
Development of Detection Method Using Dried Blood Spot with Next-Generation Sequencing and LabDroid for Gene Doping Control

 , , , , , , and
, , , , , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
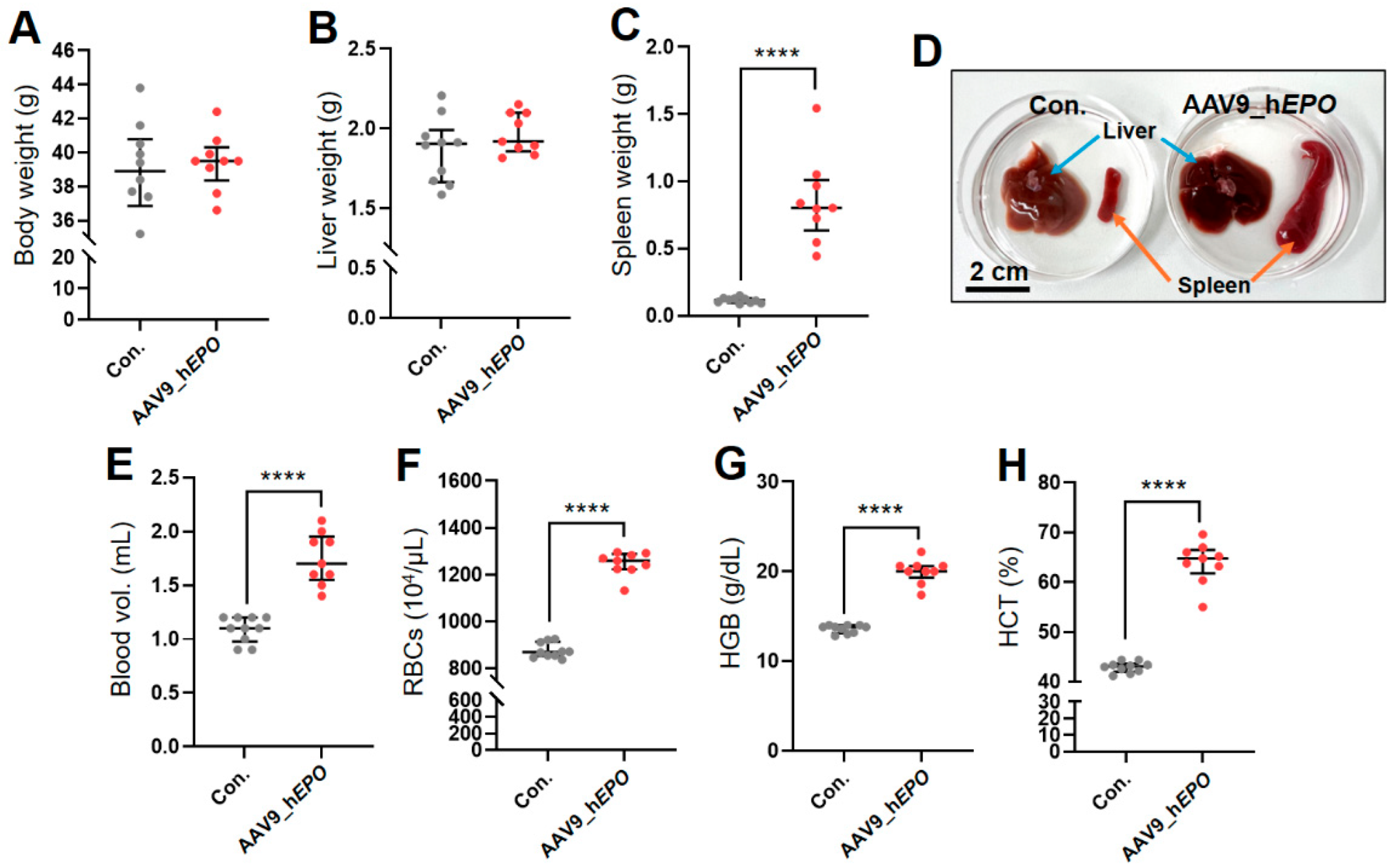
2.1. Creation of a Gene Doping Model Using AAV9_hEPO
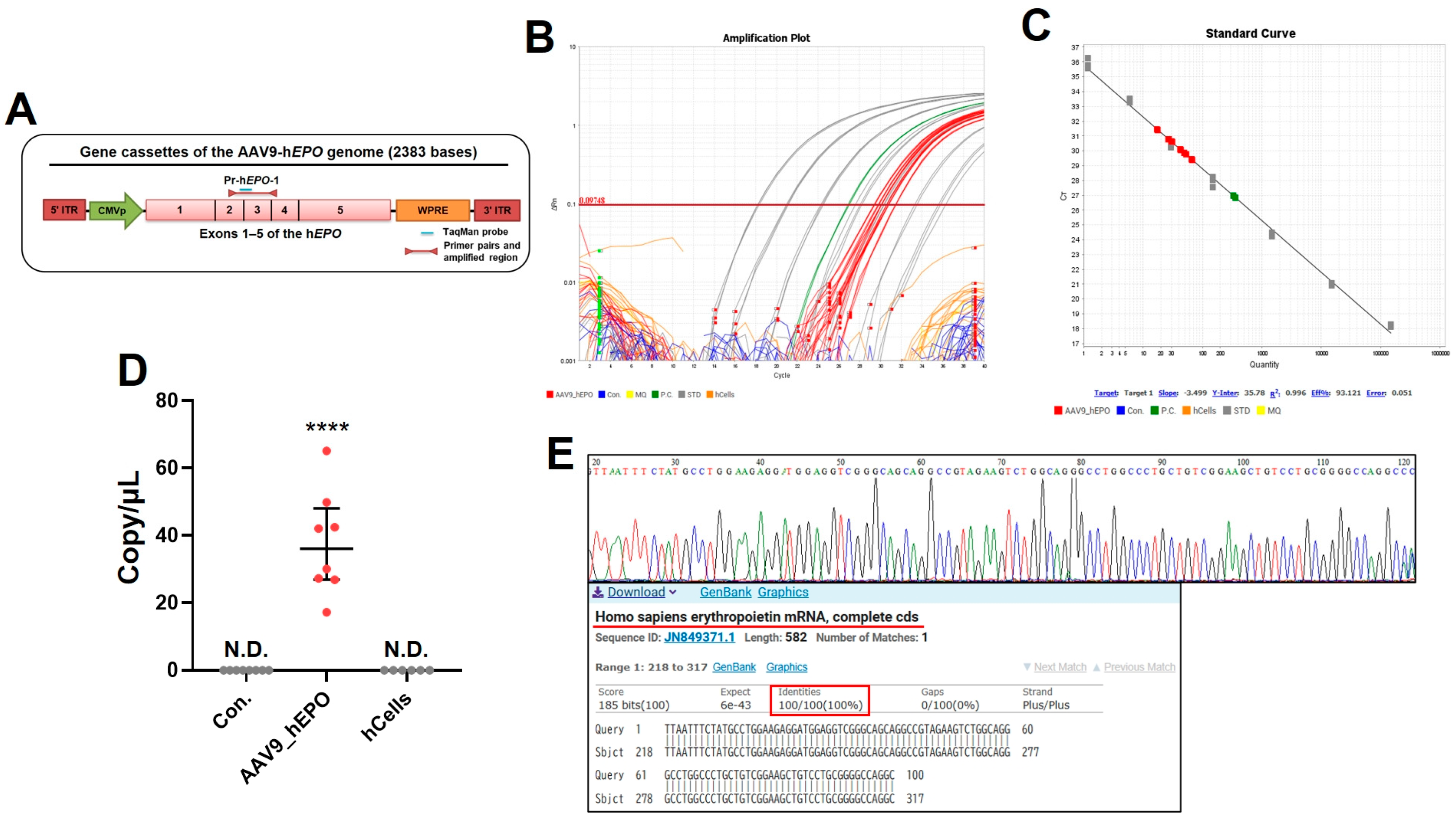
2.2. Gene Doping Was Detected from DBS Using TaqMan qPCR Assay
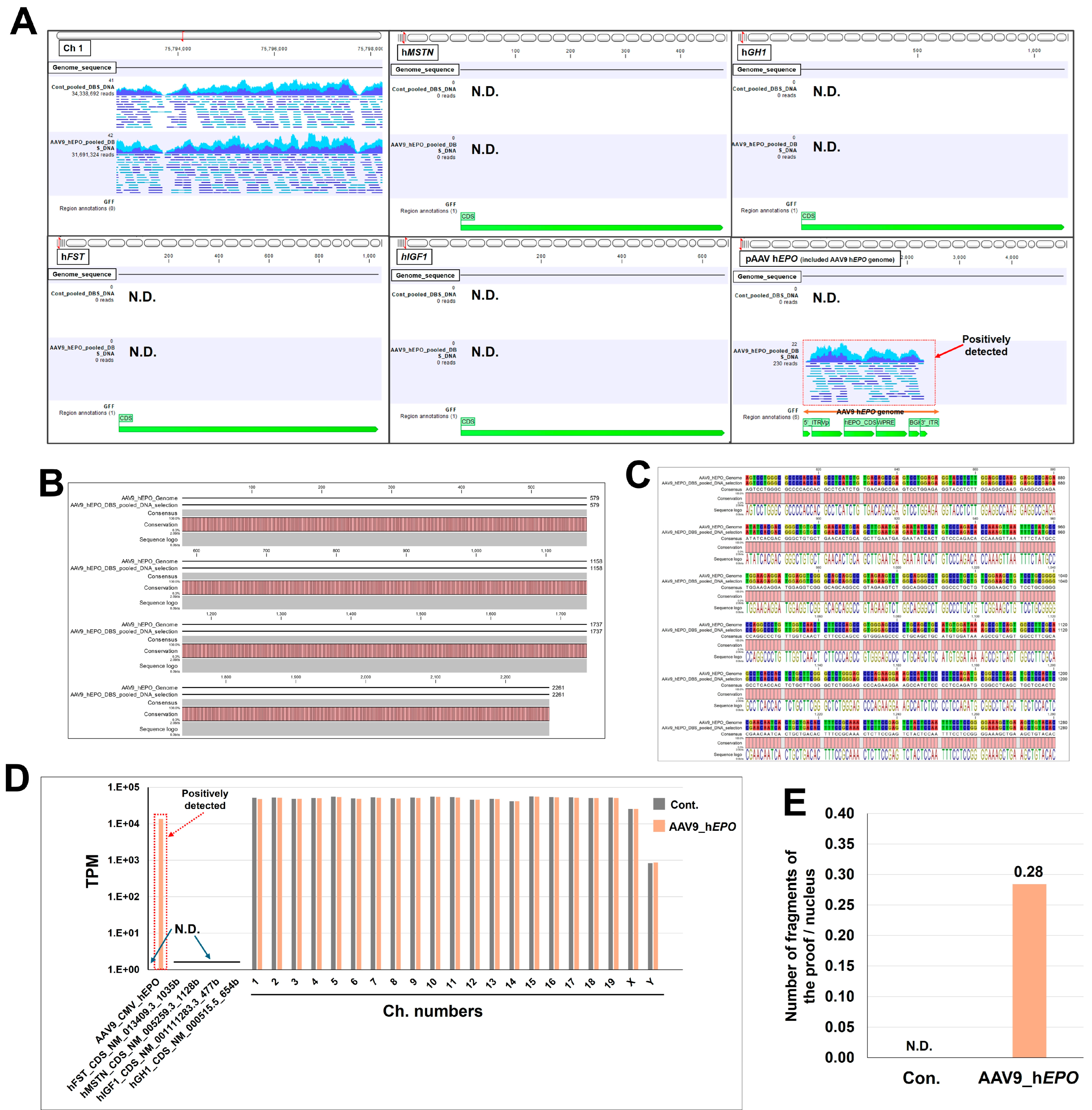
2.3. Whole-Genome Sequencing and Bioinformatic Analysis Can Detect Gene Doping
3. Discussion
4. Materials and Methods
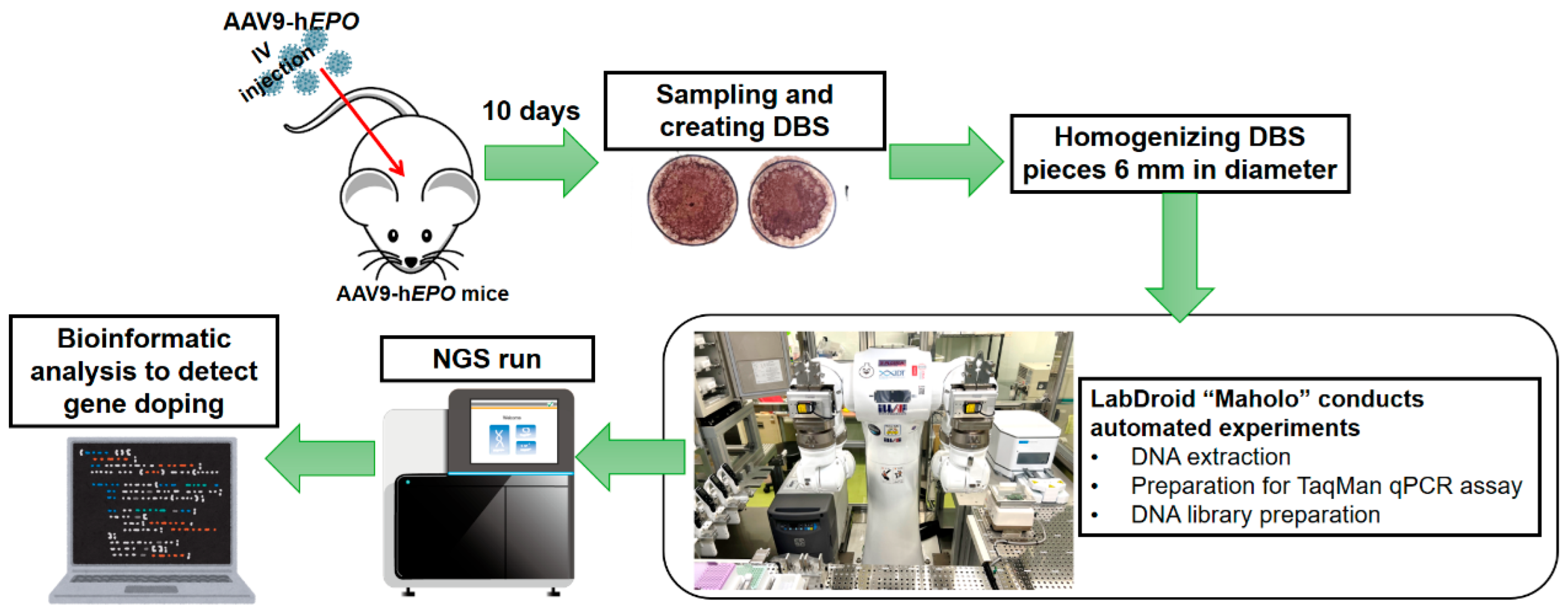
4.1. Graphical Presentation of Experimental Methods
4.2. Creation of the AAV9_hEPO Vector
4.3. Animal Experiments
4.4. Blood Test
4.5. Preparation of DBS and Homogenate
4.6. DNA Extraction and Preparation of TaqMan qPCR Assay Using Maholo
4.7. TaqMan qPCR Assay to Detect Gene Doping
4.8. Sanger Sequencing
4.9. WGS
4.10. Bioinformatic Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- A Japan Anti-Doping Agency (JADA). What Is Anti-Doping? Available online: https://www.playtruejapan.org/about/ (accessed on 7 March 2025).
- The World Anti-Doping Agency (WADA). Who We Are? Available online: https://www.wada-ama.org/en/who-we-are (accessed on 7 March 2025).
- BBC. American Man Jailed After 2020 Tokyo Olympics Doping Charge. Available online: https://www.bbc.com/sport/olympics/68365560 (accessed on 7 March 2025).
- Reuters. Olympics, Bahraini Athletes Provisionally Suspended for Blood Doping Violation. Available online: https://jp.reuters.com/article/life/sports/-idUSKBN2FA03U/ (accessed on 6 April 2025).
- Asahi Shimbun: Ikeda, Tokyo Olympic Silver Medalist in Men’s Race Walking, Provisionally Suspended for Suspected Doping Violation. Available online: https://www.asahi.com/articles/ASSC20J4CSC2UTQP004M.html (accessed on 6 April 2025).
- Division of Molecular Target and Gene Therapy Products, National Institute of Health Sciences. Approved Gene Therapy Products (Updated 27 January 2025). Available online: https://www.nihs.go.jp/mtgt/pdf/section1-1.pdf (accessed on 7 March 2025).
- Li, X.; Le, Y.; Zhang, Z.; Nian, X.; Liu, B.; Yang, X. Viral Vector-Based Gene Therapy. Int. J. Mol. Sci. 2023, 24, 7736. [Google Scholar] [CrossRef] [PubMed]
- The World Anti-Doping Agency (WADA). The World Anti-Doping Code International Standard Prohibited List 2024; WADA: Montreal, QC, Canada, 2024. [Google Scholar]
- The World Anti-Doping Agency (WADA). World Anti-Doping Code 2021; WADA: Montreal, QC, Canada, 2024. [Google Scholar]
- Sugasawa, T.; Nakano, T.; Fujita, S.-I.; Matsumoto, Y.; Ishihara, G.; Aoki, K.; Yanazawa, K.; Ono, S.; Tamai, S.; Manevich, L.; et al. Proof of Gene Doping in a Mouse Model with a Human Erythropoietin Gene Transferred Using an Adenoviral Vector. Genes 2021, 12, 1249. [Google Scholar] [CrossRef] [PubMed]
- Yanazawa, K.; Sugasawa, T.; Aoki, K.; Nakano, T.; Kawakami, Y.; Takekoshi, K. Development of a Gene Doping Detection Method to Detect Overexpressed Human Follistatin Using an Adenovirus Vector in Mice. PeerJ 2021, 9, e12285. [Google Scholar] [CrossRef]
- Sugasawa, T.; Aoki, K.; Yanazawa, K.; Takekoshi, K. Detection of Multiple Transgene Fragments in a Mouse Model of Gene Doping Based on Plasmid Vector Using TaqMan-qPCR Assay. Genes 2020, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Sugasawa, T.; Yanazawa, K.; Watanabe, K.; Takemasa, T.; Takeuchi, Y.; Aita, Y.; Yahagi, N.; Yoshida, Y.; Kuji, T.; et al. The Detection of Trans Gene Fragments of hEPO in Gene Doping Model Mice by Taqman qPCR Assay. PeerJ 2020, 8, e8595. [Google Scholar] [CrossRef]
- Sugasawa, T.; Aoki, K.; Watanabe, K.; Yanazawa, K.; Natsume, T.; Takemasa, T.; Yamaguchi, K.; Takeuchi, Y.; Aita, Y.; Yahagi, N.; et al. Detection of Transgenes in Gene Delivery Model Mice by Adenoviral Vector Using ddPCR. Genes 2019, 10, 436. [Google Scholar] [CrossRef]
- Sugasawa, T.; Hirokawa, A.; Otani, N.; Kanki, Y.; Nguyen, K.D.; Takemasa, T.; Watanabe, K.; Takeuchi, Y.; Yahagi, N.; Takahashi, Y. Detection Method for Gene Doping in a Mouse Model Expressing Human Erythropoietin from Adeno-Associated Virus Vector-9. Genes 2024, 15, 709. [Google Scholar] [CrossRef]
- Heuberger, J.A.A.C.; Cohen Tervaert, J.M.; Schepers, F.M.L.; Vliegenthart, A.D.B.; Rotmans, J.I.; Daniels, J.M.A.; Burggraaf, J.; Cohen, A.F. Erythropoietin doping in cycling: Lack of evidence for efficacy and a negative risk-benefit: Erythropoietin doping in cycling. Br. J. Clin. Pharmacol. 2013, 75, 1406–1421. [Google Scholar] [CrossRef]
- Dimeo, P. Why Lance Armstrong? Historical Context and Key Turning Points in the ‘Cleaning up’ of Professional Cycling. Int. J. Hist. Sport 2014, 31, 951–968. [Google Scholar] [CrossRef]
- Wadler, G.I. The Status of Doping and Drug Use and the Implications for Boxing. Clin. Sports Med. 2009, 28, 533–543. [Google Scholar] [CrossRef]
- Fitch, K. Proscribed Drugs at the Olympic Games: Permitted Use and Misuse (Doping) by Athletes. Clin. Med. 2012, 12, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R. Analytical Study of Doping Cases of Banned Substances during Olympics Games from 1968 to 2012. Int. J. Phys. Educ. Sports Health 2016, 3, 3–37. [Google Scholar]
- Maurya, S.; Sarangi, P.; Jayandharan, G.R. Safety of Adeno-Associated Virus-Based Vector-Mediated Gene Therapy-Impact of Vector Dose. Cancer Gene Ther. 2022, 29, 1305–1306. [Google Scholar] [CrossRef]
- Gene Therapy Clinical Trials Worldwide. Vectors. Available online: https://a873679.fmphost.com/fmi/webd/GTCT (accessed on 7 March 2025).
- The World Anti-Doping Agency (WADA). Highest Ever Proportion of Participating Athletes Tested: The ITA Summarises Its Anti-Doping Program for Paris 2024. Available online: https://ita.sport/news/highest-ever-proportion-of-participating-athletes-tested-the-ita-summarises-its-anti-doping-program-for-paris-2024/ (accessed on 7 March 2025).
- Yachie, N.; Robotic Biology Consortium; Natsume, T. Robotic Crowd Biology with Maholo LabDroids. Nat. Biotechnol. 2017, 35, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Sasamata, M.; Shimojo, D.; Fuse, H.; Nishi, Y.; Sakurai, H.; Nakahata, T.; Yamagishi, Y.; Sasaki-Iwaoka, H. Establishment of a Robust Platform for Induced Pluripotent Stem Cell Research Using Maholo LabDroid. SLAS Technol. 2021, 26, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Terada, M.; Kogawa, Y.; Shibata, Y.; Kitagawa, M.; Kato, S.; Iida, T.; Yorimitsu, T.; Kato, A.; Matsukuma, K.; Maeda, T.; et al. Robotic Cell Processing Facility for Clinical Research of Retinal Cell Therapy. SLAS Technol. 2023, 28, 449–459. [Google Scholar] [CrossRef]
- Kanda, G.N.; Tsuzuki, T.; Terada, M.; Sakai, N.; Motozawa, N.; Masuda, T.; Nishida, M.; Watanabe, C.T.; Higashi, T.; Horiguchi, S.A.; et al. Robotic Search for Optimal Cell Culture in Regenerative Medicine. eLife 2022, 11, e77007. [Google Scholar] [CrossRef]
- Sugasawa, T.; Nguyen, K.D.M.; Otani, N.; Maehara, K.; Kamiya, F.; Hirokawa, A.; Takemasa, T.; Watanabe, K.; Nishi, T.; Sato, K.; et al. Whole Mitochondrial DNA Sequencing Using Fecal Samples from Domestic Dogs. Animals 2024, 14, 2872. [Google Scholar] [CrossRef]
- Sugasawa, T.; Matsumoto, Y.; Fang, H.; Takemasa, T.; Komine, R.; Tamai, S.; Gu, W.; Tanaka, K.; Kanki, Y.; Takahashi, Y. Establishing a Sequencing Method for the Whole Mitochondrial DNA of Domestic Dogs. Animals 2023, 13, 2332. [Google Scholar] [CrossRef]
- Levernæs, M.C.S.; Solheim, S.A.; Broderstad, L.; Zandy, E.; Mørkeberg, J.; Dehnes, Y. Detection of Doping Substances in Paired Dried Blood Spots and Urine Samples Collected during Doping Controls in Danish Fitness Centers. Drug Test. Anal. 2024, 16, 1510–1527. [Google Scholar] [CrossRef]
- Thevis, M.; Walpurgis, K.; Thomas, A. DropWise: Current Role and Future Perspectives of Dried Blood Spots (DBS), Blood Microsampling, and Their Analysis in Sports Drug Testing. Crit. Rev. Clin. Lab. Sci. 2023, 60, 41–62. [Google Scholar] [CrossRef] [PubMed]
- The World Anti-Doping Agency (WADA). Dried Blood Spots (DBS) in Sports Drug Testing: Complementary Matrix Providing Unique Features and Utilities to Modern Anti-Doping Fights. Available online: https://www.wada-ama.org/en/resources/scientific-research/dried-blood-spots-dbs-sports-drug-testing-complementary-matrix (accessed on 6 April 2025).
- The World Anti-Doping Agency (WADA). DRIED BLOOD SPOTS (DBS) FOR DOPING CONTROL, Requirements and Procedures for Collection, Transport, Analytical Testing and Storage; WADA Technical Document–TD2021DBS; WADA: Montreal, QC, Canada, 2021. [Google Scholar]
- Heiland, C.E.; Martin, L.; Zhou, X.; Zhang, L.; Ericsson, M.; Marchand, A. Dried Blood Spots for Erythropoietin Analysis: Detection of Micro-Doses, EPO c.577del Variant and Comparison with in-Competition Matching Urine Samples. Drug Test. Anal. 2024, 16, 650–654. [Google Scholar] [CrossRef]
- Yuan, Y.; Xu, Y.; Lu, J. Dried Blood Spots in Doping Analysis. Bioanalysis 2021, 13, 587–604. [Google Scholar] [CrossRef] [PubMed]
- Marchand, A.; Roulland, I.; Semence, F.; Ericsson, M. EPO Transgene Detection in Dried Blood Spots for Antidoping Application. Drug Test. Anal. 2021, 13, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Colella, P.; Ronzitti, G.; Mingozzi, F. Emerging Issues in AAV-Mediated in Vivo Gene Therapy. Mol. Ther. Methods Clin. Dev. 2018, 8, 87–104. [Google Scholar] [CrossRef]
- Schmidt, M.; Foster, G.R.; Coppens, M.; Thomsen, H.; Dolmetsch, R.; Heijink, L.; Monahan, P.E.; Pipe, S.W. Molecular Evaluation and Vector Integration Analysis of HCC Complicating AAV Gene Therapy for Hemophilia B. Blood Adv. 2023, 7, 4966–4969. [Google Scholar] [CrossRef]
- Narayanasamy, S.; Markina, V.; Thorogood, A.; Blazkova, A.; Shabani, M.; Knoppers, B.M.; Prainsack, B.; Koesters, R. Genomic Sequencing Capacity, Data Retention, and Personal Access to Raw Data in Europe. Front. Genet. 2020, 11, 303. [Google Scholar] [CrossRef]
- Puchalska, M.; Witkowska-Piłaszewicz, O. Gene Doping in Horse Racing and Equine Sports: Current Landscape and Future Perspectives. Equine Vet. J. 2025, 57, 312–324. [Google Scholar] [CrossRef]
- Tozaki, T.; Ohnuma, A.; Kikuchi, M.; Ishige, T.; Hirota, K.-I.; Kakoi, H.; Kusano, K.; Nagata, S.-I. Gene Doping Control in Horseracing Industries. Dobutsu Iden Ikushu Kenkyu 2021, 49, 19–29. [Google Scholar] [CrossRef]
- Tozaki, T.; Hamilton, N.A. Control of Gene Doping in Human and Horse Sports. Gene Ther. 2022, 29, 107–112. [Google Scholar] [CrossRef]
- Ohnuma, A.; Tozaki, T.; Kikuchi, M.; Ishige, T.; Kakoi, H.; Hirota, K.-I.; Takahashi, Y.; Nagata, S.-I. Multiplex Detection of Transgenes Using πCode Technology for Gene Doping Control. Anal. Chem. 2023, 95, 10149–10154. [Google Scholar] [CrossRef] [PubMed]
- Tozaki, T.; Ohnuma, A.; Takasu, M.; Kikuchi, M.; Kakoi, H.; Hirota, K.-I.; Kusano, K.; Nagata, S.-I. Droplet Digital PCR Detection of the Erythropoietin Transgene from Horse Plasma and Urine for Gene-Doping Control. Genes 2019, 10, 243. [Google Scholar] [CrossRef]
- Tozaki, T.; Gamo, S.; Takasu, M.; Kikuchi, M.; Kakoi, H.; Hirota, K.-I.; Kusano, K.; Nagata, S.-I. Digital PCR Detection of Plasmid DNA Administered to the Skeletal Muscle of a Microminipig: A Model Case Study for Gene Doping Detection. BMC Res. Notes 2018, 11, 708. [Google Scholar] [CrossRef]
- Tozaki, T.; Ohnuma, A.; Kikuchi, M.; Ishige, T.; Kakoi, H.; Hirota, K.-I.; Kusano, K.; Nagata, S.-I. Microfluidic Quantitative PCR Detection of 12 Transgenes from Horse Plasma for Gene Doping Control. Genes 2020, 11, 457. [Google Scholar] [CrossRef] [PubMed]
- Tozaki, T.; Ohnuma, A.; Kikuchi, M.; Ishige, T.; Kakoi, H.; Hirota, K.-I.; Hamilton, N.A.; Kusano, K.; Nagata, S.-I. Whole-Genome Resequencing Using Genomic DNA Extracted from Horsehair Roots for Gene-Doping Control in Horse Sports. J. Equine Sci. 2020, 31, 75–83. [Google Scholar] [CrossRef]
- Furukawa, R.; Tozaki, T.; Kikuchi, M.; Ishige, T.; Takahashi, Y.; Fukui, E.; Kakoi, H. A Method for Detecting Gene Doping in Horse Sports without DNA Extraction. Drug Test. Anal. 2024, 17, 427–435. [Google Scholar] [CrossRef]
- Rausch, T.; Zichner, T.; Schlattl, A.; Stütz, A.M.; Benes, V.; Korbel, J.O. DELLY: Structural Variant Discovery by Integrated Paired-End and Split-Read Analysis. Bioinformatics 2012, 28, i333–i339. [Google Scholar] [CrossRef] [PubMed]
- Tozaki, T.; Ohnuma, A.; Kikuchi, M.; Ishige, T.; Kakoi, H.; Hirota, K.; Kusano, K.; Nagata, S. Simulated Validation of Intron-Less Transgene Detection Using DELLY for Gene-Doping Control in Horse Sports. Anim. Genet. 2021, 52, 759–761. [Google Scholar] [CrossRef]
- Tozaki, T.; Ohnuma, A.; Kikuchi, M.; Ishige, T.; Kakoi, H.; Hirota, K.-I.; Kusano, K.; Nagata, S.-I. Identification of Processed Pseudogenes in the Genome of Thoroughbred Horses: Possibility of Gene-Doping Detection Considering the Presence of Pseudogenes. Anim. Genet. 2022, 53, 183–192. [Google Scholar] [CrossRef]
- Tozaki, T.; Ohnuma, A.; Takasu, M.; Nakamura, K.; Kikuchi, M.; Ishige, T.; Kakoi, H.; Hirora, K.-I.; Tamura, N.; Kusano, K.; et al. Detection of Non-Targeted Transgenes by Whole-Genome Resequencing for Gene-Doping Control. Gene Ther. 2021, 28, 199–205. [Google Scholar] [CrossRef]
- Otani, N.; Kanki, Y.; Nguyen, K.D.M.; Sugasawa, T. Detection of Gene Doping Using Dried Blood Spots from a Mouse Model with rAAV9 Vector-Mediated Human Erythropoietin Expression as a Pilot Study. Analytica 2024, 5, 263–272. [Google Scholar] [CrossRef]
- Vector Builder. The Gene Delivery Revolution. Available online: https://www.vectorbuilder.jp (accessed on 6 April 2025).
- The World Anti-Doping Agency (WADA). Gene Doping Detection based on Polymerase Chain Reaction (PCR); Laboratory Guidelines; WADA: Montreal, QC, Canada, 2021. [Google Scholar]
- Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 6 April 2025).
- Mus Musculus Genome Assembly GRCm39. Available online: https://www.ncbi.nlm.nih.gov/datasets/genome/GCF_000001635.27/ (accessed on 6 April 2025).
- Homo Sapiens Myostatin (MSTN), mRNA–Nucleotide–NCBI. Available online: https://www.ncbi.nlm.nih.gov/nuccore/NM_005259.3 (accessed on 6 April 2025).
- Homo Sapiens Growth Hormone 1 (GH1), Transcript Variant 1, mRNA–Nucleotide–NCBI. Available online: https://www.ncbi.nlm.nih.gov/nuccore/NM_000515.5 (accessed on 6 April 2025).
- Homo Sapiens Follistatin (FST), Transcript Variant FST344, mRNA–Nucleotide–NCBI. Available online: https://www.ncbi.nlm.nih.gov/nuccore/NM_013409 (accessed on 6 April 2025).
- Homo Sapiens Insulin Like Growth Factor 1 (IGF1), Transcript Variant 1–Nucleotide–NCBI. Available online: https://www.ncbi.nlm.nih.gov/nuccore/NM_001111283.3 (accessed on 6 April 2025).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maehara, K.; Hirokawa, A.; Watanabe, H.; Otani, N.; Wan, J.; Shirai, T.; Takemasa, T.; Watanabe, K.; Nishi, T.; Sato, K.; et al. Development of Detection Method Using Dried Blood Spot with Next-Generation Sequencing and LabDroid for Gene Doping Control. Int. J. Mol. Sci. 2025, 26, 6129. https://doi.org/10.3390/ijms26136129
Maehara K, Hirokawa A, Watanabe H, Otani N, Wan J, Shirai T, Takemasa T, Watanabe K, Nishi T, Sato K, et al. Development of Detection Method Using Dried Blood Spot with Next-Generation Sequencing and LabDroid for Gene Doping Control. International Journal of Molecular Sciences. 2025; 26(13):6129. https://doi.org/10.3390/ijms26136129
Chicago/Turabian StyleMaehara, Kiyoshi, Atsushi Hirokawa, Hinata Watanabe, Norihiro Otani, Jiawei Wan, Takanaga Shirai, Tohru Takemasa, Koichi Watanabe, Takeki Nishi, Ken Sato, and et al. 2025. "Development of Detection Method Using Dried Blood Spot with Next-Generation Sequencing and LabDroid for Gene Doping Control" International Journal of Molecular Sciences 26, no. 13: 6129. https://doi.org/10.3390/ijms26136129
APA StyleMaehara, K., Hirokawa, A., Watanabe, H., Otani, N., Wan, J., Shirai, T., Takemasa, T., Watanabe, K., Nishi, T., Sato, K., Shimmura, S., Nguyen, K. D. M., Takahashi, Y., & Sugasawa, T. (2025). Development of Detection Method Using Dried Blood Spot with Next-Generation Sequencing and LabDroid for Gene Doping Control. International Journal of Molecular Sciences, 26(13), 6129. https://doi.org/10.3390/ijms26136129