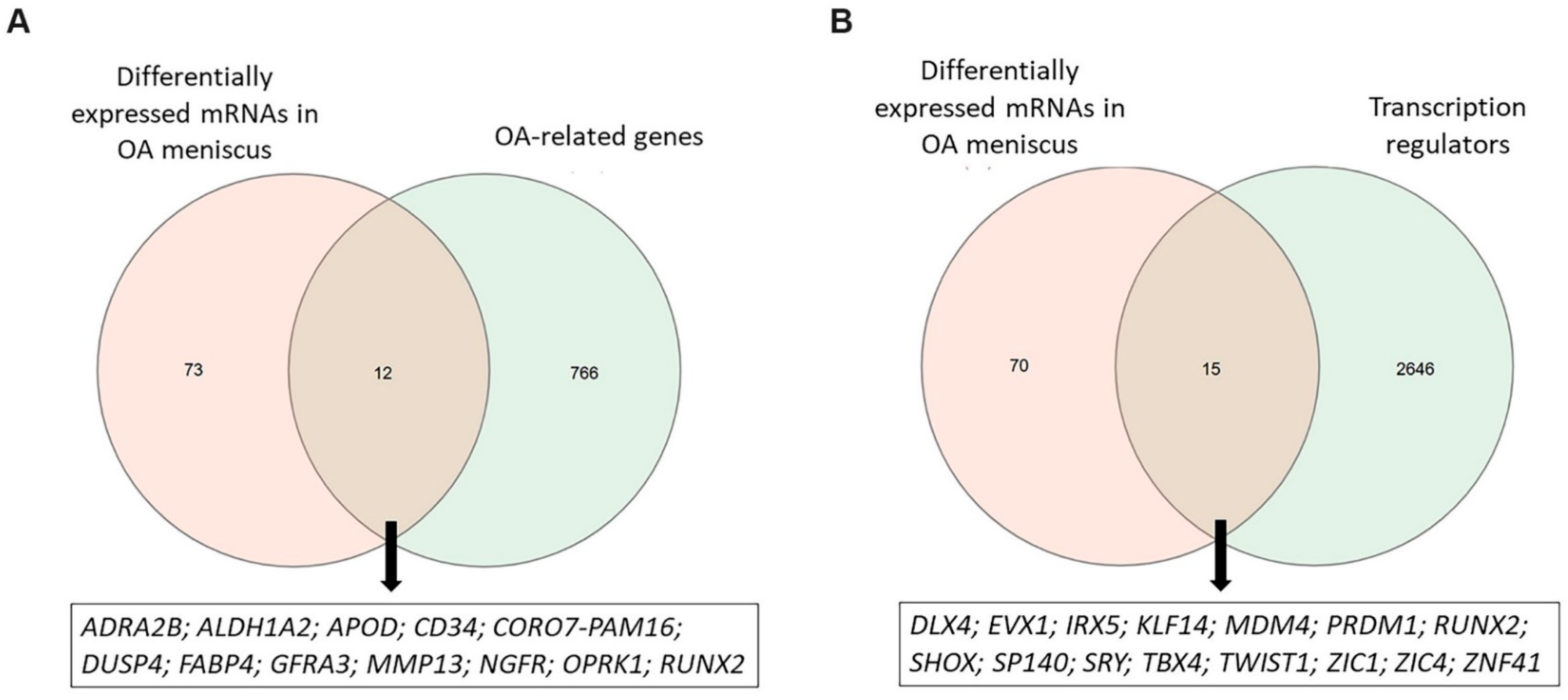
2.2. Functional Enrichment Analysis of Differentially Expressed mRNAs (DEMs) in OA Meniscus
2.3. Protein–Protein Interactions of Differentially Expressed mRNAs (DEMs) in OA Meniscus
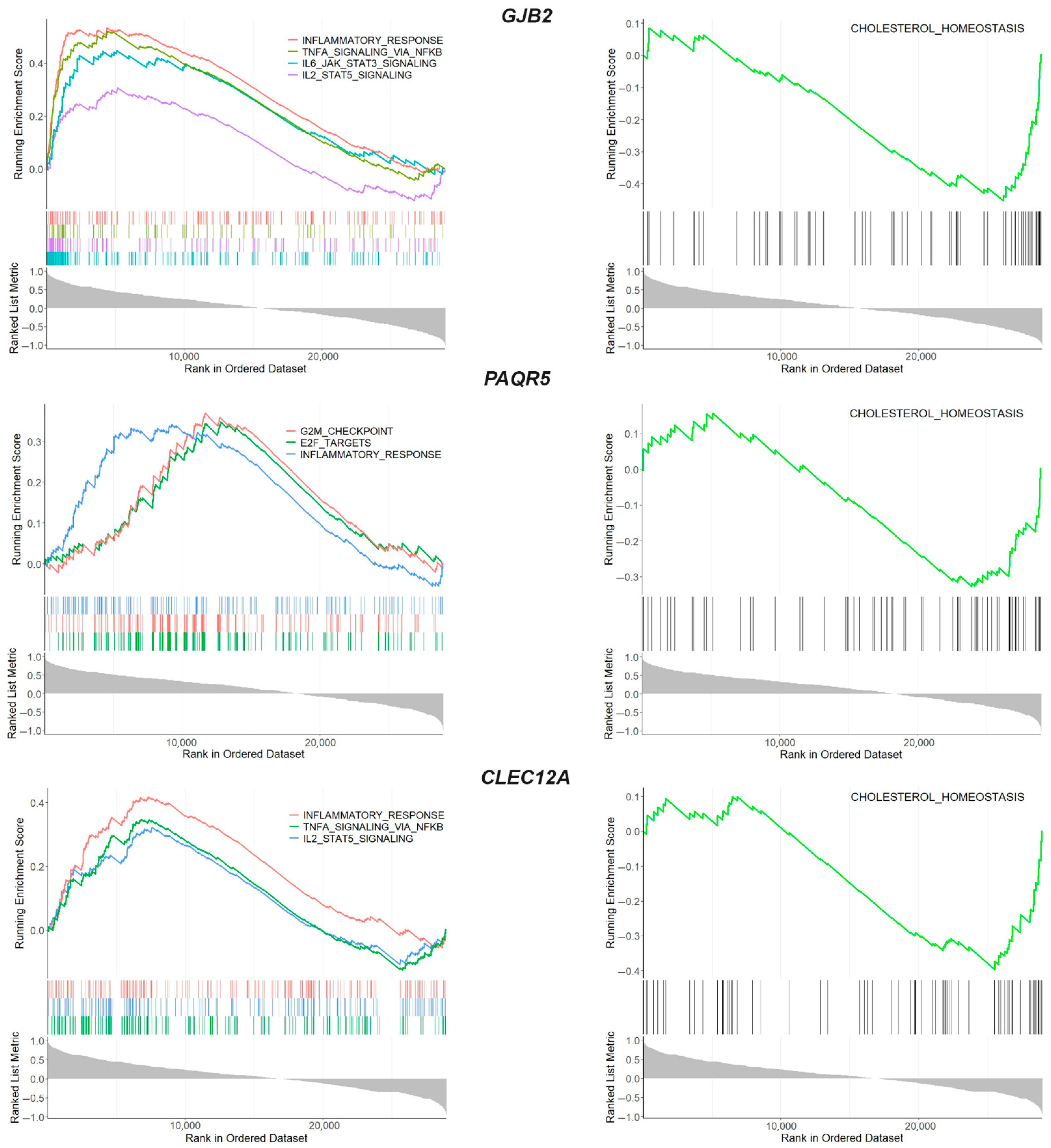
2.4. Common Transcriptional Changes in Meniscus, Synovium and Cartilage in OA
2.5. Long Non-Coding RNA (lncRNA) Profile in OA Meniscus
2.6. Drug Targets
3. Discussion
4. Materials and Methods
4.1. Datasets
4.2. Differential Gene Expression Analysis
4.3. Construction of Protein–Protein Interaction (PPI) Network
4.4. Enrichment Analysis
4.5. Identification of OA-Related Genes, Transcription Regulators and lncRNA Targets
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.Z.; Liang, X.Z.; Sun, Y.Q.; Jia, H.F.; Li, J.C.; Li, G. Global, regional, and national burdens of osteoarthritis from 1990 to 2021: Findings from the 2021 global burden of disease study. Front. Med. 2024, 11, 1476853. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Wang, X.Q.; Zhang, X.A. Exercise for Osteoarthritis: A Literature Review of Pathology and Mechanism. Front. Aging Neurosci. 2022, 14, 854026. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Kloppenburg, M.; Namane, M.; Cicuttini, F. Osteoarthritis. Lancet 2025, 405, 71–85. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Coras, R.; Torres, A.; Lane, N.E.; Guma, M. Synovial Inflammation in Osteoarthritis Progression. Nat. Rev. Rheumatol. 2022, 18, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hunter, D.J.; Jin, X.; Ding, C. The importance of synovial inflammation in osteoarthritis: Current evidence from imaging assessments and clinical trials. Osteoarthr. Cartil. 2018, 26, 165–174. [Google Scholar] [CrossRef]
- Fox, A.J.; Wanivenhaus, F.; Burge, A.J.; Warren, R.F.; Rodeo, S.A. The human meniscus: A review of anatomy, function, injury, and advances in treatment. Clin. Anat. 2015, 28, 269–287. [Google Scholar] [CrossRef]
- Englund, M.; Guermazi, A.; Lohmander, S.L. The role of the meniscus in knee osteoarthritis: A cause or consequence? Radiol. Clin. N. Am. 2009, 47, 703–712. [Google Scholar] [CrossRef]
- Ozeki, N.; Koga, H.; Sekiya, I. Degenerative Meniscus in Knee Osteoarthritis: From Pathology to Treatment. Life 2022, 12, 603. [Google Scholar] [CrossRef]
- Englund, M.; Roemer, F.W.; Hayashi, D.; Crema, M.D.; Guermazi, A. Meniscus pathology, osteoarthritis and the treatment controversy. Nat. Rev. Rheumatol. 2012, 8, 412–419. [Google Scholar] [CrossRef]
- Papalia, G.F.; Za, P.; Saccone, L.; Franceschetti, E.; Zampogna, B.; Vasta, S.; Papalia, R. Meniscal extrusion: Risk factors and diagnostic tools to predict early osteoarthritis. Orthop. Rev. 2023, 15, 74881. [Google Scholar] [CrossRef] [PubMed]
- He, A.; Ning, Y.; Wen, Y.; Cai, Y.; Xu, K.; Cai, Y.; Han, J.; Liu, L.; Du, Y.; Liang, X.; et al. Use of integrative epigenetic and mRNA expression analyses to identify significantly changed genes and functional pathways in osteoarthritic cartilage. Bone Jt. Res. 2018, 7, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Cen, X.; Huang, X.Q.; Sun, W.T.; Liu, Q.; Liu, J. Long noncoding RNAs: A new regulatory code in osteoarthritis. Am. J. Transl. Res. 2017, 9, 4747–4755. [Google Scholar] [PubMed] [PubMed Central]
- Hu, K.; Wen, H.; Song, T.; Che, Z.; Song, Y.; Song, M. Deciphering the Role of LncRNAs in Osteoarthritis: Inflammatory Pathways Unveiled. J. Inflamm. Res. 2024, 17, 6563–6581. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Z.; Ma, X.; Liu, X. Advances in Research on the Regulatory Roles of lncRNAs in Osteoarthritic Cartilage. Biomolecules 2023, 13, 580. [Google Scholar] [CrossRef]
- Shi, D.; Mei, Y.; Hao, W.; Li, J.; Liu, S.; Lin, X. Biological functions and applications of LncRNAs in the regulation of the extracellular matrix in osteoarthritis. Front. Cell Dev. Biol. 2024, 11, 1330624. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, K.; Tang, C.; Shi, Z.; Jing, K.; Zheng, J. Long non-coding RNA XIST contributes to osteoarthritis progression via miR-149-5p/DNMT3A axis. Biomed. Pharmacother. 2020, 128, 110349. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Q.; Zhang, J.; Song, C.; Han, Z.; Wang, J.; Shu, L.; Liu, W.; He, J.; Wang, P. The emerging role of lncRNAs in osteoarthritis development and potential therapy. Front. Genet. 2023, 14, 1273933. [Google Scholar] [CrossRef]
- Deng, M.; Tang, C.; Yin, L.; Jiang, Y.; Huang, Y.; Feng, Y.; Chen, C. Clinical and omics biomarkers in osteoarthritis diagnosis and treatment. J. Orthop. Transl. 2025, 50, 295–305. [Google Scholar] [CrossRef]
- Boer, C.G. Osteoarthritis year in review 2024: Genetics, genomics, and epigenetics. Osteoarthr. Cartil. 2025, 33, 50–57. [Google Scholar] [CrossRef]
- Brophy, R.H.; Zhang, B.; Cai, L.; Wright, R.W.; Sandell, L.J.; Rai, M.F. Transcriptome comparison of meniscus from patients with and without osteoarthritis. Osteoarthr. Cartil. 2018, 26, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, J.; Zou, X.; Liu, Y.; Zhao, B.; Chen, L.; Li, B.; Chen, B. Identifying autophagy-related mRNAs and potential ceRNA networks in meniscus degeneration based on RNA sequencing and experimental validation. Heliyon 2024, 10, e32782. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Du, X.; Wen, X.; Li, H.; Zeng, A.; Sun, H.; Hu, S.; He, Q.; Liao, W.; Zhang, Z. Whole-Transcriptome Sequence of Degenerative Meniscus Cells Unveiling Diagnostic Markers and Therapeutic Targets for Osteoarthritis. Front. Genet. 2021, 12, 754421. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wen, X.; Li, H.; Wu, P.; Gu, M.; Zhao, X.; Zhang, Z.; Hu, S.; Mao, G.; Ma, R.; et al. Single-cell RNA-seq analysis identifies meniscus progenitors and reveals the progression of meniscus degeneration. Ann. Rheum. Dis. 2020, 79, 408–417. [Google Scholar] [CrossRef]
- Zeng, J.; Jiang, X.; Jiang, M.; Cao, Y.; Jiang, Y. Bioinformatics analysis of hub genes as osteoarthritis prognostic biomarkers. Sci. Rep. 2023, 13, 22894. [Google Scholar] [CrossRef]
- Zhao, Y.; Lv, J.; Zhang, H.; Xie, J.; Dai, H.; Zhang, X. Gene Expression Profiles Analyzed Using Integrating RNA Sequencing, and Microarray Reveals Increased Inflammatory Response, Proliferation, and Osteoclastogenesis in Pigmented Villonodular Synovitis. Front. Immunol. 2021, 12, 665442. [Google Scholar] [CrossRef]
- Vaillancourt, M.; Desaulniers, P.; Paré, G.; Pagé, N.; Lachhab, A.; Kerever, A.; Julien, A.S.; Amiable, N.; Pelletier, M.; Tessier, P.A.; et al. Expression of the myeloid inhibitory receptor CLEC12A correlates with disease activity and cytokines in early rheumatoid arthritis. Sci. Rep. 2021, 11, 11248. [Google Scholar] [CrossRef]
- Yoshida, C.A.; Yamamoto, H.; Fujita, T.; Furuichi, T.; Ito, K.; Inoue, K.; Yamana, K.; Zanma, A.; Takada, K.; Ito, Y.; et al. Runx2 and Runx3 are essential for chondrocyte maturation, and Runx2 regulates limb growth through induction of Indian hedgehog. Genes Dev. 2004, 18, 952–963. [Google Scholar] [CrossRef]
- Zheng, Q.; Zhou, G.; Morello, R.; Chen, Y.; Garcia-Rojas, X.; Lee, B. Type X collagen gene regulation by Runx2 contributes directly to its hypertrophic chondrocyte-specific expression in vivo. J. Cell Biol. 2003, 162, 833–842. [Google Scholar] [CrossRef]
- Katsoula, G.; Lawrence, J.E.G.; Arruda, A.L.; Tutino, M.; Balogh, P.; Southam, L.; Swift, D.; Behjati, S.; Teichmann, S.A.; Wilkinson, J.M.; et al. Primary cartilage transcriptional signatures reflect cell-type-specific molecular pathways underpinning osteoarthritis. Am. J. Hum. Genet. 2024, 111, 2735–2755. [Google Scholar] [CrossRef]
- Guzzo, R.M.; Alaee, F.; Paglia, D.; Gibson, J.D.; Spicer, D.; Drissi, H. Aberrant expression of Twist1 in diseased articular cartilage and a potential role in the modulation of osteoarthritis severity. Genes Dis. 2016, 3, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Bobick, B.E.; Cobb, J. Shox2 regulates progression through chondrogenesis in the mouse proximal limb. J. Cell Sci. 2012, 125 Pt 24, 6071–6083. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Aprile, M.; Esposito, R.; Ciccodicola, A. RNA-Seq and human complex diseases: Recent accomplishments and future perspectives. Eur. J. Hum. Genet. 2013, 21, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Porcu, E.; Sadler, M.C.; Lepik, K.; Auwerx, C.; Wood, A.R.; Weihs, A.; Sleiman, M.S.B.; Ribeiro, D.M.; Bandinelli, S.; Tanaka, T.; et al. Differentially expressed genes reflect disease-induced rather than disease-causing changes in the transcriptome. Nat. Commun. 2021, 12, 5647. [Google Scholar] [CrossRef]
- Yang, C.Y.; Chanalaris, A.; Troeberg, L. ADAMTS and ADAM metalloproteinases in osteoarthritis—Looking beyond the ‘usual suspects’. Osteoarthr. Cartil. 2017, 25, 1000–1009. [Google Scholar] [CrossRef]
- Davidson, R.K.; Waters, J.G.; Kevorkian, L.; Darrah, C.; Cooper, A.; Donell, S.T.; Clark, I.M. Expression profiling of metalloproteinases and their inhibitors in synovium and cartilage. Arthritis Res. Ther. 2006, 8, R124. [Google Scholar] [CrossRef]
- Surridge, A.K.; Rodgers, U.R.; Swingler, T.E.; Davidson, R.K.; Kevorkian, L.; Norton, R.; Waters, J.G.; Goldring, M.B.; Parker, A.E.; Clark, I.M. Characterization and regulation of ADAMTS-16. Matrix Biol. 2009, 28, 416–424. [Google Scholar] [CrossRef]
- Belluzzi, E.; Olivotto, E.; Toso, G.; Cigolotti, A.; Pozzuoli, A.; Biz, C.; Trisolino, G.; Ruggieri, P.; Grigolo, B.; Ramonda, R.; et al. Conditioned media from human osteoarthritic synovium induces inflammation in a synoviocyte cell line. Connect. Tissue Res. 2019, 60, 136–145. [Google Scholar] [CrossRef]
- Olivotto, E.; Belluzzi, E.; Pozzuoli, A.; Cigolotti, A.; Scioni, M.; Goldring, S.R.; Goldring, M.B.; Ruggieri, P.; Ramonda, R.; Grigolo, B.; et al. Do synovial inflammation and meniscal degeneration impact clinical outcomes of patients undergoing arthroscopic partial meniscectomy? A histological study. Int. J. Mol. Sci. 2022, 23, 3903. [Google Scholar] [CrossRef]
- Hellio Le Graverand, M.P.; Eggerer, J.; Sciore, P.; Reno, C.; Vignon, E.; Otterness, I.; Hart, D.A. Matrix metalloproteinase-13 expression in rabbit knee joint connective tissues: Influence of maturation and response to injury. Matrix Biol. 2000, 19, 431–441. [Google Scholar] [CrossRef]
- Lei, P.; Sun, R.; Li, K.; Hu, Y.; Liao, Z. Morphological changes and expression of MMPs and TIMPs in rabbit degenerated lateral meniscus after PCL-transection. Int. J. Clin. Exp. Med. 2015, 8, 17950–17958. [Google Scholar] [PubMed] [PubMed Central]
- Baran, K.; Czechowska, A.; Kopacz, K.; Padula, G.; Migdalska-Sęk, M.; Tomaszewski, W.; Nowak, K.; Domżalski, M.; Brzeziańska-Lasota, E. MMP13 mRNA Expression Level as a Potential Marker for Knee OA Progression-An Observational Study. J. Clin. Med. 2025, 14, 1263. [Google Scholar] [CrossRef]
- D’Angelo, M.; Yan, Z.; Nooreyazdan, M.; Pacifici, M.; Sarment, D.S.; Billings, P.C.; Leboy, P.S. MMP-13 is induced during chondrocyte hypertrophy. J. Cell. Biochem. 2000, 77, 678–693. [Google Scholar] [CrossRef]
- Loeser, R.F. Integrins and chondrocyte-matrix interactions in articular cartilage. Matrix Biol. 2014, 39, 11–16. [Google Scholar] [CrossRef]
- Sun, Y.; Mauerhan, D.R.; Honeycutt, P.R.; Kneisl, J.S.; Norton, J.H.; Hanley, E.N., Jr.; Gruber, H.E. Analysis of meniscal degeneration and meniscal gene expression. BMC Musculoskelet. Disord. 2010, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, N. Genome-wide expression and methylation profiles reveal candidate genes and biological processes underlying synovial inflammatory tissue of patients with osteoarthritis. Int. J. Rheum. Dis. 2015, 18, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Li, Z. Expression and diagnostic significance of integrin beta-2 in synovial fluid of patients with osteoarthritis. J. Orthop. Surg. 2023, 31, 10225536221147213. [Google Scholar] [CrossRef]
- Selim, A.M.; Elsabagh, Y.A.; El-Sawalhi, M.M.; Ismail, N.A.; Senousy, M.A. Association of integrin-β2 polymorphism and expression with the risk of rheumatoid arthritis and osteoarthritis in Egyptian patients. BMC Med. Genom. 2023, 16, 204. [Google Scholar] [CrossRef]
- Hu, T.; Zhang, Z.; Deng, C.; Ma, X.; Liu, X. Effects of β2 Integrins on Osteoclasts, Macrophages, Chondrocytes, and Synovial Fibroblasts in Osteoarthritis. Biomolecules 2022, 12, 1653. [Google Scholar] [CrossRef]
- Chawla, S.; Mainardi, A.; Majumder, N.; Dönges, L.; Kumar, B.; Occhetta, P.; Martin, I.; Egloff, C.; Ghosh, S.; Bandyopadhyay, A.; et al. Chondrocyte Hypertrophy in Osteoarthritis: Mechanistic Studies and Models for the Identification of New Therapeutic Strategies. Cells 2022, 11, 4034. [Google Scholar] [CrossRef]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Otto, F.; Thornell, A.P.; Crompton, T.; Denzel, A.; Gilmour, K.C.; Rosewell, I.R.; Stamp, G.W.; Beddington, R.S.; Mundlos, S.; Olsen, B.R.; et al. Cbfa1, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell 1997, 89, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Kim, D.J.; Shen, J.; Zou, Z.; O’Keefe, R.J. Runx2 plays a central role in Osteoarthritis development. J. Orthop. Transl. 2019, 23, 132–139. [Google Scholar] [CrossRef]
- Komori, T. Molecular Mechanism of Runx2-Dependent Bone Development. Mol. Cells 2020, 43, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Boer, C.G.; Hatzikotoulas, K.; Southam, L.; Stefánsdóttir, L.; Zhang, Y.; Coutinho de Almeida, R.; Wu, T.T.; Zheng, J.; Hartley, A.; Teder-Laving, M.; et al. Deciphering osteoarthritis genetics across 826,690 individuals from 9 populations. Cell 2021, 184, 4784–4818.e17. [Google Scholar] [CrossRef]
- Alvarez-Garcia, O.; Fisch, K.M.; Wineinger, N.E.; Akagi, R.; Saito, M.; Sasho, T.; Su, A.I.; Lotz, M.K. Increased DNA Methylation and Reduced Expression of Transcription Factors in Human Osteoarthritis Cartilage. Arthritis Rheumatol. 2016, 68, 1876–1886. [Google Scholar] [CrossRef]
- Duboc, V.; Sulaiman, F.A.; Feneck, E.; Kucharska, A.; Bell, D.; Holder-Espinasse, M.; Logan, M.P.O. Tbx4 function during hindlimb development reveals a mechanism that explains the origins of proximal limb defects. Development 2021, 148, dev199580. [Google Scholar] [CrossRef]
- Liu, L.; Luo, P.; Yang, M.; Wang, J.; Hou, W.; Xu, P. The role of oxidative stress in the development of knee osteoarthritis: A comprehensive research review. Front. Mol. Biosci. 2022, 9, 1001212. [Google Scholar] [CrossRef]
- Ahmad, N.; Ansari, M.Y.; Haqqi, T.M. Role of iNOS in osteoarthritis: Pathological and therapeutic aspects. J. Cell. Physiol. 2020, 235, 6366–6376. [Google Scholar] [CrossRef]
- Abramson, S.B. Nitric oxide in inflammation and pain associated with osteoarthritis. Arthritis Res. Ther. 2008, 10 (Suppl. S2), S2. [Google Scholar] [CrossRef]
- Koike, M.; Nojiri, H.; Kanazawa, H.; Yamaguchi, H.; Miyagawa, K.; Nagura, N.; Banno, S.; Iwase, Y.; Kurosawa, H.; Kaneko, K. Superoxide dismutase activity is significantly lower in end-stage osteoarthritic cartilage than non-osteoarthritic cartilage. PLoS ONE 2018, 13, e0203944. [Google Scholar] [CrossRef]
- Jiang, H.; Ji, P.; Shang, X.; Zhou, Y. Connection between Osteoarthritis and Nitric Oxide: From Pathophysiology to Therapeutic Target. Molecules 2023, 28, 1683. [Google Scholar] [CrossRef]
- Dbouk, H.A.; Mroue, R.M.; El-Sabban, M.E.; Talhouk, R.S. Connexins: A myriad of functions extending beyond assembly of gap junction channels. Cell Commun. Signal. 2009, 7, 4. [Google Scholar] [CrossRef]
- Li, C.; Zheng, Z. Identification of novel targets of knee osteoarthritis shared by cartilage and synovial tissue. Int. J. Mol. Sci. 2020, 21, 6033. [Google Scholar] [CrossRef]
- Zappalà, A.; Romano, I.R.; D’Angeli, F.; Musumeci, G.; Lo Furno, D.; Giuffrida, R.; Mannino, G. Functional roles of connexins and gap junctions in osteo-chondral cellular components. Int. J. Mol. Sci. 2023, 24, 4156. [Google Scholar] [CrossRef] [PubMed]
- Mayan, M.D.; Carpintero-Fernandez, P.; Gago-Fuentes, R.; Martinez-de-Ilarduya, O.; Wang, H.-Z.; Valiunas, V.; Brink, P.; Blanco, F.J. Human articular chondrocytes express multiple gap junction proteins: Differential expression of connexins in normal and osteoarthritic cartilage. Am. J. Pathol. 2013, 182, 1337–1346. [Google Scholar] [CrossRef]
- Stains, J.P.; Civitelli, R. Gap junctions in skeletal development and function. Biochim. Biophys. Acta 2005, 1719, 69–81. [Google Scholar] [CrossRef]
- Ammons, M.C.B.; Siemsen, D.W.; Nelson-Overton, L.K.; Quinn, M.T.; Gauss, K.A. Binding of pleomorphic adenoma gene-like 2 to the tumor necrosis factor (TNF)-alpha-responsive region of the NCF2 promoter regulates p67phox expression and NADPH oxidase activity. J. Biol. Chem. 2007, 282, 17941–17952. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Wang, H.; Wu, C.; Shi, Y.; Li, H.; Bao, X.; Yang, Y.; Han, S.; Yang, X.; Tao, J.; et al. PAQR5 drives the malignant progression and shapes the immunosuppressive microenvironment of hepatocellular carcinoma by activating the NF-κB signaling. Biomark. Res. 2025, 13, 70. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Liu, W.; Yan, X.; Yang, M.; Yao, S.; Shu, Q.; Li, B.; Zhu, R. PAQR5 expression is suppressed by TGFβ1 and associated with a poor survival outcome in renal clear cell carcinoma. Front. Oncol. 2022, 11, 827344. [Google Scholar] [CrossRef] [PubMed]
- Neumann, K.; Castiñeiras-Vilariño, M.; Höckendorf, U.; Hannesschläger, N.; Lemeer, S.; Kupka, D.; Meyermann, S.; Lech, M.; Anders, H.-J.; Kuster, B.; et al. Clec12a is an inhibitory receptor for uric acid crystals that regulates inflammation in response to cell death. Immunity 2014, 40, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Barra, L.; Saunders, S.; Mangion, M.; Paré, G.; Maaroufi, H.; Garnier, A.; Cairns, E.; Fernandes, M.J. Identification of autoantibodies targeting citrullinated CLEC12A in rheumatoid arthritis patients. J. Transl. Autoimmun. 2025, 10, 100287. [Google Scholar] [CrossRef]
- Knights, A.J.; Redding, S.J.; Maerz, T. Inflammation in osteoarthritis: The latest progress and ongoing challenges. Curr. Opin. Rheumatol. 2023, 35, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Papathanasiou, I.; Anastasopoulou, L.; Tsezou, A. Cholesterol metabolism related genes in osteoarthritis. Bone 2021, 152, 116076. [Google Scholar] [CrossRef]
- Wang, T.; Liu, Y.; Wang, Y.; Huang, X.; Zhao, W.; Zhao, Z. Long non-coding RNA XIST promotes extracellular matrix degradation by functioning as a competing endogenous RNA of miR-1277-5p in osteoarthritis. Int. J. Mol. Med. 2019, 44, 630–642. [Google Scholar] [CrossRef]
- Sun, P.; Wu, Y.; Li, X.; Jia, Y. miR-142-5p protects against osteoarthritis through competing with lncRNA XIST. J. Gene Med. 2020, 22, e3158. [Google Scholar] [CrossRef]
- Li, L.; Lv, G.; Wang, B.; Kuang, L. The role of lncRNA XIST/miR-211 axis in modulating the proliferation and apoptosis of osteoarthritis chondrocytes through CXCR4 and MAPK signaling. Biochem. Biophys. Res. Commun. 2018, 503, 2555–2562. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Mondanelli, N.; Seccafico, I.; Corsaro, R.; Moretti, E.; Collodel, G.; Fioravanti, A. Role of lncRNA XIST/miR-146a axis in matrix degradation and apoptosis of osteoarthritic chondrocytes through regulation of MMP-13 and BCL2. Biology 2025, 14, 221. [Google Scholar] [CrossRef]
- Wang, C.; Lu, C.; Zou, L.; He, D. Bioinformatic analysis of lncRNA-associated competing endogenous RNA regulatory networks in synovial tissue of temporomandibular joint osteoarthritis. BIOCELL 2023, 47, 1293–1306. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, L.; Zheng, Y.; Liu, W.; Xu, Y. Kaempferol attenuates the effects of XIST/miR-130a/STAT3 on inflammation and extracellular matrix degradation in osteoarthritis. Future Med. Chem. 2021, 13, 1451–1464. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, J.; Xu, F.; Ji, E.; Wang, C.; Pan, Z. Long noncoding RNA H19 alleviates inflammation in osteoarthritis through interactions between TP53, IL-38, and IL-36 receptor. Bone Jt. Res. 2022, 11, 594–607. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Xu, L.; Wang, S. Regulation of lncRNA-H19/miR-140-5p in cartilage matrix degradation and calcification in osteoarthritis. Ann. Palliat. Med. 2020, 9, 1896–1904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, X.; Ni, X.; Feng, P.; Wang, Y.U. Long non-coding RNA H19 modulates proliferation and apoptosis in osteoarthritis via regulating miR-106a-5p. J. Biosci. 2019, 44, 128. [Google Scholar] [CrossRef]
- Steck, E.; Boeuf, S.; Gabler, J.; Werth, N.; Schnatzer, P.; Diederichs, S.; Richter, W. Regulation of H19 and its encoded microRNA-675 in osteoarthritis and under anabolic and catabolic in vitro conditions. J. Mol. Med. 2012, 90, 1185–1195. [Google Scholar] [CrossRef]
- Dai, G.; Xiao, H.; Zhao, C.; Chen, H.; Liao, J.; Huang, W. LncRNA H19 regulates BMP2-induced hypertrophic differentiation of mesenchymal stem cells by promoting Runx2 phosphorylation. Front. Cell Dev. Biol. 2020, 8, 580. [Google Scholar] [CrossRef]
- Liao, J.; Chen, B.; Zhu, Z.; Du, C.; Gao, S.; Zhao, G.; Zhao, P.; Wang, Y.; Wang, A.; Schwartz, Z.; et al. Long noncoding RNA (lncRNA) H19: An essential developmental regulator with expanding roles in cancer, stem cell differentiation, and metabolic diseases. Genes Dis. 2023, 10, 1351–1366. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zou, M.; Li, J.; Wang, B.; Zhang, Q.; Liu, F.; Lü, G. LncRNA H19 targets miR-22 to modulate H2O2-induced deregulation in nucleus pulposus cell senescence, proliferation, and ECM synthesis through Wnt signaling. J. Cell. Biochem. 2018, 119, 4990–5002. [Google Scholar] [CrossRef]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a Promising Target for the Treatment of Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 1742. [Google Scholar] [CrossRef]
- Xie, J.; Xu, X.; Yang, M.; Yu, H.; Hao, J.; Yang, D.; Xu, P. New Insights on the Therapeutic Potential of Runt-Related Transcription Factor 2 for Osteoarthritis: Evidence from Mendelian Randomization. Rheumatol. Ther. 2024, 11, 1001–1009. [Google Scholar] [CrossRef]
- Cecil, D.L.; Appleton, C.T.; Polewski, M.D.; Mort, J.S.; Schmidt, A.M.; Bendele, A.; Beier, F.; Terkeltaub, R. The pattern recognition receptor CD36 is a chondrocyte hypertrophy marker associated with suppression of catabolic responses and promotion of repair responses to inflammatory stimuli. J. Immunol. 2009, 182, 5024–5031. [Google Scholar] [CrossRef]
- Kuda, O.; Pietka, T.A.; Demianova, Z.; Kudova, E.; Cvacka, J.; Kopecky, J.; Abumrad, N.A. Sulfo-N-succinimidyl oleate (SSO) inhibits fatty acid uptake and signaling for intracellular calcium via binding CD36 lysine 164: SSO also inhibits oxidized low density lipoprotein uptake by macrophages. J. Biol. Chem. 2013, 288, 15547–15555. [Google Scholar] [CrossRef] [PubMed]
- Mansor, L.S.; Fialho, M.d.L.S.; Yea, G.; Coumans, W.A.; West, J.A.; Kerr, M.; Carr, C.A.; Luiken, J.J.F.P.; Glatz, J.F.C.; Evans, R.D.; et al. Inhibition of sarcolemmal FAT/CD36 by sulfo-N-succinimidyl oleate rapidly corrects metabolism and restores function in the diabetic heart following hypoxia/reoxygenation. Cardiovasc. Res. 2017, 113, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Sagar, D.; Singh, N.P.; Ginwala, R.; Huang, X.; Philip, R.; Nagarkatti, M.; Nagarkatti, P.; Neumann, K.; Ruland, J.; Andrews, A.M.; et al. Antibody blockade of CLEC12A delays EAE onset and attenuates disease severity by impairing myeloid cell CNS infiltration and restoring positive immunity. Sci. Rep. 2017, 7, 2707. [Google Scholar] [CrossRef] [PubMed]
- Nickbarg, E.B.; Spencer, K.B.; Mortison, J.D.; Lee, J.T. Targeting RNA with small molecules: Lessons learned from Xist RNA. RNA 2023, 29, 463–472. [Google Scholar] [CrossRef]
- Fisch, K.M.; Gamini, R.; Alvarez-Garcia, O.; Akagi, R.; Saito, M.; Muramatsu, Y.; Sasho, T.; Koziol, J.A.; Su, A.I.; Lotz, M.K. Identification of transcription factors responsible for dysregulated networks in human osteoarthritis cartilage by global gene expression analysis. Osteoarthr. Cartil. 2018, 26, 1531–1538. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wickham, H.; Navarro, D.; Lin Pedersen, T. ggplot2: Elegant Graphics for Data Analysis, 3rd ed.; Springer: New York, NY, USA, 2016; Available online: https://ggplot2-book.org/ (accessed on 3 June 2025).
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Sherman, B.T.; Panzade, G.; Imamichi, T.; Chang, W. DAVID Ortholog: An integrative tool to enhance functional analysis through orthologs. Bioinformatics 2024, 40, btae615. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Koscielny, G.; An, P.; Carvalho-Silva, D.; Cham, J.A.; Fumis, L.; Gasparyan, R.; Hasan, S.; Karamanis, N.; Maguire, M.; Papa, E.; et al. Open Targets: A platform for therapeutic target identification and validation. Nucleic Acids Res. 2017, 45, D985–D994. [Google Scholar] [CrossRef]
- Shen, W.K.; Chen, S.Y.; Gan, Z.Q.; Zhang, Y.Z.; Yue, T.; Chen, M.M.; Xue, Y.; Hu, H.; Guo, A.Y. AnimalTFDB 4.0: A comprehensive animal transcription factor database updated with variation and expression annotations. Nucleic Acids Res. 2023, 51, D39–D45. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, P.; Tian, R.; Wang, S.; Guo, Q.; Luo, M.; Zhou, W.; Liu, G.; Jiang, H.; Jiang, Q. LncRNA2Target v2.0: A comprehensive database for target genes of lncRNAs in human and mouse. Nucleic Acids Res. 2019, 47, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | p-Value | Genes |
|---|---|---|
| A. GO Biological Processes | ||
| Retinol metabolic process | 0.001 | CYP2C8, RBP4, ALDH1A2, ADH7 |
| Ossification | 0.006 | IGSF10, CORO7-PAM16, TWIST1, RUNX2 |
| Embryonic skeletal system development | 0.01 | KIAA1217, RBP4, DLX4 |
| Bone mineralization | 0.03 | MMP13, PKDCC, RUNX2 |
| B. GO Cellular Components | ||
| Extracellular region | 1.14 × 10−4 | NGFR, PKDCC, CPXM2, HEG1, WNT7B, CFI, F13A1, AGT, ADAMTS16, IGSF10, MFAP4, RBP4, MMP13, GDNF, SCG5, AMTN, FAM180B, LCN2, CNTN3, APOD, CD34, F2RL2 |
| Plasma membrane | 0.05 | RTN4R, VIPR2, CLEC12A, GPR88, CFI, ITGB2, ADH7, RGS1, CD36, CD34, SLC18A2, LRRC4B, NGFR, MCOLN3, PAQR5, WNT7B, TNFRSF10C, KCNK15, OPRK1, SORBS2, GFRA3, ADRA2B, GJB2, CYP2C8, PLPP4, ACKR4, CNTN3, NECTIN4, DSC2, F2RL2 |
| Specific granule membrane | 0.05 | CLEC12A, ITGB2, CD36 |
| C. GO Molecular Functions | ||
| DNA-binding transcription factor activity, RNA polymerase II-specific | 0.008 | DLX4, IRX5, SP140, TWIST1, ZIC4, EVX1, KLF14, TBX4, RUNX2, ZNF41, ZIC1, SHOX, SRY |
| Cytoskeletal motor activity | 0.016 | GPR88, KIF5A, MYH11 |
| D. Pathway Enrichment Analysis | ||
| PPAR signaling pathway | 0.046 | FABP4, PLIN1, CD36 |
| Interleukin-4 and interleukin-13 signaling | 0.0022 | ITGB2, LCN2, F13A1, TWIST1, CD36 |
| Metabolism of lipids | 0.037 | CYP2C8, FABP4, PNPLA3, PLIN1, CD36, TNFAIP8L2, CYP4F11, AGT, GLB1L3 |
| Extracellular matrix organization | 0.05 | ADAMTS16, MFAP4, MMP13, CAPN6, ITGB2 |
| Meniscus | Synovium | |||
|---|---|---|---|---|
| Gene Symbol | Log2FC | Adj. p-Value | Log2FC | Adj. p-Value |
| CORO7-PAM16 | −4.088 | 0.0149 | 3.091 | 9.40 × 10−9 |
| GDF10 | −3.756 | 0.0397 | 4.715 | 0.0003 |
| GJB2 | 3.689 | 0.0481 | 7.299 | 1.09 × 10−10 |
| PAQR5 | 4.240 | 0.0092 | 3.366 | 0.0266 |
| CLEC12A | 6.475 | 3.45 × 10−7 | 3.729 | 0.0097 |
| Meniscus | Cartilage | |||
|---|---|---|---|---|
| Gene Symbol | Log2FC | Adj. p-Value | Log2FC | Adj. p-Value |
| TBX4 | −4.973 | 0.0006 | −1.552 | 1.95 × 10−6 |
| C1orf226 | −4.277 | 0.0082 | −1.166 | 0.0051 |
| APOD | −5.988 | 4.62 × 10−6 | −1.646 | 0.0400 |
| OPRK1 | −5.090 | 0.0004 | −2.461 | 0.0170 |
| GLB1L3 | 6.139 | 0.0040 | 1.599 | 4.43 × 10−5 |
| WNT7B | 5.558 | 0.0220 | 1.880 | 0.0063 |
| AMTN | 5.543 | 0.0007 | 4.517 | 8.24 × 10−9 |
| TNFAIP8L2 | 5.482 | 0.0496 | 1.446 | 0.0169 |
| ALDH1A2 | 4.224 | 3.26 × 10−8 | 1.039 | 0.0022 |
| ADAMTS16 | 4.178 | 0.0373 | 1.303 | 0.0088 |
| MMP13 | 3.587 | 0.0091 | 3.699 | 2.94 × 10−12 |
| SCG5 | 3.310 | 0.0220 | 1.285 | 0.0284 |
| GDNF | 3.025 | 0.0095 | 2.961 | 0.0002 |
| CFI | 2.738 | 0.0096 | 3.324 | 5.09 × 10−29 |
| GPR88 | 2.356 | 0.0457 | 1.134 | 0.0117 |
| ITGB2 | 2.337 | 0.0399 | 3.373 | 3.14 × 10−6 |
| HEG1 | 1.962 | 0.0220 | 1.436 | 9.48 × 10−7 |
| TWIST1 | 1.527 | 0.0352 | 1.773 | 1.89 × 10−5 |
| RGS1 | 6.919 | 0.0334 | −1.316 | 1.62 × 10−6 |
| CYP4F11 | 6.586 | 0.0054 | −1.375 | 0.0280 |
| F13A1 | 4.190 | 0.0034 | −1.819 | 3.41 × 10−6 |
| KIF5A | −1.969 | 0.0480 | 1.521 | 0.0145 |
| PKDCC | −2.290 | 0.0149 | 1.565 | 0.0008 |
| CNTN3 | −3.167 | 0.0227 | 2.232 | 0.0328 |
| GJB2 | 3.689 | 0.0481 | 3.388 | 7.59 × 10−6 |
| PAQR5 | 4.240 | 0.0092 | 1.774 | 0.0210 |
| CLEC12A | 8.168 | 3.45 × 10−7 | 2.688 | 0.0234 |
| Dataset | Platform | Manufacturer | Group | Tissue | |
|---|---|---|---|---|---|
| Normal (Mean Age) | OA (Mean Age) | ||||
| GSE186064 | GPL24676 | Illumina NovaSeq 6000 | 4 (38.6) | 4 (unknown) | meniscus |
| GSE114007 | GPL11154 GPL18573 | Illumina HiSeq 2000 Illumina NextSeq 500 | 18 (38 years) | 20 (66 years) | cartilage |
| GSE143514 | GPL20795 | Illumina HiSeq X Ten | 3 (unknown) | 5 (unknown) | synovium |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papageorgiou, A.-A.; Balis, C.; Papathanasiou, I. Identification of Genes Linked to Meniscal Degeneration in Osteoarthritis: An In Silico Analysis. Int. J. Mol. Sci. 2025, 26, 6651. https://doi.org/10.3390/ijms26146651
Papageorgiou A-A, Balis C, Papathanasiou I. Identification of Genes Linked to Meniscal Degeneration in Osteoarthritis: An In Silico Analysis. International Journal of Molecular Sciences. 2025; 26(14):6651. https://doi.org/10.3390/ijms26146651
Chicago/Turabian StylePapageorgiou, Aliki-Alexandra, Charalampos Balis, and Ioanna Papathanasiou. 2025. "Identification of Genes Linked to Meniscal Degeneration in Osteoarthritis: An In Silico Analysis" International Journal of Molecular Sciences 26, no. 14: 6651. https://doi.org/10.3390/ijms26146651
APA StylePapageorgiou, A.-A., Balis, C., & Papathanasiou, I. (2025). Identification of Genes Linked to Meniscal Degeneration in Osteoarthritis: An In Silico Analysis. International Journal of Molecular Sciences, 26(14), 6651. https://doi.org/10.3390/ijms26146651