
The Therapeutic Effect of GPR81 in Autoimmune Hepatitis and Hepatocellular Carcinoma via Regulating the Immune Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
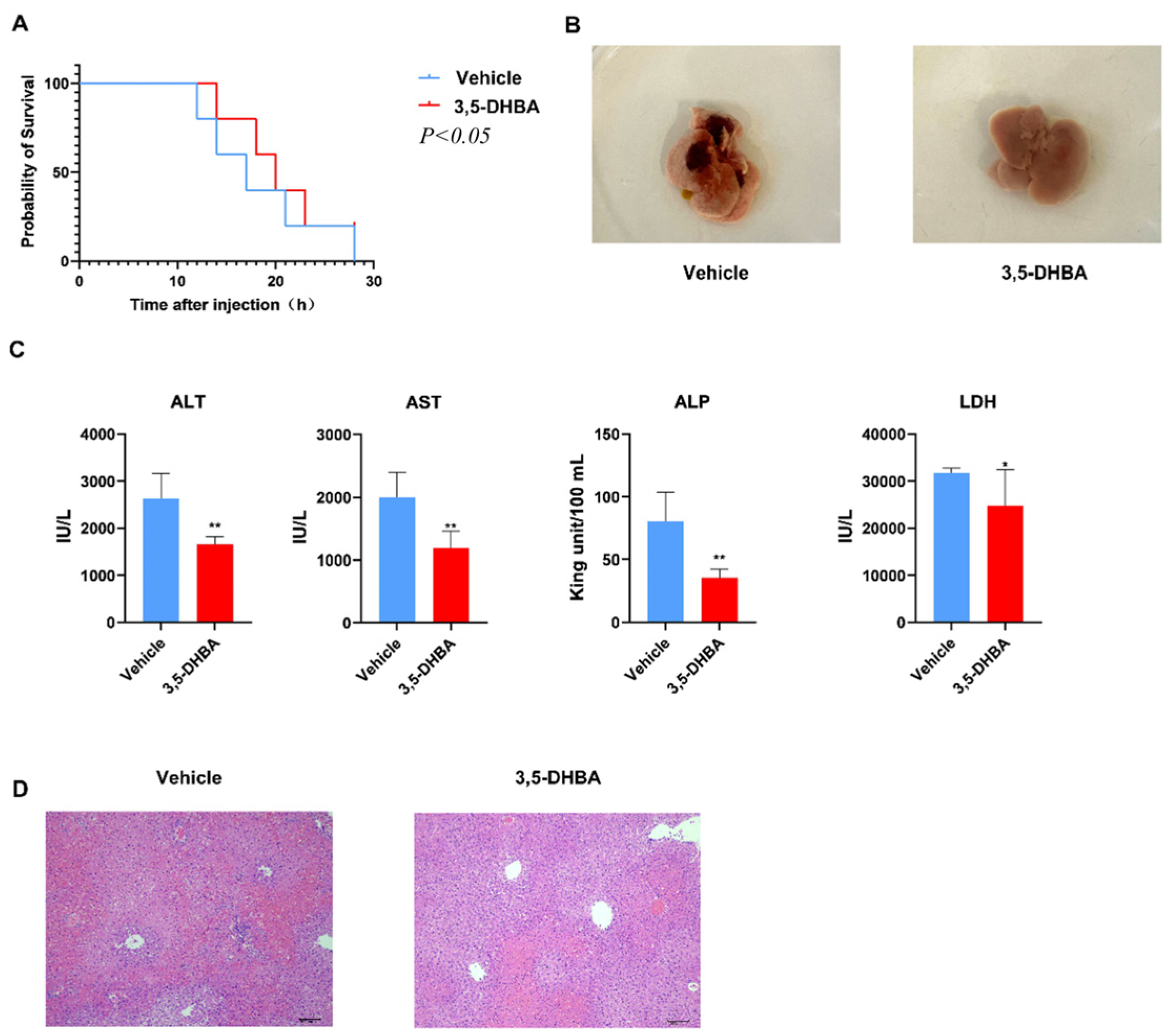
2.1. GPR81 Agonist 3,5-DHBA Effectively Alleviates Con A-Induced AIH
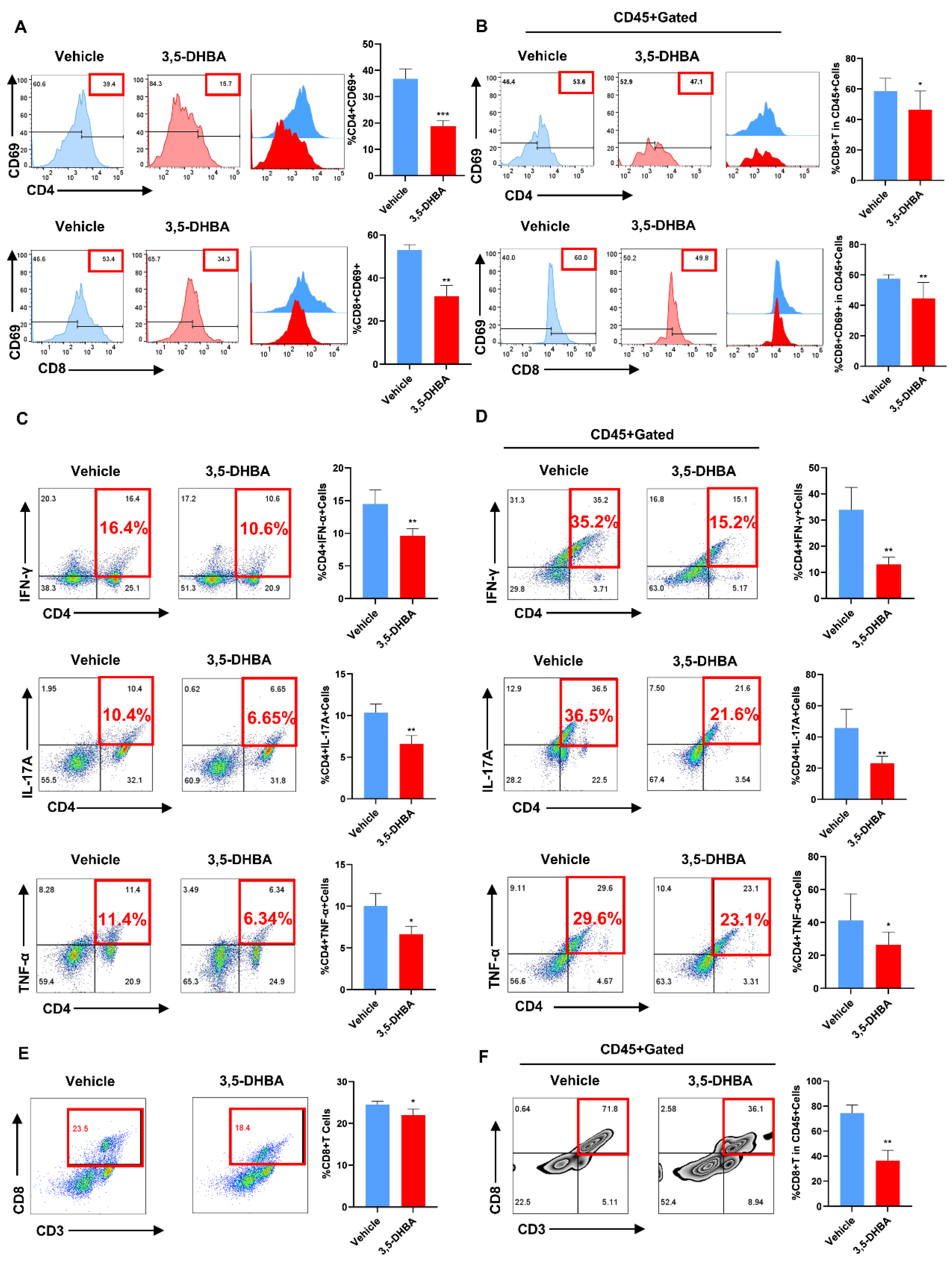
2.2. Pharmacological Activation of GPR81 Inhibits T Cell Activation In Vivo
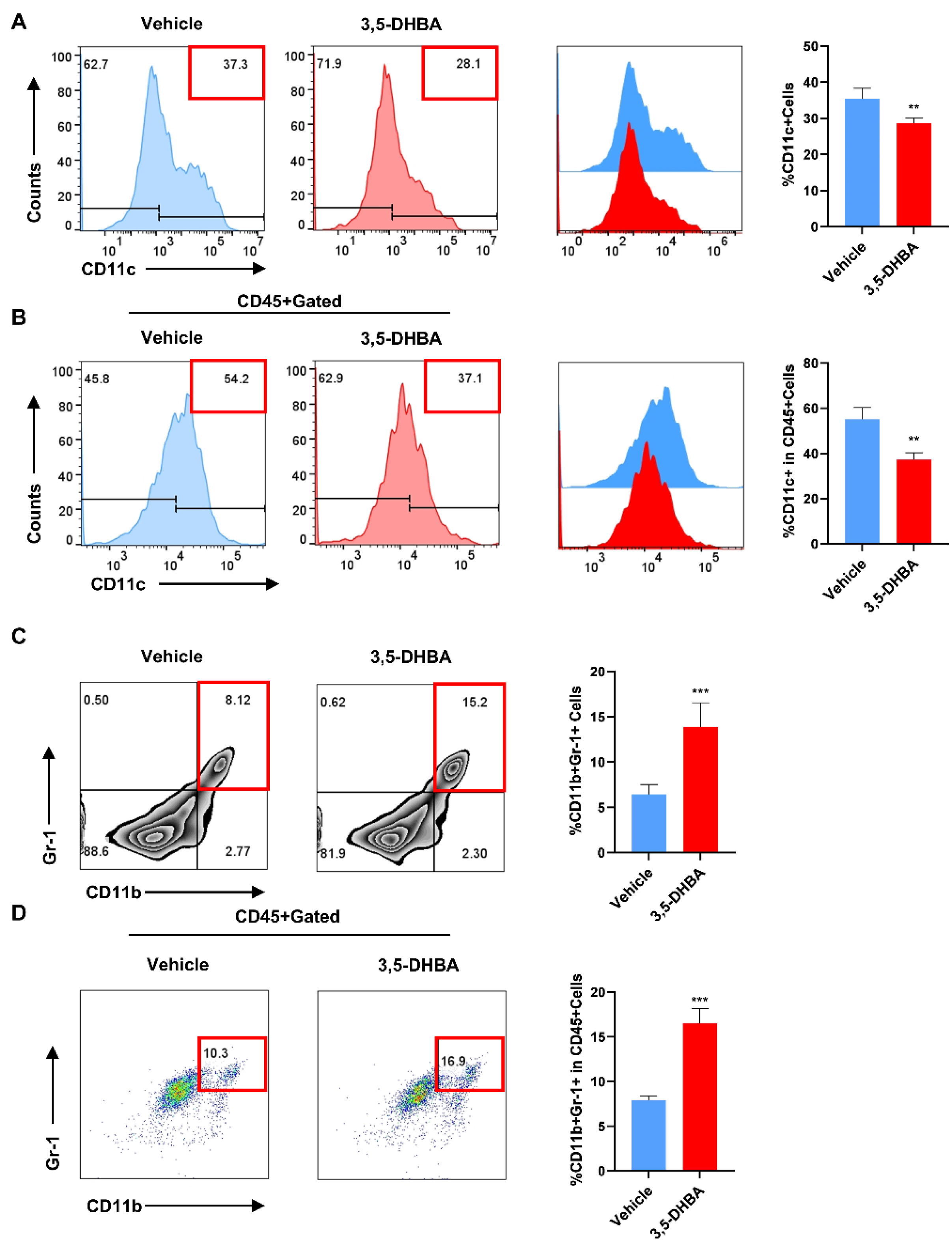
2.3. Pharmacological Activation of GPR81 Reduces the Secretion of DCs and Promotes the Release of MDSCs to Enhance Immune Regulatory Capabilities
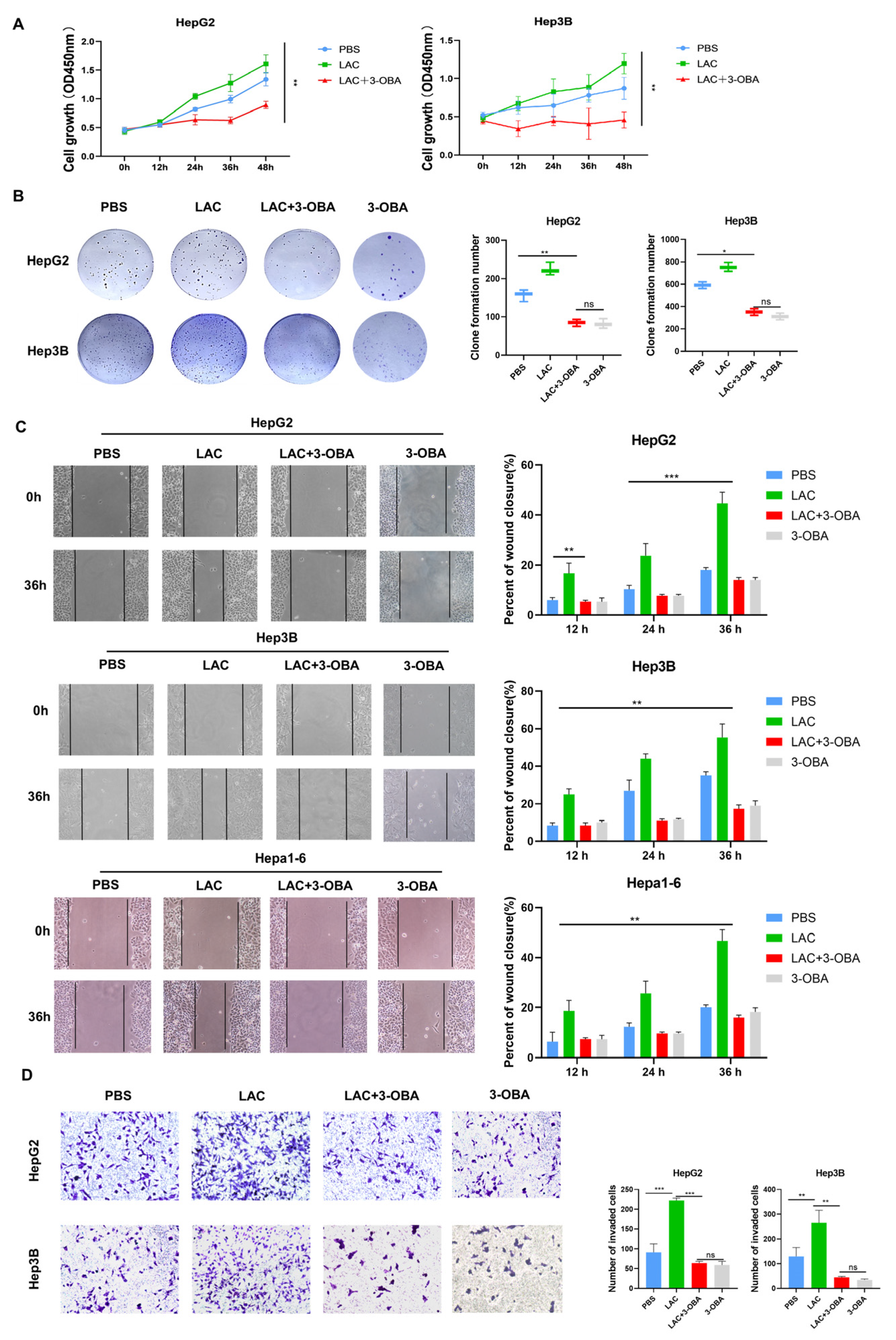
2.4. Inhibition of GPR81 Delays Malignant Progression of Hepatoblastoma Cells
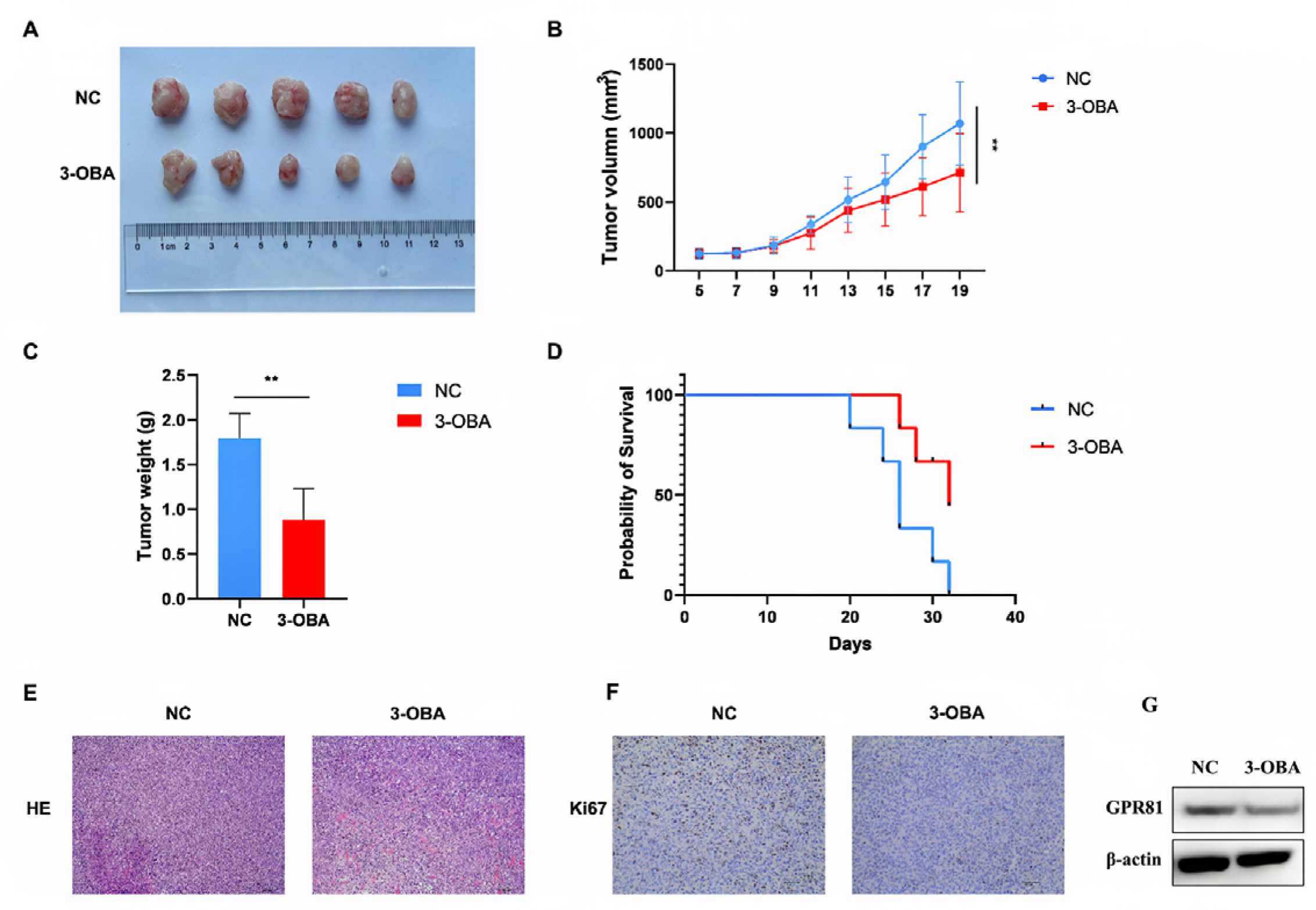
2.5. GPR81 Inhibitors Show Similar Antitumor Effects in Hepa1-6 Cell Bearing Mice
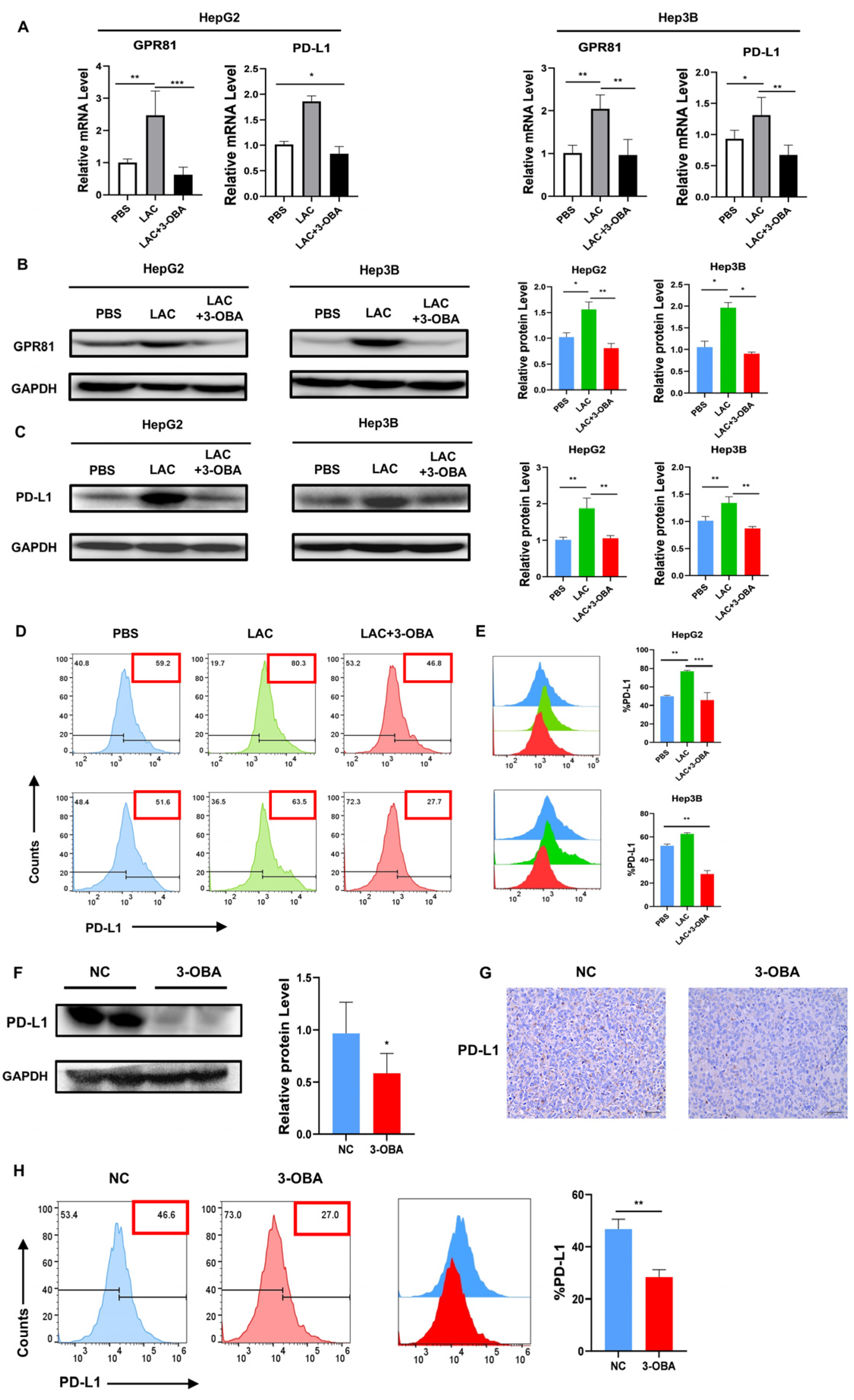
2.6. Inhibition of GPR81 in HepG2 and Hep3B Cells, as Well as in Mouse Tumor Tissues, Leads to a Downregulation of PD-L1 Expression
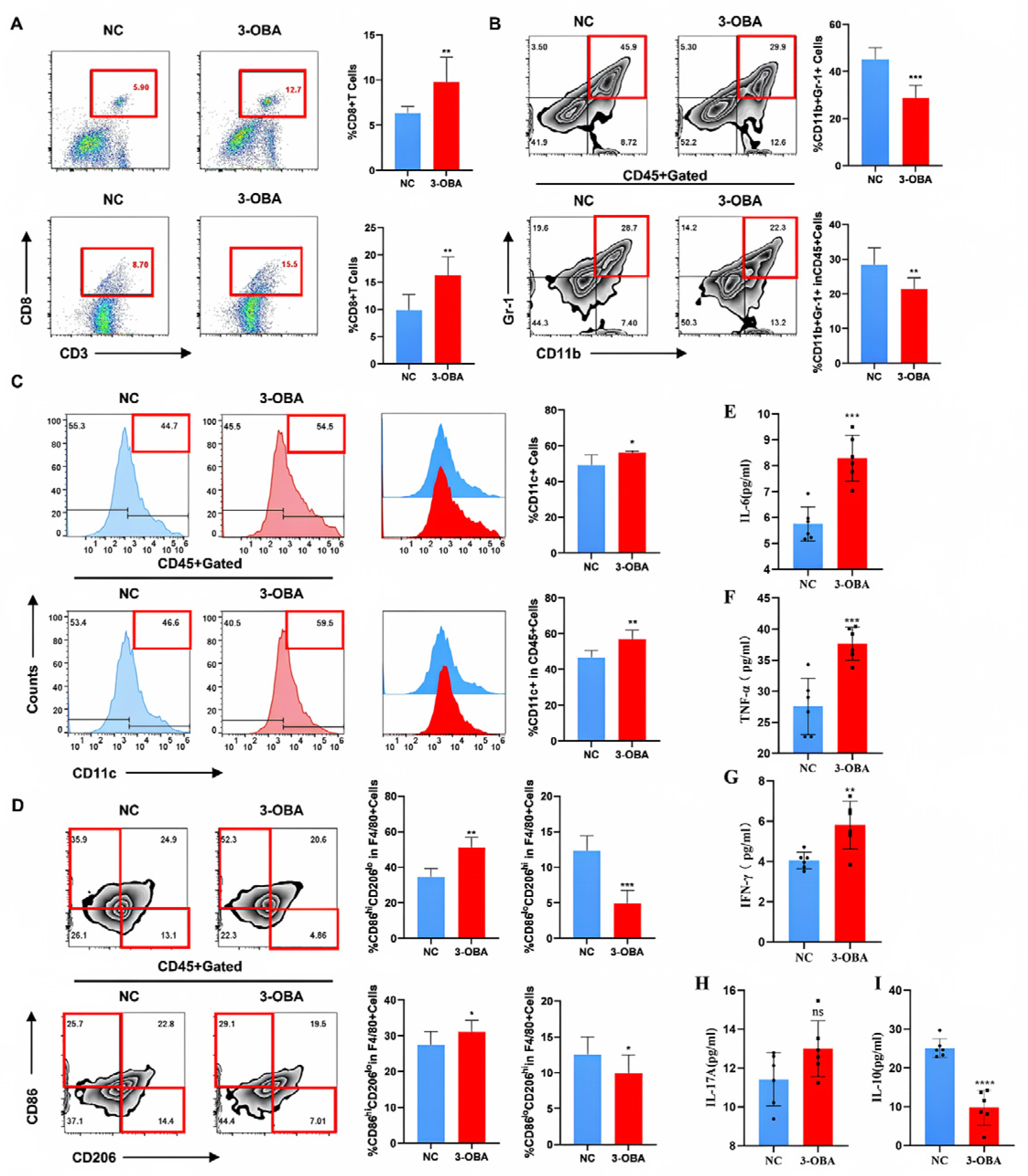
2.7. 3-OBA Remodels the Tumor Immune Microenvironment and Anti-Tumor Immune Response in Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cell Culture and Treatment
4.3. Induction of the AIH
4.4. Xenograft Tumor Growth
4.5. ALT, AST, LDH and ALP Assessment
4.6. Hematoxylin and Eosin (H&E) Staining and Immunohistochemistry
4.7. Flow Cytometry Analysis
4.8. Western Blot Analysis
4.9. RNA Extraction and Real-Time PCR Analysis
| Gene | Forward Primers (5′-3′) | Reverse Primers (5′-3′) |
|---|---|---|
| GPR81 | CCGAGGAGGAACAGCGAAGC | TGATGCCGTAGAGGAGCGATTG |
| PD-L1 | GCCTCCACTCAATGCCTCAATTTG | TTCACAACCACACTCACATGACAAG |
| GAPDH | AATCCCATCACCATCTTCC | TTGAGGCTGTTGTCATACTTCT |
4.10. CCK-8 Assay
4.11. Colony Formation Assay
4.12. Wound Healing Assay
4.13. Transwell Invasion Assay
4.14. Measurement of Cytokines
4.15. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| HCC | Hepatocellular carcinoma |
| AIH | Autoimmune hepatitis |
| 3,5-DHBA | 3,5-Dihydroxybenzoic acid |
| 3-OBA | 3-Hydroxybutyric acid |
| Con A | Concanavalin A |
| LAC | Lactate-treated |
| TME | Tumor microenvironment |
References
- Nault, J.C.; Paradis, V.; Ronot, M.; Zucman-Rossi, J. Benign liver tumours: Understanding molecular physiology to adapt clinical management. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 703–716. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Künzli, M.; Masopust, D. CD4(+) T cell memory. Nat. Immunol. 2023, 24, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Weninger, W.; Manjunath, N.; von Andrian, U.H. Migration and differentiation of CD8+ T cells. Immunol. Rev. 2002, 186, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Chen, S.; Eisenbarth, S.C. Dendritic Cell Regulation of T Helper Cells. Annu. Rev. Immunol. 2021, 39, 759–790. [Google Scholar] [CrossRef] [PubMed]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef]
- Ivashkiv, L.B. IFNγ: Signalling, epigenetics and roles in immunity, metabolism, disease and cancer immunotherapy. Nat. Rev. Immunol. 2018, 18, 545–558. [Google Scholar] [CrossRef]
- Sanjabi, S.; Oh, S.A.; Li, M.O. Regulation of the Immune Response by TGF-β: From Conception to Autoimmunity and Infection. Cold Spring Harb. Perspect. Biol. 2017, 9, a022236. [Google Scholar] [CrossRef]
- Mills, K.H.G. IL-17 and IL-17-producing cells in protection versus pathology. Nat. Rev. Immunol. 2023, 23, 38–54. [Google Scholar] [CrossRef]
- Afonina, I.S.; Cullen, S.P.; Martin, S.J. Cytotoxic and non-cytotoxic roles of the CTL/NK protease granzyme B. Immunol. Rev. 2010, 235, 105–116. [Google Scholar] [CrossRef]
- Richardson, N.; Wootton, G.E.; Bozward, A.G.; Oo, Y.H. Challenges and opportunities in achieving effective regulatory T cell therapy in autoimmune liver disease. Semin. Immunopathol. 2022, 44, 461–474. [Google Scholar] [CrossRef]
- Czaja, A.J.; Manns, M.P. Advances in the diagnosis, pathogenesis, and management of autoimmune hepatitis. Gastroenterology 2010, 139, 58–72.e54. [Google Scholar] [CrossRef] [PubMed]
- Harrington, C.; Krishnan, S.; Mack, C.L.; Cravedi, P.; Assis, D.N.; Levitsky, J. Noninvasive biomarkers for the diagnosis and management of autoimmune hepatitis. Hepatology 2022, 76, 1862–1879. [Google Scholar] [CrossRef] [PubMed]
- Lohse, A.W.; Mieli-Vergani, G. Autoimmune hepatitis. J. Hepatol. 2011, 55, 171–182. [Google Scholar] [CrossRef]
- Ahmed, K. Biological roles and therapeutic potential of hydroxy-carboxylic Acid receptors. Front. Endocrinol. 2011, 2, 51. [Google Scholar] [CrossRef]
- Sun, S.; Li, H.; Chen, J.; Qian, Q. Lactic Acid: No Longer an Inert and End-Product of Glycolysis. Physiology 2017, 32, 453–463. [Google Scholar] [CrossRef]
- Hoque, R.; Farooq, A.; Ghani, A.; Gorelick, F.; Mehal, W.Z. Lactate reduces liver and pancreatic injury in Toll-like receptor- and inflammasome-mediated inflammation via GPR81-mediated suppression of innate immunity. Gastroenterology 2014, 146, 1763–1774. [Google Scholar] [CrossRef]
- Khatib-Massalha, E.; Bhattacharya, S.; Massalha, H.; Biram, A.; Golan, K.; Kollet, O.; Kumari, A.; Avemaria, F.; Petrovich-Kopitman, E.; Gur-Cohen, S.; et al. Lactate released by inflammatory bone marrow neutrophils induces their mobilization via endothelial GPR81 signaling. Nat. Commun. 2020, 11, 3547. [Google Scholar] [CrossRef] [PubMed]
- Madaan, A.; Nadeau-Vallée, M.; Rivera, J.C.; Obari, D.; Hou, X.; Sierra, E.M.; Girard, S.; Olson, D.M.; Chemtob, S. Lactate produced during labor modulates uterine inflammation via GPR81 (HCA(1)). Am. J. Obstet. Gynecol. 2017, 216, 60.e1–60.e17. [Google Scholar] [CrossRef]
- Ranganathan, P.; Shanmugam, A.; Swafford, D.; Suryawanshi, A.; Bhattacharjee, P.; Hussein, M.S.; Koni, P.A.; Prasad, P.D.; Kurago, Z.B.; Thangaraju, M.; et al. GPR81, a Cell-Surface Receptor for Lactate, Regulates Intestinal Homeostasis and Protects Mice from Experimental Colitis. J. Immunol. 2018, 200, 1781–1789. [Google Scholar] [CrossRef]
- Wang, X.; Xie, Z.; Yuan, J.; Jin, E.; Lian, W.; Chang, S.; Sun, G.; Feng, Z.; Xu, H.; Du, C.; et al. Sodium oligomannate disrupts the adherence of Ribhigh bacteria to gut epithelia to block SAA-triggered Th1 inflammation in 5XFAD transgenic mice. Cell Discov. 2024, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, I.; Prasad, P.D.; Thangaraju, M.; Manicassamy, S. Lactate-Dependent Regulation of Immune Responses by Dendritic Cells and Macrophages. Front. Immunol. 2021, 12, 691134. [Google Scholar] [CrossRef]
- Raychaudhuri, D.; Bhattacharya, R.; Sinha, B.P.; Liu, C.S.C.; Ghosh, A.R.; Rahaman, O.; Bandopadhyay, P.; Sarif, J.; D’Rozario, R.; Paul, S.; et al. Lactate Induces Pro-tumor Reprogramming in Intratumoral Plasmacytoid Dendritic Cells. Front. Immunol. 2019, 10, 1878. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Xu, J.; Fan, M.; Tu, F.; Wang, X.; Ha, T.; Williams, D.L.; Li, C. Lactate Suppresses Macrophage Pro-Inflammatory Response to LPS Stimulation by Inhibition of YAP and NF-κB Activation via GPR81-Mediated Signaling. Front. Immunol. 2020, 11, 587913. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Wu, Y.; Li, M.; An, F.; Yao, L.; Wang, M.; Wang, X.; Yuan, J.; Jiang, K.; Li, W.; et al. Lactobacillus spp. create a protective micro-ecological environment through regulating the core fucosylation of vaginal epithelial cells against cervical cancer. Cell Death Dis. 2021, 12, 1094. [Google Scholar] [CrossRef]
- Ishihara, S.; Hata, K.; Hirose, K.; Okui, T.; Toyosawa, S.; Uzawa, N.; Nishimura, R.; Yoneda, T. The lactate sensor GPR81 regulates glycolysis and tumor growth of breast cancer. Sci. Rep. 2022, 12, 6261. [Google Scholar] [CrossRef]
- Xie, Q.; Zhu, Z.; He, Y.; Zhang, Z.; Zhang, Y.; Wang, Y.; Luo, J.; Peng, T.; Cheng, F.; Gao, J.; et al. A lactate-induced Snail/STAT3 pathway drives GPR81 expression in lung cancer cells. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165576. [Google Scholar] [CrossRef]
- Yang, X.; Lu, Y.; Hang, J.; Zhang, J.; Zhang, T.; Huo, Y.; Liu, J.; Lai, S.; Luo, D.; Wang, L.; et al. Lactate-Modulated Immunosuppression of Myeloid-Derived Suppressor Cells Contributes to the Radioresistance of Pancreatic Cancer. Cancer Immunol. Res. 2020, 8, 1440–1451. [Google Scholar] [CrossRef]
- Brown, T.P.; Bhattacharjee, P.; Ramachandran, S.; Sivaprakasam, S.; Ristic, B.; Sikder, M.O.F.; Ganapathy, V. The lactate receptor GPR81 promotes breast cancer growth via a paracrine mechanism involving antigen-presenting cells in the tumor microenvironment. Oncogene 2020, 39, 3292–3304. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.; Yao, X.; Fei, Y.; Lin, Z.; Li, Z.; Cai, K.; Zhao, Y.; Luo, Z. HCAR1/MCT1 Regulates Tumor Ferroptosis through the Lactate-Mediated AMPK-SCD1 Activity and Its Therapeutic Implications. Cell Rep. 2020, 33, 108487. [Google Scholar] [CrossRef]
- Liu, X.; Li, S.; Cui, Q.; Guo, B.; Ding, W.; Liu, J.; Quan, L.; Li, X.; Xie, P.; Jin, L.; et al. Activation of GPR81 by lactate drives tumour-induced cachexia. Nat. Metab. 2024, 6, 708–723. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yang, H.; Zhang, Y.; Wei, H.; Zhu, Z.; Zhu, B.; Yang, M.; Cao, W.; Wang, L.; Wu, Z. Tumor cell-derived lactate induces TAZ-dependent upregulation of PD-L1 through GPR81 in human lung cancer cells. Oncogene 2017, 36, 5829–5839. [Google Scholar] [CrossRef]
- Yang, K.; Fan, M.; Wang, X.; Xu, J.; Wang, Y.; Tu, F.; Gill, P.S.; Ha, T.; Liu, L.; Williams, D.L.; et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ. 2022, 29, 133–146. [Google Scholar] [CrossRef]
- Tarao, K.; Nozaki, A.; Ikeda, T.; Sato, A.; Komatsu, H.; Komatsu, T.; Taguri, M.; Tanaka, K. Real impact of liver cirrhosis on the development of hepatocellular carcinoma in various liver diseases-meta-analytic assessment. Cancer Med. 2019, 8, 1054–1065. [Google Scholar] [CrossRef]
- Mack, C.L.; Adams, D.; Assis, D.N.; Kerkar, N.; Manns, M.P.; Mayo, M.J.; Vierling, J.M.; Alsawas, M.; Murad, M.H.; Czaja, A.J. Diagnosis and Management of Autoimmune Hepatitis in Adults and Children: 2019 Practice Guidance and Guidelines From the American Association for the Study of Liver Diseases. Hepatology 2020, 72, 671–722. [Google Scholar] [CrossRef] [PubMed]
- Hino-Arinaga, T.; Ide, T.; Kuromatsu, R.; Miyajima, I.; Ogata, K.; Kuwahara, R.; Hisamochi, A.; Torimura, T.; Sata, M. Risk factors for hepatocellular carcinoma in Japanese patients with autoimmune hepatitis type 1. J. Gastroenterol. 2012, 47, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Harun-Or-Rashid, M.; Inman, D.M. Reduced AMPK activation and increased HCAR activation drive anti-inflammatory response and neuroprotection in glaucoma. J. Neuroinflammation 2018, 15, 313. [Google Scholar] [CrossRef]
- Roland, C.L.; Arumugam, T.; Deng, D.; Liu, S.H.; Philip, B.; Gomez, S.; Burns, W.R.; Ramachandran, V.; Wang, H.; Cruz-Monserrate, Z.; et al. Cell surface lactate receptor GPR81 is crucial for cancer cell survival. Cancer Res. 2014, 74, 5301–5310. [Google Scholar] [CrossRef]
- Li, X.; Chen, Y.; Wang, T.; Liu, Z.; Yin, G.; Wang, Z.; Sui, C.; Zhu, L.; Chen, W. GPR81-mediated reprogramming of glucose metabolism contributes to the immune landscape in breast cancer. Discov. Oncol. 2023, 14, 140. [Google Scholar] [CrossRef]
- Gantner, F.; Leist, M.; Lohse, A.W.; Germann, P.G.; Tiegs, G. Concanavalin A-induced T-cell-mediated hepatic injury in mice: The role of tumor necrosis factor. Hepatology 1995, 21, 190–198. [Google Scholar]
- Ohta, A.; Sitkovsky, M. Role of G-protein-coupled adenosine receptors in downregulation of inflammation and protection from tissue damage. Nature 2001, 414, 916–920. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Song, W.; Wu, X.; He, J.; Su, M.; Hu, R.; Zhao, Y. The Therapeutic Effect of GPR81 in Autoimmune Hepatitis and Hepatocellular Carcinoma via Regulating the Immune Response. Int. J. Mol. Sci. 2025, 26, 6308. https://doi.org/10.3390/ijms26136308
Wu Y, Song W, Wu X, He J, Su M, Hu R, Zhao Y. The Therapeutic Effect of GPR81 in Autoimmune Hepatitis and Hepatocellular Carcinoma via Regulating the Immune Response. International Journal of Molecular Sciences. 2025; 26(13):6308. https://doi.org/10.3390/ijms26136308
Chicago/Turabian StyleWu, Yongmei, Wenqian Song, Xuxian Wu, Jing He, Min Su, Rong Hu, and Youbo Zhao. 2025. "The Therapeutic Effect of GPR81 in Autoimmune Hepatitis and Hepatocellular Carcinoma via Regulating the Immune Response" International Journal of Molecular Sciences 26, no. 13: 6308. https://doi.org/10.3390/ijms26136308
APA StyleWu, Y., Song, W., Wu, X., He, J., Su, M., Hu, R., & Zhao, Y. (2025). The Therapeutic Effect of GPR81 in Autoimmune Hepatitis and Hepatocellular Carcinoma via Regulating the Immune Response. International Journal of Molecular Sciences, 26(13), 6308. https://doi.org/10.3390/ijms26136308