
Preclinical Evaluation of the Systemic Safety, Efficacy, and Biodistribution of a Recombinant AAV8 Vector Expressing FIX-TripleL in Hemophilia B Mice: Implications for Human Gene Therapy
 , , , and
, , , and
Abstract
1. Introduction
2. Results
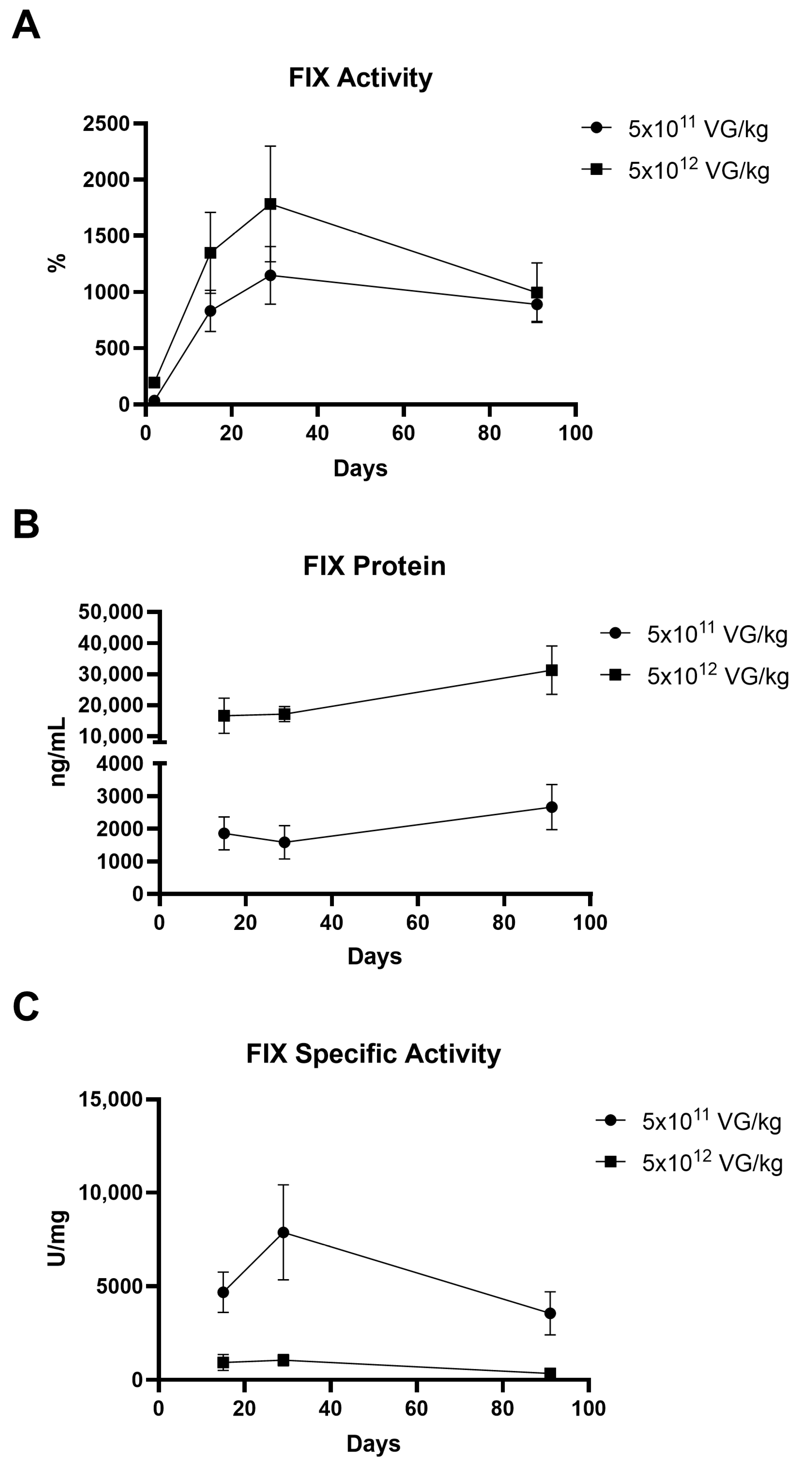
2.1. Evaluation of FIX Activity and Protein Expression Following AAV8-FIX-TripleL Administration in Hemophilia B Mice
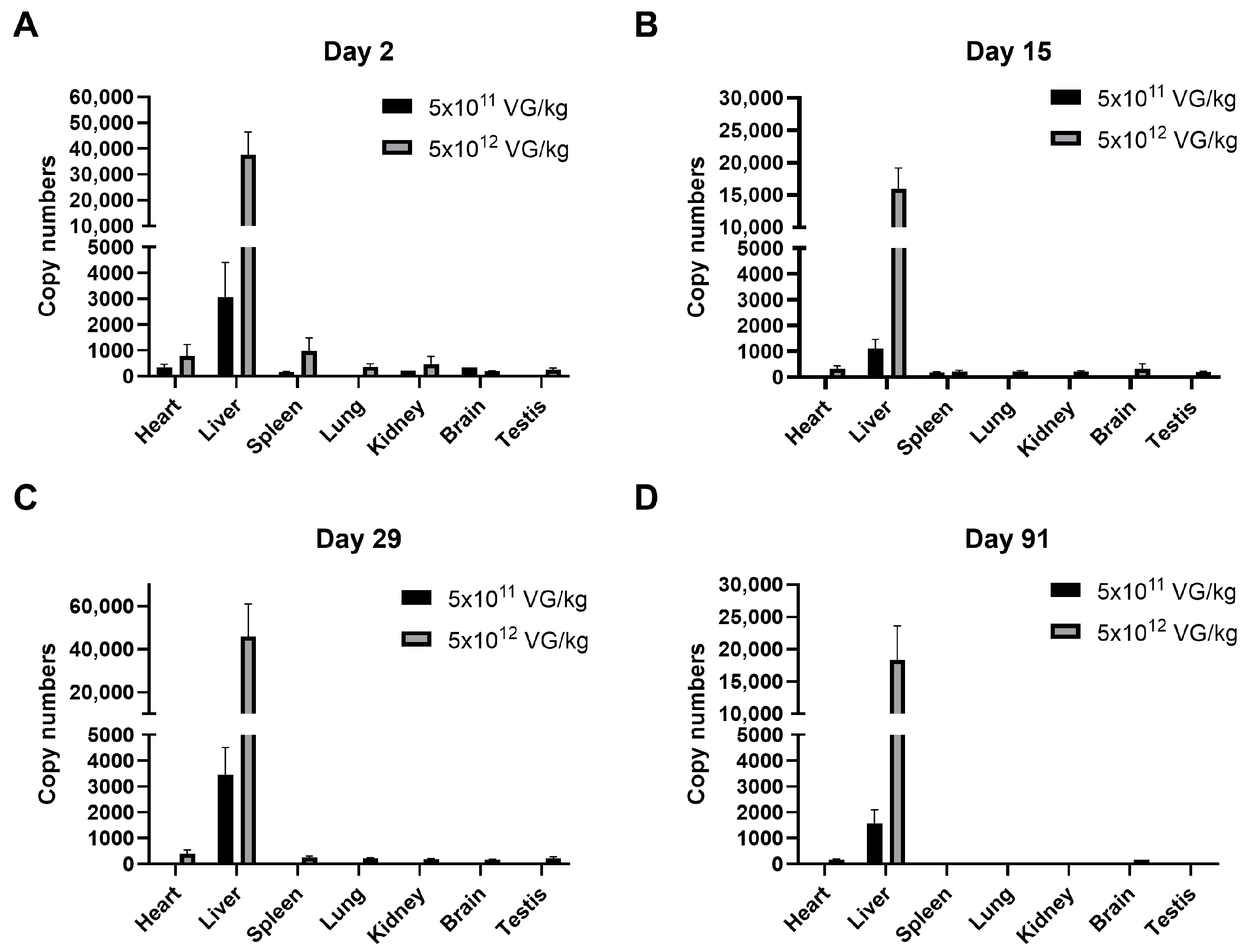
2.2. Biodistribution of AAV8-FIX-TripleL in Hemophilia B Mice
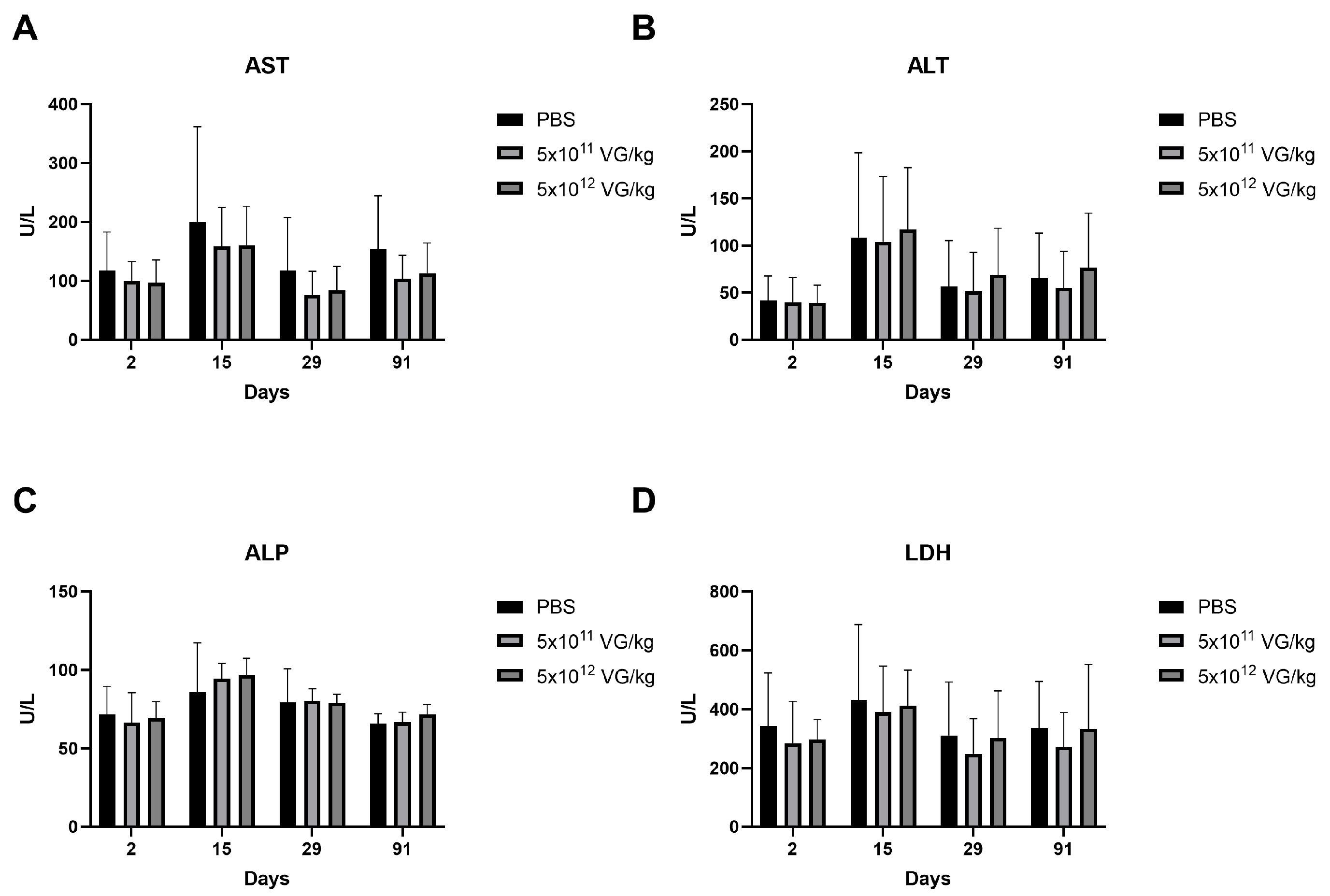
2.3. Safety Evaluation of AAV8-FIX-TripleL in Hemophilia B Mice
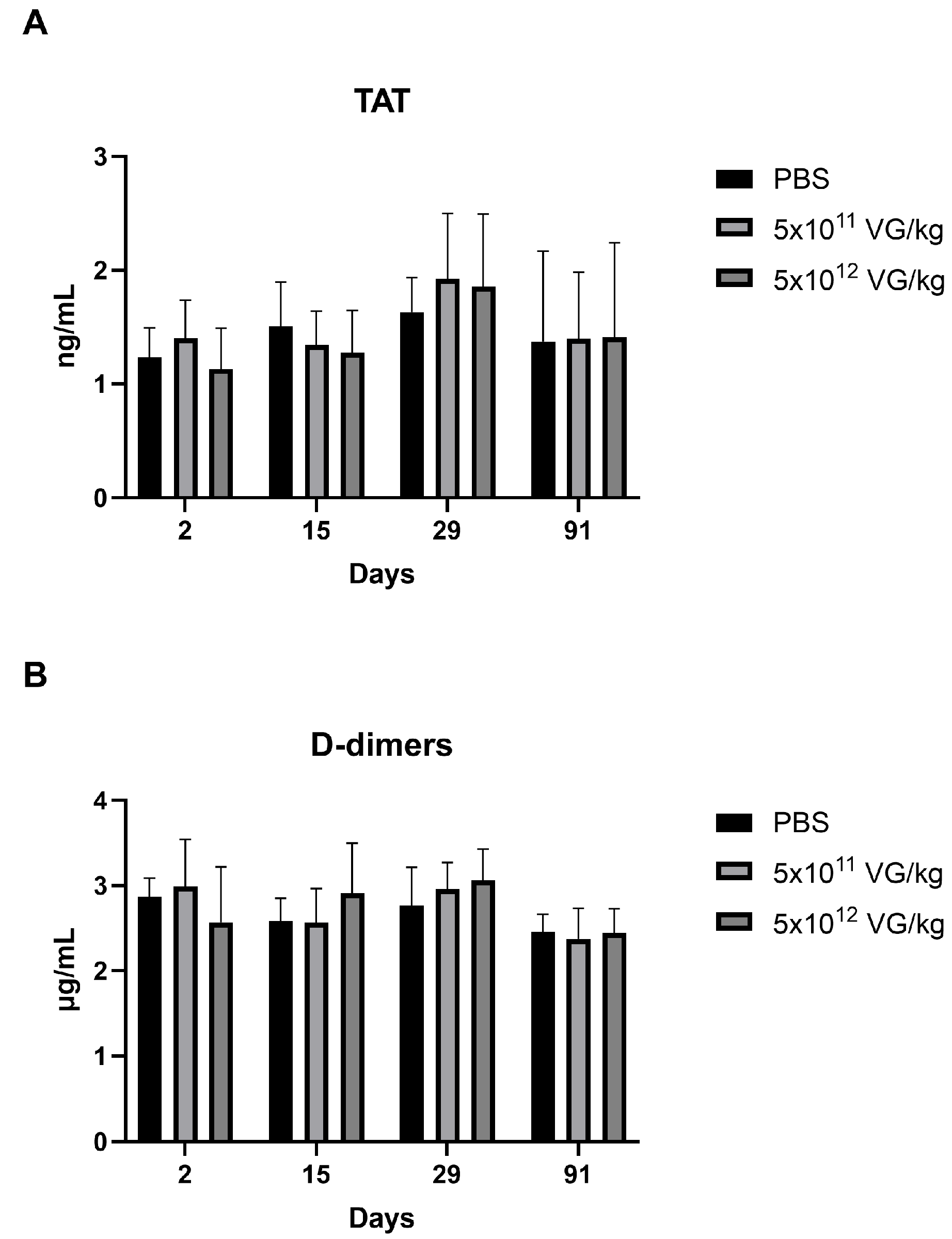
2.4. AAV8-FIX-TripleL Does Not Increase the Levels of the Prothrombotic Markers Thrombin-Antithrombin Complex and D-Dimers
2.5. Immunogenicity Evaluation of AAV8-FIX-TripleL in Hemophilia B Mice
3. Discussion
4. Materials and Methods
4.1. Recombinant AAV Vectors
4.2. Animal Experiments
4.3. ELISA for Quantitating hFIX in Plasma
4.4. FIX Activity Assay
4.5. Detection of hFIX DNA in Organs
4.6. Measurements of TAT and D-Dimer Concentrations
4.7. Detection of the Anti-AAV8 Capsid IgG Antibody
4.8. Detection of Anti-FIX IgG Antibodies
4.9. FIX-Neutralizing Antibody Detection (Bethesda Inhibitor)
4.10. Safety Assessments
4.11. Histopathological Examination
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pipe, S. Consideration in hemophilia therapy selection. Semin. Hematol. 2006, 43, S23–S27. [Google Scholar] [CrossRef] [PubMed]
- Goodeve, A.C. Hemophilia B: Molecular pathogenesis and mutation analysis. J. Thromb. Haemost. 2015, 13, 1184–1195. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Brewer, A.K.; Mauser-Bunschoten, E.P.; Key, N.S.; Kitchen, S.; Llinas, A.; Ludlam, C.A.; Mahlangu, J.N.; Mulder, K.; Poon, M.C.; et al. Guidelines for the management of hemophilia. Haemophilia 2013, 19, e1–e47. [Google Scholar] [CrossRef] [PubMed]
- Manco-Johnson, M.J.; Abshire, T.C.; Shapiro, A.D.; Riske, B.; Hacker, M.R.; Kilcoyne, R.; Ingram, J.D.; Manco-Johnson, M.L.; Funk, S.; Jacobson, L.; et al. Prophylaxis versus episodic treatment to prevent joint disease in boys with severe hemophilia. N. Engl. J. Med. 2007, 357, 535–544. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chang, C.Y.; Cheng, S.N.; Pan, R.Y.; Shih, Y.L.; Li, T.Y.; Wang, S.H. Evolution of congenital haemophilia care in Taiwan. J. Formos. Med. Assoc. 2022, 121, 582–591. [Google Scholar] [CrossRef]
- Castaman, G.; Matino, D. Hemophilia A and B: Molecular and clinical similarities and differences. Haematologica 2019, 104, 1702–1709. [Google Scholar] [CrossRef]
- DiMichele, D. Inhibitor development in haemophilia B: An orphan disease in need of attention. Br. J. Haematol. 2007, 138, 305–315. [Google Scholar] [CrossRef]
- Gater, A.; Thomson, T.A.; Strandberg-Larsen, M. Haemophilia B: Impact on patients and economic burden of disease. Thromb. Haemost. 2011, 106, 398–404. [Google Scholar] [CrossRef]
- High, K.A. Gene therapy for haemophilia: A long and winding road. J. Thromb. Haemost. 2011, 9 (Suppl. S1), 2–11. [Google Scholar] [CrossRef]
- Chou, S.C.; Hsu, Y.C.; Lin, S.W. Gene therapy for hemophilia, a clinical viewpoint. J. Formos. Med. Assoc. 2023, 122, 1101–1110. [Google Scholar] [CrossRef]
- Nathwani, A.C.; Tuddenham, E.G.; Rangarajan, S.; Rosales, C.; McIntosh, J.; Linch, D.C.; Chowdary, P.; Riddell, A.; Pie, A.J.; Harrington, C.; et al. Adenovirus-associated virus vector-mediated gene transfer in hemophilia B. N. Engl. J. Med. 2011, 365, 2357–2365. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, A.C.; Reiss, U.M.; Tuddenham, E.G.; Rosales, C.; Chowdary, P.; McIntosh, J.; Della Peruta, M.; Lheriteau, E.; Patel, N.; Raj, D.; et al. Long-term safety and efficacy of factor IX gene therapy in hemophilia B. N. Engl. J. Med. 2014, 371, 1994–2004. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.P.; Alvira, M.R.; Wang, L.; Calcedo, R.; Johnston, J.; Wilson, J.M. Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 11854–11859. [Google Scholar] [CrossRef]
- Nathwani, A.C.; Gray, J.T.; Ng, C.Y.; Zhou, J.; Spence, Y.; Waddington, S.N.; Tuddenham, E.G.; Kemball-Cook, G.; McIntosh, J.; Boon-Spijker, M.; et al. Self-complementary adeno-associated virus vectors containing a novel liver-specific human factor IX expression cassette enable highly efficient transduction of murine and nonhuman primate liver. Blood 2006, 107, 2653–2661. [Google Scholar] [CrossRef]
- Haas, J.; Park, E.C.; Seed, B. Codon usage limitation in the expression of HIV-1 envelope glycoprotein. Curr. Biol. 1996, 6, 315–324. [Google Scholar] [CrossRef]
- Pierce, G.F.; Iorio, A. Past, present and future of haemophilia gene therapy: From vectors and transgenes to known and unknown outcomes. Haemophilia 2018, 24 (Suppl. S6), 60–67. [Google Scholar] [CrossRef]
- Cuker, A.; Kavakli, K.; Frenzel, L.; Wang, J.D.; Astermark, J.; Cerqueira, M.H.; Iorio, A.; Katsarou-Fasouli, O.; Klamroth, R.; Shapiro, A.D.; et al. Gene Therapy with Fidanacogene Elaparvovec in Adults with Hemophilia B. N. Engl. J. Med. 2024, 391, 1108–1118. [Google Scholar] [CrossRef]
- Pipe, S.W.; Leebeek, F.W.G.; Recht, M.; Key, N.S.; Castaman, G.; Miesbach, W.; Lattimore, S.; Peerlinck, K.; Van der Valk, P.; Coppens, M.; et al. Gene Therapy with Etranacogene Dezaparvovec for Hemophilia B. N. Engl. J. Med. 2023, 388, 706–718. [Google Scholar] [CrossRef]
- Chang, J.; Jin, J.; Lollar, P.; Bode, W.; Brandstetter, H.; Hamaguchi, N.; Straight, D.L.; Stafford, D.W. Changing residue 338 in human factor IX from arginine to alanine causes an increase in catalytic activity. J. Biol. Chem. 1998, 273, 12089–12094. [Google Scholar] [CrossRef]
- Simioni, P.; Tormene, D.; Tognin, G.; Gavasso, S.; Bulato, C.; Iacobelli, N.P.; Finn, J.D.; Spiezia, L.; Radu, C.; Arruda, V.R. X-linked thrombophilia with a mutant factor IX (factor IX Padua). N. Engl. J. Med. 2009, 361, 1671–1675. [Google Scholar] [CrossRef]
- Kao, C.Y.; Yang, S.J.; Tao, M.H.; Jeng, Y.M.; Yu, I.S.; Lin, S.W. Incorporation of the factor IX Padua mutation into FIX-Triple improves clotting activity in vitro and in vivo. Thromb. Haemost. 2013, 110, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Monahan, P.E.; Sun, J.; Gui, T.; Hu, G.; Hannah, W.B.; Wichlan, D.G.; Wu, Z.; Grieger, J.C.; Li, C.; Suwanmanee, T.; et al. Employing a gain-of-function factor IX variant R338L to advance the efficacy and safety of hemophilia B human gene therapy: Preclinical evaluation supporting an ongoing adeno-associated virus clinical trial. Hum. Gene. Ther. 2015, 26, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.; Bell, P.; McCarter, R.J.; He, J.; Calcedo, R.; Vandenberghe, L.H.; Morizono, H.; Batshaw, M.L.; Wilson, J.M. Systematic evaluation of AAV vectors for liver directed gene transfer in murine models. Mol. Ther. 2010, 18, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kang, K.; Chen, S.; Su, Q.; Zhang, W.; Zeng, L.; Lin, X.; Peng, F.; Lin, J.; Chai, D. High serum lactate dehydrogenase as a predictor of cardiac insufficiency at follow-up in elderly patients with acute myocardial infarction. Arch. Gerontol. Geriatr. 2024, 117, 105253. [Google Scholar] [CrossRef]
- Brancaccio, P.; Lippi, G.; Maffulli, N. Biochemical markers of muscular damage. Clin. Chem. Lab. Med. 2010, 48, 757–767. [Google Scholar] [CrossRef]
- Speiser, W.; Mallek, R.; Koppensteiner, R.; Stumpflen, A.; Kapiotis, S.; Minar, E.; Ehringer, H.; Lechner, K. D-dimer and TAT measurement in patients with deep venous thrombosis: Utility in diagnosis and judgement of anticoagulant treatment effectiveness. Thromb. Haemost. 1990, 64, 196–201. [Google Scholar] [CrossRef]
- Pulivarthi, S.; Gurram, M.K. Effectiveness of d-dimer as a screening test for venous thromboembolism: An update. N. Am. J. Med. Sci. 2014, 6, 491–499. [Google Scholar] [CrossRef]
- Greig, J.A.; Limberis, M.P.; Bell, P.; Chen, S.J.; Calcedo, R.; Rader, D.J.; Wilson, J.M. Non-Clinical Study Examining AAV8.TBG.hLDLR Vector-Associated Toxicity in Chow-Fed Wild-Type and LDLR(+/−) Rhesus Macaques. Hum. Gene Ther. Clin. Dev. 2017, 28, 39–50. [Google Scholar] [CrossRef]
- Kloosterman, F.; Zwagemaker, A.F.; Abdi, A.; Gouw, S.; Castaman, G.; Fijnvandraat, K. Hemophilia management: Huge impact of a tiny difference. Res. Pract. Thromb. Haemost. 2020, 4, 377–385. [Google Scholar] [CrossRef]
- Crudele, J.M.; Finn, J.D.; Siner, J.I.; Martin, N.B.; Niemeyer, G.P.; Zhou, S.; Mingozzi, F.; Lothrop, C.D., Jr.; Arruda, V.R. AAV liver expression of FIX-Padua prevents and eradicates FIX inhibitor without increasing thrombogenicity in hemophilia B dogs and mice. Blood 2015, 125, 1553–1561. [Google Scholar] [CrossRef]
- Kohn, D.B.; Chen, Y.Y.; Spencer, M.J. Successes and challenges in clinical gene therapy. Gene Ther. 2023, 30, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, A.C. Gene therapy for hemophilia. Hematol. Am. Soc. Hematol. Educ. Program 2022, 2022, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Soroka, A.B.; Feoktistova, S.G.; Mityaeva, O.N.; Volchkov, P.Y. Gene Therapy Approaches for the Treatment of Hemophilia B. Int. J. Mol. Sci. 2023, 24, 10766. [Google Scholar] [CrossRef] [PubMed]
- Chowdary, P.; Shapiro, S.; Makris, M.; Evans, G.; Boyce, S.; Talks, K.; Dolan, G.; Reiss, U.; Phillips, M.; Riddell, A.; et al. Phase 1-2 Trial of AAVS3 Gene Therapy in Patients with Hemophilia B. N. Engl. J. Med. 2022, 387, 237–247. [Google Scholar] [CrossRef]
- Bowyer, A.E.; Gosselin, R.C. Factor VIII and Factor IX Activity Measurements for Hemophilia Diagnosis and Related Treatments. Semin. Thromb. Hemost. 2023, 49, 609–620. [Google Scholar] [CrossRef]
- Chen, V.P.; Gao, Y.; Geng, L.; Steele, M.; Jenks, N.; Peng, K.W.; Brimijoin, S. Systemic Safety of a Recombinant AAV8 Vector for Human Cocaine Hydrolase Gene Therapy: A Good Laboratory Practice Preclinical Study in Mice. Hum. Gene Ther. 2020, 31, 70–79. [Google Scholar] [CrossRef]
- Miller, C.H. Laboratory testing for factor VIII and IX inhibitors in haemophilia: A review. Haemophilia 2018, 24, 186–197. [Google Scholar] [CrossRef]
- Shackelford, C.; Long, G.; Wolf, J.; Okerberg, C.; Herbert, R. Qualitative and quantitative analysis of nonneoplastic lesions in toxicology studies. Toxicol. Pathol. 2002, 30, 93–96. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Vector Treatment | Dose (VG/kg) | Harvest Day | Animal Number/Sex |
|---|---|---|---|---|
| 1 | PBS | - | Day 2 | 15/Male |
| 2 | AAV8-FIX-TripleL | 5 × 1011 | Day 2 | 15/Male |
| 3 | AAV8-FIX-TripleL | 5 × 1012 | Day 2 | 15/Male |
| 4 | PBS | - | Day 15 | 15/Male |
| 5 | AAV8-FIX-TripleL | 5 × 1011 | Day 15 | 15/Male |
| 6 | AAV8-FIX-TripleL | 5 × 1012 | Day 15 | 15/Male |
| 7 | PBS | - | Day 29 | 15/Male |
| 8 | AAV8-FIX-TripleL | 5 × 1011 | Day 29 | 15/Male |
| 9 | AAV8-FIX-TripleL | 5 × 1012 | Day 29 | 15/Male |
| 10 | PBS | - | Day 91 | 15/Male |
| 11 | AAV8-FIX-TripleL | 5 × 1011 | Day 91 | 15/Male |
| 12 | AAV8-FIX-TripleL | 5 × 1012 | Day 91 | 15/Male |
| Days | Day 2 | Day 15 | Day 29 | Day 91 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose (VG/kg) | - | 5 × 1011 | 5 × 1012 | - | 5 × 1011 | 5 × 1012 | - | 5 × 1011 | 5 × 1012 | - | 5 × 1011 | 5 × 1012 | |
| Lung | Osseous metaplasia, alveoli, focal, minimal | 1/15 | - | - | - | - | - | - | - | - | - | - | - |
| Score value | 0.07 ± 0.26 | - | - | - | - | - | - | - | - | - | - | - | |
| Infiltration, mononuclear cell, focal, minimal | - | - | 1/15 | - | - | 1/15 | - | - | 1/15 | 1/15 | - | 1/15 | |
| Score value | - | - | 0.07 ± 0.26 | - | - | 0.07 ± 0.26 | - | - | 0.07 ± 0.26 | 0.07 ± 0.26 | - | 0.07 ± 0.26 | |
| Heart | Infiltration, mononuclear cell, focal, minimal | 1/15 | - | - | - | - | - | 1/15 | - | - | - | - | - |
| Score value | 0.07 ± 0.26 | - | - | - | - | - | 0.07 ± 0.26 | - | - | - | - | - | |
| Mineralization, focal, minimal | - | - | - | 1/15 | - | - | - | - | - | - | - | - | |
| Score value | - | - | - | 0.07 ± 0.26 | - | - | - | - | - | - | - | - | |
| Cardiomyopathy, focal, minimal | - | - | - | - | - | - | - | - | 1/15 | - | - | - | |
| Score value | - | - | - | - | - | - | - | - | 0.07 ± 0.26 | - | - | - | |
| Spleen | Pigment, focal, minimal | 1/15 | 1/15 | - | - | - | - | - | - | - | - | - | - |
| Score value | 0.07 ± 0.26 | 0.07 ± 0.26 | - | - | - | - | - | - | - | - | - | - | |
| Pancreas | Vacuolation, focal, minimal | 1/15 | - | - | - | - | - | - | - | - | - | - | - |
| Score value | 0.07 ± 0.26 | - | - | - | - | - | - | - | - | - | - | - | |
| Liver | Infiltration, mononuclear cell, focal, minimal | 8/15 | 5/15 | 7/15 | 9/15 | 9/15 | 6/15 | 5/15 | 9/15 | 8/15 | 8/15 | 7/15 | 2/15 |
| Score value | 0.53 ± 0.52 | 0.33 ± 0.49 | 0.47 ± 0.52 | 0.60 ± 0.51 | 0.60 ± 0.51 | 0.40 ± 0.51 | 0.33 ± 0.49 | 0.60 ± 0.51 | 0.60 ± 0.63 | 0.53 ± 0.52 | 0.47 ± 0.52 | 0.13 ± 0.35 * | |
| Necrosis, focal, minimal | - | - | - | 2/15 | - | - | - | - | - | - | - | - | |
| Score value | - | - | - | 0.13 ± 0.35 | - | - | - | - | - | - | - | - | |
| Fibrosis, focal, mild | - | - | - | - | - | - | - | - | 1/15 | - | - | - | |
| Score value | - | - | - | - | - | - | - | - | 0.13 ± 0.52 | - | - | - | |
| Kidneys | Infiltration, mononuclear cell, cortex, focal, minimal | 1/15 | 3/15 | 1/15 | 3/15 | - | 3/15 | 4/15 | - | 3/15 | 4/15 | 3/15 | 5/15 |
| Score value | 0.07 ± 0.26 | 0.20 ± 0.41 | 0.07 ± 0.26 | 0.20 ± 0.41 | - | 0.20 ± 0.41 | 0.27 ± 0.46 | - | 0.20 ± 0.41 | 0.27 ± 0.46 | 0.27 ± 0.59 | 0.33 ± 0.49 | |
| Vacuolation, glomerulus, focal, minimal | - | - | 1/15 | - | - | - | - | - | - | - | - | - | |
| Score value | - | - | 0.07 ± 0.26 | - | - | - | - | - | - | - | - | - | |
| Cyst, cortex/medulla, focal, minimal to moderate | 3/15 | - | - | - | 1/15 | - | - | - | - | - | - | - | |
| Score value | 0.40 ± 0.91 | - | - | - | 0.07 ± 0.26 | - | - | - | - | - | - | - | |
| Cast, renal tubule, cortex, focal, minimal | 2/15 | - | - | - | 1/15 | - | 1/15 | 1/15 | - | - | 1/15 | 2/15 | |
| Score value | 0.13 ± 0.35 | - | - | - | 0.07 ± 0.26 | - | 0.07 ± 0.26 | 0.07 ± 0.26 | - | - | 0.07 ± 0.26 | 0.13 ± 0.35 | |
| Basophilia, tubule, cortex, focal, minimal | - | - | - | - | 1/15 | - | - | - | - | - | - | - | |
| Score value | - | - | - | - | 0.07 ± 0.26 | - | - | - | - | - | - | - | |
| Dilation, tubule, cortex, focal, minimal | - | - | - | - | - | - | 1/15 | - | - | - | - | - | |
| Score value | - | - | - | - | - | - | 0.07 ± 0.26 | - | - | - | - | - | |
| Adrenals | Hyperplasia, subcapsular, cortex, focal, minimal | 2/15 | 1/15 | 1/15 | - | 1/15 | - | 1/15 | - | - | - | 2/15 | - |
| Score value | 0.13 ± 0.35 | 0.07 ± 0.26 | 0.07 ± 0.26 | - | 0.07 ± 0.26 | - | 0.07 ± 0.26 | - | - | - | 0.13 ± 0.35 | - | |
| Accessory adrenocortical nodule, focal, minimal to mild | - | 2/15 | - | - | - | - | - | - | - | 1/15 | 1/15 | - | |
| Score value | - | 0.20 ± 0.56 | - | - | - | - | - | - | - | 0.07 ± 0.26 | 0.07 ± 0.26 | ||
| Testis | Degeneration, germ cell, focal, moderate | - | - | - | - | - | - | 1/15 | 1/15 | - | - | - | - |
| Score value | - | - | - | - | - | - | 0.20 ± 0.77 | 0.20 ± 0.77 | - | - | - | - | |
| Days | Day 2 | Day 15 | Day 29 | Day 91 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose (VG/kg) | - | 5 × 1011 | 5 × 1012 | - | 5 × 1011 | 5 × 1012 | - | 5 × 1011 | 5 × 1012 | - | 5 × 1011 | 5 × 1012 |
| AAV8 antibody/total no. of animals | 0/15 | 0/15 | 0/15 | 0/15 | 15/15 | 15/15 | 0/15 | 15/15 | 15/15 | 0/15 | 15/15 | 15/15 |
| Range (μg/mL) | - | - | - | - | 5.21 ± 2.38 | 10.99 ± 4.51 | - | 11.70 ± 5.51 | 32.79 ± 12.10 | - | 24.78 ± 25.15 | 42.50 ± 22.36 |
| FIX antibody/total no. of animals | 0/15 | 0/15 | 0/15 | 0/15 | 1/15 | 0/15 | 0/15 | 0/15 | 4/15 | 0/15 | 0/15 | 4/15 |
| Range (μg/mL) | - | - | - | - | 7.61 | - | - | - | 3.60 ± 5.48 | - | - | 0.57 ± 0.31 |
| Bethesda inhibitor/total no. of animals | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 | 0/15 |
| Titer (BIU) | - | - | - | - | - | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, S.-C.; Huang, C.-P.; Su, Y.-H.; Yu, C.-H.; Yang, Y.-L.; Wang, S.-C.; Lin, Y.-H.; Chen, Y.-T.; Li, J.-Y.; Chang, Y.-T.; et al. Preclinical Evaluation of the Systemic Safety, Efficacy, and Biodistribution of a Recombinant AAV8 Vector Expressing FIX-TripleL in Hemophilia B Mice: Implications for Human Gene Therapy. Int. J. Mol. Sci. 2025, 26, 6073. https://doi.org/10.3390/ijms26136073
Chou S-C, Huang C-P, Su Y-H, Yu C-H, Yang Y-L, Wang S-C, Lin Y-H, Chen Y-T, Li J-Y, Chang Y-T, et al. Preclinical Evaluation of the Systemic Safety, Efficacy, and Biodistribution of a Recombinant AAV8 Vector Expressing FIX-TripleL in Hemophilia B Mice: Implications for Human Gene Therapy. International Journal of Molecular Sciences. 2025; 26(13):6073. https://doi.org/10.3390/ijms26136073
Chicago/Turabian StyleChou, Sheng-Chieh, Cheng-Po Huang, Ying-Hui Su, Chih-Hsiang Yu, Yung-Li Yang, Ssu-Chia Wang, Yi-Hsiu Lin, Yen-Ting Chen, Jia-Yi Li, Yen-Ting Chang, and et al. 2025. "Preclinical Evaluation of the Systemic Safety, Efficacy, and Biodistribution of a Recombinant AAV8 Vector Expressing FIX-TripleL in Hemophilia B Mice: Implications for Human Gene Therapy" International Journal of Molecular Sciences 26, no. 13: 6073. https://doi.org/10.3390/ijms26136073
APA StyleChou, S.-C., Huang, C.-P., Su, Y.-H., Yu, C.-H., Yang, Y.-L., Wang, S.-C., Lin, Y.-H., Chen, Y.-T., Li, J.-Y., Chang, Y.-T., Chen, S.-Y., & Lin, S.-W. (2025). Preclinical Evaluation of the Systemic Safety, Efficacy, and Biodistribution of a Recombinant AAV8 Vector Expressing FIX-TripleL in Hemophilia B Mice: Implications for Human Gene Therapy. International Journal of Molecular Sciences, 26(13), 6073. https://doi.org/10.3390/ijms26136073