
Estradiol Promotes Myelin Repair in the Spinal Cord of Female Mice in a CXCR4 Chemokine Receptor-Independent Manner

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
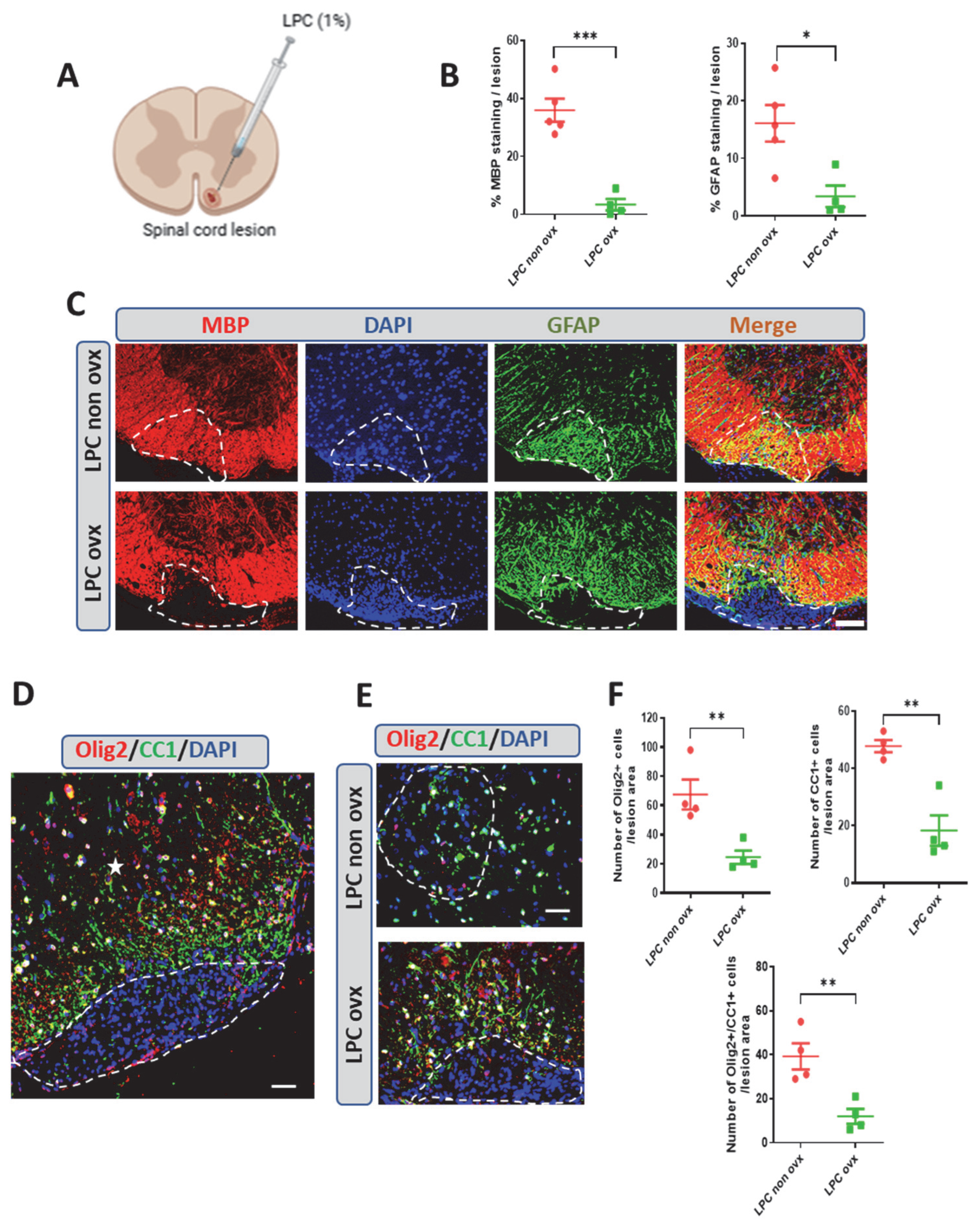
2.1. Spontaneous Remyelination in Female Mice and the Effect of Ovariectomy
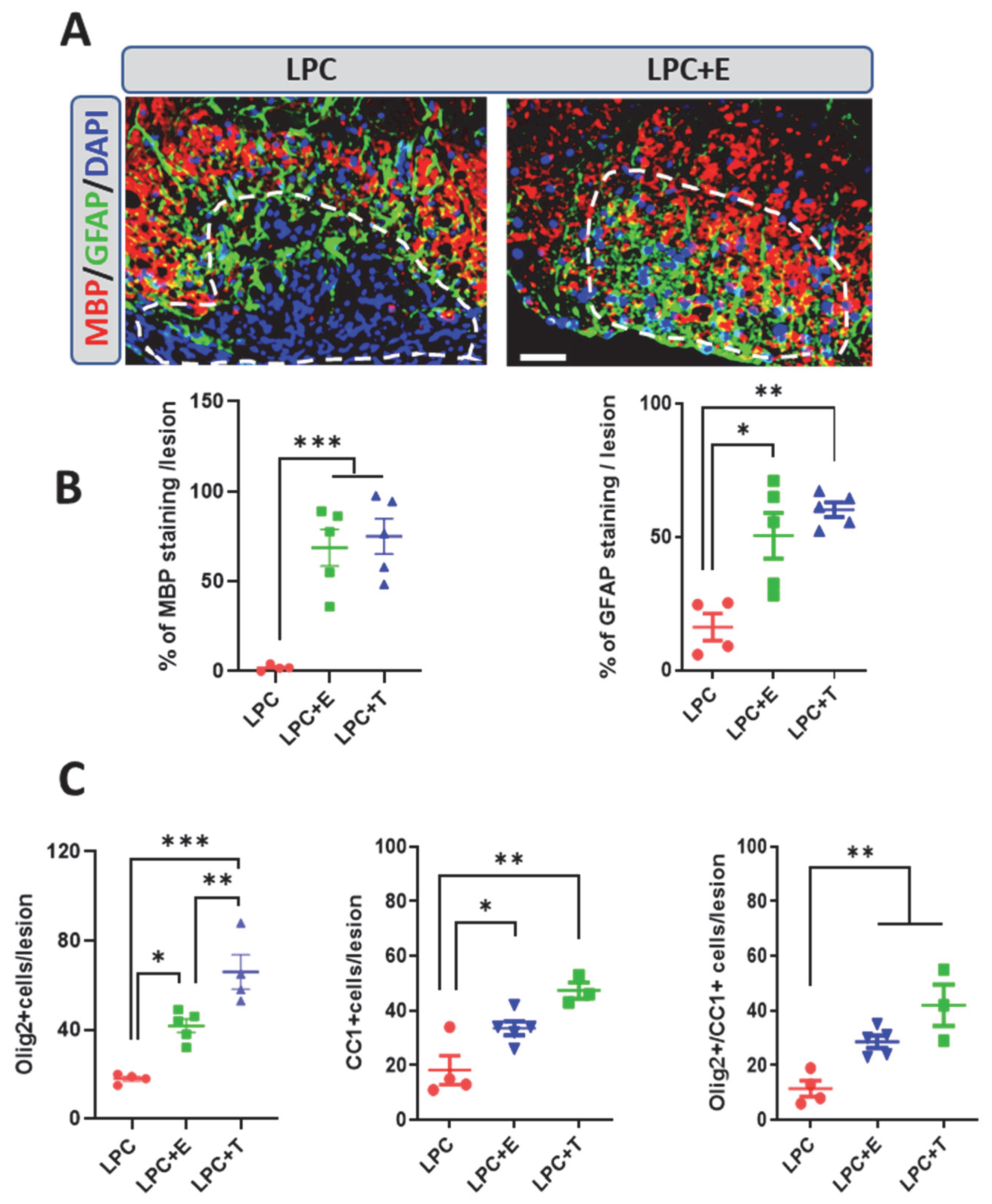
2.2. Treatment of Ovariectomized Females with Estradiol or Testosterone Restores Remyelination
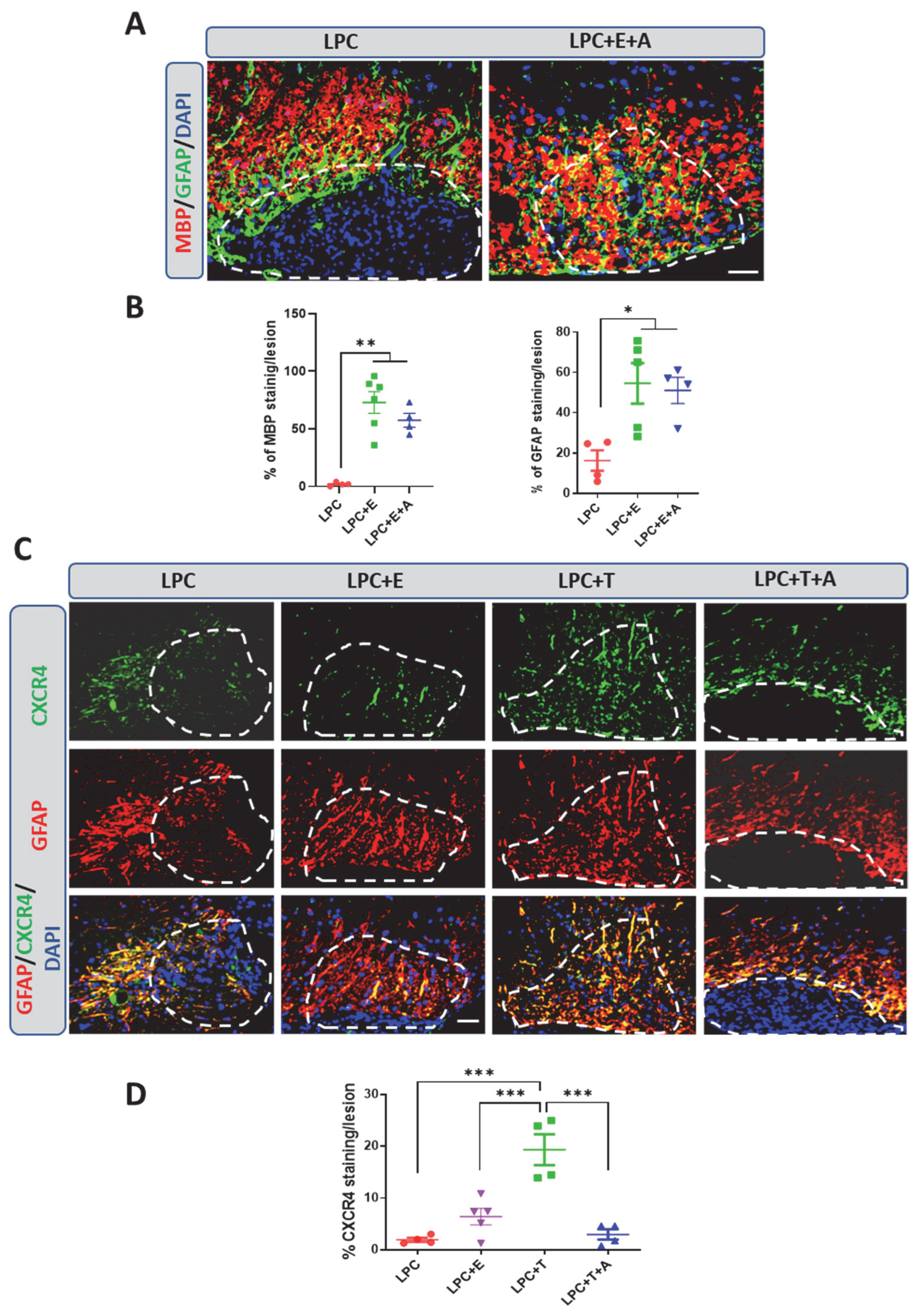
2.3. Inhibition of CXCR4 Blocks Testosterone-Dependent Remyelination—But Not Estradiol-Dependent Remyelination
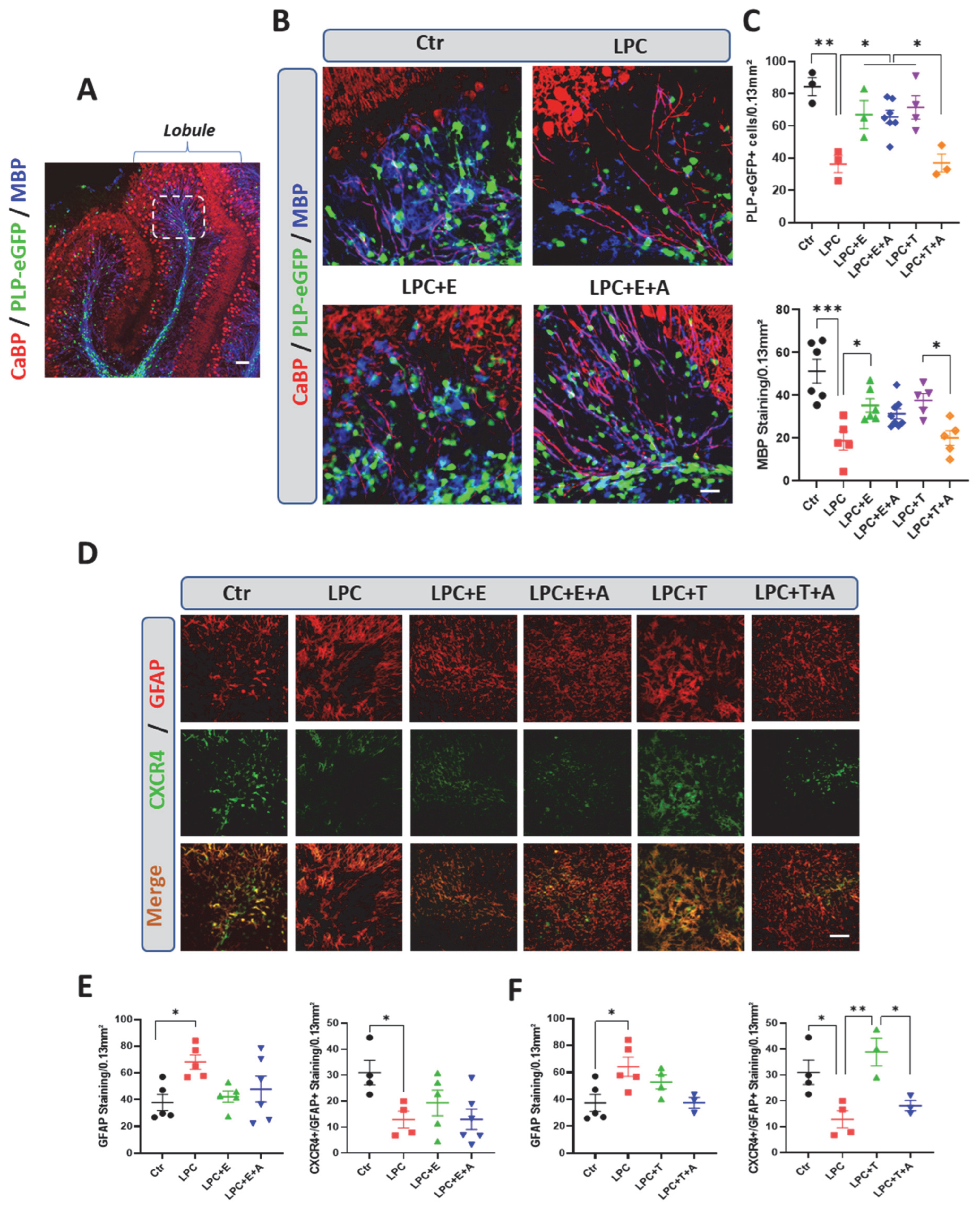
2.4. Estradiol Increases the Number of Oligodendrocytes and Promotes Remyelination in Organotypic Cultures in a CXCR4-Independent Manner
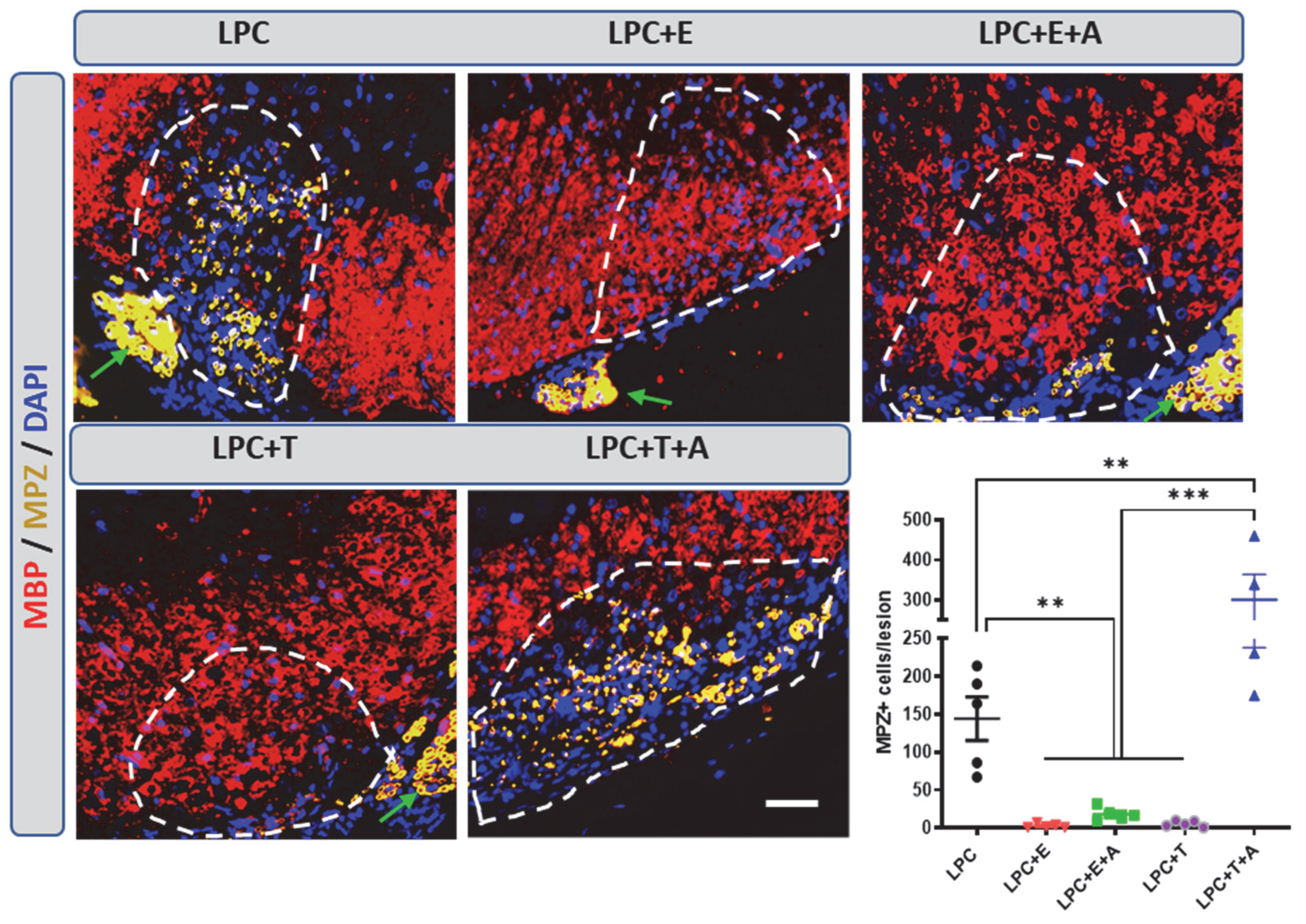
2.5. In the Absence of Estradiol and Oligodendrocyte Remyelination, Schwann Cells Invade and Remyelinate the Demyelinated Lesion
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Focal Demyelination with Lysolecithin (LPC)
4.3. Organotypic Cerebellar Slice Cultures
4.4. Immunohistochemistry
4.5. Immunohistological Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Duncan, G.J.; Simkins, T.J.; Emery, B. Neuron-Oligodendrocyte Interactions in the Structure and Integrity of Axons. Front. Cell Dev. Biol. 2021, 9, 653101. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Nave, K.-A. Oligodendrocytes: Myelination and Axonal Support. Cold Spring Harb. Perspect. Biol. 2015, 8, a020479. [Google Scholar] [CrossRef]
- Wilkins, A.; Majed, H.; Layfield, R.; Compston, A.; Chandran, S. Oligodendrocytes Promote Neuronal Survival and Axonal Length by Distinct Intracellular Mechanisms: A Novel Role for Oligodendrocyte-Derived Glial Cell Line-Derived Neurotrophic Factor. J. Neurosci. 2003, 23, 4967–4974. [Google Scholar] [CrossRef]
- Doyle, S.; Hansen, D.B.; Vella, J.; Bond, P.; Harper, G.; Zammit, C.; Valentino, M.; Fern, R. Vesicular Glutamate Release from Central Axons Contributes to Myelin Damage. Nat. Commun. 2018, 9, 1032. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.J.M.; Ffrench-Constant, C.; Edgar, J.M.; Smith, K.J. Neuroprotection and Repair in Multiple Sclerosis. Nat. Rev. Neurol. 2012, 8, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ren, C.; Luo, Y.; Ding, Y.; Ji, X.; Li, S. New Insights into the Roles of Oligodendrocytes Regulation in Ischemic Stroke Recovery. Neurobiol. Dis. 2023, 184, 106200. [Google Scholar] [CrossRef]
- Crawford, A.H.; Stockley, J.H.; Tripathi, R.B.; Richardson, W.D.; Franklin, R.J.M. Oligodendrocyte Progenitors: Adult Stem Cells of the Central Nervous System? Exp. Neurol. 2014, 260, 50–55. [Google Scholar] [CrossRef]
- Cheng, Y.-J.; Wang, F.; Feng, J.; Yu, B.; Wang, B.; Gao, Q.; Wang, T.-Y.; Hu, B.; Gao, X.; Chen, J.-F.; et al. Prolonged Myelin Deficits Contribute to Neuron Loss and Functional Impairments after Ischaemic Stroke. Brain 2024, 147, 1294–1311. [Google Scholar] [CrossRef]
- Franklin, R.J.M.; Bodini, B.; Goldman, S.A. Remyelination in the Central Nervous System. Cold Spring Harb. Perspect. Biol. 2024, 16, a041371. [Google Scholar] [CrossRef]
- Bove, R.; Chitnis, T. The Role of Gender and Sex Hormones in Determining the Onset and Outcome of Multiple Sclerosis. Mult. Scler. 2014, 20, 520–526. [Google Scholar] [CrossRef]
- Pakpoor, J.; Goldacre, R.; Schmierer, K.; Giovannoni, G.; Goldacre, M.J. Testicular Hypofunction and Multiple Sclerosis Risk: A Record-Linkage Study. Ann. Neurol. 2014, 76, 625–628. [Google Scholar] [CrossRef]
- Asbelaoui, N.; Abi-Ghanem, C.; Schlecht-Louf, G.; Oukil, H.; Degerny, C.; Bank, T.N.B.; Schumacher, M.; Ghoumari, A.M. Interplay between Androgen and CXCR4 Chemokine Signaling in Myelin Repair. Acta Neuropathol. Commun. 2024, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Bielecki, B.; Mattern, C.; Ghoumari, A.M.; Javaid, S.; Smietanka, K.; Abi Ghanem, C.; Mhaouty-Kodja, S.; Ghandour, M.S.; Baulieu, E.-E.; Franklin, R.J.M.; et al. Unexpected Central Role of the Androgen Receptor in the Spontaneous Regeneration of Myelin. Proc. Natl. Acad. Sci. USA 2016, 113, 14829–14834. [Google Scholar] [CrossRef]
- Dalal, M.; Kim, S.; Voskuhl, R.R. Testosterone Therapy Ameliorates Experimental Autoimmune Encephalomyelitis and Induces a T Helper 2 Bias in the Autoantigen-Specific T Lymphocyte Response. J. Immunol. 1997, 159, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Laouarem, Y.; Kassoussi, A.; Zahaf, A.; Hutteau-Hamel, T.; Mellouk, A.; Bobé, P.; Mattern, C.; Schumacher, M.; Traiffort, E. Functional Cooperation of the Hedgehog and Androgen Signaling Pathways during Developmental and Repairing Myelination. Glia 2021, 69, 1369–1392. [Google Scholar] [CrossRef] [PubMed]
- Carbajal, K.S.; Miranda, J.L.; Tsukamoto, M.R.; Lane, T.E. CXCR4 Signaling Regulates Remyelination by Endogenous Oligodendrocyte Progenitor Cells in a Viral Model of Demyelination. Glia 2011, 59, 1813–1821. [Google Scholar] [CrossRef]
- Patel, J.R.; McCandless, E.E.; Dorsey, D.; Klein, R.S. CXCR4 Promotes Differentiation of Oligodendrocyte Progenitors and Remyelination. Proc. Natl. Acad. Sci. USA 2010, 107, 11062–11067. [Google Scholar] [CrossRef]
- Talbott, J.F.; Cao, Q.; Enzmann, G.U.; Benton, R.L.; Achim, V.; Cheng, X.X.; Mills, M.D.; Rao, M.S.; Whittemore, S.R. Schwann Cell-like Differentiation by Adult Oligodendrocyte Precursor Cells Following Engraftment into the Demyelinated Spinal Cord Is BMP-Dependent. Glia 2006, 54, 147–159. [Google Scholar] [CrossRef]
- Zawadzka, M.; Rivers, L.E.; Fancy, S.P.J.; Zhao, C.; Tripathi, R.; Jamen, F.; Young, K.; Goncharevich, A.; Pohl, H.; Rizzi, M.; et al. CNS-Resident Glial Progenitor/Stem Cells Produce Schwann Cells as Well as Oligodendrocytes during Repair of CNS Demyelination. Cell Stem Cell 2010, 6, 578–590. [Google Scholar] [CrossRef]
- Ghezzi, L.; Bollman, B.; De Feo, L.; Piccio, L.; Trapp, B.D.; Schmidt, R.E.; Cross, A.H. Schwann Cell Remyelination in the Multiple Sclerosis Central Nervous System. Lab. Investig. 2023, 103, 100128. [Google Scholar] [CrossRef]
- Itoyama, Y.; Ohnishi, A.; Tateishi, J.; Kuroiwa, Y.; Webster, H.D. Spinal Cord Multiple Sclerosis Lesions in Japanese Patients: Schwann Cell Remyelination Occurs in Areas That Lack Glial Fibrillary Acidic Protein (GFAP). Acta Neuropathol. 1985, 65, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Hussain, R.; Ghoumari, A.M.; Bielecki, B.; Steibel, J.; Boehm, N.; Liere, P.; Macklin, W.B.; Kumar, N.; Habert, R.; Mhaouty-Kodja, S.; et al. The Neural Androgen Receptor: A Therapeutic Target for Myelin Repair in Chronic Demyelination. Brain 2013, 136, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Bui, H.N.; Struys, E.A.; Martens, F.; de Ronde, W.; Thienpont, L.M.; Kenemans, P.; Verhoeven, M.O.; Jakobs, C.; Dijstelbloem, H.M.; Blankenstein, M.A. Serum Testosterone Levels Measured by Isotope Dilution-Liquid Chromatography-Tandem Mass Spectrometry in Postmenopausal Women versus Those in Women Who Underwent Bilateral Oophorectomy. Ann. Clin. Biochem. 2010, 47, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Davio, A.; Woolcock, H.; Nanba, A.T.; Rege, J.; O’Day, P.; Ren, J.; Zhao, L.; Ebina, H.; Auchus, R.; Rainey, W.E.; et al. Sex Differences in 11-Oxygenated Androgen Patterns Across Adulthood. J. Clin. Endocrinol. Metab. 2020, 105, e2921–e2929. [Google Scholar] [CrossRef]
- Frederiksen, H.; Johannsen, T.H.; Andersen, S.E.; Albrethsen, J.; Landersoe, S.K.; Petersen, J.H.; Andersen, A.N.; Vestergaard, E.T.; Schorring, M.E.; Linneberg, A.; et al. Sex-Specific Estrogen Levels and Reference Intervals from Infancy to Late Adulthood Determined by LC-MS/MS. J. Clin. Endocrinol. Metab. 2020, 105, 754–768. [Google Scholar] [CrossRef]
- Abi Ghanem, C.; Degerny, C.; Hussain, R.; Liere, P.; Pianos, A.; Tourpin, S.; Habert, R.; Macklin, W.B.; Schumacher, M.; Ghoumari, A.M. Long-Lasting Masculinizing Effects of Postnatal Androgens on Myelin Governed by the Brain Androgen Receptor. PLoS Genet. 2017, 13, e1007049. [Google Scholar] [CrossRef]
- Hussain, R.; El-Etr, M.; Gaci, O.; Rakotomamonjy, J.; Macklin, W.B.; Kumar, N.; Sitruk-Ware, R.; Schumacher, M.; Ghoumari, A.M. Progesterone and Nestorone Facilitate Axon Remyelination: A Role for Progesterone Receptors. Endocrinology 2011, 152, 3820–3831. [Google Scholar] [CrossRef]
- Mallon, B.S.; Shick, H.E.; Kidd, G.J.; Macklin, W.B. Proteolipid Promoter Activity Distinguishes Two Populations of NG2-Positive Cells throughout Neonatal Cortical Development. J. Neurosci. 2002, 22, 876–885. [Google Scholar] [CrossRef]
- Gorter, R.P.; Baron, W. Recent Insights into Astrocytes as Therapeutic Targets for Demyelinating Diseases. Curr. Opin. Pharmacol. 2022, 65, 102261. [Google Scholar] [CrossRef]
- Rawji, K.S.; Gonzalez Martinez, G.A.; Sharma, A.; Franklin, R.J.M. The Role of Astrocytes in Remyelination. Trends Neurosci. 2020, 43, 596–607. [Google Scholar] [CrossRef]
- Talbott, J.F.; Loy, D.N.; Liu, Y.; Qiu, M.S.; Bunge, M.B.; Rao, M.S.; Whittemore, S.R. Endogenous Nkx2.2+/Olig2+ Oligodendrocyte Precursor Cells Fail to Remyelinate the Demyelinated Adult Rat Spinal Cord in the Absence of Astrocytes. Exp. Neurol. 2005, 192, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Moore, S.; Crawford, D.K.; Hannsun, G.; Sasidhar, M.V.; Tan, K.; Molaie, D.; Tiwari-Woodruff, S.K. Attenuation of Corpus Callosum Axon Myelination and Remyelination in the Absence of Circulating Sex Hormones. Brain Pathol. 2013, 23, 462–475. [Google Scholar] [CrossRef]
- Spence, R.D.; Voskuhl, R.R. Neuroprotective Effects of Estrogens and Androgens in CNS Inflammation and Neurodegeneration. Front. Neuroendocrinol. 2012, 33, 105–115. [Google Scholar] [CrossRef]
- Assinck, P.; Duncan, G.J.; Plemel, J.R.; Lee, M.J.; Stratton, J.A.; Manesh, S.B.; Liu, J.; Ramer, L.M.; Kang, S.H.; Bergles, D.E.; et al. Myelinogenic Plasticity of Oligodendrocyte Precursor Cells Following Spinal Cord Contusion Injury. J. Neurosci. 2017, 37, 8635–8654. [Google Scholar] [CrossRef] [PubMed]
- Afshari, F.T.; Kwok, J.C.; Fawcett, J.W. Astrocyte-Produced Ephrins Inhibit Schwann Cell Migration via VAV2 Signaling. J. Neurosci. 2010, 30, 4246–4255. [Google Scholar] [CrossRef]
- Garcia-Diaz, B.; Baron-Van Evercooren, A. Schwann Cells: Rescuers of Central Demyelination. Glia 2020, 68, 1945–1956. [Google Scholar] [CrossRef]
- Woodruff, R.H.; Franklin, R.J. Demyelination and Remyelination of the Caudal Cerebellar Peduncle of Adult Rats Following Stereotaxic Injections of Lysolecithin, Ethidium Bromide, and Complement/Anti-Galactocerebroside: A Comparative Study. Glia 1999, 25, 216–228. [Google Scholar] [CrossRef]
- Itoyama, Y.; Webster, H.D.; Richardson, E.P.; Trapp, B.D. Schwann Cell Remyelination of Demyelinated Axons in Spinal Cord Multiple Sclerosis Lesions. Ann. Neurol. 1983, 14, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Dahlman-Wright, K.; Gustafsson, J.-Å. Estrogen Receptor Alpha and Beta in Health and Disease. Best. Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 557–568. [Google Scholar] [CrossRef]
- Spence, R.D.; Wisdom, A.J.; Cao, Y.; Hill, H.M.; Mongerson, C.R.L.; Stapornkul, B.; Itoh, N.; Sofroniew, M.V.; Voskuhl, R.R. Estrogen Mediates Neuroprotection and Anti-Inflammatory Effects during EAE through ER Signaling on Astrocytes But Not through ER Signaling on Astrocytes or Neurons. J. Neurosci. 2013, 33, 10924–10933. [Google Scholar] [CrossRef]
- Khalaj, A.J.; Yoon, J.; Nakai, J.; Winchester, Z.; Moore, S.M.; Yoo, T.; Martinez-Torres, L.; Kumar, S.; Itoh, N.; Tiwari-Woodruff, S.K. Estrogen Receptor (ER) β Expression in Oligodendrocytes Is Required for Attenuation of Clinical Disease by an ERβ Ligand. Proc. Natl. Acad. Sci. USA 2013, 110, 19125–19130. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.D.; Hamby, M.E.; Umeda, E.; Itoh, N.; Du, S.; Wisdom, A.J.; Cao, Y.; Bondar, G.; Lam, J.; Ao, Y.; et al. Neuroprotection Mediated through Estrogen Receptor-Alpha in Astrocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 8867–8872. [Google Scholar] [CrossRef]
- Laffont, S.; Garnier, L.; Lélu, K.; Guéry, J.-C. Estrogen-Mediated Protection of Experimental Autoimmune Encephalomyelitis: Lessons from the Dissection of Estrogen Receptor-Signaling In Vivo. Biomed. J. 2015, 38, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, M.-A.; Azcoitia, I.; Garcia-Segura, L.M. The Neuroprotective Actions of Oestradiol and Oestrogen Receptors. Nat. Rev. Neurosci. 2015, 16, 17–29. [Google Scholar] [CrossRef]
- Voskuhl, R.R.; Itoh, N.; Tassoni, A.; Matsukawa, M.A.; Ren, E.; Tse, V.; Jang, E.; Suen, T.T.; Itoh, Y. Gene Expression in Oligodendrocytes during Remyelination Reveals Cholesterol Homeostasis as a Therapeutic Target in Multiple Sclerosis. Proc. Natl. Acad. Sci. USA 2019, 116, 10130–10139. [Google Scholar] [CrossRef]
- Karim, H.; Kim, S.H.; Lapato, A.S.; Yasui, N.; Katzenellenbogen, J.A.; Tiwari-Woodruff, S.K. Increase in Chemokine CXCL1 by ERβ Ligand Treatment Is a Key Mediator in Promoting Axon Myelination. Proc. Natl. Acad. Sci. USA 2018, 115, 6291–6296. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Itoh, Y.; Meyer, C.E.; Suen, T.T.; Cortez-Delgado, D.; Rivera Lomeli, M.; Wendin, S.; Somepalli, S.S.; Golden, L.C.; MacKenzie-Graham, A.; et al. Estrogen Receptor Beta in Astrocytes Modulates Cognitive Function in Mid-Age Female Mice. Nat. Commun. 2023, 14, 6044. [Google Scholar] [CrossRef]
- Tiwari-Woodruff, S.; Morales, L.B.J.; Lee, R.; Voskuhl, R.R. Differential Neuroprotective and Antiinflammatory Effects of Estrogen Receptor (ER)α and ERβ Ligand Treatment. Proc. Natl. Acad. Sci. USA 2007, 104, 14813–14818. [Google Scholar] [CrossRef]
- Wu, W.; Tan, X.; Dai, Y.; Krishnan, V.; Warner, M.; Gustafsson, J.-Å. Targeting Estrogen Receptor β in Microglia and T Cells to Treat Experimental Autoimmune Encephalomyelitis. Proc. Natl. Acad. Sci. USA 2013, 110, 3543–3548. [Google Scholar] [CrossRef]
- Labombarda, F.; Pianos, A.; Liere, P.; Eychenne, B.; Gonzalez, S.; Cambourg, A.; De Nicola, A.F.; Schumacher, M.; Guennoun, R. Injury Elicited Increase in Spinal Cord Neurosteroid Content Analyzed by Gas Chromatography Mass Spectrometry. Endocrinology 2006, 147, 1847–1859. [Google Scholar] [CrossRef]
- Schumacher, M.; Mattern, C.; Ghoumari, A.; Oudinet, J.P.; Liere, P.; Labombarda, F.; Sitruk-Ware, R.; De Nicola, A.F.; Guennoun, R. Revisiting the Roles of Progesterone and Allopregnanolone in the Nervous System: Resurgence of the Progesterone Receptors. Prog. Neurobiol. 2014, 113, 6–39. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Fréchou, M.; Liere, P.; Zhang, S.; Pianos, A.; Fernandez, N.; Denier, C.; Mattern, C.; Schumacher, M.; Guennoun, R. A Role of Endogenous Progesterone in Stroke Cerebroprotection Revealed by the Neural-Specific Deletion of Its Intracellular Receptors. J. Neurosci. 2017, 37, 10998–11020. [Google Scholar] [CrossRef] [PubMed]
- El-Etr, M.; Rame, M.; Boucher, C.; Ghoumari, A.M.; Kumar, N.; Liere, P.; Pianos, A.; Schumacher, M.; Sitruk-Ware, R. Progesterone and Nestorone Promote Myelin Regeneration in Chronic Demyelinating Lesions of Corpus Callosum and Cerebral Cortex. Glia 2015, 63, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Ghoumari, A.M.; Ibanez, C.; El-Etr, M.; Leclerc, P.; Eychenne, B.; O’Malley, B.W.; Baulieu, E.E.; Schumacher, M. Progesterone and Its Metabolites Increase Myelin Basic Protein Expression in Organotypic Slice Cultures of Rat Cerebellum. J. Neurochem. 2003, 86, 848–859. [Google Scholar] [CrossRef]
- Acs, P.; Kipp, M.; Norkute, A.; Johann, S.; Clarner, T.; Braun, A.; Berente, Z.; Komoly, S.; Beyer, C. 17β-estradiol and Progesterone Prevent Cuprizone Provoked Demyelination of Corpus Callosum in Male Mice. Glia 2009, 57, 807–814. [Google Scholar] [CrossRef]
- Garay, L.; Gonzalez Deniselle, M.C.; Gierman, L.; Meyer, M.; Lima, A.; Roig, P.; De Nicola, A.F. Steroid Protection in the Experimental Autoimmune Encephalomyelitis Model of Multiple Sclerosis. Neuroimmunomodulation 2008, 15, 76–83. [Google Scholar] [CrossRef]
- Kipp, M.; Amor, S.; Krauth, R.; Beyer, C. Multiple Sclerosis: Neuroprotective Alliance of Estrogen-Progesterone and Gender. Front. Neuroendocrinol. 2012, 33, 1–16. [Google Scholar] [CrossRef]
- Palaszynski, K.M.; Liu, H.; Loo, K.K.; Voskuhl, R.R. Estriol Treatment Ameliorates Disease in Males with Experimental Autoimmune Encephalomyelitis: Implications for Multiple Sclerosis. J. Neuroimmunol. 2004, 149, 84–89. [Google Scholar] [CrossRef]
- Zahaf, A.; Kassoussi, A.; Hutteau-Hamel, T.; Mellouk, A.; Marie, C.; Zoupi, L.; Tsouki, F.; Mattern, C.; Bobé, P.; Schumacher, M.; et al. Androgens Show Sex-Dependent Differences in Myelination in Immune and Non-Immune Murine Models of CNS Demyelination. Nat. Commun. 2023, 14, 1592. [Google Scholar] [CrossRef]
- Petheram, K.; Dobson, R. All Women with Multiple Sclerosis Should Start Hormone Replacement Therapy at Menopause Unless Contraindicated: Commentary. Mult. Scler. 2024, 30, 1111–1112. [Google Scholar] [CrossRef]
- Voskuhl, R. All Women with Multiple Sclerosis Should Start Hormone Replacement Therapy at Menopause Unless Contraindicated: Yes. Mult. Scler. 2024, 30, 1107–1109. [Google Scholar] [CrossRef] [PubMed]
- Canonico, M.; Carcaillon, L.; Plu-Bureau, G.; Oger, E.; Singh-Manoux, A.; Tubert-Bitter, P.; Elbaz, A.; Scarabin, P.-Y. Postmenopausal Hormone Therapy and Risk of Stroke: Impact of the Route of Estrogen Administration and Type of Progestogen. Stroke 2016, 47, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- L’Hermite, M. Bioidentical Menopausal Hormone Therapy: Registered Hormones (Non-Oral Estradiol ± Progesterone) Are Optimal. Climacteric 2017, 20, 331–338. [Google Scholar] [CrossRef]
- Lorite, M.I.; Cuadros, A.M.; Rivera-Izquierdo, M.; Sanchez-Martin, V.; Cuadros, M. Benefits for Cardiovascular System, Bone Density, and Quality of Life of a Long-Term Hormone Therapy in Hysterectomized Women: A 20-Year Follow-up Study. Menopause 2023, 30, 995–1001. [Google Scholar] [CrossRef]
- Mirkin, S.; Ryan, K.A.; Chandran, A.B.; Komm, B.S. Bazedoxifene/Conjugated Estrogens for Managing the Burden of Estrogen Deficiency Symptoms. Maturitas 2014, 77, 24–31. [Google Scholar] [CrossRef]
- Itoh, N.; Kim, R.; Peng, M.; DiFilippo, E.; Johnsonbaugh, H.; MacKenzie-Graham, A.; Voskuhl, R.R. Bedside to Bench to Bedside Research: Estrogen Receptor Beta Ligand as a Candidate Neuroprotective Treatment for Multiple Sclerosis. J. Neuroimmunol. 2017, 304, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Voskuhl, R.R.; Wang, H.; Wu, T.C.J.; Sicotte, N.L.; Nakamura, K.; Kurth, F.; Itoh, N.; Bardens, J.; Bernard, J.T.; Corboy, J.R.; et al. Estriol Combined with Glatiramer Acetate for Women with Relapsing-Remitting Multiple Sclerosis: A Randomised, Placebo-Controlled, Phase 2 Trial. Lancet Neurol. 2016, 15, 35–46. [Google Scholar] [CrossRef]
- Escande, A.; Pillon, A.; Servant, N.; Cravedi, J.-P.; Larrea, F.; Muhn, P.; Nicolas, J.-C.; Cavaillès, V.; Balaguer, P. Evaluation of Ligand Selectivity Using Reporter Cell Lines Stably Expressing Estrogen Receptor Alpha or Beta. Biochem. Pharmacol. 2006, 71, 1459–1469. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Häggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the Ligand Binding Specificity and Transcript Tissue Distribution of Estrogen Receptors Alpha and Beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Ali, E.S.; Mangold, C.; Peiris, A.N. Estriol: Emerging Clinical Benefits. Menopause 2017, 24, 1081–1085. [Google Scholar] [CrossRef]
- Ghoumari, A.M.; Baulieu, E.E.; Schumacher, M. Progesterone Increases Oligodendroglial Cell Proliferation in Rat Cerebellar Slice Cultures. Neuroscience 2005, 135, 47–58. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bardy-Lagarde, M.; Asbelaoui, N.; Schumacher, M.; Ghoumari, A.M. Estradiol Promotes Myelin Repair in the Spinal Cord of Female Mice in a CXCR4 Chemokine Receptor-Independent Manner. Int. J. Mol. Sci. 2025, 26, 4752. https://doi.org/10.3390/ijms26104752
Bardy-Lagarde M, Asbelaoui N, Schumacher M, Ghoumari AM. Estradiol Promotes Myelin Repair in the Spinal Cord of Female Mice in a CXCR4 Chemokine Receptor-Independent Manner. International Journal of Molecular Sciences. 2025; 26(10):4752. https://doi.org/10.3390/ijms26104752
Chicago/Turabian StyleBardy-Lagarde, Marianne, Narimene Asbelaoui, Michael Schumacher, and Abdel Mouman Ghoumari. 2025. "Estradiol Promotes Myelin Repair in the Spinal Cord of Female Mice in a CXCR4 Chemokine Receptor-Independent Manner" International Journal of Molecular Sciences 26, no. 10: 4752. https://doi.org/10.3390/ijms26104752
APA StyleBardy-Lagarde, M., Asbelaoui, N., Schumacher, M., & Ghoumari, A. M. (2025). Estradiol Promotes Myelin Repair in the Spinal Cord of Female Mice in a CXCR4 Chemokine Receptor-Independent Manner. International Journal of Molecular Sciences, 26(10), 4752. https://doi.org/10.3390/ijms26104752