
Identification and Expression Analysis of G-Protein-Coupled Receptors Provide Insights into Functional and Mechanistic Responses to Herbivore-Induced Plant Volatiles of Paracarophenax alternatus


Abstract
1. Introduction
2. Results
2.1. Identification and Phylogenetic Analysis of GPCR Genes in P. alternatus
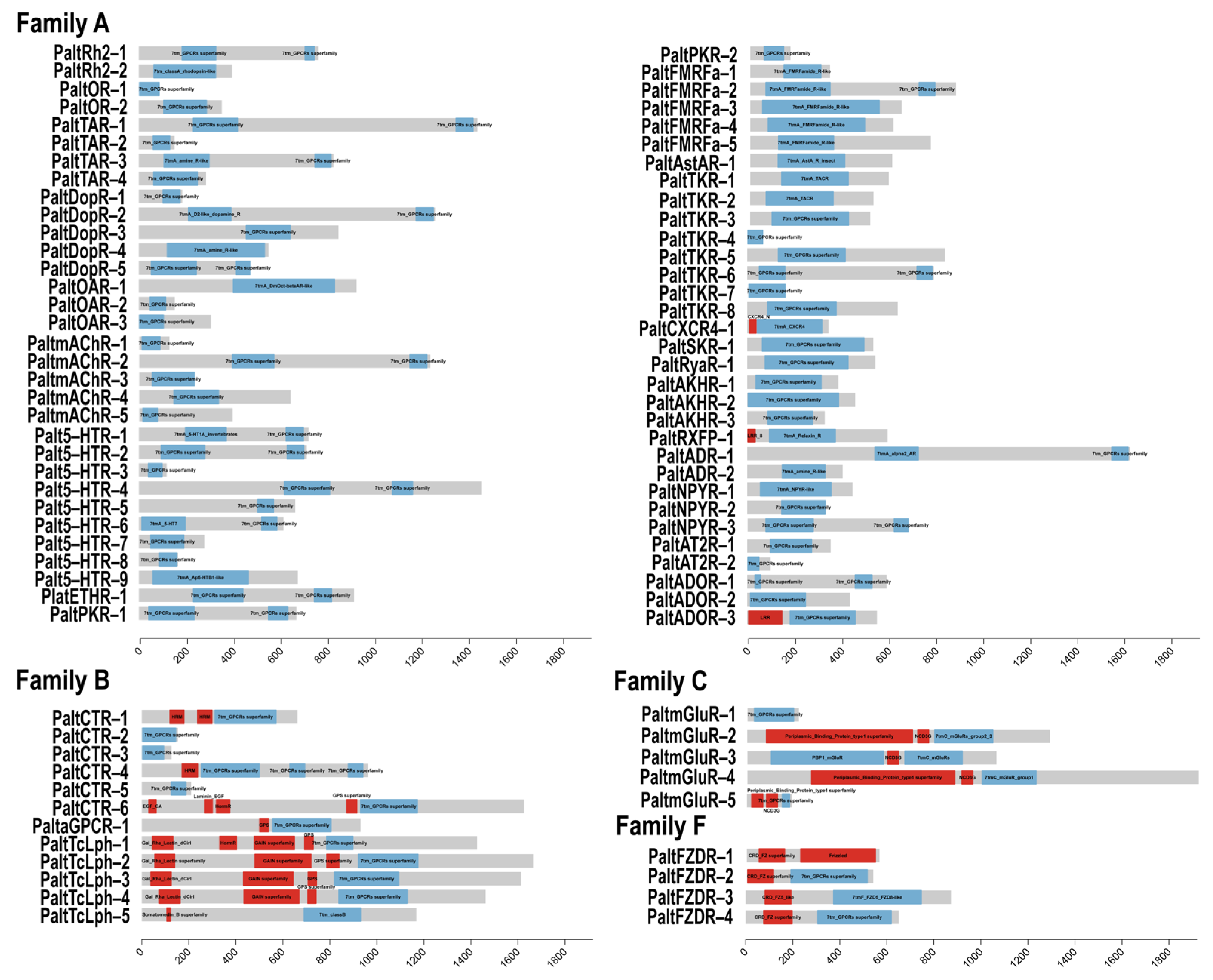
2.2. Identification of Conserved Motifs and Domain Analysis
2.3. Expression Profiles of GPCR Genes in Different Physiological States
2.4. Molecular Dynamics Simulation
3. Discussion
4. Materials and Methods
4.1. Insect and Mite Rearing
4.2. Identification and Analysis of P. alternatus GPCRs
4.3. Phylogenetic Analysis
4.4. RNA Extraction and cDNA Preparation
4.5. Real-Time Quantitative PCR
4.6. Structure Prediction and Molecular Docking
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Primer Sequences |
|---|---|---|
| GAPDH | GAPDH-F | ATCGAAAACGCAGCTTGCAG |
| GAPDH-R | GTCGTGGTGCAGGACAAAAC | |
| PaltRh2-1 | PaltRh2-1-F | GTTAGGTATCACCGTATTGCCCT |
| PaltRh2-1-R | GAAGCAGTAGATAGCCAAACGTC | |
| PaltRh2-2 | PaltRh2-2-F | TATGGTCAACGGATGTGCGTAA |
| PaltRh2-2-R | GTCCTTAGTTGTGGTTGCATTGG | |
| PaltOR-1 | PaltOR-1-F | CATGACACTTTTCACCTATGCCC |
| PaltOR-1-R | TAAATGATGGGGTTGAGGAGTGG | |
| PaltOR-2 | PaltOR-2-F | TGCTCTTTATAGTCGAGTTTGTGT |
| PaltOR-2-R | GCAACAGCTTGACCATTTATTCG | |
| PaltTAR-1 | PaltTAR-1-F | GCCTTTATCTGTGGCGTACAATG |
| PaltTAR-1-R | GGCTTGATACCTATCTACCGCAA | |
| PaltTAR-2 | PaltTAR-2-F | GTTCACATATCCTAGCCAGGTGA |
| PaltTAR-2-R | CTCAGGTACTTGGCATTGTGATG | |
| PaltTAR-3 | PaltTAR-3-F | ATCCAGTTTACGTGGTGTTTGC |
| PaltTAR-3-R | CTAGATCGAGCAAACAGATGGC | |
| PaltTAR-4 | PaltTAR-4-F | TGTGTGAAAATGATTTGTGCCCT |
| PaltTAR-4-R | ATTCACCAAACCCATCACCATTG | |
| PaltDopR-1 | PaltDopR-1-F | CTTCTCATGCCGTCTGTTTAAGC |
| PaltDopR-1-R | AAAACCAGGCAGGCTTAAGACT | |
| PaltDopR-2 | PaltDopR-2-F | CTGAACAGCAGCCTTTTAACACC |
| PaltDopR-2-R | GTCAGTTACTTCTTGCGAATCCG | |
| PaltDopR-3 | PaltDopR-3-F | GCAGTAAGCCCAGAAAATAAGCC |
| PaltDopR-3-R | CACCATGACTACCAAAGGACCAT | |
| PaltDopR-4 | PaltDopR-4-F | TGATGCCATTTGGTGTTATTGCT |
| PaltDopR-4-R | TCAGAATTGATGCAGTACAGGCA | |
| PaltDopR-5 | PaltDopR-5-F | TCAATAAAGCCGCAAGTGAATGG |
| PaltDopR-5-R | TTTCAGCTTCAACTTGCATTTGC | |
| PaltOAR-1 | PaltOAR-1-F | CCGTGTTGTCTGCATATTCGTC |
| PaltOAR-1-R | AGTCGGGAAACAATGGTGATGA | |
| PaltOAR-2 | PaltOAR-2-F | CGAAGAAGATGGTGTCAACGC |
| PaltOAR-2-R | ACTTTTCCACCACTAGACGGAG | |
| PaltOAR-3 | PaltOAR-3-F | CATGCCGTAGCTATGAGTTCAAC |
| PaltOAR-3-R | CTAGGTCCAGTTCGAAAACAAGG | |
| PaltmAChR-1 | PaltmAChR-1-F | GACATGGACACCGTATAATGTGG |
| PaltmAChR-1-R | GCATAGCAAAGGGGATTTACAGTG | |
| PaltmAChR-2 | PaltmAChR-2-F | GAAGCATCTTTTCCAGAGATGGC |
| PaltmAChR-2-R | GAGCTTGAGTACACAGGTGATCT | |
| PaltmAChR-3 | PaltmAChR-3-F | GGCTGAACAGGAAGTAGTTGTTG |
| PaltmAChR-3-R | GAGACAGTGGAGATGGTGTTCAT | |
| PaltmAChR-4 | PaltmAChR-4-F | GTTCGACGTACTACTAGACGAGC |
| PaltmAChR-4-R | AGTACGTTCACCCTCAATGTAGG | |
| PaltmAChR-5 | PaltmAChR-5-F | CGAGTAATGGCCACACATATCAC |
| PaltmAChR-5-R | CTGTGAGGTAGTAGGTGATTGCA | |
| Palt5-HTR-1 | Palt5-HTR-1-F | ATTGCCGCTGTTTTAAGTGACC |
| Palt5-HTR-1-R | AAAACGCCTAATGTCCATTGGC | |
| Palt5-HTR-2 | Palt5-HTR-2-F | GCTGGCAACTTGGTTCAGAATT |
| Palt5-HTR-2-R | TCCAGTACTTCGACCCTGACTAT | |
| Palt5-HTR-3 | Palt5-HTR-3-F | TGTAATGGGTGTTTTCGGCA |
| Palt5-HTR-3-R | ACAACATGAGGAATGGCATCAC | |
| Palt5-HTR-4 | Palt5-HTR-4-F | AAGAGCAGTGAGGTTCCTACAAG |
| Palt5-HTR-4-R | GTGGACAAAGACCAATCTGCTAC | |
| Palt5-HTR-5 | Palt5-HTR-5-F | CTTAACTTGGTTGGGGGTAATCG |
| Palt5-HTR-5-R | GGACGCAATGGAACTGCTTAAT | |
| Palt5-HTR-6 | Palt5-HTR-6-F | GCATTAAGTGATCTTGGTGTGGC |
| Palt5-HTR-6-R | CAGCAAACGACATCTAAACCCAG | |
| Palt5-HTR-7 | Palt5-HTR-7-F | CACCGTGTTTAAATCATCTCGTGT |
| Palt5-HTR-7-R | ACAATTCCATTTCCAAAAACTGCG | |
| Palt5-HTR-8 | Palt5-HTR-8-F | CCGCAAACTAGTAGTGTAGACCA |
| Palt5-HTR-8-R | ACTTTGAAAATCGCTTTTTGCCG | |
| Palt5-HTR-9 | Palt5-HTR-9-F | GCAGTGGTCAGTTCTTGCATTAG |
| Palt5-HTR-9-R | GGCTTGAGGGCTAATATTAATGGC | |
| PlatETHR-1 | PlatETHR-1-F | TTTCACACTGGCTTTGCTATGC |
| PlatETHR-1-R | CATACACATACAGCTGAACGCG | |
| PaltPKR-1 | PaltPKR-1-F | CCAATATCGGGATCCCAGTCTTT |
| PaltPKR-1-R | TCCATGATTTGAATCGCCAGGA | |
| PaltPKR-2 | PaltPKR-2-F | TTTGAGCTTCAGTAGGCTGAGTG |
| PaltPKR-2-R | AAAGATGTTAGCTGCCGTGGTA | |
| PaltFMRFa-1 | PaltFMRFa-1-F | TGATGCCATTTGGTGTTATTGCT |
| PaltFMRFa-1-R | TCAGAATTGATGCAGTACAGGCA | |
| PaltFMRFa-2 | PaltFMRFa-2-F | ACGGTATAGTAAGACATGGGCTG |
| PaltFMRFa-2-R | CCATGTAACGCCATACACCTAGT | |
| PaltFMRFa-3 | PaltFMRFa-3-F | TGGCACAGAAAGTATTGCTTCG |
| PaltFMRFa-3-R | AACAAGACAAGAATGCCGGAAG | |
| PaltFMRFa-4 | PaltFMRFa-4-F | GCCTCTTGTCAGACAACTGTTTC |
| PaltFMRFa-4-R | AAAGCATGCGGTTGATGTACAG | |
| PaltFMRFa-5 | PaltFMRFa-5-F | CCTTGTTACTTGCAGGTCAAAGG |
| PaltFMRFa-5-R | GTACTCTGATGGAGAAATGCAGC | |
| PaltAstAR-1 | PaltAstAR-1-F | GGTCATAGTAGTGGTTTGCAATCC |
| PaltAstAR-1-R | CTGAAGCAGTAAACGGTACACA | |
| PaltTKR-1 | PaltTKR-1-F | CCAGATGGTGTAGCACAAGAATC |
| PaltTKR-1-R | CCCCATAGTACAATGGCTACTCG | |
| PaltTKR-2 | PaltTKR-2-F | TGGTGACTCTCGTGTAATCTGT |
| PaltTKR-2-R | AGGCACCATGTACGTTACAAGT | |
| PaltTKR-3 | PaltTKR-3-F | AATGTTAGAACGGCATACCAGC |
| PaltTKR-3-R | AATGTGCACAGCTTGGAATGTC | |
| PaltTKR-4 | PaltTKR-4-F | GTACTTCAATGTGCACAGCTTGG |
| PaltTKR-4-R | CAATGTTAGAACGGCATACCAGC | |
| PaltTKR-5 | PaltTKR-5-F | GCCTCTGCGGTTACTACTGTTAA |
| PaltTKR-5-R | TGTAGAGTCGAAACTAGCATCCG | |
| PaltTKR-6 | PaltTKR-6-F | CTGGGTGCTTCCATACTTTCTCT |
| PaltTKR-6-R | GACCACTAAATAACGATCTGCTGC | |
| PaltTKR-7 | PaltTKR-7-F | TGCAACGTAACGGTCGAAAC |
| PaltTKR-7-R | TGGAAAGCTGGCCTTATGGG | |
| PaltTKR-8 | PaltTKR-8-F | CGACCGTTACGTTGCAATTGTA |
| PaltTKR-8-R | GGCATAGCTACAATCAACGAACC | |
| PaltCXCR4-1 | PaltCXCR4-1-F | GCCCACCATCTACTTCATCATCT |
| PaltCXCR4-1-R | TGTGATGACAAAGAGGAGGTCAG | |
| PaltRyaR-1 | PaltRyaR-1-F | CAGTATTGGCCTTTTGGTTCTGTC |
| PaltRyaR-1-R | TCTGTCTATGCTGATTGCCACA | |
| PaltAKHR-1 | PaltAKHR-1-F | TCGAGCAGCACCTCAAAATACA |
| PaltAKHR-1-R | CTGTTCGGTTAAAGGCAAATCCTC | |
| PaltAKHR-2 | PaltAKHR-2-F | GCATCTCGCAATCAGTGACTTAC |
| PaltAKHR-2-R | TAGAGCCATATGTCACAAGCACC | |
| PaltAKHR-3 | PaltAKHR-3-F | TCAAGAAGGTCAGCTAAATGGCA |
| PaltAKHR-3-R | ACAACACCAGCAAATTACCAGTT | |
| PaltRXFP-1 | PaltRXFP-1-F | GACAAGTAGGTCGTCAAATGGCT |
| PaltRXFP-1-R | GGGAAGAATAAATGCTGCAGTCC | |
| PaltADR-1 | PaltADR-1-F | GATGGAATCAGGGTTCGGCTTAA |
| PaltADR-1-R | CGTCGTTTTGCCCTCTTGATTAG | |
| PaltADR-2 | PaltADR-2-F | CTTACGAGTTTGGCCATTTGGAG |
| PaltADR-2-R | GACTTGTGATTGCCCAGTACCTA | |
| PaltNPYR-1 | PaltNPYR-1-F | TAGTAGCTGGTCTAGAAGCCACA |
| PaltNPYR-1-R | TGATCTCAATGGTTGGCACGAT | |
| PaltNPYR-2 | PaltNPYR-2-F | GGTGTTACGGGTAATGGTCTAGT |
| PaltNPYR-2-R | AGAGGTGTTATTGGTACGGCAA | |
| PaltNPYR-3 | PaltNPYR-3-F | AAAACATCTGTGCCTTCTTCACA |
| PaltNPYR-3-R | AGGTGGCCAATTGCAGAATATTG | |
| PaltAT2R-1 | PaltAT2R-1-F | AGCCACGGGAATTTAGAGTTGT |
| PaltAT2R-1-R | CAATCCAAGGCACAAGCATCTC | |
| PaltAT2R-2 | PaltAT2R-2-F | GTCTACCAAGTTTCGAAGCCAAC |
| PaltAT2R-2-R | CAAGTTGAAGCGTAGTTCGTCTG | |
| PaltADOR-1 | PaltADOR-1-F | GAGGCAGTGGGTACTTCTTTTTG |
| PaltADOR-1-R | TGATAACTCGTATTATCCGCTGGG | |
| PaltADOR-2 | PaltADOR-2-F | GATAGTGGTTTAGGAGTGCACCA |
| PaltADOR-2-R | TAACCGCCTAGATTTGAACCTCC | |
| PaltADOR-3 | PaltADOR-3-F | CTATGGCTGCCCCTAAAGAAATG |
| PaltADOR-3-R | TGAAGTTCTGCCTGAGGGATATG | |
| PaltCTR-1 | PaltCTR-1-F | GTGGATTCCATCGAAGTGCAAC |
| PaltCTR-1-R | GCTCCAACCATCAAAAGTAGCC | |
| PaltCTR-2 | PaltCTR-2-F | GGCAATTCTCATTATTGGCCAACTAT |
| PaltCTR-2-R | ACCCCATCCAAGAATTACGTATTTG | |
| PaltCTR-3 | PaltCTR-3-F | TGCTGCAGTTAAAGCATCAATTACA |
| PaltCTR-3-R | TTCCCAAATAAGATAAAACTTCTTGCA | |
| PaltCTR-4 | PaltCTR-4-F | GTTTGCTGGCTCGCTGAATTAT |
| PaltCTR-4-R | ACTTGCTGTACCTCGTTAGTGG | |
| PaltCTR-5 | PaltCTR-5-F | TATCACTACGCGCTGCATCC |
| PaltCTR-5-R | AATTCAGCGAGCCAGCAAAC | |
| PaltCTR-6 | PaltCTR-6-F | GACAGTGCCATTGTAAACCAGG |
| PaltCTR-6-R | TATCATAGACGACTTCGCAGCC | |
| PaltaGPCR-1 | PaltaGPCR-1-F | TGTGCGAATTACATAAACCTCGA |
| PaltaGPCR-1-R | ACTGTAGGAGCGATAGTAGCTT | |
| PaltTcLph-1 | PaltTcLph-1-F | CTGGAATAGCGCAATGGTTCTG |
| PaltTcLph-1-R | ACATTGAGCGAATATTGGCTGC | |
| PaltTcLph-2 | PaltTcLph-2-F | AGGCAGTTTTCCTGTTAACCCA |
| PaltTcLph-2-R | CAGGTGGGTTATGGTGCCTTAT | |
| PaltTcLph-3 | PaltTcLph-3-F | GAACACCATAAGTTTGCACCCC |
| PaltTcLph-3-R | CAGCTACTTCCTGTGGTTGCTA | |
| PaltTcLph-4 | PaltTcLph-4-F | TTGAACAGCCATGTCCAGAAGA |
| PaltTcLph-4-R | GGTTTTGAGCTTGGCATCCATT | |
| PaltTcLph-5 | PaltTcLph-5-F | GTTCGAAACACTTGTGCAGACA |
| PaltTcLph-5-R | GGCACAATGCGGATTCCAATAA | |
| PaltmGluR-1 | PaltmGluR-1-F | CATGGAGATTTACAGGTGCCTTTT |
| PaltmGluR-1-R | CCAAGTAATGTAGTGCCAACATTGT | |
| PaltmGluR-2 | PaltmGluR-2-F | TGGTTGGCTTTTGTACCCATCT |
| PaltmGluR-2-R | AAGCCACATATGCAGACAGTGA | |
| PaltmGluR-3 | PaltmGluR-3-F | GATTTGCCTGGGGATTTATCGC |
| PaltmGluR-3-R | GAGCCTGTATACCACCTTGAGG | |
| PaltmGluR-4 | PaltmGluR-4-F | TTTTTGCGAAGGAATGACGGTC |
| PaltmGluR-4-R | TCTGTCTGACCAACCATCACTG | |
| PaltmGluR-5 | PaltmGluR-5-F | CTTTTCTGTTGCCCGCTCTAAC |
| PaltmGluR-5-R | TTGCTTACGCCTGTTCTAAGGT | |
| PaltFZDR-1 | PaltFZDR-1-F | CCATGCAACTCAATCACAAGCA |
| PaltFZDR-1-R | TGCATGGAGGTATGGGACTATC | |
| PaltFZDR-2 | PaltFZDR-2-F | AACAAGGAGCACAAGGAAATGC |
| PaltFZDR-2-R | AGTCCTGCTTCTAGATACCAGGT | |
| PaltFZDR-3 | PaltFZDR-3-F | CCGGATTTTGGATTTGGTCTGG |
| PaltFZDR-3-R | TGCGATGCTTTTTCCCATTCTC | |
| PaltFZDR-4 | PaltFZDR-4-F | GACCTTTGTGTGATACAGTGCG |
| PaltFZDR-4-R | TGGCCCTTCCATACACATTTGA |
References
- Yang, H.; Ciais, P.; Frappart, F.; Li, X.; Brandt, M.; Fensholt, R.; Fan, L.; Saatchi, S.; Besnard, S.; Deng, Z. Global increase in biomass carbon stock dominated by growth of northern young forests over past decade. Nat. Geosci. 2023, 16, 886–892. [Google Scholar] [CrossRef]
- Zhou, J.; Du, J.; Bonifácio, L.; Yin, W.; Huang, L.; Ning, J.; Han, D.; Hu, J.; Song, W.; Zhao, L. Vulnerability of Global Pine Forestry’s Carbon Sink to an Invasive Pathogen–Vector System. Glob. Change Biol. 2024, 30, e17614. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Liu, L.; Fan, S.; Zheng, W.; Liu, R.; Zhang, Z.; Huang, R.; Zhao, L.; Shi, J. Occurrence and potential diffusion of pine wilt disease mediated by insect vectors in China under climate change. Pest. Manag. Sci. 2024, 80, 6068–6081. [Google Scholar] [CrossRef] [PubMed]
- Ryss, A.Y.; Kulinich, O.A.; Sutherland, J.R. Pine wilt disease: A short review of worldwide research. For. Stud. China 2011, 13, 132–138. [Google Scholar] [CrossRef]
- Wu, S.; Gao, Y.; Xu, X.; Wang, E.; Lei, Z. Evaluation of Stratiolaelaos scimitus and Neoseiulus barkeri for biological control of thrips on greenhouse cucumbers. Biocontrol Sci. Technol. 2014, 24, 1110–1121. [Google Scholar] [CrossRef]
- Li, L.; He, L.; Yu, L.; He, X.Z.; Xu, C.; Jiao, R.; Zhang, L.; Liu, J. Preliminary study on the potential of Pyemotes zhonghuajia (Acari: Pyemotidae) in biological control of Aphis citricola (Hemiptera: Aphididae). Syst. Appl. Acarol. 2019, 24, 1116–1120. [Google Scholar] [CrossRef]
- Xu, Y.; Li, Y.C.; Huang, B.R.; Cai, M.L.; Wu, S.Q.; Zhang, F.P.; Wu, J.Q. First record of the genus Paracarophenax (Acari: Acarophenacidae) from China, with description of a new species. Syst. Appl. Acarol. 2018, 23, 2411–2419. [Google Scholar] [CrossRef]
- Li, J.N.; Zhu, Y.Z.; Chen, R.K.; Fu, Y.; Cai, M.L.; Huang, B.R.; Xu, Y.; Wu, S.Q.; Zhang, F.P. Research on Paracarophenax alternatus Ⅰ. Biological Characteristics. Sci. Silvae Sin. 2022, 58, 79–87. [Google Scholar]
- Li, J.N.; Chen, R.K.; Fu, Y.; Cai, M.L.; Huang, B.R.; Xu, Y.; Wu, S.Q.; Zhang, F.P. Research on Paracarophenax alternatus III. Phoretic Characteristics by Monochamus alternatus Adults. Sci. Silvae Sin. 2022, 58, 128–137. [Google Scholar]
- Li, J.N.; Chen, R.K.; Fu, Y.; Cai, M.L.; Huang, B.R.; Xu, Y.; Wu, S.Q.; Zhang, F.P. Research on Paracarophenax alternatus II. Mobility and Tropism of Female Adults. Sci. Silvae Sin. 2022, 58, 149–156. [Google Scholar]
- Wu, Y.X. Olfactory Sensory Mechanism of Neoseiulus Barkeri Tocowpea Defense Responses Induced by Tetranych Usurticae Infestation. Ph.D. Thesis, Southwest University, Chongqing, China, 2023. [Google Scholar]
- Qu, S.X.; Ma, L.; Li, H.P.; Song, J.D.; Hong, X.Y. Chemosensory proteins involved in host recognition in the stored food mite Tyrophagus putrescentiae. Pest. Manag. Sci. 2015, 72, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Song, C.C. Olfactory Perception and Application of Neoseiulus californicus to Citrus Leaf Volatiles Induced by Panonychus citri. Ph.D. Thesis, Southwest University, Chongqing, China, 2024. [Google Scholar]
- Ignell, R.; Root, C.M.; Birse, R.T.; Wang, J.W.; Nässel, D.R.; Winther, A.M. Presynaptic peptidergic modulation of olfactory receptor neurons in Drosophila. Pans 2009, 106, 13070–13075. [Google Scholar] [CrossRef] [PubMed]
- Lü, S.M.; Zhao, Z.; Li, K.; Zhang, Y.L.; Xi, G.S. Cloning and expression analysis of a muscarinic cholinergic receptor from the brain of ant, Polyrhachis vicina. Arch. Insect Biochem. Physiol. 2011, 78, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, Z.; Cheng, M.; Cheng, L.; Li, S.; Lei, S.; Ding, L.; Pan, Q.; Ran, C. Identification and Expression Pattern Analysis of Octopamine Receptor Genes of Neoseiulus barkeri (Acari: Phytoseiidae). J. Southwest Univ. Nat. Sci. Ed. 2023, 45, 100–109. [Google Scholar]
- Hoy, M.A.; Yu, F.; Meyer, J.M.; Tarazona, O.A.; Jeyaprakash, A.; Wu, K. Transcriptome sequencing and annotation of the predatory mite Metaseiulus occidentalis (Acari: Phytoseiidae): A cautionary tale about possible contamination by prey sequences. Exp. Appl. Acarol. 2013, 59, 283–296. [Google Scholar] [CrossRef]
- Liu, N.; Li, T.; Wang, Y.; Liu, S. G-Protein Coupled Receptors (GPCRs) in Insects-A Potential Target for New Insecticide Development. Molecules 2021, 26, 2993. [Google Scholar] [CrossRef]
- Li, J.; Lv, H.; Li, X.; Yao, Y.; Li, J.; Ma, K. Identification and expression analysis of G protein-coupled receptors in the cotton aphid, Aphis gossypii Glover. Int. J. Biol. Macromol. 2023, 224, 115–124. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Y.; Li, T.; Feng, X. G-Protein Coupled Receptors (GPCRs): Signaling Pathways, Characterization, and Functions in Insect Physiology and Toxicology. Int. J. Mol. Sci. 2021, 22, 5260. [Google Scholar] [CrossRef]
- Veenstra, J.A.; Rombauts, S.; Grbić, M. In silico cloning of genes encoding neuropeptides, neurohormones and their putative G-protein coupled receptors in a spider mite. Insect Biochem. Mol. Biol. 2012, 42, 277–295. [Google Scholar] [CrossRef]
- Brody, T.; Cravchik, A. Drosophila melanogasterG Protein–Coupled Receptors. J. Cell Biol. 2000, 150, 83–88. [Google Scholar] [CrossRef]
- Hanlon, C.D.; Andrew, D.J. Outside-in signaling–a brief review of GPCR signaling with a focus on the Drosophila GPCR family. J. Cell Sci. 2015, 128, 3533–3542. [Google Scholar]
- Hill, C.A.; Fox, A.N.; Pitts, R.J.; Kent, L.B.; Zwiebel, L.J. G Protein-Coupled Receptors in Anopheles gambiae. Science 2002, 298, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Liu, X.; Wang, S.; Wang, Y.; Bai, L.; Peng, X.; Chen, M. A relaxin receptor gene RpGPCR41 is involved in the resistance of Rhopalosiphum padi to pyrethroids. Pestic. Biochem. Physiol. 2024, 201, 105894. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zeng, B.; Shang, T.; Zhou, S. Identification of G protein-coupled receptors required for vitellogenesis and egg development in an insect with panoistic ovary. Insect Sci. 2021, 28, 1005–1017. [Google Scholar] [CrossRef]
- Riemensperger, T.; Vller, T.; Stock, P.; Buchner, E.; Fiala, A. Punishment Prediction by Dopaminergic Neurons in Drosophila. Curr. Biol. 2005, 15, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.L.; Ma, H.T.; Wu, H.Q.; Wu, S.P.; Yan, L.Q.; Zhang, C.S.; Nur, F.A.; Wang, Z.Y.; Cao, C.W. A tachykinin receptor affects starvation tolerance and feeding behavior in the fall webworm, Hyphantria cunea. Entomol. Gen. 2022, 42, 621–630. [Google Scholar] [CrossRef]
- Shabbir, S.; Deng, M.-g.; Nawaz, M.; Lin, Q.-S. Genome-wide identification of G-protein coupled receptors (GPCRs) and their expression profile in response to β-cypermethrin stress in Zeugodacus cucurbitae. Pestic. Biochem. Physiol. 2024, 202, 105919. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Venthur, H.; Mutis, A.; Zhou, J.J.; Quiroz, A. Ligand binding and homology modelling of insect odorant-binding proteins. Physiol. Entomol. 2014, 39, 183–198. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Fu, X.B.; Cui, H.C.; Zhao, L.; Yu, J.Z.; Li, H.L. Functional characteristics, electrophysiological and antennal immunolocalization of general odorant-binding protein 2 in tea geometrid, Ectropis obliqua. Int. J. Mol. Sci. 2018, 19, 875. [Google Scholar] [CrossRef]
- Anstey, M.L.; Rogers, S.M.; Ott, S.R.; Burrows, M.; Simpson, S.J. Serotonin mediates behavioral gregarization underlying swarm formation in desert locusts. Science 2009, 323, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Winther, Å.M.; Acebes, A.; Ferrús, A. Tachykinin-related peptides modulate odor perception and locomotor activity in Drosophila. Mol. Cell. Neurosci. 2006, 31, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chen, H.; He, J.; Zeng, X.; Lei, H.; Liu, J. Dual roles of dopaminergic pathways in olfactory learning and memory in the oriental fruit fly, Bactrocera dorsalis. Pestic. Biochem. Physiol. 2024, 200, 105825. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.Q.; Linghu, J.H.; Xie, H.Y.; Li, G.; Zhu, F.; Smagghe, G.; Gui, S.H.; Liu, T.X. Characterization of sulfakinin and its role in larval feeding and molting in Spodoptera frugiperda. Insect Sci. 2024, 1–13. [Google Scholar] [CrossRef]
- Brigaud, I.; Grosmaître, X.; François, M.-C.; Jacquin-Joly, E. Cloning and expression pattern of a putative octopamine/tyramine receptor in antennae of the noctuid moth Mamestra brassicae. Cell Tissue Res. 2009, 335, 455–463. [Google Scholar] [CrossRef]
- Gao, H.; Tian, Y.; Zhang, H.; Li, Y.; Li, C.; Li, B. Species-Specific Duplicated FMRFaR-like Gene A62 Regulates Spontaneous Locomotion in Apolygus lucorum. Pest. Manag. Sci. 2024, 80, 3358–3368. [Google Scholar] [CrossRef]
- Lenz, O.; Xiong, J.; Nelson, M.D.; Raizen, D.M.; Williams, J.A. FMRFamide signaling promotes stress-induced sleep in Drosophila. Brain Behav. Immun. 2015, 47, 141–148. [Google Scholar] [CrossRef]
- Katanaev, V.L.; Ponzielli, R.; Sémériva, M.; Tomlinson, A. Trimeric G protein-dependent frizzled signaling in Drosophila. Cell 2005, 120, 111–122. [Google Scholar] [CrossRef]
- He, X.; Zang, J.; Li, X.; Shao, J.; Yang, H.; Yang, J.; Huang, H.; Chen, L.; Shi, L.; Zhu, C. Activation of BNGR-A24 by direct interaction with tachykinin-related peptides from the silkworm Bombyx mori leads to the Gq-and Gs-coupled signaling cascades. Biochemistry 2014, 53, 6667–6678. [Google Scholar] [CrossRef]
- Kamareddine, L.; Robins, W.P.; Berkey, C.D.; Mekalanos, J.J.; Watnick, P.I. The Drosophila immune deficiency pathway modulates enteroendocrine function and host metabolism. Cell Metab. 2018, 28, 449–462.e5. [Google Scholar] [CrossRef]
- Chowański, S.; Pacholska-Bogalska, J.; Rosiński, G. Cholinergic agonists and antagonists have an effect on the metabolism of the beetle Tenebrio molitor. Molecules 2018, 24, 17. [Google Scholar] [CrossRef] [PubMed]
- Futahashi, R.; Kawahara-Miki, R.; Kinoshita, M.; Yoshitake, K.; Yajima, S.; Arikawa, K.; Fukatsu, T. Extraordinary diversity of visual opsin genes in dragonflies. Proc. Natl. Acad. Sci. USA 2015, 112, E1247–E1256. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Liu, X.; Liu, J.; Xiong, W.; Song, X.; Wu, W.; Wei, L.; Li, B. Identification and evolution of latrophilin receptor gene involved in Tribolium castaneum devolopment and female fecundity. Genesis 2017, 55, e23081. [Google Scholar] [CrossRef] [PubMed]
- Seeman, O.D.; Walter, D.E. Phoresy and Mites: More Than Just a Free Ride. Annu. Rev. Entomol. 2023, 68, 69–88. [Google Scholar] [CrossRef]
- Birse, R.T.; Söderberg, J.A.; Luo, J.; Winther, Å.M.; Nässel, D.R. Regulation of insulin-producing cells in the adult Drosophila brain via the tachykinin peptide receptor DTKR. J. Exp. Biol. 2011, 214, 4201–4208. [Google Scholar] [CrossRef]
- Söderberg, J.A.; Birse, R.T.; Nässel, D.R. Insulin production and signaling in renal tubules of Drosophila is under control of tachykinin-related peptide and regulates stress resistance. PLoS ONE 2011, 6, e19866. [Google Scholar] [CrossRef]
- Nian, X.; Wang, B.; Holford, P.; Beattie, G.A.C.; Tan, S.; Yuan, W.; Cen, Y.; He, Y.; Zhang, S. Neuropeptide Ecdysis-Triggering Hormone and Its Receptor Mediate the Fecundity Improvement of ‘Candidatus Liberibacter Asiaticus’-Infected Diaphorina citri Females and CLas Proliferation. Adv. Sci. 2025, 12, 2412384. [Google Scholar] [CrossRef]
- Huang, J.; Wu, S.-F.; Li, X.-H.; Adamo, S.A.; Ye, G.-Y. The characterization of a concentration-sensitive α-adrenergic-like octopamine receptor found on insect immune cells and its possible role in mediating stress hormone effects on immune function. Brain Behav. Immun. 2012, 26, 942–950. [Google Scholar] [CrossRef]
- Nässel, D.R.; Wu, S.F. Cholecystokinin/sulfakinin peptide signaling: Conserved roles at the intersection between feeding, mating and aggression. Cell. Mol. Life Sci. 2022, 79, 188. [Google Scholar] [CrossRef]
- Wu, S.; Guo, C.; Zhao, H.; Sun, M.; Chen, J.; Han, C.; Peng, Q.; Qiao, H.; Peng, P.; Liu, Y.; et al. Drosulfakinin signaling in fruitless circuitry antagonizes P1 neurons to regulate sexual arousal in Drosophila. Nat. Commun. 2019, 10, 4770. [Google Scholar] [CrossRef]
- Nichols, R.; Egle, J.P.; Langan, N.R.; Palmer, G.C. The different effects of structurally related sulfakinins on Drosophila melanogaster odor preference and locomotion suggest involvement of distinct mechanisms. Peptides 2008, 29, 2128–2135. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Jiang, Y.; Chen, S.; Chen, F.; Chen, F. Herbivory-Induced Emission of Volatile Terpenes in Chrysanthemum morifolium Functions as an Indirect Defense against Spodoptera litura Larvae by Attracting Natural Enemies. J. Agric. Food Chem. 2021, 69, 9743–9753. [Google Scholar] [CrossRef]
- Wang, S.; Han, M.; Li, H.; Xie, J.; Wei, K.; Wang, X. Testing the host range of Cyanopterus ninghais (Hymenoptera: Braconidae), a candidate for the biological control of Monochamus alternatus, the vector of pine wilt disease in Asia. Biol. Control 2024, 195, 105547. [Google Scholar] [CrossRef]
- Chen, R.; He, X.; Chen, J.; Gu, T.; Liu, P.; Xu, T.; Teale, S.A.; Hao, D. Traumatic resin duct development, terpenoid formation, and related synthase gene expression in Pinus massoniana under feeding pressure of Monochamus alternatus. Plant Growth Regul. 2019, 38, 897–908. [Google Scholar] [CrossRef]
- Mattila, T.M.; Bechsgaard, J.S.; Hansen, T.T.; Schierup, M.H.; Bilde, T. Orthologous genes identified by transcriptome sequencing in the spider genus Stegodyphus. BMC Genom. 2012, 13, 70. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Aron, M.B.; Yu, B.; Lianyi, H.; Jane, H.; Lanczycki, C.J.; Shennan, L.; Farideh, C.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar]
- Lam-Tung, N.; Schmidt, H.A.; Arndt, V.H.; Quang, M.B. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.V.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]






| Receptor No. in the Text | Accession Number | Length (aa) | Receptor No. in the Text | Accession Number | Length (aa) |
|---|---|---|---|---|---|
| Family A | PaltTKR-6 | XP_015794716.1 | 806 | ||
| PaltRh2-1 | RWS15172.1 | 775 | PaltTKR-7 | XP_003737058.1 | 166 |
| PaltRh2-2 | RWS03304.1 | 402 | PaltTKR-8 | RWS16120.1 | 649 |
| PaltOR-1 | RWS28020.1 | 88 | PaltSKR-1 | RWS08800.1 | 543 |
| PaltOR-2 | RWS04419.1 | 359 | PaltCXCR4-1 | XP_015795307.1 | 351 |
| PaltTAR-1 | RWS23218.1 | 1462 | PaltRyaR-1 | XP_015785080.1 | 553 |
| PaltTAR-2 | XP_003741345.1 | 152 | PaltAKHR-1 | RWS20534.1 | 393 |
| PaltTAR-3 | RWS23218.1 | 840 | PaltAKHR-2 | XP_053206171.1 | 465 |
| PaltTAR-4 | RWS06560.1 | 288 | PaltAKHR-3 | XP_053210923.1 | 334 |
| PaltDopR-1 | XP_028969126.1 | 187 | PaltRXFP-1 | RWS24200.1 | 605 |
| PaltDopR-2 | RWS31985.1 | 1282 | PaltADR-1 | RWS24614.1 | 1655 |
| PaltDopR-3 | XP_028969125.1 | 862 | PaltADR-2 | RWS29702.1 | 412 |
| PaltDopR-4 | OQR76146.1 | 560 | PaltNPYR-1 | XP_015792452.1 | 455 |
| PaltDopR-5 | RWS11398.1 | 482 | PaltNPYR-2 | OQR71898.1 | 342 |
| PaltOAR-1 | XP_015785358.1 | 939 | PaltNPYR-3 | XP_015792844.1 | 696 |
| PaltOAR-2 | XP_053214250.1 | 153 | PaltAT2R-1 | RWS08012.1 | 359 |
| PaltOAR-3 | XP_028966328.1 | 311 | PaltAT2R-2 | RWS13054.1 | 100 |
| PaltmAChR-1 | RWS02783.1 | 129 | PaltADOR-1 | RWS07176.1 | 601 |
| PaltmAChR-2 | XP_003747881.1 | 1256 | PaltADOR-2 | RWS06158.1 | 444 |
| PaltmAChR-3 | XP_053206256.1 | 240 | PaltADOR-3 | RWS20623.1 | 560 |
| PaltmAChR-4 | XP_025016167.1 | 654 | Family B | ||
| PaltmAChR-5 | OQR68484.1 | 401 | PaltCTR-1 | XP_015788638.1 | 660 |
| Palt5-HTR-1 | RWS30558.1 | 733 | PaltCTR-2 | XP_053207328.1 | 151 |
| Palt5-HTR-2 | XP_015781282.1 | 723 | PaltCTR-3 | RWS26867.1 | 124 |
| Palt5-HTR-3 | XP_022654876.1 | 118 | PaltCTR-4 | XP_015789508.1 | 961 |
| Palt5-HTR-4 | XP_003742827.1 | 1481 | PaltCTR-5 | XP_022663537.1 | 208 |
| Palt5-HTR-5 | RWS07087.1 | 674 | PaltCTR-6 | XP_022645609.1 | 1625 |
| Palt5-HTR-6 | RWS01106.1 | 624 | PaltaGPCR-1 | XP_015788882.1 | 944 |
| Palt5-HTR-7 | XP_053203942.1 | 283 | PaltTcLph-1 | XP_053202594.1 | 1425 |
| Palt5-HTR-8 | RWS27206.1 | 166 | PaltTcLph-2 | XP_028966344.1 | 1665 |
| Palt5-HTR-9 | XP_053207194.1 | 685 | PaltTcLph-3 | XP_022656988.1 | 1612 |
| PaltETHR-1 | AZL90164.1 | 928 | PaltTcLph-4 | XP_022656975.1 | 1460 |
| PaltPKR-1 | XP_053210255.1 | 680 | PaltTcLph-5 | XP_053202593.1 | 1166 |
| PaltPKR-2 | XP_053207649.1 | 173 | Family C | ||
| PaltFMRFa-1 | RWS24014.1 | 343 | PaltmGluR-1 | RWS08505.1 | 221 |
| PaltFMRFa-2 | XP_022663222.1 | 888 | PaltmGluR-2 | XP_053202653.1 | 1309 |
| PaltFMRFa-3 | RWS28231.1 | 653 | PaltmGluR-3 | RWS26253.1 | 1075 |
| PaltFMRFa-4 | RWS31476.1 | 618 | PaltmGluR-4 | RWS26411.1 | 1949 |
| PaltFMRFa-5 | OQR77613.1 | 779 | PaltmGluR-5 | XP_015793762.1 | 193 |
| PaltAstAR-1 | XP_015789363.1 | 614 | Family D | ||
| PaltTKR-1 | RWS14979.1 | 597 | PaltFZDR-1 | XP_015786657.1 | 572 |
| PaltTKR-2 | RWS09339.1 | 532 | PaltFZDR-2 | XP_053209274.1 | 544 |
| PaltTKR-3 | XP_015790474.1 | 518 | PaltFZDR-3 | XP_015790096.1 | 886 |
| PaltTKR-4 | RWS27691.1 | 159 | PaltFZDR-4 | XP_003739708.2 | 660 |
| PaltTKR-5 | XP_053211289.1 | 853 |
| α-Pinene | β-Pinene | Longifolene | β-Caryophyllene | β-Phellandrene | |
|---|---|---|---|---|---|
| PaltDopR-5 | −4.45 | −4.56 | −5.17 | −5.37 | −4.38 |
| PaltOR-1 | −4.37 | −4.43 | −5.62 | −5.19 | −5.83 |
| PaltOR-2 | −4.38 | −4.38 | −5.15 | −5.17 | −4.68 |
| PaltSKR-1 | −5.06 | −5.22 | −5.73 | −5.35 | −4.38 |
| PaltTAR-3 | −4.14 | −4.05 | −4.69 | −4.99 | −4.11 |
| PaltTKR-2 | −4.70 | −5.00 | −5.35 | −5.80 | −4.72 |
| Compounds | Molecular Formula | Structural Formula | CAS NO. |
|---|---|---|---|
| α-Pinene | C10H16 |  | 80-56-8 |
| β-pinene | C10H16 |  | 127-91-3 |
| Longifolene | C15H24 |  | 475-20-7 |
| β-Caryophyllene | C15H24 |  | 87-44-5 |
| β-Phellandrene | C10H16 |  | 99-83-2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, R.; Chu, X.; Zhang, Y.; Ke, S.; Zheng, Y.; Yu, W.; Zhang, F.; Wu, S. Identification and Expression Analysis of G-Protein-Coupled Receptors Provide Insights into Functional and Mechanistic Responses to Herbivore-Induced Plant Volatiles of Paracarophenax alternatus. Int. J. Mol. Sci. 2025, 26, 5890. https://doi.org/10.3390/ijms26125890
Lin R, Chu X, Zhang Y, Ke S, Zheng Y, Yu W, Zhang F, Wu S. Identification and Expression Analysis of G-Protein-Coupled Receptors Provide Insights into Functional and Mechanistic Responses to Herbivore-Induced Plant Volatiles of Paracarophenax alternatus. International Journal of Molecular Sciences. 2025; 26(12):5890. https://doi.org/10.3390/ijms26125890
Chicago/Turabian StyleLin, Ruiheng, Xu Chu, Yangming Zhang, Sikai Ke, Yunfeng Zheng, Wei Yu, Feiping Zhang, and Songqing Wu. 2025. "Identification and Expression Analysis of G-Protein-Coupled Receptors Provide Insights into Functional and Mechanistic Responses to Herbivore-Induced Plant Volatiles of Paracarophenax alternatus" International Journal of Molecular Sciences 26, no. 12: 5890. https://doi.org/10.3390/ijms26125890
APA StyleLin, R., Chu, X., Zhang, Y., Ke, S., Zheng, Y., Yu, W., Zhang, F., & Wu, S. (2025). Identification and Expression Analysis of G-Protein-Coupled Receptors Provide Insights into Functional and Mechanistic Responses to Herbivore-Induced Plant Volatiles of Paracarophenax alternatus. International Journal of Molecular Sciences, 26(12), 5890. https://doi.org/10.3390/ijms26125890