
Phase-Separated Multienzyme Condensates for Efficient Synthesis of Imines from Carboxylic Acids with Enhanced Dual-Cofactor Recycling



Abstract
1. Introduction
2. Results and Discussion
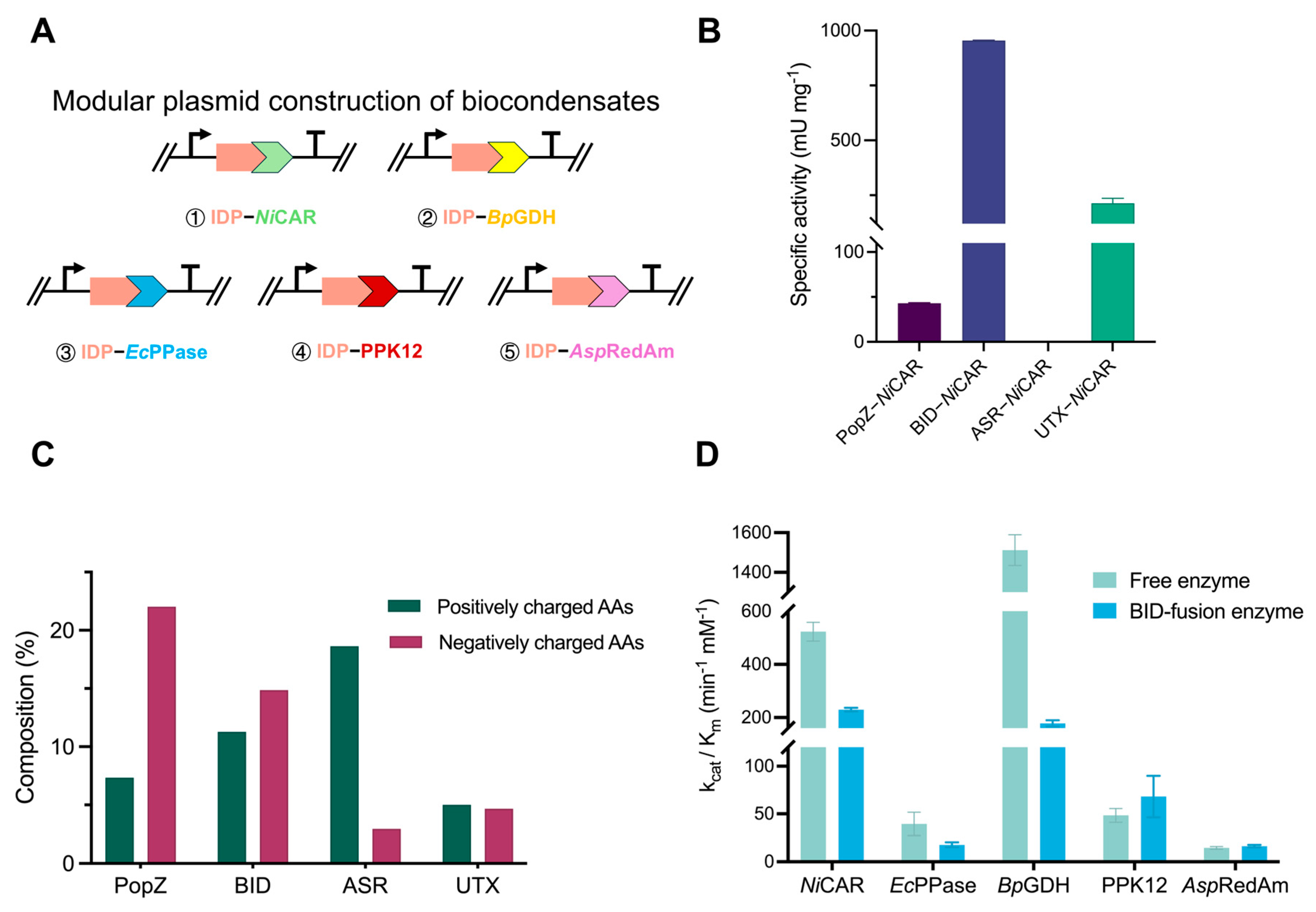
2.1. Construction of IDP-Induced Self-Assembled Multienzyme Condensates
2.2. Characterization of Fluorescent Protein Localization with Enzyme Condensates
2.3. Aldehyde Production via a Phase-Separated NiCAR System
2.4. Impact of ATP and NADPH Recycling on the Conversion of Benzoic Acid 1a
2.5. Substrate Scope and Scale-Up Amine Production
3. Materials and Methods
3.1. General Information
3.2. Preparation of Fusion Proteins
3.2.1. Carboxylate Reductase
3.2.2. Polyphosphate Kinase
3.2.3. Pyrophosphatase
3.2.4. Glucose Dehydrogenase
3.2.5. Reductive Aminase
3.3. Phase Separation Assay
3.3.1. Fluorescent-Tagged Plasmid Construction
3.3.2. Phase Separation Confocal Imaging
3.4. Fluorescent Detection of ATP and NADPH Recycling in Condensates
3.4.1. Preparation of ATP and NADPH Fluorescent Probe
3.4.2. Fluorescent Detection of Cofactors in Condensates
3.5. Enzyme-Catalyzed Reactions for Aldehyde Production
3.6. HPLC and GC Analysis
3.7. Preparation of Scale-Up Amine Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nugent, T.C.; El-Shazly, M. Chiral Amine Synthesis—Recent Developments and Trends for Enamide Reduction, Reductive Amination, and Imine Reduction. Adv. Synth. Catal. 2010, 352, 753–819. [Google Scholar] [CrossRef]
- Gunanathan, C.; Milstein, D. Applications of Acceptorless Dehydrogenation and Related Transformations in Chemical Synthesis. Science 2013, 341, 1229712. [Google Scholar] [CrossRef] [PubMed]
- Afanasyev, O.I.; Kuchuk, E.; Usanov, D.L.; Chusov, D. Reductive Amination in the Synthesis of Pharmaceuticals. Chem. Rev. 2019, 119, 11857–11911. [Google Scholar] [CrossRef] [PubMed]
- Hollmann, F.; Opperman, D.J.; Paul, C.E. Biocatalytic Reduction Reactions from a Chemist’s Perspective. Angew. Chem. Int. Ed. 2021, 60, 5644–5665. [Google Scholar] [CrossRef]
- Murugesan, K.; Senthamarai, T.; Chandrashekhar, V.G.; Natte, K.; Kamer, P.C.J.; Beller, M.; Jagadeesh, R.V. Catalytic Reductive Aminations Using Molecular Hydrogen for Synthesis of Different Kinds of Amines. Chem. Soc. Rev. 2020, 49, 6273–6328. [Google Scholar] [CrossRef]
- Elangovan, S.; Neumann, J.; Sortais, J.-B.; Junge, K.; Darcel, C.; Beller, M. Efficient and Selective N-Alkylation of Amines with Alcohols Catalysed by Manganese Pincer Complexes. Nat. Commun. 2016, 7, 12641. [Google Scholar] [CrossRef]
- Liu, H.; Chuah, G.-K.; Jaenicke, S. N-Alkylation of Amines with Alcohols over Alumina-Entrapped Ag Catalysts Using the “Borrowing Hydrogen” Methodology. J. Catal. 2012, 292, 130–137. [Google Scholar] [CrossRef]
- Luo, R.; Tong, J.; Ouyang, L.; Liu, L.; Liao, J. One-Pot Reductive Amination of Carbonyl Compounds and Nitro Compounds via Ir-Catalyzed Transfer Hydrogenation. RSC Adv. 2023, 13, 29607–29612. [Google Scholar] [CrossRef]
- Irrgang, T.; Kempe, R. Transition-Metal-Catalyzed Reductive Amination Employing Hydrogen. Chem. Rev. 2020, 120, 9583–9674. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2018, 118, 801–838. [Google Scholar] [CrossRef]
- Pavlidis, I.V.; Weiß, M.S.; Genz, M.; Spurr, P.; Hanlon, S.P.; Wirz, B.; Iding, H.; Bornscheuer, U.T. Identification of (S)-Selective Transaminases for the Asymmetric Synthesis of Bulky Chiral Amines. Nat. Chem. 2016, 8, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Ferrandi, E.E.; Monti, D. Amine Transaminases in Chiral Amines Synthesis: Recent Advances and Challenges. World J. Microbiol. Biotechnol. 2018, 34, 13. [Google Scholar] [CrossRef] [PubMed]
- Gomm, A.; O’Reilly, E. Transaminases for Chiral Amine Synthesis. Curr. Opin. Chem. Biol. 2018, 43, 106–112. [Google Scholar] [CrossRef]
- Aleku, G.A.; Hollfelder, F. Expanding the Repertoire of Imine Reductases by Mining Divergent Biosynthetic Pathways for Promiscuous Reactivity. Chem. Catal. 2024, 4, 101160. [Google Scholar] [CrossRef]
- Zhu, X.-X.; Zheng, W.-Q.; Xia, Z.-W.; Chen, X.-R.; Jin, T.; Ding, X.-W.; Chen, F.-F.; Chen, Q.; Xu, J.-H.; Kong, X.-D.; et al. Evolutionary Insights into the Stereoselectivity of Imine Reductases Based on Ancestral Sequence Reconstruction. Nat. Commun. 2024, 15, 10330. [Google Scholar] [CrossRef]
- Wu, K.; Yan, J.; Liu, Q.; Wang, X.; Wu, P.; Cao, Y.; Lu, X.; Xu, Y.; Huang, J.; Shao, L. Computational Design of an Imine Reductase: Mechanism-Guided Stereoselectivity Reversion and Interface Stabilization. Chem. Sci. 2024, 15, 1431–1440. [Google Scholar] [CrossRef]
- Kumar, R.; Karmilowicz, M.J.; Burke, D.; Burns, M.P.; Clark, L.A.; Connor, C.G.; Cordi, E.; Do, N.M.; Doyle, K.M.; Hoagland, S.; et al. Biocatalytic Reductive Amination from Discovery to Commercial Manufacturing Applied to Abrocitinib JAK1 Inhibitor. Nat. Catal. 2021, 4, 775–782. [Google Scholar] [CrossRef]
- Li, B.B.; Zhang, J.; Chen, F.F.; Chen, Q.; Xu, J.H.; Zheng, G.W. Direct Reductive Amination of Ketones with Amines by Reductive Aminases. Green. Synth. Catal. 2021, 2, 345–349. [Google Scholar] [CrossRef]
- Aleku, G.A.; France, S.P.; Man, H.; Mangas-Sanchez, J.; Montgomery, S.L.; Sharma, M.; Leipold, F.; Hussain, S.; Grogan, G.; Turner, N.J. A Reductive Aminase from Aspergillus Oryzae. Nat. Chem. 2017, 9, 961–969. [Google Scholar] [CrossRef]
- Cosgrove, S.C.; Brzezniak, A.; France, S.P.; Ramsden, J.I.; Mangas-Sanchez, J.; Montgomery, S.L.; Heath, R.S.; Turner, N.J. Imine Reductases, Reductive Aminases, and Amine Oxidases for the Synthesis of Chiral Amines: Discovery, Characterization, and Synthetic Applications. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 608, pp. 131–149. [Google Scholar]
- Aleku, G.A.; Man, H.; France, S.P.; Leipold, F.; Hussain, S.; Toca-Gonzalez, L.; Marchington, R.; Hart, S.; Turkenburg, J.P.; Grogan, G.; et al. Stereoselectivity and Structural Characterization of an Imine Reductase (IRED) from Amycolatopsis orientalis. ACS Catal. 2016, 6, 3880–3889. [Google Scholar] [CrossRef]
- Citoler, J.; Derrington, S.R.; Galman, J.L.; Bevinakatti, H.; Turner, N.J. A Biocatalytic Cascade for the Conversion of Fatty Acids to Fatty Amines. Green Chem. 2019, 21, 4932–4935. [Google Scholar] [CrossRef]
- Ramsden, J.I.; Heath, R.S.; Derrington, S.R.; Montgomery, S.L.; Mangas-Sanchez, J.; Mulholland, K.R.; Turner, N.J. Biocatalytic N-Alkylation of Amines Using Either Primary Alcohols or Carboxylic Acids via Reductive Aminase Cascades. J. Am. Chem. Soc. 2019, 141, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- France, S.P.; Hussain, S.; Hill, A.M.; Hepworth, L.J.; Howard, R.M.; Mulholland, K.R.; Flitsch, S.L.; Turner, N.J. One-Pot Cascade Synthesis of Mono- and Disubstituted Piperidines and Pyrrolidines Using Carboxylic Acid Reductase (CAR), ω-Transaminase (ω-TA), and Imine Reductase (IRED) Biocatalysts. ACS Catal. 2016, 6, 3753–3759. [Google Scholar] [CrossRef]
- Alberti, S.; Gladfelter, A.; Mittag, T. Considerations and Challenges in Studying Liquid-Liquid Phase Separation and Biomolecular Condensates. Cell 2019, 176, 419–434. [Google Scholar] [CrossRef]
- Hyman, A.A.; Weber, C.A.; Jülicher, F. Liquid-Liquid Phase Separation in Biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular Condensates: Organizers of Cellular Biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef]
- Dignon, G.L.; Best, R.B.; Mittal, J. Biomolecular Phase Separation: From Molecular Driving Forces to Macroscopic Properties. Annu. Rev. Phys. Chem. 2020, 71, 53–75. [Google Scholar] [CrossRef]
- Li, W.; Jiang, H. Nuclear Protein Condensates and Their Properties in Regulation of Gene Expression. J. Mol. Biol. 2022, 434, 167151. [Google Scholar] [CrossRef]
- Ambadipudi, S.; Biernat, J.; Riedel, D.; Mandelkow, E.; Zweckstetter, M. Liquid–Liquid Phase Separation of the Microtubule-Binding Repeats of the Alzheimer-Related Protein Tau. Nat. Commun. 2017, 8, 275. [Google Scholar] [CrossRef]
- Roden, C.; Gladfelter, A.S. RNA Contributions to the Form and Function of Biomolecular Condensates. Nat. Rev. Mol. Cell Biol. 2021, 22, 183–195. [Google Scholar] [CrossRef]
- Sabari, B.R.; Dall’Agnese, A.; Young, R.A. Biomolecular Condensates in the Nucleus. Trends Biochem. Sci. 2020, 45, 961–977. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N. Intrinsically Disordered Proteins in Overcrowded Milieu: Membrane-Less Organelles, Phase Separation, and Intrinsic Disorder. Curr. Opin. Struc Biol. 2017, 44, 18–30. [Google Scholar] [CrossRef]
- Wright, P.E.; Dyson, H.J. Intrinsically Disordered Proteins in Cellular Signalling and Regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Zhu, Y.; Zhang, W.; Mu, W. Phase-Separated Synthetic Organelles Based on Intrinsically Disordered Protein Domain for Metabolic Pathway Assembly in Escherichia coli. ACS Nano 2023, 17, 10806–10816. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, M.; Wei, Q.; Wu, W.; He, Y.; Gao, J.; Zhou, R.; Jiang, L.; Qu, J.; Xia, J. Phase-Separated Multienzyme Compartmentalization for Terpene Biosynthesis in a Prokaryote. Angew. Chem. Int. Ed. 2022, 61, e202203909. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.; Zhang, X.; Fan, D.; Bai, Y. Enhanced Cofactor Recycling and Accelerated Reaction Rate via Liquid-liquid Phase Separation in Dual-enzyme Condensates. Chin. J. Catal. 2025, 69, 135–148. [Google Scholar] [CrossRef]
- Faltova, L.; Küffner, A.M.; Hondele, M.; Weis, K.; Arosio, P. Multifunctional Protein Materials and Microreactors Using Low Complexity Domains as Molecular Adhesives. ACS Nano 2018, 12, 9991–9999. [Google Scholar] [CrossRef]
- Liu, M.; He, S.; Cheng, L.; Qu, J.; Xia, J. Phase-Separated Multienzyme Biosynthesis. Biomacromolecules 2020, 21, 2391–2399. [Google Scholar] [CrossRef]
- Venkitasubramanian, P.; Daniels, L.; Rosazza, J.P.N. Reduction of Carboxylic Acids by Nocardia Aldehyde Oxidoreductase Requires a Phosphopantetheinylated Enzyme. J. Biol. Chem. 2007, 282, 478–485. [Google Scholar] [CrossRef]
- Tavanti, M.; Hosford, J.; Lloyd, R.C.; Brown, M.J.B. ATP Regeneration by a Single Polyphosphate Kinase Powers Multigram-Scale Aldehyde Synthesis in Vitro. Green Chem. 2021, 23, 828–837. [Google Scholar] [CrossRef]
- Strohmeier, G.A.; Eiteljörg, I.C.; Schwarz, A.; Winkler, M. Enzymatic One-Step Reduction of Carboxylates to Aldehydes with Cell-Free Regeneration of ATP and NADPH. Chem. Eur. J. 2019, 25, 6119–6123. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Li, W.; Song, Y.; Wang, Z.; Ju, R.; Ulman, A.; Hu, J.; Palomba, F.; Zhao, Y.; Le, J.P.; et al. UTX Condensation Underlies Its Tumour-Suppressive Activity. Nature 2021, 597, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zheng, Y.; Liu, C.; Mencius, J.; Wu, Z.; Quan, S. Molecular Characterization of an Intrinsically Disordered Chaperone Reveals Net-Charge Regulation in Chaperone Action. J. Mol. Biol. 2022, 434, 167405. [Google Scholar] [CrossRef] [PubMed]
- Holmes, J.A.; Follett, S.E.; Wang, H.; Meadows, C.P.; Varga, K.; Bowman, G.R. Caulobacter PopZ Forms an Intrinsically Disordered Hub in Organizing Bacterial Cell Poles. Proc. Natl. Acad. Sci. USA 2016, 113, 12490–12495. [Google Scholar] [CrossRef]
- Lasker, K.; Boeynaems, S.; Lam, V.; Scholl, D.; Stainton, E.; Briner, A.; Jacquemyn, M.; Daelemans, D.; Deniz, A.; Villa, E.; et al. The Material Properties of a Bacterial-Derived Biomolecular Condensate Tune Biological Function in Natural and Synthetic Systems. Nat. Commun. 2022, 13, 5643. [Google Scholar] [CrossRef]
- Chou, J.J.; Li, H.; Salvesen, G.S.; Yuan, J.; Wagner, G. Solution Structure of BID, an Intracellular Amplifier of Apoptotic Signaling. Cell 1999, 96, 615–624. [Google Scholar] [CrossRef]
- Li, J.; Chen, C.-H.; O’Neill, K.L.; Fousek-Schuller, V.J.; Black, A.R.; Black, J.D.; Zhang, J.; Luo, X. Combined Inhibition of Aurora Kinases and Bcl-xL Induces Apoptosis through Select BH3-Only Proteins. J. Biol. Chem. 2023, 299, 102875. [Google Scholar] [CrossRef]
- Strohmeier, G.A.; Schwarz, A.; Andexer, J.N.; Winkler, M. Co-Factor Demand and Regeneration in the Enzymatic One-Step Reduction of Carboxylates to Aldehydes in Cell-Free Systems. J. Biotechnol. 2020, 307, 202–207. [Google Scholar] [CrossRef]
- Kunjapur, A.M.; Cervantes, B.; Prather, K.L.J. Coupling Carboxylic Acid Reductase to Inorganic Pyrophosphatase Enhances Cell-Free in Vitro Aldehyde Biosynthesis. Biochem. Eng. J. 2016, 109, 19–27. [Google Scholar] [CrossRef]
- Pan, X.; Zhao, Y.; Cheng, T.; Zheng, A.; Ge, A.; Zang, L.; Xu, K.; Tang, B. Monitoring NAD(P)H by an Ultrasensitive Fluorescent Probe to Reveal Reductive Stress Induced by Natural Antioxidants in HepG2 Cells under Hypoxia. Chem. Sci. 2019, 10, 8179–8186. [Google Scholar] [CrossRef]
- Xu, Z.; Singh, N.J.; Lim, J.; Pan, J.; Kim, H.N.; Park, S.; Kim, K.S.; Yoon, J. Unique Sandwich Stacking of Pyrene-Adenine-Pyrene for Selective and Ratiometric Fluorescent Sensing of ATP at Physiological pH. J. Am. Chem. Soc. 2009, 131, 15528–15533. [Google Scholar] [CrossRef] [PubMed]
- Nocek, B.; Kochinyan, S.; Proudfoot, M.; Brown, G.; Evdokimova, E.; Osipiuk, J.; Edwards, A.M.; Savchenko, A.; Joachimiak, A.; Yakunin, A.F. Polyphosphate-Dependent Synthesis of ATP and ADP by the Family-2 Polyphosphate Kinases in Bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 17730–17735. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Product | Catalyst (U/mL) | System | Time (h) | Conv. (%) | Isolated Yield (%) | STY (mg L−1 h−1) |
|---|---|---|---|---|---|---|---|
| 1 |  | 1 a | free enzymes | 8 | 83 | 58 | 144 |
| 2 | 1 b | penta-enzyme condensates | 6 | >99 | 79 | 232 | |
| 3 |  | 1.25 c | free enzymes | 8 | 79 | 50 | 132 |
| 4 | 1.25 d | penta-enzyme condensates | 6 | 90 | 64 | 212 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, T.; Zeng, L.; Liu, J.; Zhang, X.; Bai, Y. Phase-Separated Multienzyme Condensates for Efficient Synthesis of Imines from Carboxylic Acids with Enhanced Dual-Cofactor Recycling. Int. J. Mol. Sci. 2025, 26, 4795. https://doi.org/10.3390/ijms26104795
Guo T, Zeng L, Liu J, Zhang X, Bai Y. Phase-Separated Multienzyme Condensates for Efficient Synthesis of Imines from Carboxylic Acids with Enhanced Dual-Cofactor Recycling. International Journal of Molecular Sciences. 2025; 26(10):4795. https://doi.org/10.3390/ijms26104795
Chicago/Turabian StyleGuo, Tingxiao, Lifang Zeng, Jiaxu Liu, Xiaoyan Zhang, and Yunpeng Bai. 2025. "Phase-Separated Multienzyme Condensates for Efficient Synthesis of Imines from Carboxylic Acids with Enhanced Dual-Cofactor Recycling" International Journal of Molecular Sciences 26, no. 10: 4795. https://doi.org/10.3390/ijms26104795
APA StyleGuo, T., Zeng, L., Liu, J., Zhang, X., & Bai, Y. (2025). Phase-Separated Multienzyme Condensates for Efficient Synthesis of Imines from Carboxylic Acids with Enhanced Dual-Cofactor Recycling. International Journal of Molecular Sciences, 26(10), 4795. https://doi.org/10.3390/ijms26104795