
Functional Characterization of Two β-Hexosaminidase A Isoforms During Ovarian Development in Macrobrachium nipponense

 , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Full-Length Sequence Analysis of Mn-HexA1 and Mn-HexA2
2.2. Similarity Comparison and Phylogenetic Analysis
2.3. Expression Analysis of Mn-HexA1 and Mn-HexA2
2.3.1. Expression Analysis of Mn-HexA1 and Mn-HexA2 at Different Developmental Stages
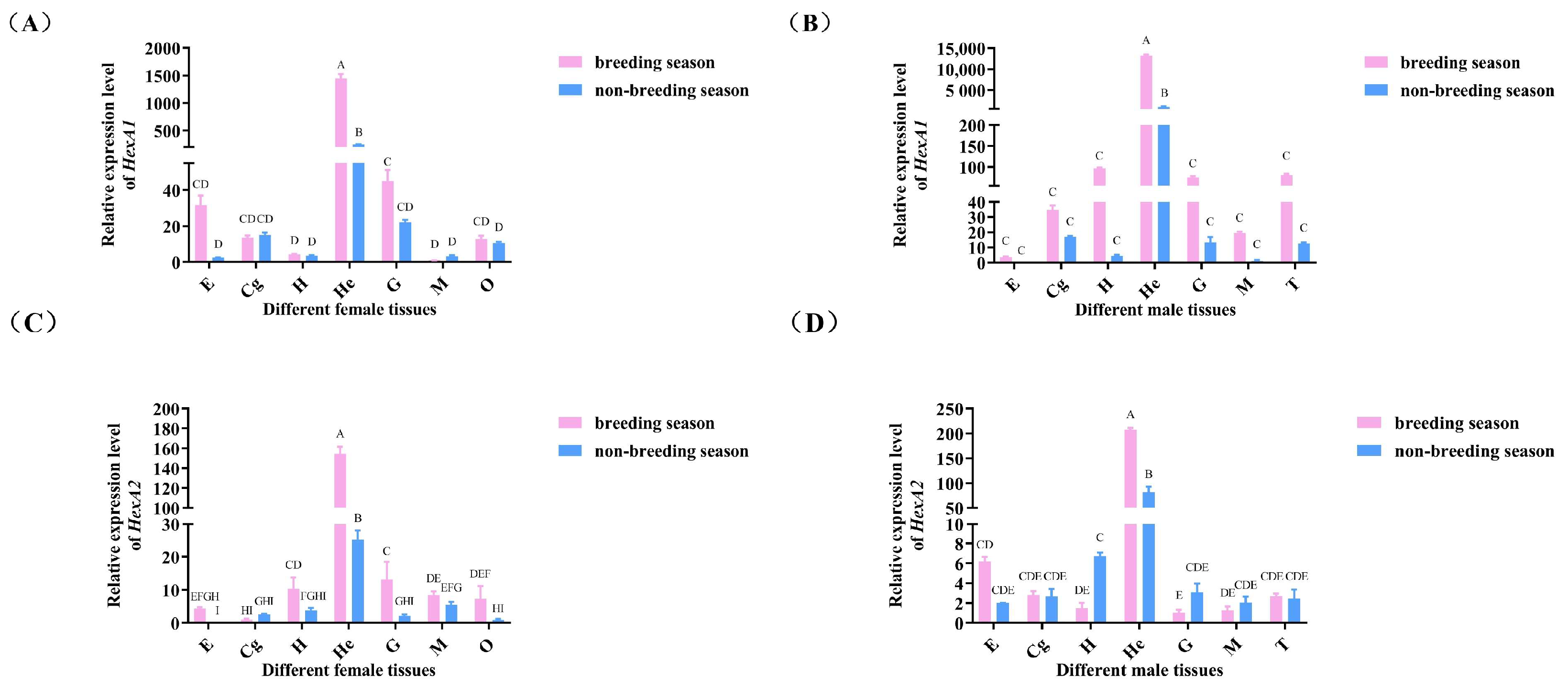
2.3.2. Expression Analysis of Mn-HexA1 and Mn-HexA2 in Different Tissue
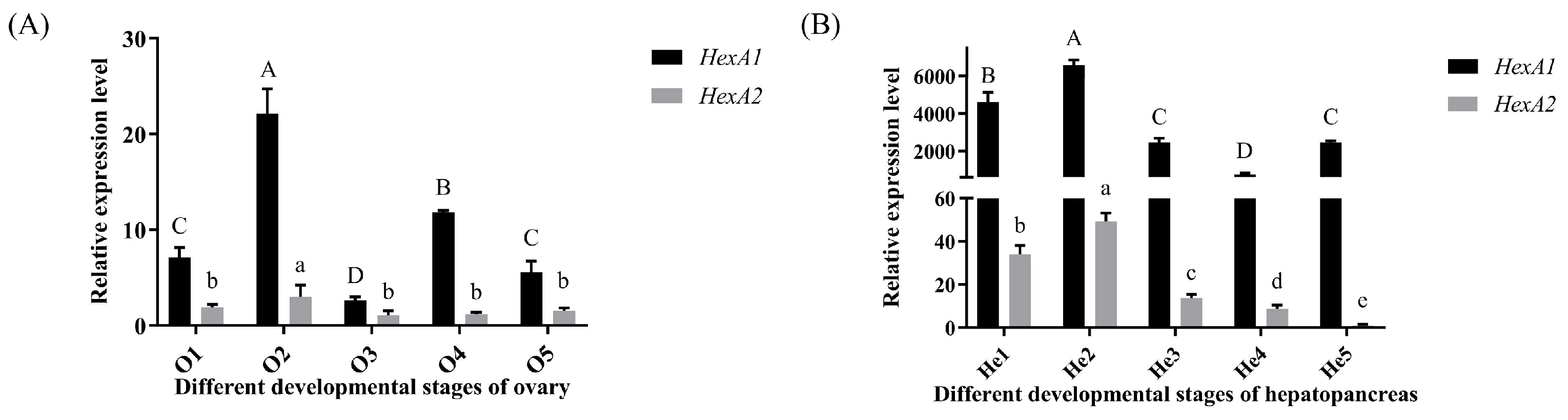
2.3.3. Expression Analysis of Mn-HexA1 and Mn-HexA2 in Hepatopancreas and Ovaries at Different Stages
2.4. Localization of Mn-HexA1 and Mn-HexA2 in Different Tissues
2.5. Functional Analysis of Mn-HexA1 and Mn-HexA2
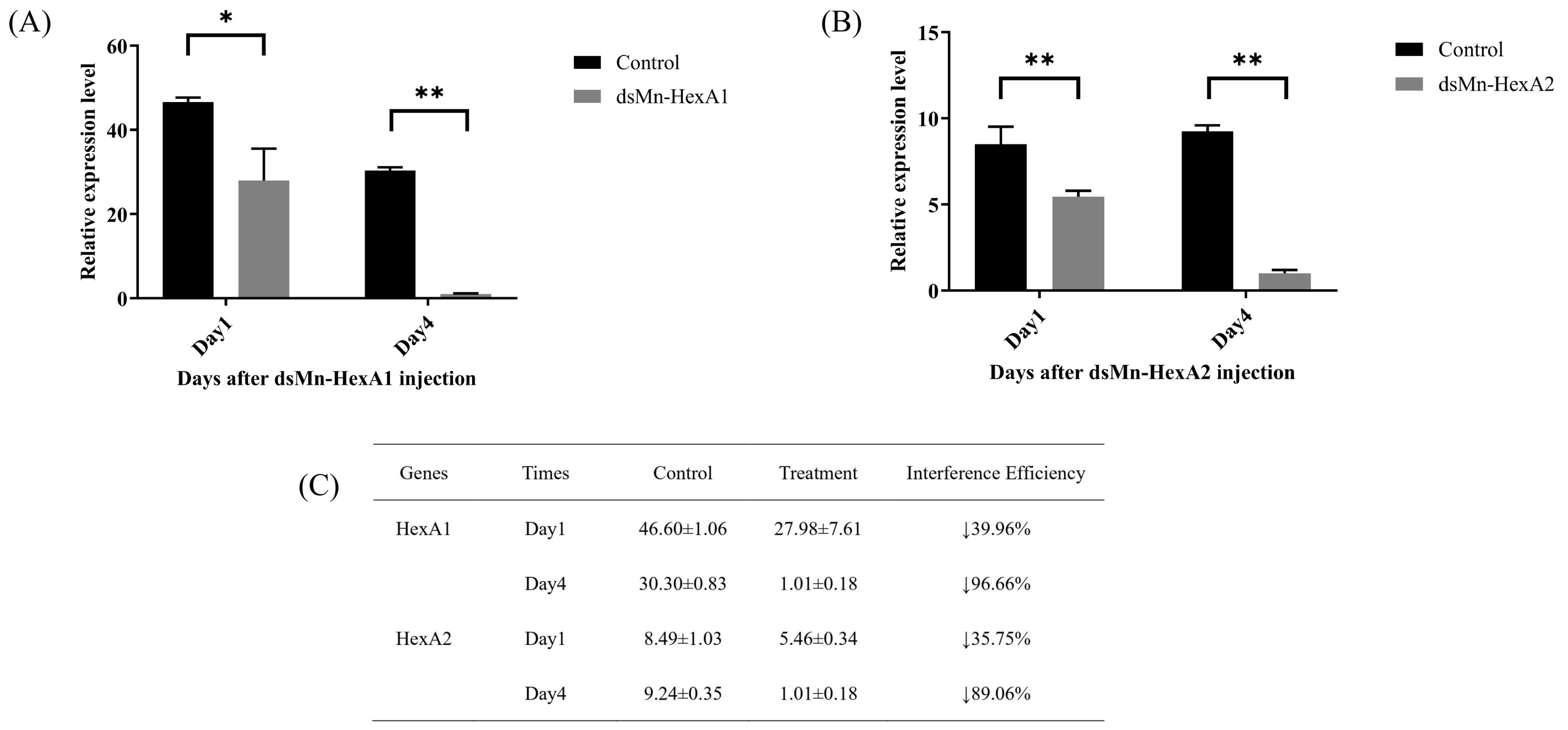
2.5.1. Interference Efficiency
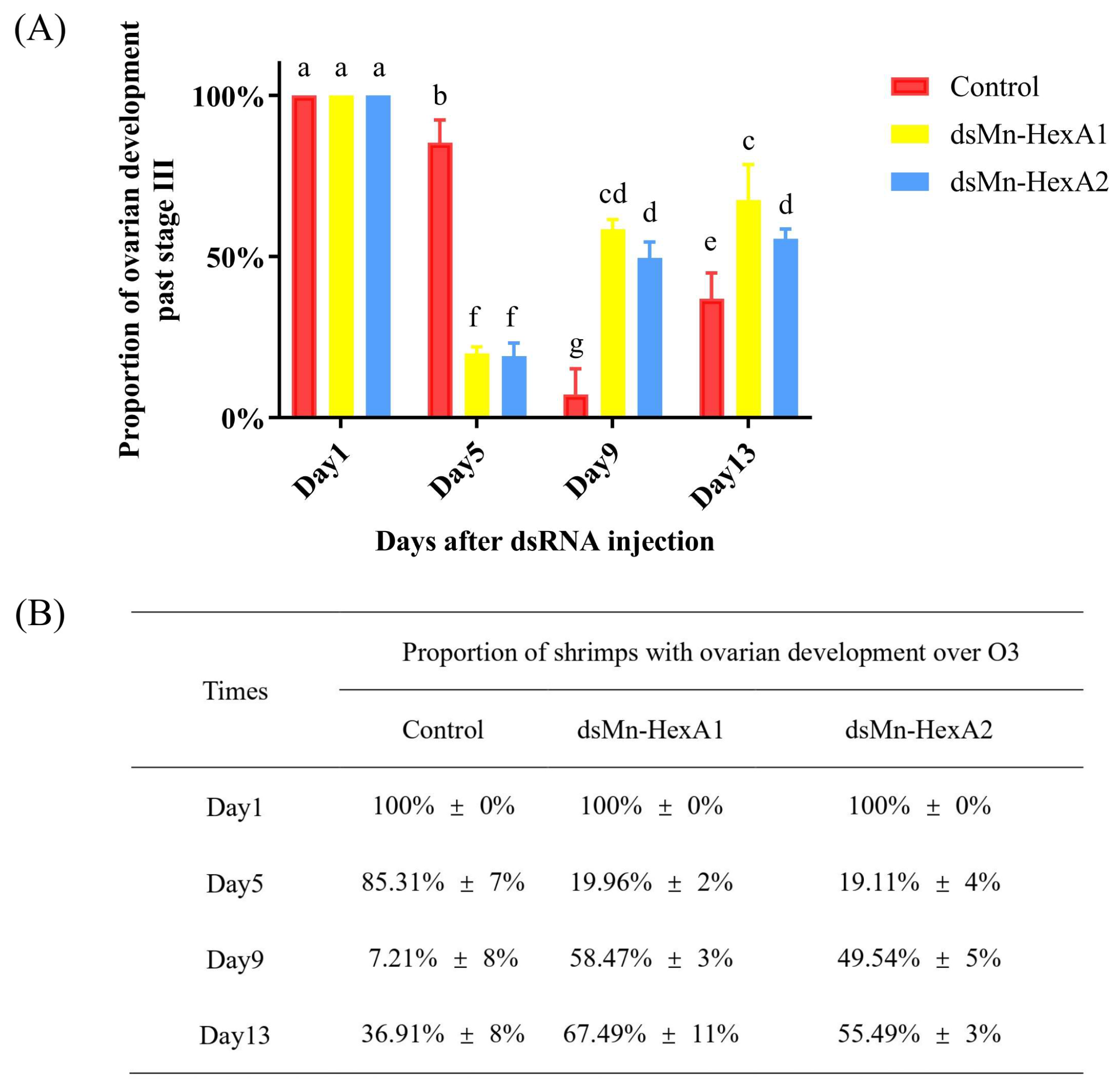
2.5.2. Effect of Mn-HexA1 and Mn-HexA2 Knockdown on Ovarian Development of M. nipponense
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Breeding Conditions
4.2. Tissue Sample Collection
4.3. Genes Cloning
4.4. Bioinformatics Analysis
4.5. In Situ Hybridization
4.6. RNAi Experiment
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, G.; Xue, Y.; Huang, X. Temperature-Induced Sex Differentiation in River Prawn (Macrobrachium nipponense): Mechanisms and Effects. Int. J. Mol. Sci. 2024, 25, 1207. [Google Scholar] [CrossRef]
- Jiang, H.; Liu, X.; Li, Y.; Zhang, R.; Liu, H.; Ma, X.; Wu, L.; Qiao, Z.; Li, X. Identification of Ribosomal Protein L24 (RPL24) from the Oriental River Prawn, Macrobrachium nipponense, and Its Roles in Ovarian Development. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 266, 111154. [Google Scholar] [CrossRef]
- Qiao, H.; Fu, H.; Xiong, Y.; Jiang, S.; Zhang, W.; Sun, S.; Jin, S.; Gong, Y.; Wang, Y.; Shan, D.; et al. Molecular Insights into Reproduction Regulation of Female Oriental River Prawns Macrobrachium nipponense through Comparative Transcriptomic Analysis. Sci. Rep. 2017, 7, 12161. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Li, X.; Li, Y.; Liu, X.; Zhang, S.; Li, H.; Zhang, M.; Wang, L.; Yu, M.; Qiao, Z. Molecular and Functional Characterization of Ribosome Protein S24 in Ovarian Development of Macrobrachium nipponense. Int. J. Biol. Macromol. 2024, 254, 127934. [Google Scholar] [CrossRef] [PubMed]
- Fatima, H.; Ayub, Z.; Ali, S.; Siddiqui, G. Biochemical Composition of the Hemolymph, Hepatopancreas, Ovary, and Muscle during Ovarian Maturation in the Penaeid Shrimps Fenneropenaeus Merguiensis and F. penicillatus (Crustacea: Decapoda). Turk. J. Zool. 2013, 37, 334–347. [Google Scholar] [CrossRef]
- Feng, W.; Zhao, Z.; Wang, J.; Han, T. Nutrient Composition of Ovary, Hepatopancreas and Muscle Tissues in Relation to Ovarian Development Stage of Female Swimming Crab, Portunus Trituberculatus. Animals 2023, 13, 3220. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, W.; Xiong, Y.; Cheng, D.; Wang, J.; Jin, S.; Gong, Y.; Wu, Y.; Qiao, H.; Fu, H. Hepatopancreas Transcriptome Analyses Provide New Insights into the Molecular Regulatory Mechanism of Fast Ovary Maturation in Macrobrachium nipponense. BMC Genom. 2022, 23, 625. [Google Scholar] [CrossRef]
- Cheng, D.; Zhang, W.; Jiang, S.; Xiong, Y.; Jin, S.; Pan, F.; Zhu, J.; Gong, Y.; Wu, Y.; Qiao, H.; et al. Cathepsin D Plays a Vital Role in Macrobrachium nipponense of Ovary Maturation: Identification, Characterization, and Function Analysis. Genes 2022, 13, 1495. [Google Scholar] [CrossRef]
- Menendez-Helman, R.J.; Sanjurjo, C.; Miranda, P.V. Seminal Plasma Hexosaminidase in Patients with Normal and Abnormal Spermograms. Iran. J. Reprod. Med. 2015, 13, 541–548. [Google Scholar]
- Szczubiał, M.; Kankofer, M.; Wawrzykowski, J.; Dąbrowski, R.; Bochniarz, M.; Brodzki, P. Activity of the Glycosidases β-Galactosidase, α-l-Fucosidase, β-N-Acetyl-Hexosaminidase, and Sialidase in Uterine Tissues from Female Dogs in Diestrus with and without Pyometra. Theriogenology 2022, 177, 133–139. [Google Scholar] [CrossRef]
- Intra, J.; Cenni, F.; Pavesi, G.; Pasini, M.; Perotti, M.-E. Interspecific Analysis of the Glycosidases of the Sperm Plasma Membrane in Drosophila. Mol. Reprod. Dev. 2009, 76, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, F.; Pasini, M.E.; Intra, J.; Matsumoto, M.; Briani, F.; Hoshi, M.; Perotti, M.E. Identification and Expression Analysis of Drosophilamelanogaster Genes Encoding β-Hexosaminidases of the Sperm Plasma Membrane. Glycobiology 2006, 16, 786–800. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, F.; Ogiso, M.; Hoshi, M.; Perotti, M.-E.; Pasini, M.E. Purification and Characterization of the Plasma Membrane Glycosidases of Drosophila melanogaster Spermatozoa. Insect Biochem. Mol. Biol. 2002, 32, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Perez Martinez, S.L.; Menendez Helman, R.J.; Zitta, K.S.; Brandelli, A.; Miranda, P.V. Characterization of Human Sperm N-Acetylglucosaminidase. Int. J. Androl. 2008, 31, 315–324. [Google Scholar] [CrossRef]
- Zitta, K.; Wertheimer, E.V.; Miranda, P.V. Sperm N-Acetylglucosaminidase Is Involved in Primary Binding to the Zona Pellucida. Mol. Hum. Reprod. 2006, 12, 557–563. [Google Scholar] [CrossRef]
- Koyanagi, R.; Honegger, T.G. Molecular Cloning and Sequence Analysis of an Ascidian Egg β-N-Acetylhexosaminidase with a Potential Role in Fertilization. Dev. Growth Differ. 2003, 45, 209–218. [Google Scholar] [CrossRef]
- Intra, J.; Pavesi, G.; Horner, D.S. Phylogenetic Analyses Suggest Multiple Changes of Substrate Specificity within the Glycosyl Hydrolase 20 Family. BMC Evol. Biol. 2008, 8, 214. [Google Scholar] [CrossRef]
- Venugopal, A.; Mondal, S.; Ranganatha, K.S.; Datta, D.; Kumar, N.S.; Swamy, M.J. Purification and Biochemical/Biophysical Characterization of Two Hexosaminidases from the Fresh Water Mussel, Lamellidens Corrianus. Int. J. Biol. Macromol. 2020, 149, 754–766. [Google Scholar] [CrossRef]
- Wendeler, M.; Sandhoff, K. Hexosaminidase Assays. Glycoconj. J. 2009, 26, 945–952. [Google Scholar] [CrossRef]
- Mark, B.L.; Mahuran, D.J.; Cherney, M.M.; Zhao, D.; Knapp, S.; James, M.N.G. Crystal Structure of Human β-Hexosaminidase B: Understanding the Molecular Basis of Sandhoff and Tay–Sachs Disease. J. Mol. Biol. 2003, 327, 1093–1109. [Google Scholar] [CrossRef]
- Mahuran, D.J. Biochemical Consequences of Mutations Causing the GM2 Gangliosidoses. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 1999, 1455, 105–138. [Google Scholar] [CrossRef]
- Lemieux, M.J.; Mark, B.L.; Cherney, M.M.; Withers, S.G.; Mahuran, D.J.; James, M.N.G. Crystallographic Structure of Human β-Hexosaminidase A: Interpretation of Tay-Sachs Mutations and Loss of GM2 Ganglioside Hydrolysis. J. Mol. Biol. 2006, 359, 913–929. [Google Scholar] [CrossRef] [PubMed]
- Calzoni, E.; Cerrotti, G.; Sagini, K.; Delo, F.; Buratta, S.; Pellegrino, R.M.; Alabed, H.B.R.; Fratini, F.; Emiliani, C.; Urbanelli, L. Evidence of Lysosomal β-Hexosaminidase Enzymatic Activity Associated with Extracellular Vesicles: Potential Applications for the Correction of Sandhoff Disease. J. Funct. Biomater. 2024, 15, 153. [Google Scholar] [CrossRef]
- Lee, J.; Boo, J.; Kim, Y.-H.; Roh, J.; Ko, S.-K.; Shin, I. A Fluorescent Probe for Selective Detection of Lysosomal β-Hexosaminidase in Live Cells. Talanta 2024, 271, 125715. [Google Scholar] [CrossRef]
- Olszewska, E.; Borzym-Kluczyk, M.; Rzewnicki, I.; Rutkowska, J.; Knas, M.; Rogowski, M.; Waniewska, E.; Wielgosz, R. Hexosaminidase as a New Potential Marker for Larynx Cancer. Clin. Biochem. 2009, 42, 1187–1189. [Google Scholar] [CrossRef]
- Plucinsky, M.C.; Prorok, J.J.; Alhadeff, J.A. β-Hexosaminidase From Colon and Sera of Dukes-Classified Colorectal Cancer Patients: Activity Levels, Isozyme Patterns, and Kinetic Properties2. JNCI J. Natl. Cancer Inst. 1986, 77, 57–62. [Google Scholar] [CrossRef]
- Samarasinghe, A.E.; Penkert, R.R.; Hurwitz, J.L.; Sealy, R.E.; LeMessurier, K.S.; Hammond, C.; Dubin, P.J.; Lew, D.B. Questioning Cause and Effect: Children with Severe Asthma Exhibit High Levels of Inflammatory Biomarkers Including Beta-Hexosaminidase, but Low Levels of Vitamin A and Immunoglobulins. Biomedicines 2020, 8, 393. [Google Scholar] [CrossRef]
- Fukuishi, N.; Murakami, S.; Ohno, A.; Yamanaka, N.; Matsui, N.; Fukutsuji, K.; Yamada, S.; Itoh, K.; Akagi, M. Does β-Hexosaminidase Function Only as a Degranulation Indicator in Mast Cells? The Primary Role of β-Hexosaminidase in Mast Cell Granules. J. Immunol. 2014, 193, 1886–1894. [Google Scholar] [CrossRef]
- Emiliani, C.; Ciferri, S.; Mencarelli, S.; Mezzasoma, A.M.; Momi, S.; Orlacchio, A.; Gresele, P. Defective Platelet β-N-Acetyl Hexosaminidase Content and Release in Chronic Myeloproliferative Disorders. Platelets 2006, 17, 20–29. [Google Scholar] [CrossRef]
- Strasser, R.; Bondili, J.S.; Schoberer, J.; Svoboda, B.; Liebminger, E.; Glössl, J.; Altmann, F.; Steinkellner, H.; Mach, L. Enzymatic Properties and Subcellular Localization of Arabidopsis β-N-Acetylhexosaminidases. Plant Physiol. 2007, 145, 5–16. [Google Scholar] [CrossRef]
- Oikawa, A.; Itoh, E.; Ishihara, A.; Iwamura, H. Purification and Characterization of β-N-Acetylhexosaminidase from Maize Seedlings. J. Plant Physiol. 2003, 160, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Tsujibo, H.; Miyamoto, K.; Yoshimura, M.; Takata, M.; Miyamoto, J.; Inamori, Y. Molecular Cloning of the Gene Encoding a Novel β-N-Acetylhexosaminidase from a Marine Bacterium, Alteromonas sp. Strain O-7, and Characterization of the Cloned Enzyme. Biosci. Biotechnol. Biochem. 2002, 66, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Horsch, M.; Mayer, C.; Sennhauser, U.; Rast, D.M. β-N-Acetylhexosaminidase: A Target for the Design of Antifungal Agents. Pharmacol. Ther. 1997, 76, 187–218. [Google Scholar] [CrossRef] [PubMed]
- Tomiya, N.; Narang, S.; Park, J.; Abdul-Rahman, B.; Choi, O.; Singh, S.; Hiratake, J.; Sakata, K.; Betenbaugh, M.J.; Palter, K.B.; et al. Purification, Characterization, and Cloning of a Spodoptera Frugiperda Sf9 β-N-Acetylhexosaminidase That Hydrolyzes Terminal N-Acetylglucosamine on the N-Glycan Core*. J. Biol. Chem. 2006, 281, 19545–19560. [Google Scholar] [CrossRef]
- Hogenkamp, D.G.; Arakane, Y.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Characterization and Expression of the β-N-Acetylhexosaminidase Gene Family of Tribolium Castaneum. Insect Biochem. Mol. Biol. 2008, 38, 478–489. [Google Scholar] [CrossRef]
- Guo, P.-P.; Yang, X.-B.; Yang, H.; Zhou, C.; Long, G.-Y.; Jin, D.-C. Knockdown of the β-N-Acetylhexosaminidase Genes by RNA Interference Inhibited the Molting and Increased the Mortality of the White-Backed Planthopper, Sogatella furcifera. Pestic. Biochem. Physiol. 2025, 207, 106216. [Google Scholar] [CrossRef]
- Jiang, X.; Yang, Q. Recent Advances in Glycoside Hydrolase Family 20 and 84 Inhibitors: Structures, Inhibitory Mechanisms and Biological Activities. Bioorganic Chem. 2024, 142, 106870. [Google Scholar] [CrossRef]
- Shi, W.; Hu, R.; Zhao, R.; Zhu, J.; Shen, H.; Li, H.; Wang, L.; Yang, Z.; Jiang, Q.; Qiao, Y.; et al. Transcriptome Analysis of Hepatopancreas and Gills of Palaemon gravieri under Salinity Stress. Gene 2023, 851, 147013. [Google Scholar] [CrossRef]
- Jiang, S.; Xiong, Y.; Zhang, W.; Zhu, J.; Cheng, D.; Gong, Y.; Wu, Y.; Qiao, H.; Fu, H. Molecular Characterization of a Novel Cathepsin L in Macrobrachium nipponense and Its Function in Ovary Maturation. Front. Endocrinol. 2022, 12, 816813. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, W.; Xiong, Y.; Zhang, M.; Yuan, H.; Niu, Y.; Qiao, H.; Fu, H. NPC Intracellular Cholesterol Transporter 1 Regulates Ovarian Maturation and Molting in Female Macrobrachium nipponense. Int. J. Mol. Sci. 2024, 25, 6049. [Google Scholar] [CrossRef]
- Cai, P.; Zhang, W.; Jiang, S.; Xiong, Y.; Qiao, H.; Yuan, H.; Gao, Z.; Zhou, Y.; Jin, S.; Fu, H. Role of Mn-LIPA in Sex Hormone Regulation and Gonadal Development in the Oriental River Prawn, Macrobrachium nipponense. Int. J. Mol. Sci. 2024, 25, 1399. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Andersen, C.L.; Ye, X. Functions of Lysosomes in Mammalian Female Reproductive System. Reprod. Dev. Med. 2020, 4, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilian, Y.; Hela, F.; Bildik, G.; İltumur, E.; Yusufoglu, S.; Yakin, K.; Oktem, O. Discovery of Autophagy as a Universal Mechanism for Sex Steroid Synthesis in Human Ovary and Testis. Autophagy Rep. 2023, 2, 2251804. [Google Scholar] [CrossRef] [PubMed]
- Niswender, G.D. Response of the Corpus Luteum to Luteinizing Hormone. Environ. Health Perspect. 1981, 38, 47–50. [Google Scholar] [CrossRef]
- Krishnamurthy, H.; Kishi, H.; Shi, M.; Galet, C.; Bhaskaran, R.S.; Hirakawa, T.; Ascoli, M. Postendocytotic Trafficking of the Follicle-Stimulating Hormone (FSH)-FSH Receptor Complex. Mol. Endocrinol. 2003, 17, 2162–2176. [Google Scholar] [CrossRef]
- Mitra, S.; Rao, C. Receptors for Gonadotropins and Prostaglandins in Lysosomes of Bovine Corpora Lutea. Arch. Biochem. Biophys. 1978, 185, 126–133. [Google Scholar] [CrossRef]
- Jin, S.; Zhang, W.; Xiong, Y.; Jiang, S.; Qiao, H.; Gong, Y.; Wu, Y.; Fu, H. Identification of Important Genes Involved in the Sex-Differentiation Mechanism of Oriental River Prawn, Macrobrachium nipponense, During the Gonad Differentiation and Development Period. Front. Genet. 2022, 13, 797796. [Google Scholar] [CrossRef]
- Jiang, S.; Qiao, H.; Fu, H.; Gu, Z. Hepatopancreas Proteomic Analysis Reveals Key Proteins and Pathways in Regulatory of Ovary Maturation of Macrobrachium nipponense. Animals 2023, 13, 977. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, M.; Xiong, J.; Zhang, M.; Wu, X. Sequence Characteristics, Evolutionary History and Expression Pattern of BCO2 in Chinese Mitten Crab Eriocheir Sinensis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2025, 56, 101524. [Google Scholar] [CrossRef]
- Zhang, Y.; Kan, D.; Zhou, Y.; Lian, H.; Ge, L.; Shen, J.; Dai, Z.; Shi, Y.; Han, C.; Liu, X.; et al. Efficient RNA Interference Method by Feeding in Brachionus Plicatilis (Rotifera). Biotechnol. Lett. 2024, 46, 961–971. [Google Scholar] [CrossRef]
- Qiao, H.; Xiong, Y.; Zhang, W.; Fu, H.; Jiang, S.; Sun, S.; Bai, H.; Jin, S.; Gong, Y. Characterization, Expression, and Function Analysis of Gonad-Inhibiting Hormone in Oriental River Prawn, Macrobrachium nipponense and Its Induced Expression by Temperature. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 185, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Bayliss, J.; Nie, S.; de Nardo, W.; Keenan, S.N.; Anari, M.; Taddese, A.Z.; Williamson, N.A.; Ooi, G.J.; Brown, W.A.; et al. Liver-Secreted Hexosaminidase A Regulates Insulin-Like Growth Factor Signaling and Glucose Transport in Skeletal Muscle. Diabetes 2022, 72, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Chen, L.; Wu, P.; Zhao, W.; Li, E.; Qin, J. cDNA Cloning and Expression of Ubc9 in the Developing Embryo and Ovary of Oriental River Prawn, Macrobrachium nipponense. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 155, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Fu, Y.; Jin, S.; Fu, H.; Qiao, H.; Zhang, W.; Jiang, S.; Gong, Y.; Xiong, Y.; Wu, Y.; et al. Comparative Transcriptome Analysis of Lethality in Response to RNA Interference of the Oriental River Prawn (Macrobrachium nipponense). Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 38, 100802. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jin, S.; Fu, H.; Jiang, S.; Xiong, Y.; Sun, S.; Qiao, H.; Zhang, W.; Gong, Y.; Wu, Y. Molecular Cloning, Expression, and In Situ Hybridization Analysis of Forkhead Box Protein L2 during Development in Macrobrachium nipponense. J. World Aquac. Soc. 2018, 49, 429–440. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Jiang, S.; Zhang, W.; Jin, S.; Xiong, Y.; Xu, M.; Gao, Z.; Xu, M.; Qiao, H.; Fu, H. Functional Characterization of Two β-Hexosaminidase A Isoforms During Ovarian Development in Macrobrachium nipponense. Int. J. Mol. Sci. 2025, 26, 5459. https://doi.org/10.3390/ijms26125459
Wang Z, Jiang S, Zhang W, Jin S, Xiong Y, Xu M, Gao Z, Xu M, Qiao H, Fu H. Functional Characterization of Two β-Hexosaminidase A Isoforms During Ovarian Development in Macrobrachium nipponense. International Journal of Molecular Sciences. 2025; 26(12):5459. https://doi.org/10.3390/ijms26125459
Chicago/Turabian StyleWang, Zhiming, Sufei Jiang, Wenyi Zhang, Shubo Jin, Yiwei Xiong, Ming Xu, Zijian Gao, Mingjia Xu, Hui Qiao, and Hongtuo Fu. 2025. "Functional Characterization of Two β-Hexosaminidase A Isoforms During Ovarian Development in Macrobrachium nipponense" International Journal of Molecular Sciences 26, no. 12: 5459. https://doi.org/10.3390/ijms26125459
APA StyleWang, Z., Jiang, S., Zhang, W., Jin, S., Xiong, Y., Xu, M., Gao, Z., Xu, M., Qiao, H., & Fu, H. (2025). Functional Characterization of Two β-Hexosaminidase A Isoforms During Ovarian Development in Macrobrachium nipponense. International Journal of Molecular Sciences, 26(12), 5459. https://doi.org/10.3390/ijms26125459