
Extracellular Vesicles as Biomarkers in Chronic Hepatobiliary Diseases: An Overview of Their Interplay

 ,
,  , ,
, , 
 ,
,
Abstract
1. Introduction
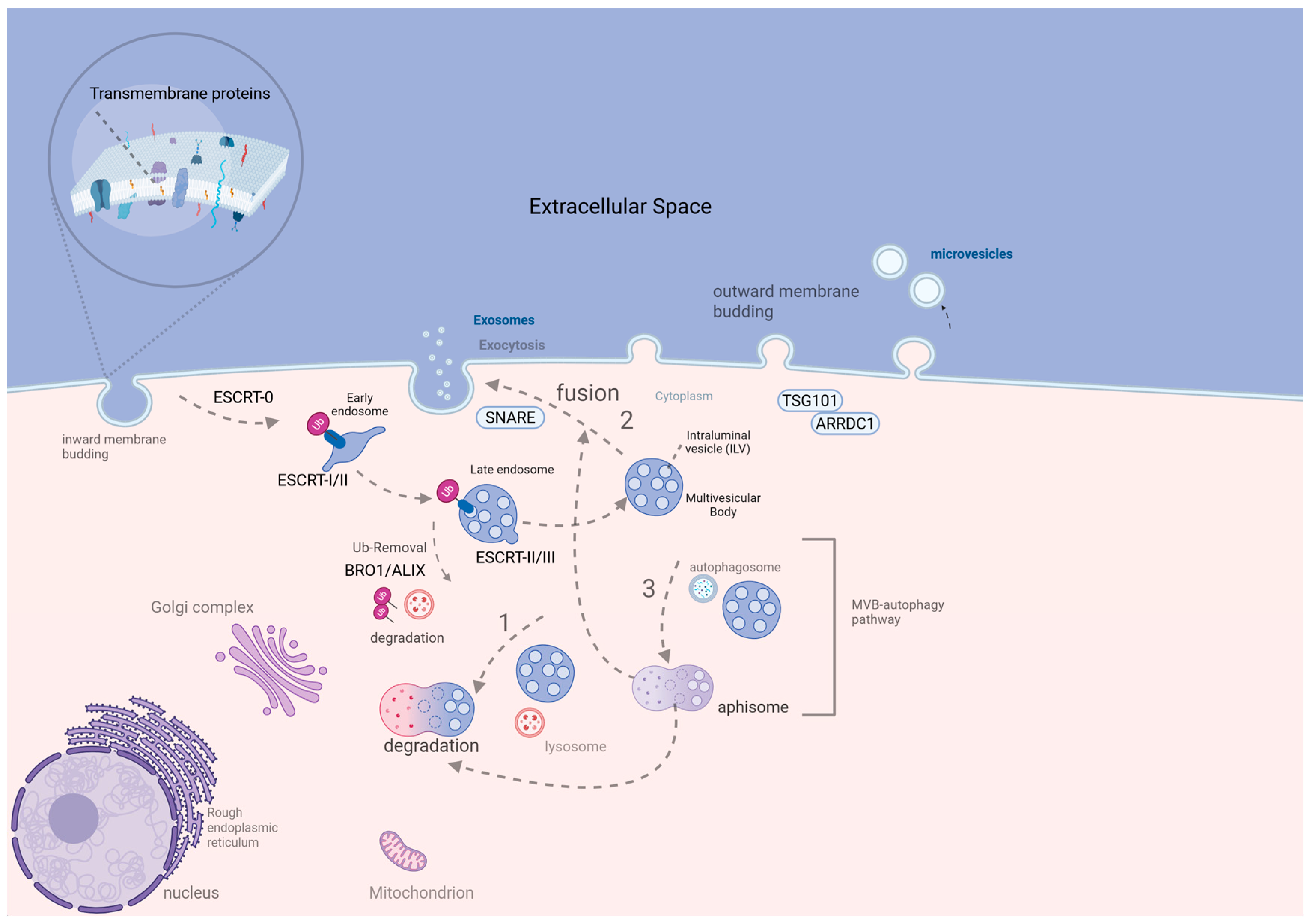
2. Review of EV Biogenesis
3. An Overview of the Interplay Between EV and Hepatobiliary Diseases and Their Potential Role as Biomarkers
3.1. Chronic Viral Hepatitis B and C
3.2. Metabolic Diseases
3.2.1. MASLD
3.2.2. Alcohol-Associated Liver Disease (ALD)
3.2.3. Hereditary Hemochromatosis (HH)
3.3. Autoimmune Hepatobiliary Diseases
3.3.1. PBC and PSC
3.3.2. Autoimmune Hepatitis (AIH)
3.4. Complications of Chronic Hepatobiliary Diseases
3.5. Fibrotic Injury and Cirrhosis
3.6. Portal Hypertension and Hepatopulmonary and Porto-Pulmonary Syndromes
3.7. Coagulation Disorders
3.8. Ascites and Hepatic Encephalopathy
3.9. Biliary Tract Stenosis and Chronic Cholecystitis
3.10. Gallbladder Cancer (GC)
3.11. Hepatocellular Cancer
3.12. Cholangiocarcinoma (CCA)
4. Limitations of EVs Utilization as Biomarkers
5. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| A | |
| AFP | Alpha-fetoprotein |
| AIH | Autoimmune hepatitis |
| ALD | Alcohol-associated liver disease |
| AMPK | AMP-activated protein kinase |
| ANA | Anti-nuclear antibodies |
| ApoEVs | Apoptotic extracellular vesicles |
| ARF6 | ADP-ribosylation factor 6 |
| ARRDC1 | Arrestin domain-containing protein 1 |
| ASGPR1 | Asialoglycoprotein receptor 1 |
| ASMA | Anti-smooth muscle antibody |
| B | |
| B4GALT3 | Beta-1,4-galactosyltransferase 3 |
| BMI1 | B lymphoma Mo-MLV insertion region 1 homolog |
| BM-EVs | Bone marrow-derived extracellular vesicles (context inferred) |
| C | |
| CAFs | Cancer-associated fibroblasts |
| CA19-9 | Carbohydrate antigen 19-9 |
| CCL | Chemokine (C-C motif) ligand |
| CCA | Cholangiocarcinoma |
| CDKN2D | Cyclin-dependent kinase inhibitor 2D |
| CEA | Carcinoembryonic antigen |
| circRNA | Circular RNA |
| CK18 | Cytokeratin 18 |
| CP | Child |
| Pugh | |
| CXCL | C-X-C motif chemokine ligand |
| D | |
| DM | Diabetes mellitus |
| DMT1 | Divalent metal transporter 1 |
| dCCA | Distal cholangiocarcinoma |
| E | |
| ECM | Extracellular matrix |
| EGFR | Epidermal growth factor receptor |
| EMT | Epithelial-to-mesenchymal transition |
| ERK | Extracellular signal-regulated kinases |
| ESCRT | Endosomal sorting complex required for transport |
| EV | Extracellular vesicle |
| F | |
| FAP | Fibroblast activation protein |
| FZD10 | Frizzled class receptor 10 |
| F1–F4 | Fibrosis/cirrhosis staging |
| G | |
| GC | Gallbladder cancer |
| GI | Gastrointestinal |
| GMP | Good manufacturing practice |
| (γ-GT) | Gamma-glutamyl transferase (inferred) |
| H | |
| HBcAg | Hepatitis B core antigen |
| HBsAg | Hepatitis B surface antigen |
| HBV | Hepatitis B virus |
| HCC | Hepatocellular carcinoma |
| HCV | Hepatitis C virus |
| HE | Hepatic encephalopathy |
| HH | Hereditary hemochromatosis |
| HMGA2 | High-mobility group AT-Hook 2 |
| HSCs | Hepatic stellate cells |
| HSP | Heat shock protein |
| HUVECs | Human umbilical vein endothelial cells |
| HuCCT1 | HuCCT1 cholangiocarcinoma cell line |
| I | |
| IL | Interleukin |
| ILVs | Intraluminal vesicles |
| INF-γ | Interferon gamma |
| iCCA | Intrahepatic cholangiocarcinoma |
| ITGβ1 | Integrin beta 1 |
| ITGB4 | Integrin beta 4 |
| J | |
| JAK2 | Janus kinase 2 |
| K | |
| Kupffer cells | liver macrophages; no specific abbreviation |
| L | |
| LSECs | Liver sinusoidal endothelial cells |
| lncRNA | Long non-coding RNA |
| LRP6 | Low-density lipoprotein receptor-related protein 6 |
| LY6E | Lymphocyte antigen 6 family member E |
| M | |
| MASH | Metabolic dysfunction-associated steatohepatitis |
| MASLD | Metabolic dysfunction-associated steatotic liver disease |
| MIF | Macrophage migration inhibitory factor |
| miRNA | MicroRNA |
| MLK3 | Mixed lineage kinase 3 |
| MMP | Matrix metalloproteinases |
| MSCs | Mesenchymal stem cells |
| mTOR | Mammalian target of rapamycin |
| MVBs | Multivesicular bodies |
| N | |
| NAFLD | Nonalcoholic fatty liver disease (inferred) |
| NASH | Nonalcoholic steatohepatitis (inferred) |
| NF-κB | Nuclear factor kappa B |
| NK | Natural killer |
| P | |
| PBC | Primary biliary cholangitis |
| PC | Phosphatidylcholine |
| PDGFRα | Platelet-derived growth factor receptor alpha |
| PD-L1 | Programmed death-ligand 1 |
| PE | Phosphatidylethanolamine |
| PGE2 | Prostaglandin E2 |
| PI3K | Phosphoinositide 3-kinase |
| POU5F1 | POU class 5 homeobox 1 |
| R | |
| ROS | Reactive oxygen species |
| RBBP4 | Rb-binding protein 4 |
| Rb | Retinoblastoma protein |
| S | |
| STAT | Signal transducer and activator of transcription |
| siRNA | Small interfering RNA (inferred) |
| snRNA | Small nuclear RNA (possible) |
| snoRNA | Small nucleolar RNA (possible) |
| S1P | Sphingosine 1-phosphate |
| SK1 | Sphingosine kinase 1 |
| SLC27A5 | Solute carrier family 27 member 5 |
| SENP3–EIF4A1 | SUMO-specific protease 3–eukaryotic initiation factor 4A1 complex |
| T | |
| TAM | Tumor-associated macrophage |
| TME | Tumor microenvironment |
| TNF-α | Tumor necrosis factor alpha |
| TGF-β | Transforming growth factor beta |
| Twist1 | Twist family basic helix-loop-helix transcription factor 1 |
| U | |
| Ub | Ubiquitin (inferred) |
| V | |
| VEGF | Vascular endothelial growth factor |
| VEGFR | Vascular endothelial growth factor receptor |
| VAMP | Vesicle-associated membrane protein |
| Vps4A | Vacuolar protein sorting-associated protein 4A |
| VWF | von Willebrand factor |
| Y | |
| YY1 | Yin yang 1 |
References
- Gan, C.; Yuan, Y.; Shen, H.; Gao, J.; Kong, X.; Che, Z.; Guo, Y.; Wang, H.; Dong, E.; Xiao, J. Liver diseases: Epidemiology, causes, trends and predictions. Signal Transduct. Target. Ther. 2025, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Nagalli, S. Chronic Liver Disease. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2025. [Google Scholar]
- Jiang, Y.; Jiang, L.; Li, F.; Li, Q.; Yuan, S.; Huang, S.; Fu, Y.; Yan, X.; Chen, J.; Li, H.; et al. The epidemiological trends of biliary tract cancers in the United States of America. BMC Gastroenterol. 2022, 22, 546. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Sane, S.; Kim, J.E.; Yun, S.; Kim, H.J.; Jo, K.B.; Wright, J.P.; Khoshdoozmasouleh, N.; Lee, K.; Oh, H.T.; et al. Biogenesis and delivery of extracellular vesicles: Harnessing the power of EVs for diagnostics and therapeutics. Front. Mol. Biosci. 2023, 10, 1330400. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef]
- Kostallari, E.; Valainathan, S.; Biquard, L.; Shah, V.H.; Rautou, P.E. Role of extracellular vesicles in liver diseases and their therapeutic potential. Adv. Drug Deliv. Rev. 2021, 175, 113816. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.F.; Li, W.J.; Hu, K.S.; Gao, J.; Zhai, W.L.; Yang, J.H.; Zhang, S.J. Exosome biogenesis: Machinery, regulation, and therapeutic implications in cancer. Mol. Cancer 2022, 21, 207. [Google Scholar] [CrossRef]
- Tricarico, C.; Clancy, J.; D’Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small GTPases 2017, 8, 220–232. [Google Scholar] [CrossRef]
- Battistelli, M.; Falcieri, E. Apoptotic Bodies: Particular Extracellular Vesicles Involved in Intercellular Communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef]
- Rey-Cadilhac, F.; Rachenne, F.; Missé, D.; Pompon, J. Viral Components Trafficking with(in) Extracellular Vesicles. Viruses 2023, 15, 2333. [Google Scholar] [CrossRef]
- Kakizaki, M.; Yamamoto, Y.; Otsuka, M.; Kitamura, K.; Ito, M.; Kawai, H.D.; Muramatsu, M.; Kagawa, T.; Kotani, A. Extracellular vesicles secreted by HBV-infected cells modulate HBV persistence in hydrodynamic HBV transfection mouse model. J. Biol. Chem. 2020, 295, 12449–12460. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.K.; Jeffrey, G.P.; Ramm, G.A.; Soekmadji, C. Pathogenesis of Viral Hepatitis-Induced Chronic Liver Disease: Role of Extracellular Vesicles. Front. Cell. Infect. Microbiol. 2020, 10, 587628. [Google Scholar] [CrossRef] [PubMed]
- Karamichali, E.; Foka, P.; Papadopoulou, G.; Loukaki-Gkountara, D.; Andresaki, K.; Koskinas, I.; Georgopoulou, U. Hepatitis Viruses Control Host Immune Responses by Modifying the Exosomal Biogenesis Pathway and Cargo. Int. J. Mol. Sci. 2022, 23, 10862. [Google Scholar] [CrossRef] [PubMed]
- Tahir, F.; Farooq, M.; Malik, M.A.; Manzoor, S. Extracellular Vesicles Contribute to Viral-Induced Hepatocellular Carcinoma: Understanding Their Involvement in Viral Hepatitis and Their Potential as Biomarkers for Early Hepatocellular Carcinoma Detection. Viral Immunol. 2024, 37, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Bi, Y.; Huang, Z.; Liao, C.; Li, X.; Hu, H.; Xie, H.; Huang, Y. CD69 Expression is Negatively Associated With T-Cell Immunity and Predicts Antiviral Therapy Response in Chronic Hepatitis B. Ann. Lab. Med. 2025, 45, 185–198. [Google Scholar] [CrossRef]
- Zhang, Q.; Qu, Y.; Zhang, Q.; Li, F.; Li, B.; Li, Z.; Dong, Y.; Lu, L.; Cai, X. Exosomes derived from hepatitis B virus-infected hepatocytes promote liver fibrosis via miR-222/TFRC axis. Cell Biol. Toxicol. 2023, 39, 467–481. [Google Scholar] [CrossRef]
- Yin, Y.; Zhao, Y.; Chen, Q.; Chen, Y.; Mao, L. Dual roles and potential applications of exosomes in HCV infections. Front. Microbiol. 2022, 13, 1044832. [Google Scholar] [CrossRef]
- Lee, C.; Han, J.; Jung, Y. Pathological Contribution of Extracellular Vesicles and Their MicroRNAs to Progression of Chronic Liver Disease. Biology 2022, 11, 637. [Google Scholar] [CrossRef]
- Grossini, E.; Smirne, C.; Venkatesan, S.; Tonello, S.; D’Onghia, D.; Minisini, R.; Cantaluppi, V.; Sainaghi, P.P.; Comi, C.; Tanzi, A.; et al. Plasma Pattern of Extracellular Vesicles Isolated from Hepatitis C Virus Patients and Their Effects on Human Vascular Endothelial Cells. Int. J. Mol. Sci. 2023, 24, 10197. [Google Scholar] [CrossRef]
- Ye, B.; Shen, Y.; Chen, H.; Lin, S.; Mao, W.; Dong, Y.; Li, X. Differential proteomic analysis of plasma-derived exosomes as diagnostic biomarkers for chronic HBV-related liver disease. Sci. Rep. 2022, 12, 14428. [Google Scholar] [CrossRef]
- Cairoli, V.; Valle-Millares, D.; Terrón-Orellano, M.C.; Luque, D.; Ryan, P.; Dominguez, L.; Martín-Carbonero, L.; De Los Santos, I.; De Matteo, E.; Ameigeiras, B.; et al. MicroRNA signature from extracellular vesicles of HCV/HIV co-infected individuals differs from HCV mono-infected. J. Mol. Med. 2023, 101, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Kalligeros, M.; Henry, L. Epidemiology of metabolic dysfunction-associated steatotic liver disease. Clin. Mol. Hepatol. 2025, 31, S32–S50. [Google Scholar] [CrossRef]
- Rodriguez, L.A.; Schmittdiel, J.A.; Liu, L.; Macdonald, B.A.; Balasubramanian, S.; Chai, K.P.; Seo, S.I.; Mukhtar, N.; Levin, T.R.; Saxena, V. Hepatocellular Carcinoma in Metabolic Dysfunction-Associated Steatotic Liver Disease. JAMA Netw. Open 2024, 7, e2421019. [Google Scholar] [CrossRef]
- Sanyal, A.J.; Jha, P.; Kleiner, D.E. Digital pathology for nonalcoholic steatohepatitis assessment. Nat. reviews. Gastroenterol. Hepatol. 2024, 21, 57–69. [Google Scholar] [CrossRef]
- Trifylli, E.M.; Kriebardis, A.G.; Koustas, E.; Papadopoulos, N.; Deutsch, M.; Aloizos, G.; Fortis, S.P.; Papageorgiou, E.G.; Tsagarakis, A.; Manolakopoulos, S. The Emerging Role of Extracellular Vesicles and Autophagy Machinery in NASH-Future Horizons in NASH Management. Int. J. Mol. Sci. 2022, 23, 12185. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Chen, C.; Liu, M. The Role of Extracellular Vesicles in Liver Fibrosis: Friends or Foes? Biomedicines 2024, 12, 2665. [Google Scholar] [CrossRef] [PubMed]
- Newman, L.A.; Muller, K.; Rowland, A. Circulating cell-specific extracellular vesicles as biomarkers for the diagnosis and monitoring of chronic liver diseases. Cell. Mol. Life Sci. CMLS 2022, 79, 232. [Google Scholar] [CrossRef] [PubMed]
- Povero, D.; Eguchi, A.; Li, H.; Johnson, C.D.; Papouchado, B.G.; Wree, A.; Messer, K.; Feldstein, A.E. Circulating extracellular vesicles with specific proteome and liver microRNAs are potential biomarkers for liver injury in experimental fatty liver disease. PLoS ONE 2014, 9, e113651. [Google Scholar] [CrossRef]
- Wang, J.; Bao, S.; An, Q.; Li, C.; Feng, J. Roles of extracellular vesicles from different origins in metabolic-associated fatty liver disease: Progress and perspectives. Front. Immunol. 2025, 16, 1544012. [Google Scholar] [CrossRef]
- Iturbe-Rey, S.; Maccali, C.; Arrese, M.; Aspichueta, P.; Oliveira, C.P.; Castro, R.E.; Lapitz, A.; Izquierdo-Sanchez, L.; Bujanda, L.; Perugorria, M.J.; et al. Lipotoxicity-driven metabolic dysfunction-associated steatotic liver disease (MASLD). Atherosclerosis 2025, 400, 119053. [Google Scholar] [CrossRef]
- Hammerich, L.; Tacke, F. Hepatic inflammatory responses in liver fibrosis. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 633–646. [Google Scholar] [CrossRef]
- Boccatonda, A.; Del Cane, L.; Marola, L.; D’Ardes, D.; Lessiani, G.; di Gregorio, N.; Ferri, C.; Cipollone, F.; Serra, C.; Santilli, F.; et al. Platelet, Antiplatelet Therapy and Metabolic Dysfunction-Associated Steatotic Liver Disease: A Narrative Review. Life 2024, 14, 473. [Google Scholar] [CrossRef]
- Li, J.; Yuan, Y.; Fu, Q.; Chen, M.; Liang, H.; Chen, X.; Long, X.; Zhang, B.; Zhao, J.; Chen, Q. Novel insights into the role of immunomodulatory extracellular vesicles in the pathogenesis of liver fibrosis. Biomark. Res. 2024, 12, 119. [Google Scholar] [CrossRef] [PubMed]
- Mathiesen, A.; Hamilton, T.; Carter, N.; Brown, M.; McPheat, W.; Dobrian, A. Endothelial Extracellular Vesicles: From Keepers of Health to Messengers of Disease. Int. J. Mol. Sci. 2021, 22, 4640. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Buelna, M.; Ramirez-Lopez, I.G.; San Juan-Garcia, C.A.; Garcia-Regalado, J.J.; Millan-Sanchez, M.S.; de la Cruz-Mosso, U.; Haramati, J.; Pereira-Suarez, A.L.; Macias-Barragan, J. Contribution of extracellular vesicles to steatosis-related liver disease and their therapeutic potential. World J. Hepatol. 2024, 16, 1211–1228. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Saldaña, H.A.; Fernández-Garza, L.E.; Barrera-Barrera, S.A. Liquid biopsy in chronic liver disease. Ann. Hepatol. 2021, 20, 100197. [Google Scholar] [CrossRef]
- Balaphas, A.; Meyer, J.; Sadoul, K.; Fontana, P.; Morel, P.; Gonelle-Gispert, C.; Bühler, L.H. Platelets and Platelet-Derived Extracellular Vesicles in Liver Physiology and Disease. Hepatol. Commun. 2019, 3, 855–866. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Tveden-Nyborg, P. Extracellular Vesicles as Drivers of Non-Alcoholic Fatty Liver Disease: Small Particles with Big Impact. Biomedicines 2021, 9, 93. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, M.F.; Jiang, S.; Wu, J.; Liu, J.; Yuan, X.W.; Shen, D.; Zhang, J.Z.; Zhou, N.; He, J.; et al. Liver governs adipose remodelling via extracellular vesicles in response to lipid overload. Nat. Commun. 2020, 11, 719. [Google Scholar] [CrossRef]
- Tomita, K.; Freeman, B.L.; Bronk, S.F.; LeBrasseur, N.K.; White, T.A.; Hirsova, P.; Ibrahim, S.H. CXCL10-Mediates Macrophage, but not Other Innate Immune Cells-Associated Inflammation in Murine Nonalcoholic Steatohepatitis. Sci. Rep. 2016, 6, 28786. [Google Scholar] [CrossRef]
- Srinivas, A.N.; Suresh, D.; Santhekadur, P.K.; Suvarna, D.; Kumar, D.P. Extracellular Vesicles as Inflammatory Drivers in NAFLD. Front. Immunol. 2020, 11, 627424. [Google Scholar] [CrossRef]
- Svobodová, G.; Horní, M.; Velecká, E.; Boušová, I. Metabolic dysfunction-associated steatotic liver disease-induced changes in the antioxidant system: A review. Arch. Toxicol. 2025, 99, 1–22. [Google Scholar] [CrossRef]
- Zhu, C.; Huai, Q.; Zhang, X.; Dai, H.; Li, X.; Wang, H. Insights into the roles and pathomechanisms of ceramide and sphigosine-1-phosphate in nonalcoholic fatty liver disease. Int. J. Biol. Sci. 2023, 19, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Cao, H.X.; Wang, B.C.; Xin, F.Z.; Zhang, R.N.; Zhou, D.; Yang, R.X.; Zhao, Z.H.; Pan, Q.; Fan, J.G. miR-192-5p regulates lipid synthesis in non-alcoholic fatty liver disease through SCD-1. World J. Gastroenterol. 2017, 23, 8140–8151. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Furuta, K.; Lucien, F.; Gutierrez Sanchez, L.H.; Hirsova, P.; Krishnan, A.; Kabashima, A.; Pavelko, K.D.; Madden, B.; Alhuwaish, H.; et al. Integrin β(1)-enriched extracellular vesicles mediate monocyte adhesion and promote liver inflammation in murine NASH. J. Hepatol. 2019, 71, 1193–1205. [Google Scholar] [CrossRef]
- Hochreuter, M.Y.; Dall, M.; Treebak, J.T.; Barrès, R. MicroRNAs in non-alcoholic fatty liver disease: Progress and perspectives. Mol. Metab. 2022, 65, 101581. [Google Scholar] [CrossRef] [PubMed]
- Osawa, Y.; Kawai, H.; Nakashima, K.; Nakaseko, Y.; Suto, D.; Yanagida, K.; Hashidate-Yoshida, T.; Mori, T.; Yoshio, S.; Ohtake, T.; et al. Sphingosine-1-phosphate promotes liver fibrosis in metabolic dysfunction-associated steatohepatitis. PLoS ONE 2024, 19, e0303296. [Google Scholar] [CrossRef]
- Grønbæk, H.; Mellemkjær, A.; Nielsen, S.; Magkos, F. The vascular endothelial growth factor system-a new player in the pathogenesis and development of metabolic dysfunction-associated steatotic liver disease. Hepatobiliary Surg. Nutr. 2023, 12, 963–965. [Google Scholar] [CrossRef]
- Dai, Q.; Ain, Q.; Seth, N.; Rooney, M.; Zipprich, A. Liver sinusoidal endothelial cells: Friend or foe in metabolic dysfunction- associated steatotic liver disease/metabolic dysfunction-associated steatohepatitis. Dig. Liver Dis. 2025, 57, 493–503. [Google Scholar] [CrossRef]
- Povero, D.; Panera, N.; Eguchi, A.; Johnson, C.D.; Papouchado, B.G.; de Araujo Horcel, L.; Pinatel, E.M.; Alisi, A.; Nobili, V.; Feldstein, A.E. Lipid-induced hepatocyte-derived extracellular vesicles regulate hepatic stellate cell via microRNAs targeting PPAR-γ. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 646–663.e644. [Google Scholar] [CrossRef]
- Luo, X.; Luo, S.Z.; Xu, Z.X.; Zhou, C.; Li, Z.H.; Zhou, X.Y.; Xu, M.Y. Lipotoxic hepatocyte-derived exosomal miR-1297 promotes hepatic stellate cell activation through the PTEN signaling pathway in metabolic-associated fatty liver disease. World J. Gastroenterol. 2021, 27, 1419–1434. [Google Scholar] [CrossRef]
- Yang, D.; Liu, J. Targeting extracellular vesicles-mediated hepatic inflammation as a therapeutic strategy in liver diseases. Liver Int. 2020, 40, 2064–2073. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, F.; Cheng, F.; Zhu, L.; Yan, Y. Lipotoxic hepatocyte derived LIMA1 enriched small extracellular vesicles promote hepatic stellate cells activation via inhibiting mitophagy. Cell. Mol. Biol. Lett. 2024, 29, 82. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, Z.; Xia, M.; Salas, S.S.; Ospina, J.A.; Buist-Homan, M.; Harmsen, M.C.; Moshage, H. Extracellular vesicles derived from liver sinusoidal endothelial cells inhibit the activation of hepatic stellate cells and Kupffer cells in vitro. Biochim. Biophys. Acta Mol. Basis Dis. 2024, 1870, 167020. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Hernández, R.; Rojas, Á.; Gato, S.; Gallego, J.; Gil-Gómez, A.; Castro, M.J.; Ampuero, J.; Romero-Gómez, M. Extracellular Vesicles as Biomarkers in Liver Disease. Int. J. Mol. Sci. 2022, 23, 16217. [Google Scholar] [CrossRef]
- Zhang, F.; Xue, M.; Jiang, X.; Yu, H.; Qiu, Y.; Yu, J.; Yang, F.; Bao, Z. Identifying SLC27A5 as a potential prognostic marker of hepatocellular carcinoma by weighted gene co-expression network analysis and in vitro assays. Cancer Cell Int. 2021, 21, 174. [Google Scholar] [CrossRef]
- Ferro, A.; Saccu, G.; Mattivi, S.; Gaido, A.; Herrera Sanchez, M.B.; Haque, S.; Silengo, L.; Altruda, F.; Durazzo, M.; Fagoonee, S. Extracellular Vesicles as Delivery Vehicles for Non-Coding RNAs: Potential Biomarkers for Chronic Liver Diseases. Biomolecules 2024, 14, 277. [Google Scholar] [CrossRef]
- Hu, M.; Huang, H.; Jia, M.; Xu, M.; Chen, M.; Wu, J.; Gu, S.; Liang, H.; Zhou, H.; Gong, Y. A panel of miRNAs in the serum extracellular vesicles serve as novel diagnostic biomarkers for MASLD. Biomed. J. 2025, 100838. [Google Scholar] [CrossRef]
- Erceg, S.; Munjas, J.; Sopić, M.; Tomašević, R.; Mitrović, M.; Kotur-Stevuljević, J.; Mamić, M.; Vujčić, S.; Klisic, A.; Ninić, A. Expression Analysis of Circulating miR-21, miR-34a and miR-122 and Redox Status Markers in Metabolic Dysfunction-Associated Steatotic Liver Disease Patients with and Without Type 2 Diabetes. Int. J. Mol. Sci. 2025, 26, 2392. [Google Scholar] [CrossRef]
- Nie, Z.; Xiao, C.; Wang, Y.; Li, R.; Zhao, F. Heat shock proteins (HSPs) in non-alcoholic fatty liver disease (NAFLD): From molecular mechanisms to therapeutic avenues. Biomark. Res. 2024, 12, 120. [Google Scholar] [CrossRef]
- Welsh, J.A.; Scorletti, E.; Clough, G.F.; Englyst, N.A.; Byrne, C.D. Leukocyte extracellular vesicle concentration is inversely associated with liver fibrosis severity in NAFLD. J. Leukoc. Biol. 2018, 104, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Nakao, Y.; Amrollahi, P.; Parthasarathy, G.; Mauer, A.S.; Sehrawat, T.S.; Vanderboom, P.; Nair, K.S.; Nakao, K.; Allen, A.M.; Hu, T.Y.; et al. Circulating extracellular vesicles are a biomarker for NAFLD resolution and response to weight loss surgery. Nanomed. Nanotechnol. Biol. Med. 2021, 36, 102430. [Google Scholar] [CrossRef] [PubMed]
- Long, M.T.; Massaro, J.M.; Hoffmann, U.; Benjamin, E.J.; Naimi, T.S. Alcohol Use Is Associated With Hepatic Steatosis Among Persons with Presumed Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2020, 18, 1831–1841.e1835. [Google Scholar] [CrossRef]
- Saha, B.; Momen-Heravi, F.; Furi, I.; Kodys, K.; Catalano, D.; Gangopadhyay, A.; Haraszti, R.; Satishchandran, A.; Iracheta-Vellve, A.; Adejumo, A.; et al. Extracellular vesicles from mice with alcoholic liver disease carry a distinct protein cargo and induce macrophage activation through heat shock protein 90. Hepatology 2018, 67, 1986–2000. [Google Scholar] [CrossRef]
- Cho, Y.E.; Song, B.J.; Akbar, M.; Baek, M.C. Extracellular vesicles as potential biomarkers for alcohol- and drug-induced liver injury and their therapeutic applications. Pharmacol. Ther. 2018, 187, 180–194. [Google Scholar] [CrossRef]
- Rahman, M.A.; Patters, B.J.; Kodidela, S.; Kumar, S. Extracellular Vesicles: Intercellular Mediators in Alcohol-Induced Pathologies. J. Neuroimmune Pharmacol. 2020, 15, 409–421. [Google Scholar] [CrossRef]
- Eguchi, A.; Yan, R.; Pan, S.Q.; Wu, R.; Kim, J.; Chen, Y.; Ansong, C.; Smith, R.D.; Tempaku, M.; Ohno-Machado, L.; et al. Comprehensive characterization of hepatocyte-derived extracellular vesicles identifies direct miRNA-based regulation of hepatic stellate cells and DAMP-based hepatic macrophage IL-1β and IL-17 upregulation in alcoholic hepatitis mice. J. Mol. Med. 2020, 98, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.K.; Li, H.; Wang, R.; Hirsova, P.; Mushref, M.; Liu, Y.; Cao, S.; Contreras, P.C.; Malhi, H.; Kamath, P.S.; et al. Alcohol stimulates macrophage activation through caspase-dependent hepatocyte derived release of CD40L containing extracellular vesicles. J. Hepatol. 2016, 64, 651–660. [Google Scholar] [CrossRef]
- Wang, J.; Xia, J.; Huang, R.; Hu, Y.; Fan, J.; Shu, Q.; Xu, J. Mesenchymal stem cell-derived extracellular vesicles alter disease outcomes via endorsement of macrophage polarization. Stem Cell Res. Ther. 2020, 11, 424. [Google Scholar] [CrossRef]
- Babuta, M.; Furi, I.; Bala, S.; Bukong, T.N.; Lowe, P.; Catalano, D.; Calenda, C.; Kodys, K.; Szabo, G. Dysregulated Autophagy and Lysosome Function Are Linked to Exosome Production by Micro-RNA 155 in Alcoholic Liver Disease. Hepatology 2019, 70, 2123–2141. [Google Scholar] [CrossRef]
- Sehrawat, T.S.; Arab, J.P.; Liu, M.; Amrollahi, P.; Wan, M.; Fan, J.; Nakao, Y.; Pose, E.; Navarro-Corcuera, A.; Dasgupta, D.; et al. Circulating Extracellular Vesicles Carrying Sphingolipid Cargo for the Diagnosis and Dynamic Risk Profiling of Alcoholic Hepatitis. Hepatology 2021, 73, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Schnabl, B. Acute alcohol-associated hepatitis: Latest findings in non-invasive biomarkers and treatment. Liver Int. 2025, 45, e15608. [Google Scholar] [CrossRef]
- Fleming, R.E.; Britton, R.S.; Waheed, A.; Sly, W.S.; Bacon, B.R. Pathophysiology of hereditary hemochromatosis. Semin. Liver Dis. 2005, 25, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Pinyopornpanish, K.; Tantiworawit, A.; Leerapun, A.; Soontornpun, A.; Thongsawat, S. Secondary Iron Overload and the Liver: A Comprehensive Review. J. Clin. Transl. Hepatol. 2023, 11, 932–941. [Google Scholar] [CrossRef]
- Wu, J.; Li, Z.; Wu, Y.; Cui, N. The crosstalk between exosomes and ferroptosis: A review. Cell Death Discov. 2024, 10, 170. [Google Scholar] [CrossRef]
- Daou, Y.; Falabrègue, M.; Pourzand, C.; Peyssonnaux, C.; Edeas, M. Host and microbiota derived extracellular vesicles: Crucial players in iron homeostasis. Front. Med. 2022, 9, 985141. [Google Scholar] [CrossRef]
- Qu, H.; Zhou, L.; Wang, J.; Tang, D.; Zhang, Q.; Shi, J. Iron overload is closely associated with metabolic dysfunction-associated fatty liver disease in type 2 diabetes. Obesity 2025, 33, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S.; Kong, Y.; Zheng, H.; Mi, D.; Katabuchi, M.; Motooka, Y.; Ito, F. Double-edged Sword Role of Iron-loaded Ferritin in Extracellular Vesicles. J. Cancer Prev. 2021, 26, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Kim, J.H.; Kim, S.E.; Jung, J.H.; Jang, M.K.; Park, S.H.; Lee, M.S.; Kim, H.S.; Suk, K.T.; Kim, D.J. Primary Biliary Cholangitis and Primary Sclerosing Cholangitis: Current Knowledge of Pathogenesis and Therapeutics. Biomedicines 2022, 10, 1288. [Google Scholar] [CrossRef]
- Trifylli, E.M.; Kriebardis, A.G.; Koustas, E.; Papadopoulos, N.; Vasileiadi, S.; Fortis, S.P.; Tzounakas, V.L.; Anastasiadi, A.T.; Sarantis, P.; Papageorgiou, E.G.; et al. The Arising Role of Extracellular Vesicles in Cholangiocarcinoma: A Rundown of the Current Knowledge Regarding Diagnostic and Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 15563. [Google Scholar] [CrossRef]
- Olaizola, P.; Lee-Law, P.Y.; Arbelaiz, A.; Lapitz, A.; Perugorria, M.J.; Bujanda, L.; Banales, J.M. MicroRNAs and extracellular vesicles in cholangiopathies. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, R.; Wang, Y.; Zhu, W.; Zhao, D.; Wang, X.; Yang, H.; Gurley, E.C.; Chen, W.; Hylemon, P.B.; et al. Cholangiocyte-Derived Exosomal lncRNA H19 Promotes Macrophage Activation and Hepatic Inflammation under Cholestatic Conditions. Cells 2020, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Sucher, E.; Sucher, R.; Gradistanac, T.; Brandacher, G.; Schneeberger, S.; Berg, T. Autoimmune Hepatitis-Immunologically Triggered Liver Pathogenesis-Diagnostic and Therapeutic Strategies. J. Immunol. Res. 2019, 2019, 9437043. [Google Scholar] [CrossRef]
- Yang, F.; Ni, B.; Liang, X.; He, Y.; Yuan, C.; Chu, J.; Huang, Y.; Zhong, H.; Yang, L.; Lu, J.; et al. Mesenchymal stromal cell-derived extracellular vesicles as nanotherapeutics for concanavalin a-induced hepatitis: Modulating the gut-liver axis. Stem Cell Res. Ther. 2025, 16, 4. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, L.; Huang, M.; Wu, Y.; Jin, S.; Zhang, Y.; Gan, X.; Yu, T.; Yu, G.; Zhang, J.; et al. Mesenchymal Stem Cell-Derived Exosomes: Emerging as a Promising Cell-Free Therapeutic Strategy for Autoimmune Hepatitis. Biomolecules 2024, 14, 1353. [Google Scholar] [CrossRef] [PubMed]
- Boccatonda, A.; Piscaglia, F. Predictive role of microvesicles in cirrhotic patients: A promised land or a land of confusion? A narrative review. Ann. Hepatol. 2024, 30, 101563. [Google Scholar] [CrossRef]
- McConnell, M.J.; Kostallari, E.; Ibrahim, S.H.; Iwakiri, Y. The evolving role of liver sinusoidal endothelial cells in liver health and disease. Hepatology 2023, 78, 649–669. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, J.H. The emerging roles of extracellular vesicles as intercellular messengers in liver physiology and pathology. Clin. Mol. Hepatol. 2022, 28, 706–724. [Google Scholar] [CrossRef]
- Iwakiri, Y.; Trebicka, J. Portal hypertension in cirrhosis: Pathophysiological mechanisms and therapy. JHEP Rep. Innov. Hepatol. 2021, 3, 100316. [Google Scholar] [CrossRef]
- Baweja, S.; Kumari, A.; Negi, P.; Thangariyal, S.; Subudhi, P.D.; Gautam, S.; Mittal, A.; Bihari, C. Vascular Extracellular Vesicles Indicate Severe Hepatopulmonary Syndrome in Cirrhosis. Diagnostics 2023, 13, 1272. [Google Scholar] [CrossRef]
- Conti, M.; Minniti, M.; Tiné, M.; De Francesco, M.; Gaeta, R.; Nieri, D.; Semenzato, U.; Biondini, D.; Camera, M.; Cosio, M.G.; et al. Extracellular Vesicles in Pulmonary Hypertension: A Dangerous Liaison? Biology 2023, 12, 1099. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.M.; Bozbas, E.; Tannetta, S.D.; Alroqaiba, N.; Zhou, R.; Crawley, J.T.B.; Gibbins, J.M.; Jones, C.I.; Ahnström, J.; Yaqoob, P. Mode of induction of platelet-derived extracellular vesicles is a critical determinant of their phenotype and function. Sci. Rep. 2020, 10, 18061. [Google Scholar] [CrossRef] [PubMed]
- Leszczynska, A.; Stoess, C.; Sung, H.; Povero, D.; Eguchi, A.; Feldstein, A. Extracellular Vesicles as Therapeutic and Diagnostic Tools for Chronic Liver Diseases. Biomedicines 2023, 11, 2808. [Google Scholar] [CrossRef] [PubMed]
- Severino, V.; Dumonceau, J.M.; Delhaye, M.; Moll, S.; Annessi-Ramseyer, I.; Robin, X.; Frossard, J.L.; Farina, A. Extracellular Vesicles in Bile as Markers of Malignant Biliary Stenoses. Gastroenterology 2017, 153, 495–504.e498. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, M.X.; Yu, M.C.; Ma, Q.W.; Huang, M.J.; Lu, C.W.; Chen, C.B.; Chung, W.H.; Chang, C.J. Altered microbiome of serum exosomes in patients with acute and chronic cholecystitis. BMC Microbiol. 2024, 24, 133. [Google Scholar] [CrossRef] [PubMed]
- Lapitz, A.; Arbelaiz, A.; Olaizola, P.; Aranburu, A.; Bujanda, L.; Perugorria, M.J.; Banales, J.M. Extracellular Vesicles in Hepatobiliary Malignancies. Front. Immunol. 2018, 9, 2270. [Google Scholar] [CrossRef]
- Lin, H.Z.; Zhang, T.; Chen, M.Y.; Shen, J.L. Novel biomarkers for the diagnosis and prognosis of gallbladder cancer. J. Dig. Dis. 2021, 22, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Hong, D.H.; Paudel, S.; Yoon, N.E.; Jung, B.H.; Kim, M.; Kim, T.H.; Jeong, J.; Choi, D.; Lee, H. Metabolomics and miRNA profiling reveals feature of gallbladder cancer-derived biliary extracellular vesicles. Biochem. Biophys. Res. Commun. 2024, 705, 149724. [Google Scholar] [CrossRef]
- Ueta, E.; Tsutsumi, K.; Kato, H.; Matsushita, H.; Shiraha, H.; Fujii, M.; Matsumoto, K.; Horiguchi, S.; Okada, H. Extracellular vesicle-shuttled miRNAs as a diagnostic and prognostic biomarker and their potential roles in gallbladder cancer patients. Sci. Rep. 2021, 11, 12298. [Google Scholar] [CrossRef]
- Priya, R.; Jain, V.; Akhtar, J.; Saklani, N.; Sakhuja, P.; Agarwal, A.K.; Polisetty, R.V.; Sirdeshmukh, R.; Kar, S.; Gautam, P. Proteomic profiling of cell line-derived extracellular vesicles to identify candidate circulatory markers for detection of gallbladder cancer. Front. Oncol. 2022, 12, 1027914. [Google Scholar] [CrossRef]
- Huang, D.-F.; Zhang, W.-J.; Chen, J.; Jiao, Z.-G.; Wang, X.-L.; Rao, D.-Y.; Li, W.-S.; Hu, D.; Xie, F.-F.; Wang, X.-X.; et al. Hepatocellular carcinoma cell-derived small extracellular vesicle-associated CD147 serves as a diagnostic marker and promotes endothelial cell angiogenesis via the PI3K/Akt pathway. Extracell. Vesicles Circ. Nucleic Acids 2023, 4, 532–547. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Zhang, L.; Wang, X.; Wang, Y.; Yu, J.; Li, M.; Ma, Z.; Chi-Lui Ho, P.; Chen, X.; Wang, L.; et al. Extracellular vesicles in the HCC microenvironment: Implications for therapy and biomarkers. Pharmacol. Res. 2024, 209, 107419. [Google Scholar] [CrossRef] [PubMed]
- Koustas, E.; Trifylli, E.M.; Sarantis, P.; Papadopoulos, N.; Papanikolopoulos, K.; Aloizos, G.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Matthaios, D.; et al. An Insight into the Arising Role of MicroRNAs in Hepatocellular Carcinoma: Future Diagnostic and Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 7168. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, X.; Zhang, X.; Shao, T.; Luo, Y.; Wang, W.; Han, Y. Extracellular Vesicles and Hepatocellular Carcinoma: Opportunities and Challenges. Front. Oncol. 2022, 12, 884369. [Google Scholar] [CrossRef]
- Li, J.; Bao, H.; Huang, Z.; Liang, Z.; Wang, M.; Lin, N.; Ni, C.; Xu, Y. Little things with significant impact: miRNAs in hepatocellular carcinoma. Front. Oncol. 2023, 13, 1191070. [Google Scholar] [CrossRef]
- Yang, Y.; Mao, F.; Guo, L.; Shi, J.; Wu, M.; Cheng, S.; Guo, W. Tumor cells derived-extracellular vesicles transfer miR-3129 to promote hepatocellular carcinoma metastasis by targeting TXNIP. Dig. Liver Dis. 2021, 53, 474–485. [Google Scholar] [CrossRef]
- Yamada, N.O.; Heishima, K.; Akao, Y.; Senda, T. Extracellular Vesicles Containing MicroRNA-92a-3p Facilitate Partial Endothelial-Mesenchymal Transition and Angiogenesis in Endothelial Cells. Int. J. Mol. Sci. 2019, 20, 4406. [Google Scholar] [CrossRef]
- Guo, J.; Wang, K.; Sun, Q.; Liu, J.; Zheng, J. Targeting B4GALT3 in BMSCs-EVs for Therapeutic Control of HCC via NF-κB pathway inhibition. Cell Biol. Toxicol. 2025, 41, 67. [Google Scholar] [CrossRef]
- Wang, H.; Lu, Z.; Zhao, X. Tumorigenesis, diagnosis, and therapeutic potential of exosomes in liver cancer. J. Hematol. Oncol. 2019, 12, 133. [Google Scholar] [CrossRef] [PubMed]
- Shoucair, I.; Weber Mello, F.; Jabalee, J.; Maleki, S.; Garnis, C. The Role of Cancer-Associated Fibroblasts and Extracellular Vesicles in Tumorigenesis. Int. J. Mol. Sci. 2020, 21, 6837. [Google Scholar] [CrossRef]
- Hu, W.; Liu, C.; Bi, Z.Y.; Zhou, Q.; Zhang, H.; Li, L.L.; Zhang, J.; Zhu, W.; Song, Y.Y.; Zhang, F.; et al. Comprehensive landscape of extracellular vesicle-derived RNAs in cancer initiation, progression, metastasis and cancer immunology. Mol. Cancer 2020, 19, 102. [Google Scholar] [CrossRef] [PubMed]
- Gondaliya, P.; Sayyed, A.A.; Driscoll, J.; Patel, K.; Patel, T. Extracellular vesicle RNA signaling in the liver tumor microenvironment. Cancer Lett. 2023, 558, 216089. [Google Scholar] [CrossRef]
- Kumar, M.A.; Baba, S.K.; Sadida, H.Q.; Marzooqi, S.A.; Jerobin, J.; Altemani, F.H.; Algehainy, N.; Alanazi, M.A.; Abou-Samra, A.B.; Kumar, R.; et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct. Target. Ther. 2024, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fan, L.; Yu, H.; Zhang, J.; He, Y.; Feng, D.; Wang, F.; Li, X.; Liu, Q.; Li, Y.; et al. Endoplasmic Reticulum Stress Causes Liver Cancer Cells to Release Exosomal miR-23a-3p and Up-regulate Programmed Death Ligand 1 Expression in Macrophages. Hepatology 2019, 70, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Augello, G.; Cusimano, A.; Cervello, M.; Cusimano, A. Extracellular Vesicle-Related Non-Coding RNAs in Hepatocellular Carcinoma: An Overview. Cancers 2024, 16, 1415. [Google Scholar] [CrossRef]
- Wu, Z.; Zeng, Q.; Cao, K.; Sun, Y. Exosomes: Small vesicles with big roles in hepatocellular carcinoma. Oncotarget 2016, 7, 60687–60697. [Google Scholar] [CrossRef]
- Dai, W.; Wang, Y.; Yang, T.; Wang, J.; Wu, W.; Gu, J. Downregulation of exosomal CLEC3B in hepatocellular carcinoma promotes metastasis and angiogenesis via AMPK and VEGF signals. Cell Commun. Signal. CCS 2019, 17, 113. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, X.; Sun, W.; Yue, S.; Yang, J.; Li, J.; Ma, B.; Wang, J.; Yang, X.; Pu, M.; et al. Loss of exosomal miR-320a from cancer-associated fibroblasts contributes to HCC proliferation and metastasis. Cancer Lett. 2017, 397, 33–42. [Google Scholar] [CrossRef]
- Conigliaro, A.; Costa, V.; Lo Dico, A.; Saieva, L.; Buccheri, S.; Dieli, F.; Manno, M.; Raccosta, S.; Mancone, C.; Tripodi, M.; et al. CD90+ liver cancer cells modulate endothelial cell phenotype through the release of exosomes containing H19 lncRNA. Mol. Cancer 2015, 14, 155. [Google Scholar] [CrossRef]
- Wei, J.X.; Lv, L.H.; Wan, Y.L.; Cao, Y.; Li, G.L.; Lin, H.M.; Zhou, R.; Shang, C.Z.; Cao, J.; He, H.; et al. Vps4A functions as a tumor suppressor by regulating the secretion and uptake of exosomal microRNAs in human hepatoma cells. Hepatology 2015, 61, 1284–1294. [Google Scholar] [CrossRef]
- Wang, J.; Pu, J.; Zhang, Y.; Yao, T.; Luo, Z.; Li, W.; Xu, G.; Liu, J.; Wei, W.; Deng, Y. Exosome-transmitted long non-coding RNA SENP3-EIF4A1 suppresses the progression of hepatocellular carcinoma. Aging 2020, 12, 11550–11567. [Google Scholar] [CrossRef] [PubMed]
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Expert consensus document: Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef]
- Zhao, K.; Li, X.; Shi, Y.; Lu, Y.; Qiu, P.; Deng, Z.; Yao, W.; Wang, J. Exosomes in the tumor microenvironment of cholangiocarcinoma: Current status and future perspectives. J. Transl. Med. 2022, 20, 117. [Google Scholar] [CrossRef]
- Oliviero, B.; Dei Cas, M.; Zulueta, A.; Maiello, R.; Villa, A.; Martinelli, C.; Del Favero, E.; Falleni, M.; Montavoci, L.; Varchetta, S.; et al. Ceramide present in cholangiocarcinoma-derived extracellular vesicle induces a pro-inflammatory state in monocytes. Sci. Rep. 2023, 13, 7766. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, G.; Liu, C.G.; Xiang, X.; Le, M.T.N.; Sethi, G.; Wang, L.; Goh, B.C.; Ma, Z. The potential role of exosomal circRNAs in the tumor microenvironment: Insights into cancer diagnosis and therapy. Theranostics 2022, 12, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.; Trifylli, E.M. Role of exosomal circular RNAs as microRNA sponges and potential targeting for suppressing hepatocellular carcinoma growth and progression. World J. Gastroenterol. 2024, 30, 994–998. [Google Scholar] [CrossRef]
- Haga, H.; Yan, I.K.; Takahashi, K.; Wood, J.; Zubair, A.; Patel, T. Tumour cell-derived extracellular vesicles interact with mesenchymal stem cells to modulate the microenvironment and enhance cholangiocarcinoma growth. J. Extracell. Vesicles 2015, 4, 24900. [Google Scholar] [CrossRef]
- Hadad, S.; Khalaji, A.; Sarmadian, A.J.; Sarmadian, P.J.; Janagard, E.M.; Baradaran, B. Tumor-associated macrophages derived exosomes; from pathogenesis to therapeutic opportunities. Int. Immunopharmacol. 2024, 136, 112406. [Google Scholar] [CrossRef]
- Li, L.; Piontek, K.; Ishida, M.; Fausther, M.; Dranoff, J.A.; Fu, R.; Mezey, E.; Gould, S.J.; Fordjour, F.K.; Meltzer, S.J.; et al. Extracellular vesicles carry microRNA-195 to intrahepatic cholangiocarcinoma and improve survival in a rat model. Hepatology 2017, 65, 501–514. [Google Scholar] [CrossRef]
- Momen-Heravi, F.; Balaj, L.; Alian, S.; Mantel, P.Y.; Halleck, A.E.; Trachtenberg, A.J.; Soria, C.E.; Oquin, S.; Bonebreak, C.M.; Saracoglu, E.; et al. Current methods for the isolation of extracellular vesicles. Biol. Chem. 2013, 394, 1253–1262. [Google Scholar] [CrossRef]
- Allelein, S.; Medina-Perez, P.; Lopes, A.L.H.; Rau, S.; Hause, G.; Kölsch, A.; Kuhlmeier, D. Potential and challenges of specifically isolating extracellular vesicles from heterogeneous populations. Sci. Rep. 2021, 11, 11585. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, S.; Jafari, N.; Tamadon, A.; Ghaffarzadeh, A.; Rahbarghazi, R.; Mahdipour, M. Different storage and freezing protocols for extracellular vesicles: A systematic review. Stem Cell Res. Ther. 2024, 15, 453. [Google Scholar] [CrossRef] [PubMed]
- Stawarska, A.; Bamburowicz-Klimkowska, M.; Runden-Pran, E.; Dusinska, M.; Cimpan, M.R.; Rios-Mondragon, I.; Grudzinski, I.P. Extracellular Vesicles as Next-Generation Diagnostics and Advanced Therapy Medicinal Products. Int. J. Mol. Sci. 2024, 25, 6533. [Google Scholar] [CrossRef]
- Poupardin, R.; Wolf, M.; Strunk, D. Adherence to minimal experimental requirements for defining extracellular vesicles and their functions. Adv. Drug Deliv. Rev. 2021, 176, 113872. [Google Scholar] [CrossRef]
- Su, X.; Wang, H.; Li, Q.; Chen, Z. Extracellular Vesicles: A Review of Their Therapeutic Potentials, Sources, Biodistribution, and Administration Routes. Int. J. Nanomed. 2025, 20, 3175–3199. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Biomarker | Description |
|---|---|
| Serum EV-KV311 [21] | Increased levels in chronic hepatitis B versus HCC patients |
| Serum EV-CO9/SVEP1 [21] | Increased levels in chronic hepatitis B compared to healthy controls |
| Serum EV-LBP [21] | Decreased levels in chronic hepatitis B versus HCC patients |
| Serum EV-Willebrand factor [21] | Decreased levels in chronic hepatitis B versus cirrhotic patients |
| Plasma EV-hsa-miR-221/hsa-miR-1290 [22] | Downregulated in HCV-HIV coinfection |
| (a) | |||
|---|---|---|---|
| EV Cargo | Parental Cell | Target Cell(s) | Effect(s) |
| Vanin-1 [19] | Lipotoxic hepatocytes | LSECs, HSCs | Increases angiogenesis (LSECs) and fibrogenesis (HSCs) |
| miR-1 [39] | Lipotoxic hepatocytes | LSECs | Promotes angiogenesis and disease progression |
| let-7e-5p [40] | Lipotoxic hepatocytes | Pre-adipocytes (adipose tissue) | Alters lipid deposition, promotes lipogenesis |
| CXCL10 [41] | Lipotoxic hepatocytes | Monocytes/Macrophages | Enhances chemotaxis, inflammation (inhibited via MLK3) |
| TRAIL [42] | Lipotoxic hepatocytes | Monocytes/Macrophages | Activates macrophages, NF-κB-mediated immune response |
| Mitochondrial DNA (oxidized) [43] | Lipotoxic hepatocytes | Monocytes/Macrophages | Enhances chemotaxis, macrophage recruitment |
| Ceramides [26,44] | Lipotoxic hepatocytes | Monocytes/Macrophages | Enhances chemotaxis, macrophage recruitment |
| miR-192-5p [45] | Lipotoxic hepatocytes | Monocytes/Macrophages | Increases chemotaxis |
| Integrin-β1 [46] | Lipotoxic hepatocytes | Monocytes/Macrophages | Enhances inflammatory response; elevated in MASH with fibrosis |
| miR-128-3p miR-192 [47] | Lipotoxic hepatocytes | HSCs | Activates HSCs, enhances fibrogenesis |
| Sphingosine kinase 1/S1P [48] | Monocytes, HSCs, LSECs | HSCs | Activates HSCs, enhances fibrogenesis |
| VEGF [49] | Portal fibroblasts | Endothelial cells | Enhances angiogenesis |
| Pro-fibrogenic cytokines: TGF-β, CTGF, PDGF [50] | Kupffer, LSECs | HSCs | Enhance fibrosis and angiogenesis, ECM production |
| miR-128-3p [51] | Lipotoxic hepatocytes | HSCs | PPAR-γ inhibition in HSCs, activation of HSCs Increased pro-fibrogenic genes expression (a-SMA, TIMP-2, collagen type I) |
| miR-1297 [52] | Lipotoxic hepatocytes | HSCs | Effect on PTEN/PI3K/AKT signaling in HSCs, HSCs activation, and enhanced fibrogenesis |
| Hedgehog ligands [53] | Activated HSCs | LSECs | LSECs capillarization Vascular remodeling Enhanced fibrogenesis |
| LIMA1 protein [54] | Lipotoxic hepatocytes | HSCs | Mitophagy suppression HSC activation increased COL1A1/A3, α-SMA |
| LSEC-derived EVs [55] | Healthy LSECs | HSCs Kupffer cells | Decrease in HSC/Kupffer cells activation. For instance, EVs from healthy cells |
| (b) | |||
| Biomarker | Description/Relevance | Diagnostic Accuracy (AUC, Sensitivity (%), Specificity (%)) | |
| ITGβ1 [46] | From LPC-treated hepatocytes; promotes macrophage infiltration and inflammation in MASH. Suppressed by antibodies | - | |
| EV-S1P [36,48] | From hepatocytes; involved in fibrogenesis via HSC activation, increased in fibrosis | - | |
| EV-TRAIL [42] | Implicated in MASH; suppression limits disease progression in animal models | - | |
| EV-ASGPR1 [56] | Increased MASLD with advanced fibrosis | 0.83 | |
| EV-SLC27A5 [57] | Elevated in advanced MASLD and MASLD-HCC | - | |
| EV-miR-22 [58] | Levels increase proportionally with MASLD severity | ||
| EV-miR-16 [29] mir-128-3p [47,51,58], mir-192-5p [58], mir-129 [29,58] | Elevated in MASLD/MASH; not liver-specific | - | |
| miR-574-3p, miR-542-3p, miR-200a-3p [59] | Elevated in MASLD patients | - | |
| miR-542-3p, and miR-200a-3p [59] | Elevated in MASLD patients with advanced fibrosis | - | |
| EV-miR-122 [60] | Hepatocyte-specific (70% expression); elevated in advanced MASLD and extended hepatic injury | 0.77 | |
| EV-miR-34a [60] | Non-liver-specific; elevated in MASLD/MASH with fibrotic injury | - | |
| EV-heat shock proteins [61] | Reflect stress-induced hepatocyte injury | - | |
| Leukocyte-derived EVs [62] | Inversely correlated with hepatic fibrosis severity | - | |
| miR-135a-3p miR-122-5p miR-504-3p | Significantly reduced in the serum of MASLD patients vs. healthy controls | miR-135a-3p: 0.849 vs. ALT (0.672) miR-122-5p: 0.790 miR-504-3p: 0.708 | |
| Total circulating EVs (post-operation) [63] | Significantly reduced after bariatric surgery and weight loss | - | |
| miR-21-5p, miR-151a-3p, miR-126-5p | Liver stiffness, steatosis evaluation | 0.76–0.81 0.95 (miR-126-5p + leptin): best for steatosis 0.81 (miR-151a-3p + glucose) | |
| Biomarker | Description/Relevance | Diagnostic Accuracy (AUC; Sensitivity (%); Specificity (%)) |
|---|---|---|
| EV-CYP2E1 [67] | Diagnostic marker of hepatocyte injury in chronic alcohol exposure; linked to ER and oxidative stress, and monocyte toxicity | |
| Plasma EV-miR-19b [66,67] | Elevated in alcohol-associated liver fibrogenesis models | |
| Plasma EV-sphingolipids [72] | Levels correlate with AH severity; serve as prognostic biomarkers | |
| Plasma EV-CK18 (M30, M65) [73] | Diagnostic markers for AH | EV-CK18 M30: 0.75–0.863; 71.5%; 84.6% EV-CK18 M65: 0.82–0.91; 75%; 76% better for inflammation |
| EV-ASGPR1 and EV-CD34+ [56] | Increased corticosteroid non-responders in AH; function as predictive biomarkers for treatment response |
| Topic | Key Component/Process | Function/Mechanism | Outcome/Relevance |
|---|---|---|---|
| EVs in Iron Regulation | Ferritin-containing EVs | EVs carry ferritin in circulation and urine | Non-invasive biomarker of iron status |
| Mitochondria-derived EVs | May deliver ferritin to recipient cells or back to circulation via the multivesicular body–exosome pathway | Potential iron redistribution mechanism | |
| Bone marrow-derived EVs | Modulate hepcidin production | Involved in systemic iron homeostasis | |
| EVs under oxidative stress | Carry antioxidant proteins related to iron metabolism | Mitigate ROS damage; modulate ferroptosis | |
| EV-mediated iron redistribution | EVs sequester excess iron from parental cells | Protect parental cells and may harm recipient cells | |
| EVs in Hepatic Iron Overload | EVs from hepatocytes/macrophages | Altered ferritin and iron-handling enzyme expression | Reflect intracellular iron load; signal local and systemic stress |
| Macrophage-derived EVs | High ferritin content | Indicator of iron overload and inflammation | |
| Hepatocyte-derived EVs in blood/bile | Reflect liver iron load and damage | Useful in MASLD, HH, and other hepatic conditions | |
| Hepatocyte-derived EVs as biomarkers | Reflect liver stress and iron imbalance | Track disease progression in HH and related disorders | |
| EV-iron handling enzymes | Reflect intracellular iron status | EVs can serve as sensitive markers of iron-rich conditions, such as HH | |
| EV-ferritin released by hepatocytes/macrophages | Reflect intracellular iron status | Diagnostic markers of intracellular iron status, non-invasive monitoring biomarkers for HH-related liver injury |
| Condition | EV Component/Origin | Target/Function | Outcome/Associated Effect |
|---|---|---|---|
| PSC [84] | Serum EV-lncRNA H19 | Correlates with PSC severity | Fibrogenesis; disease progression |
| PBC and PSC [82] | Cholangiocyte-derived EVs | Involved in bile duct homeostasis and intercellular crosstalk | Dysregulated in cholangiopathies |
| Hepatocyte-EVs (EGFR and ITGB4) | Biliary tract oncogenesis | Oncogenic signaling | |
| Bile EVs to cholangiocyte cilia | Induce miR-15a → suppress ERK signaling | Inhibit cholangiocyte proliferation | |
| AH [85,86] | MSC-exosomes-miR-21 and miR-16 | Promote pro-inflammatory macrophage phenotype | Can worsen the inflammatory profile |
| MSC-exosomes | Inhibit T-cell proliferation and migration (↓ CCL1, CCL2, CCL21) | Reduced chemotaxis, immune suppression | |
| MSC-EVs | Promote Th1 and Th2 transition | Anti-inflammatory shift | |
| MSC-EVs | PD-L1 expression T-cell function inhibition | Immune suppression |
| Complication | Description | EV-Related Mechanisms |
|---|---|---|
| Fibrotic injury and cirrhosis [50,87,89] | Excessive ECM deposition leading to architectural liver distortion and angiogenesis. | ↑ EV-PDGFRα in circulation; ↓ EV-Twist1 and miR-214 from HSCs; hepatocyte-derived EV-miR-128-3p and miR-192 activate HSCs via PPARγ suppression; LSEC-EV-SK1 promotes fibrosis. |
| PH, hepatopulmonary and porto-pulmonary syndromes [7,90,91,92] | Results from vascular resistance and fibrosis; causes systemic vasodilation and pulmonary complications. | Large EVs involved in vasodilation (↑ in CP B/C); small EVs activate JAK2/ROCK → ↑ resistance; ↑ EV-miR-194 in hepatopulmonary syndrome; EV-VEGF from portal myofibroblasts worsens the condition. |
| Coagulation disorders [19,26,49,93] | Altered coagulation due to liver dysfunction, leading to bleeding or thrombosis. | Platelet-derived EV-annexin V ↑ in severe cirrhosis; EV-tissue factor promotes clotting; HSC, cholangiocyte, and hepatocyte EVs enhance angiogenesis; LSEC-derived EV-VEGF also promotes angiogenesis. |
| Ascites and HE [19,94] | Fluid accumulation and cognitive impairment in advanced liver disease. | ↑ Hepatocyte and endothelial EVs in ascites → associated with mortality; altered EV protein cargoes in HE models; small vesicles in ascites promote inflammation. |
| Biliary tract stenosis and chronic cholecystitis [95,96] | Narrowing of bile ducts and gallbladder inflammation; can progress to malignancy. | Bile-EV levels (e.g., Severino et al.) distinguish malignant vs. benign CBD stenosis (100% accuracy); EVs from microbial-infected cells carry bacteria → dysbiosis and cholecystitis; exosomes modulate gene expression and inflammatory signaling. |
| Hepatobiliary malignancies [97] | Includes HCC, CCA, GBC; often the final stage of chronic hepatobiliary disease. | Multiple EV biomarkers involved (e.g., miRs, lncRNAs, circRNAs, proteins, and lipids); EVs mediate tumor progression, immune evasion, angiogenesis, and chemoresistance. |
| Cancer Type | Biomarker | Description/Relevance | Diagnostic Power AUC; Sensitivity(%); Specificity (%) |
|---|---|---|---|
| GC [82,98,99,100,101] | EV-miR-451a | Decrease in GC; involved in apoptosis and tumor suppression via CDKN2D, MIF, PSMB8. Prognostic marker. | 0.664; 62.0%; 75.0% |
| EV-miR-1246 | Increased GBC; promotes tumor progression, invasion, and proliferation. | 0.646; 60.0%; 66.7% | |
| CEA + CA19-9 + miR-1246 in serum EVs | 0.816; 72.0%; 90.8% | ||
| vs. CEA | 0.770; 60.0%; 83.3% | ||
| CA19-9 | 0.729; 58.0%; 92.6% | ||
| HCC [102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122] | EV-miR-224, -221, -21, -665, -222, -18 | Elevated in HCC; diagnostic markers. | EV-miR-224: 92.5%; 90%; accuracy 94% EV-miR-221: 0.880; 86.5%; 76.7% EV-miR-21: 0.773; 61.1%; 83.3% EV-miR-222 and EV-miR-221: 0.84; 86%; 66% vs. cirrhosis/chronic hepatitis |
| EV-miR-101, -125b | Decreased; miR-125b associated with poor survival and recurrence. | EV-miR-101 0.956; 92.5%; 97.5% EV-miR-125b: 0.739; 83%; 67.9% for recurrence, 0.702; 82.5%; 53.4% survival | |
| EV-hsa_circ_0028861 | Distinguishes HCC vs. CHB | 0.83; 76.79 %; 78.95 % | |
| EV-miR-21-5p + AFP | Elevated in plasma; diagnostic and monitoring value. | 0.85;95%;50% vs. AFP | |
| EV-miR-93 | Poor prognosis; promotes progression via TIMP2, CDKN1A, TP53INP1 targeting. | ||
| EV-LINC00853 | Distinguish AFP (−) early HCC | 0.883 | |
| Distinguish AFP (+) early HCC. | 0.897 | ||
| EV-miR-25 | Associated with resistance to sorafenib therapy. | ||
| EVs-SH3BGRL3, ANGPTL3, IFITM3 | Elevated in viral-HCC; potential early diagnostic biomarkers. | ||
| EV-miR-92b | Elevated post-transplant in recurrent HCC. Predictive biomarker. | ||
| EV-miR-148a | Differentiate HCC vs. cirrhosis. | 0.891 | |
| EV-miR-19–3p | Distinguishing non-hepatitis B, non-hepatitis C-infected HCC. | 0.82 | |
| EV-miR-19-3P ± AFP | 0.92 | ||
| EV-lnc85 | Differentiate HCC vs. cirrhosis. | 0.888 | |
| EV-DANCR | Predicting post-operative recurrence in HCV-infected HCC patients. | 0.88 | |
| CCA [81,82,122,123,124,125,126,127,128,129,130] | EV-FZD10 | Promotes CCA growth/metastasis; a recurrence predictor. | |
| EV-ceramide/dihydroceramide | Correlated with tumor progression and poor prognosis. | ||
| Exosomal membrane lipids (PCs/PEs) | Decreased unsaturated phosphatidylcholines and phosphatidylethanolamines in GC, CCA; associated with loss of membrane integrity. Potential diagnostic marker. | 0.857; 71.4%; 100% (CCA vs. benign) and assay kit phosphatidylcholine: 0.839; 71.4%; 100% |
| Specific EV Components | Mechanism/Pathway | HCC Pathogenesis Role | Mechanism |
|---|---|---|---|
| miR-103, miR-210 [106] | STAT6, SMAD4 overexpression | Altered endothelial integrity | HCC-EV-miR-103, miR-210 affect endothelial cells. |
| miR-3129 [107] | EMT induction | Promotes metastasis | EV-miR-3129 targets TXNIP. |
| MMP-2, MMP-9 [105] | Pro-inflammatory signals | Proliferation, migration | EVs trigger cytokine and MMPs release. |
| let-7b, CD147 [102] | IL-6, MMP-2 upregulation | Inflammation, invasion | Macrophages uptake EV-let-7b and CD147. |
| miR-92a-3p [108] | EMT pathway | Metastasis | EV-miR-92a-3p promotes EMT. |
| miR-1247-3p [109] | Downregulates B4GALT3 | CAF activation, metastasis | EV-miR-1247-3p affects CAFs via B4GALT3. |
| CAF-EVs [109] | Inflammatory cytokines | Lung pre-metastasis niche | CAFs produce IL-6, IL-8, inducing lung niche. |
| circ-PTGR1 [110] | TME homeostasis disruption | Invasion, migration | EV-circ-PTGR1 modifies TME. |
| miR-21 [105,111,112,113] | Growth factor overproduction | CAF transformation | HSCs uptake EV-miR-21. |
| miR-23a [105,112,113] | Adipocyte crosstalk | Proliferation, migration | EV-miR-23a interacts with adipocytes. |
| Adipocyte-EVs [105,112,114] | Deubiquitination | HCC growth promotion | Adipocyte EVs suppress miR-34a. |
| miR-23a-3p, TUC339 [105,112,114] | M2 polarization, PD-L1 ↑ | Immune escape, cytokine release | Macrophages uptake EV-miR-23a-3p, TUC339. |
| miR-221 [112,113,114,117] | Cell cycle regulation | Proliferation | EV-miR-221 targets p27/Kip1. |
| miR-429 [112,113,114,117] | Gene expression control | Stemness, progression | EV-miR-429 affects RBBP4 and POU5F1. |
| circFBLIM1 [112,113,114,117] | Wnt/β-catenin axis | Proliferation | EV-circFBLIM1 involves miR-338/LRP6. |
| FAL1 [112,113,114,117] | ZEB1, AFP expression | Metastasis, tumor marker ↑ | EV-lncRNA FAL1 targets miR-1236. |
| miR-25 [112,113,114,117] | Drug resistance | Therapy resistance | EV-miR-25 mediates sorafenib resistance. |
| CD147, CFH [112,113,114,117] | Fibroblast, inflammation | Tumor progression | EV-CD147 and complement factor H are oncogenic. |
| miR-122 [112,113,114,117] | IGF-1 modulation | Tumor suppression | EV-miR-122 from Huh7 suppresses the tumor. |
| circ-0051443 [112,113,114,117] | Apoptosis promotion | Growth inhibition | EV-circ-0051443 sponges miR-331-3p. |
| VEGF-suppressors [118] | AMPK pathway | Angiogenesis suppression | EV-VEGF-suppressing proteins inhibit angiogenesis. |
| CLEC3B [118] | EGF signaling | Angiogenesis control | EV-CLEC3B suppresses EGF. |
| miR-320a [119] | Inhibits PBX3/ERK/CDK2 | Growth suppression | CAF-derived EV-miR-320a targets PBX3/ERK1/2. |
| H19 [120] | VEGF/VEGFR ↑ | Neovascularization | EV-H19 lncRNA promotes angiogenesis. |
| Vps4A [121] | Tumor signaling suppression | Metastasis inhibition | EV-Vps4A suppresses the PI3K-AKT pathway. |
| SENP3-EIF4A [122] | miR-9-5p suppression | Tumor suppression | Normal liver EVs carry SENP3-EIF4A1. |
| EV Role/Content | Key Concept | Mechanism/Effect |
|---|---|---|
| CCA-derived EVs [81] | Desmoplasia | Promotes fibrotic stroma, ECM remodeling, cytokine, and tumor-promoting molecule production |
| Immune escape | Impairs anti-tumor immune responses, inhibits CIK function (TNF-α, perforin) | |
| EV-integrin α/β, FZD10, vitronectin, lactadherin [124] | Migration/invasion | β-catenin/Wnt pathway activation, enhanced migration, proliferation, metastasis |
| EV-ceramide, dihydroceramide [125] | Distant dissemination | Blood dissemination, monocyte cytokine overproduction |
| EV-circ-CCAC1 [81,82,124] | Neoangiogenesis | Endothelial interaction, YY1 upregulation via miR-514a-5p sponging |
| EV-miR-183-5p [81,82,97,124,125] | Neoangiogenesis | Induces VEGF, PGE2, PTGER1 via mast cells |
| EV-circ-0000284 [128] | Oncogenic transformation | Sponges miR-637, upregulates LY6E |
| HuCCT1-EVs [128] | Tumor microenvironment | Contains CXCL-1, α-SMA, vimentin, FAP, CCL2, IL-6; induces CAFs from MSCs |
| TAM-derived EV-circ-0020256 [129] | Macrophage EVs | Enhances CCA proliferation, migration, and dissemination |
| EV-BMI1 | Immune modulation | Modulates CD8+ T-cell chemotaxis, promotes progression |
| EVmiR-34c, miR-183-5p, miR-200c-3p, miR-200b-3p [124] | Oncogenic EV-miRNAs | Promotes growth, PD-L1 induction, and chemoresistance |
| EV-miR-30e | Tumor suppression | Suppresses EMT, inhibits dissemination |
| HSC-derived EV-miR-195 [130] | Tumor suppression | Inhibits growth and progression in vitro |
| Limitations | Description/Impact |
|---|---|
| Isolation | Lack of standard methods (e.g., ultracentrifugation, immunoaffinity, filtration, size-exclusion); affects purity and reproducibility. |
| Heterogeneity | EVs vary in size, origin, and cargo, complicating function and interpretation. |
| Specificity | Overlapping molecular signatures across diseases can reduce diagnostic specificity. |
| Analytical complexity | Advanced tools needed for analysis are often not available in clinical labs. |
| Storage requirements | Must freeze at −80 °C; avoid repeated freeze–thaw cycles; trehalose may help preserve integrity. |
| Regulatory issues | No unified framework for diagnostic/therapeutic classification; this complicates clinical approval. |
| Ethical considerations | Concerns about donor safety, consent, and traceability—especially for engineered/donor-derived EVs. |
| Contaminants | Risk of co-purifying protein aggregates and lipoproteins, skewing omics analyses. |
| Knowledge gaps | Incomplete understanding of in vivo biodistribution, uptake, and clearance. |
| Manufacturing issues, costs, and resources | Low yield, batch variability, absence of GMP standards hinder scalability, resource-dependent, high costs for manufacturing. |
| Definition inconsistency | Inconsistent EV definitions and reporting methods reduce reproducibility and scientific consensus. |
| Lack of compatibility with other diagnostic modalities | Technological harmonization is required, an algorithmic approach and multi-modal interpretation are needed. |
| Needs for clinical translation | Standardization, mechanistic insights, ethical regulation, and clinical validation. Multi-diseased patients constitute a challenge for EV-profile interpretation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trifylli, E.M.; Fortis, S.P.; Kriebardis, A.G.; Papadopoulos, N.; Koustas, E.; Sarantis, P.; Manolakopoulos, S.; Deutsch, M. Extracellular Vesicles as Biomarkers in Chronic Hepatobiliary Diseases: An Overview of Their Interplay. Int. J. Mol. Sci. 2025, 26, 6333. https://doi.org/10.3390/ijms26136333
Trifylli EM, Fortis SP, Kriebardis AG, Papadopoulos N, Koustas E, Sarantis P, Manolakopoulos S, Deutsch M. Extracellular Vesicles as Biomarkers in Chronic Hepatobiliary Diseases: An Overview of Their Interplay. International Journal of Molecular Sciences. 2025; 26(13):6333. https://doi.org/10.3390/ijms26136333
Chicago/Turabian StyleTrifylli, Eleni Myrto, Sotirios P. Fortis, Anastasios G. Kriebardis, Nikolaos Papadopoulos, Evangelos Koustas, Panagiotis Sarantis, Spilios Manolakopoulos, and Melanie Deutsch. 2025. "Extracellular Vesicles as Biomarkers in Chronic Hepatobiliary Diseases: An Overview of Their Interplay" International Journal of Molecular Sciences 26, no. 13: 6333. https://doi.org/10.3390/ijms26136333
APA StyleTrifylli, E. M., Fortis, S. P., Kriebardis, A. G., Papadopoulos, N., Koustas, E., Sarantis, P., Manolakopoulos, S., & Deutsch, M. (2025). Extracellular Vesicles as Biomarkers in Chronic Hepatobiliary Diseases: An Overview of Their Interplay. International Journal of Molecular Sciences, 26(13), 6333. https://doi.org/10.3390/ijms26136333