
Ergosterol Peroxide Disrupts Triple-Negative Breast Cancer Mitochondrial Function and Inhibits Tumor Growth and Metastasis


 , , , , ,
, , , , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
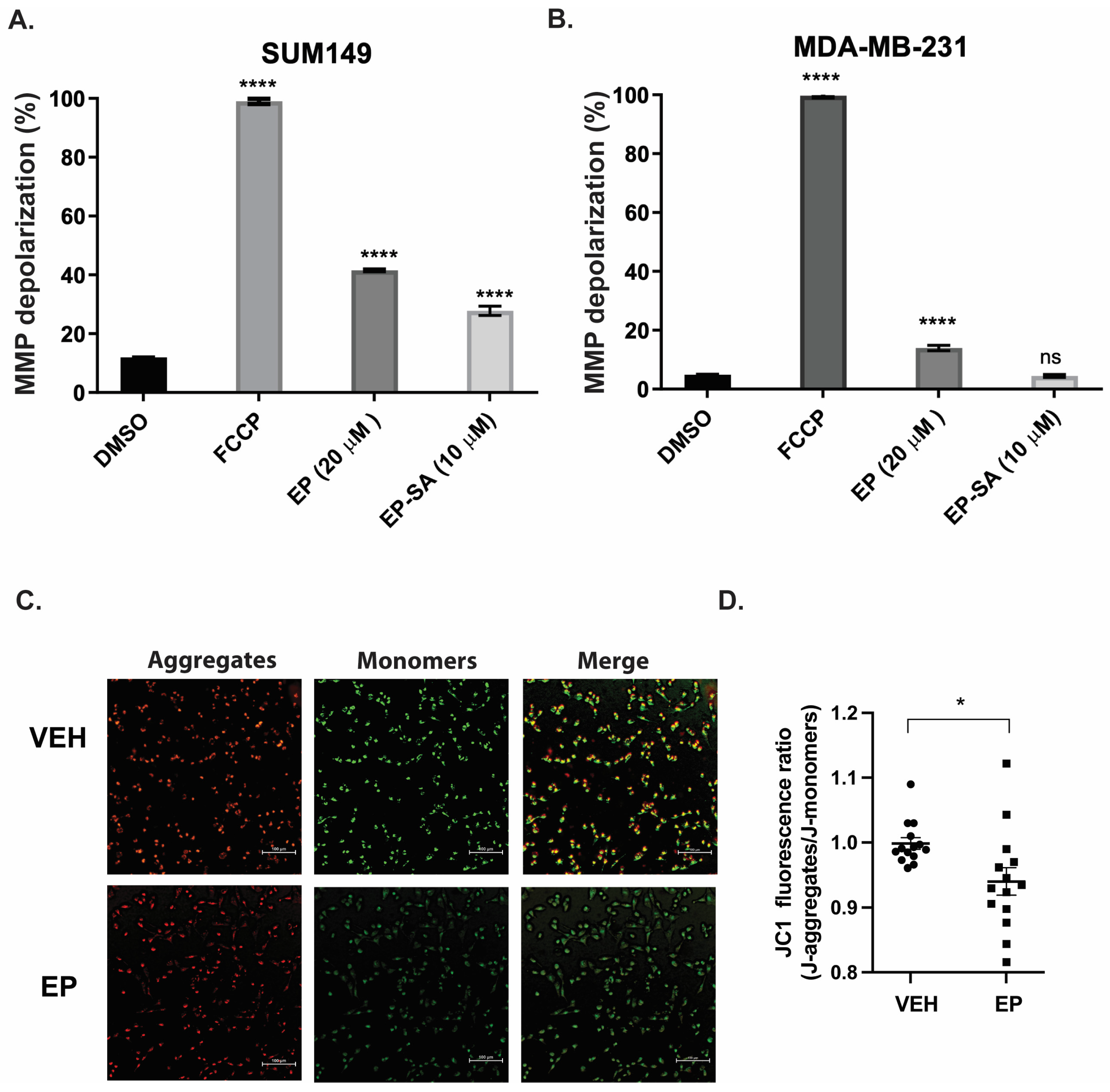
2.1. EP Affects Mitochondrial Membrane Potential in TNBC Cells
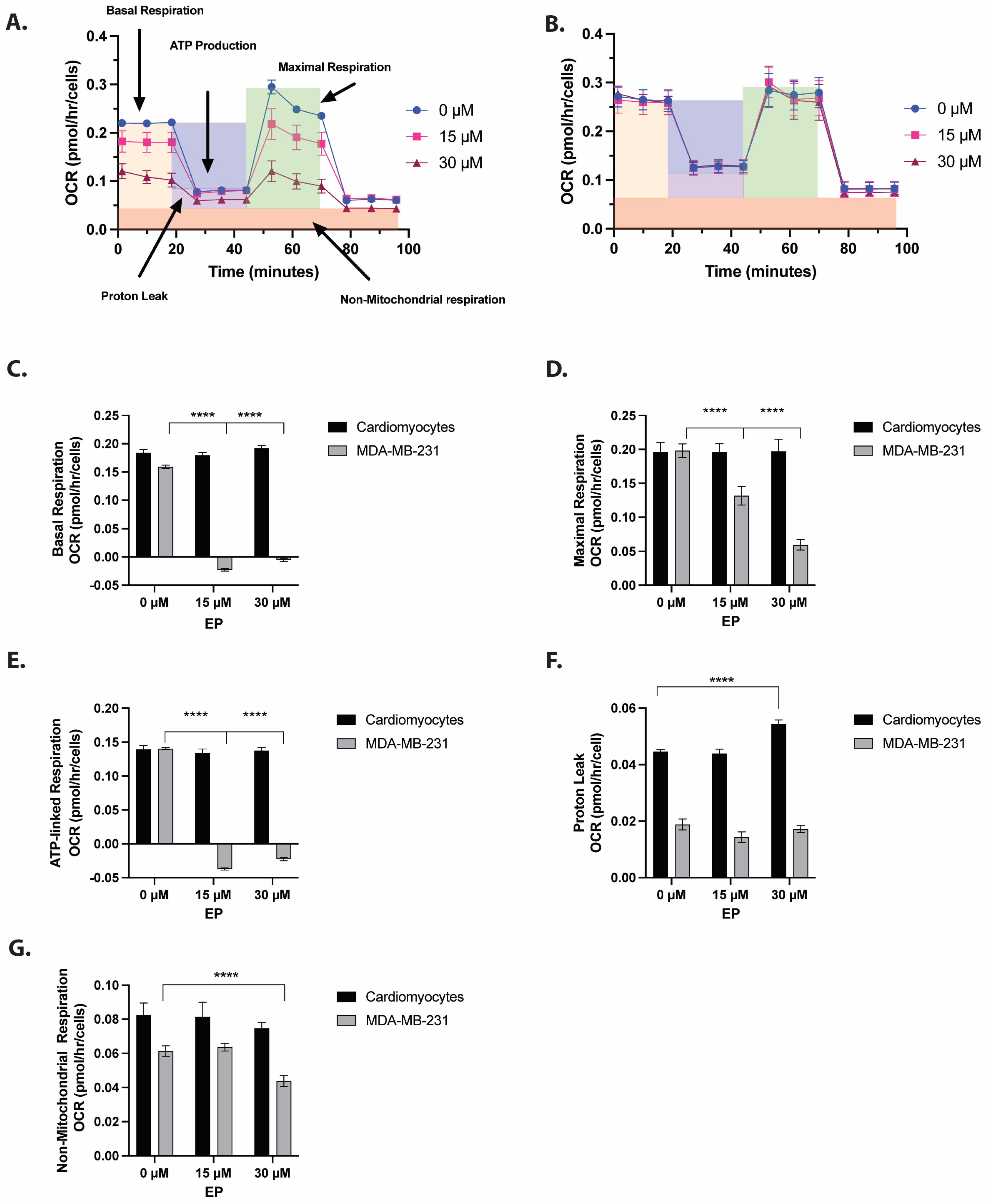
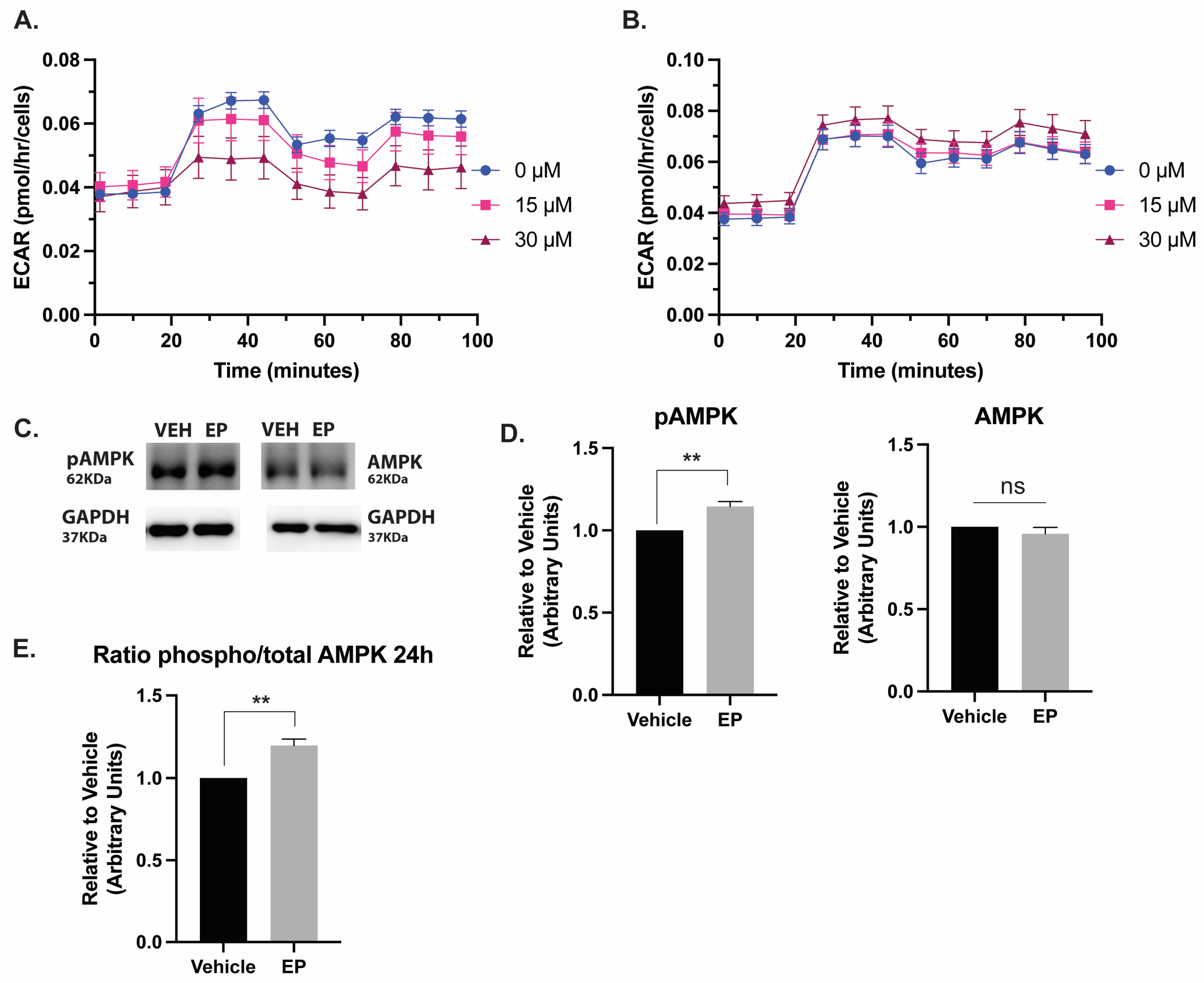
2.2. EP Selectively Reduces Cellular Respiration and Glycolysis in TNBC Cells
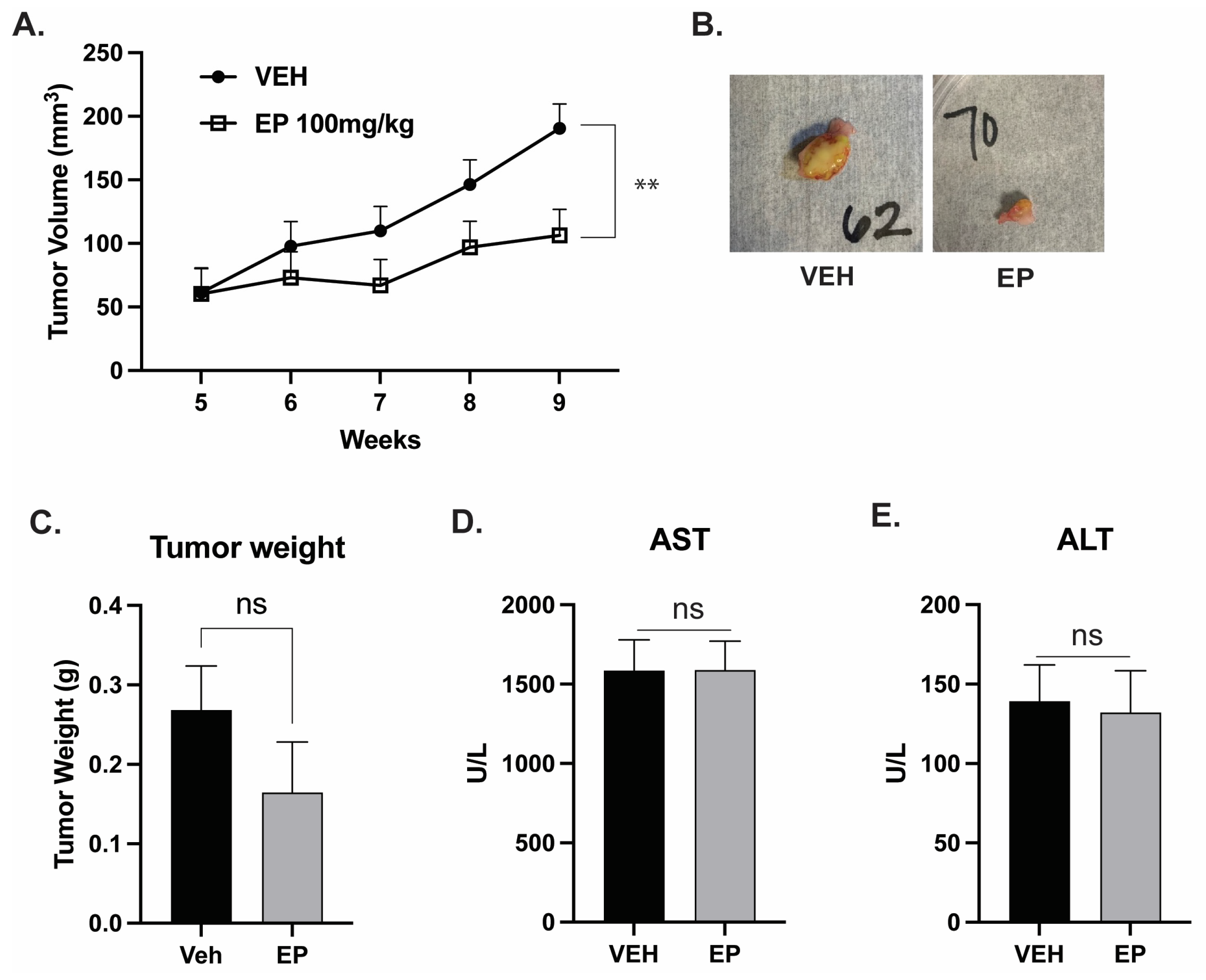
2.3. EP Reduces Tumor Volume in a Model of TNBC
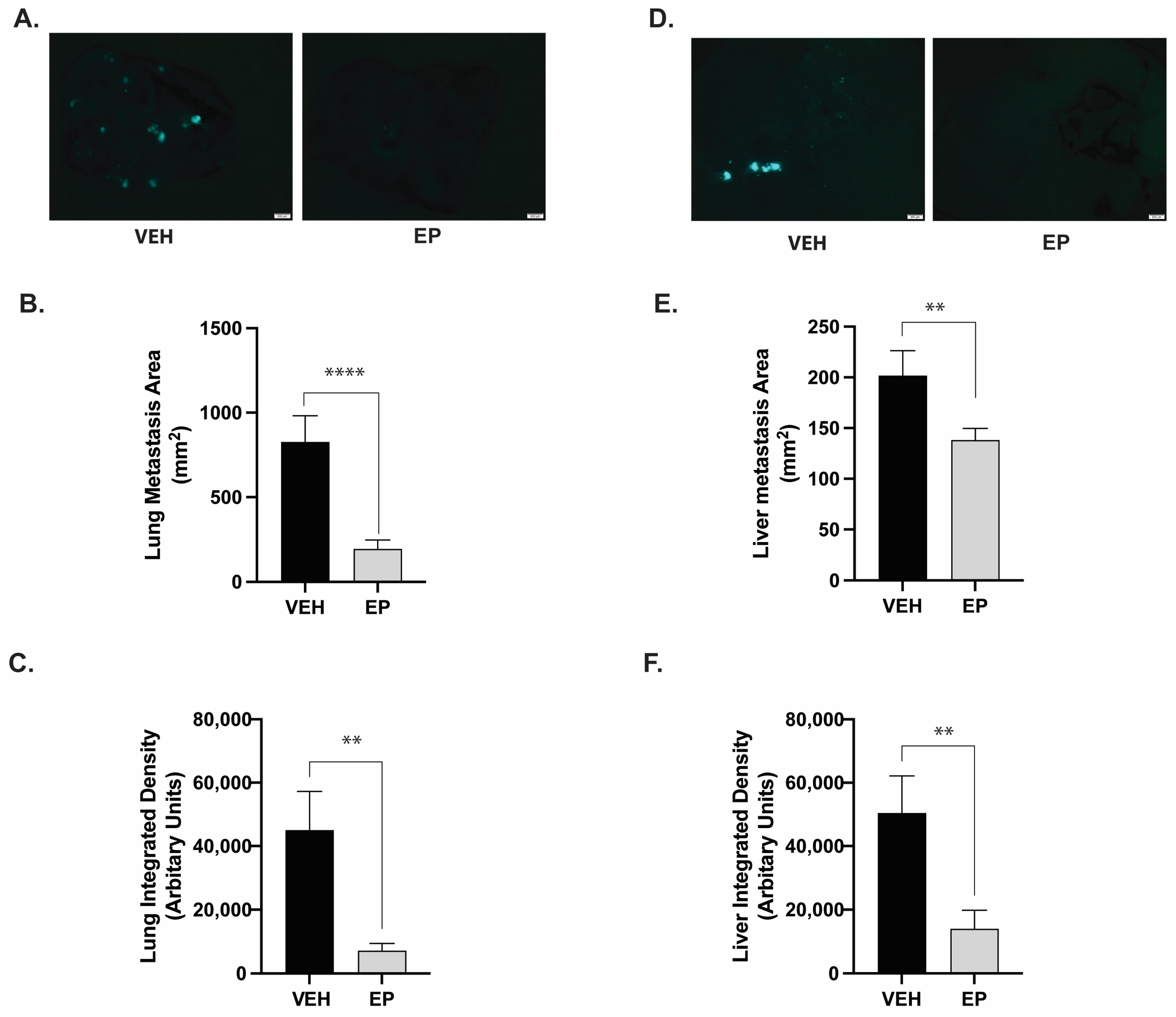
2.4. EP Significantly Reduces TNBC Metastasis in the Lungs and the Liver
3. Materials and Methods
3.1. Cell Culture and Reagents
3.2. JC-1 Mitochondrial Membrane Potential Assay
3.3. Tetramethylrhodamine Methyl Ester Perchlorate (TMRM) Assay
3.4. Seahorse XF Mito Stress Test
3.5. Immunoblots
3.6. Immunocytochemistry
3.7. In Vivo Study
3.7.1. Tumor Model
3.7.2. Treatment Administration
3.7.3. Analysis of Metastases
3.8. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EP | ergosterol peroxide |
| EP-SA | ergosterol peroxide sulfonamide |
| BC | breast cancer |
| TNBC | triple-negative breast cancer |
| ROS | reactive oxygen species |
| MMP | mitochondrial membrane potential |
| ER | estrogen receptor |
| PR | progesterone receptor |
| HER2 | human epidermal growth factor receptor |
| TMRM | tetramethylrodhamine methyl ester |
| mtDNA | mitochondrial DNA |
| FCCP | trifluoromethoxy carbonylcyanide phenylhydrazone |
| OCR | oxygen consumption rate |
| ECAR | extracellular acidification rates |
| 2-DG | 2-deoxyglucose |
| OXPHOS | oxidative phosphorylation |
| SHO-SCID | hairless outbred severe combined immunodeficient |
| BW | body weight |
| AST | aspartate aminotransferase |
| ALT | alanine aminotransferase |
| Tx | treatment |
| GLE | Ganoderma lucidum extract |
| CSCs | cancer stem cells |
| ALDH1 | aldehyde dehydrogenase 1 |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| FBS | fetal bovine serum |
| PBS | phosphate-buffered saline |
| hiPSCs | human-induced pluripotent stem cells |
| cTNT | cardiac troponin T |
| cTNI | cardiac troponin I |
| BSA | bovine serum albumin |
| RT | room temperature |
| LR | likelihood ratio |
| NIGMS | National Institutes of Health-National Institute of General Medical Sciences |
| NSF | National Science Foundation |
| CIAS | Common Instrumentation Area & Services |
| OADRGS | Office of the Associate Dean for Research and Graduate Studies |
| STCE | Science and Technology Competency Enhancement |
| INBRE | IDeA Networks of Biomedical Research Excellence |
References
- American Cancer Society. Cancer Facts & Figures 2024; American Cancer Society: Atlanta, GA, USA, 2024. [Google Scholar]
- Cuthrell, K.M.; Tzenios, N. Breast Cancer: Updated and Deep Insights. Int. Res. J. Oncol. 2023, 6, 104–118. [Google Scholar]
- Obidiro, O.; Battogtokh, G.; Akala, E.O. Triple Negative Breast Cancer Treatment Options and Limitations: Future Outlook. Pharmaceutics 2023, 15, 1796. [Google Scholar] [CrossRef] [PubMed]
- Orrantia-Borunda, E.; Anchondo-Nuñez, P.; Acuña-Aguilar, L.E.; Gómez-Valles, F.O.; Ramírez-Valdespino, C.A. Subtypes of Breast Cancer. In Breast Cancer; Mayrovitz, H.N., Ed.; Exon Publications: Brisbane, Australia, 2022. [Google Scholar]
- Lee, A.; Djamgoz, M.B.A. Triple negative breast cancer: Emerging therapeutic modalities and novel combination therapies. Cancer Treat. Rev. 2018, 62, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Gupta, S.; Chandra Gupta, S. Editorial: Targeting triple negative breast cancer by natural compounds. Front. Pharmacol. 2023, 14, 1172245. [Google Scholar] [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar]
- Weinberg, S.E.; Chandel, N.S. Targeting mitochondria metabolism for cancer therapy. Nat. Chem. Biol. 2015, 11, 9–15. [Google Scholar] [CrossRef]
- Martínez-Montemayor, M.M.; Ling, T.; Suárez-Arroyo, I.J.; Ortiz-Soto, G.; Santiago-Negrón, C.L.; Lacourt-Ventura, M.Y.; Valentín-Acevedo, A.; Lang, W.H.; Rivas, F. Identification of Biologically Active Ganoderma lucidum Compounds and Synthesis of Improved Derivatives That Confer Anti-cancer Activities in vitro. Front. Pharmacol. 2019, 10, 115. [Google Scholar] [CrossRef]
- Popgeorgiev, N.; Gil, C.; Berthenet, K.; Bertolin, G.; Ichim, G. Shedding light on mitochondrial outer-membrane permeabilization and membrane potential: State of the art methods and biosensors. Semin. Cell Dev. Biol. 2024, 156, 58–65. [Google Scholar] [CrossRef]
- Yan, B.; Dong, L.; Neuzil, J. Mitochondria: An intriguing target for killing tumour-initiating cells. Mitochondrion 2016, 26, 86–93. [Google Scholar] [CrossRef]
- Barreto-Gamarra, C.; Domenech, M. Integrin stimulation by collagen I at the progenitor stage accelerates maturation of human iPSC-derived cardiomyocytes. J. Mol. Cell Cardiol. 2025, 201, 70–86. [Google Scholar] [CrossRef]
- Ling, T.; Lang, W.H.; Martinez-Montemayor, M.M.; Rivas, F. Development of ergosterol peroxide probes for cellular localisation studies. Org. Biomol. Chem. 2019, 17, 5223–5229. [Google Scholar] [CrossRef]
- Ling, T.; Arroyo-Cruz, L.V.; Smither, W.R.; Seighman, E.K.; Martínez-Montemayor, M.M.; Rivas, F. Early Preclinical Studies of Ergosterol Peroxide and Biological Evaluation of Its Derivatives. ACS Omega 2024, 9, 37117–37127. [Google Scholar] [CrossRef] [PubMed]
- Pelicano, H.; Zhang, W.; Liu, J.; Hammoudi, N.; Dai, J.; Xu, R.H.; Pusztai, L.; Huang, P. Mitochondrial dysfunction in some triple-negative breast cancer cell lines: Role of mTOR pathway and therapeutic potential. Breast Cancer Res. 2014, 16, 434. [Google Scholar] [CrossRef] [PubMed]
- Ghiraldeli, L.; Anderson, R.; Pladna, K.; Pardee, T.S. Adenosine Monophosphate Activated Protein Kinase (AMPK) enhances chemotherapy response in Acute Myeloid Leukemia (AML). Cancer Lett. 2022, 535, 215659. [Google Scholar] [CrossRef]
- Villanueva-Paz, M.; Cotán, D.; Garrido-Maraver, J.; Oropesa-Ávila, M.; de la Mata, M.; Delgado-Pavón, A.; de Lavera, I.; Alcocer-Gómez, E.; Álvarez-Córdoba, M.; Sánchez-Alcázar, J.A. AMPK Regulation of Cell Growth, Apoptosis, Autophagy, and Bioenergetics. Exp. Suppl. 2016, 107, 45–71. [Google Scholar]
- Georgoulis, A.; Vorgias, C.E.; Chrousos, G.P.; Rogakou, E.P. Genome Instability and γH2AX. Int. J. Mol. Sci. 2017, 18, 1979. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Bao, C.; Huang, L.; Wei, J.F. Current Therapeutic Strategies for Metastatic Triple-Negative Breast Cancer: From Pharmacists’ Perspective. J. Clin. Med. 2022, 11, 6021. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Castillo-Pichardo, L.; Martinez-Montemayor, M.M.; Martinez, J.E.; Wall, K.M.; Cubano, L.A.; Dharmawardhane, S. Inhibition of mammary tumor growth and metastases to bone and liver by dietary grape polyphenols. Clin. Exp. Metastasis 2009, 26, 505–516. [Google Scholar] [CrossRef]
- Ling, T.; Lang, W.H.; Craig, J.; Potts, M.B.; Budhraja, A.; Opferman, J.; Bollinger, J.; Maier, J.; Marsico, T.D.; Rivas, F. Studies of Jatrogossone A as a Reactive Oxygen Species Inducer in Cancer Cellular Models. J. Nat. Prod. 2019, 82, 1301–1311. [Google Scholar] [CrossRef]
- Rios-Fuller, T.; Ortiz-Soto, G.; Lacourt-Ventura, M.; Maldonado-Martinez, G.; Cubano, L.; Schneider, R.; Martinez-Montemayor, M. Ganoderma lucidum extract (GLE) impairs breast cancer stem cells by targeting the STAT3 pathway. Oncotarget 2018, 9, 35907–35921. [Google Scholar] [CrossRef]
- Cruz-Collazo, A.M.; Katsara, O.; Grafals-Ruiz, N.; Colon Gonzalez, J.; Dorta-Estremera, S.; Carlo, V.P.; Chorna, N.; Schneider, R.J.; Dharmawardhane, S. Novel Inhibition of Central Carbon Metabolism Pathways by Rac and CDC42 inhibitor MBQ167 and Paclitaxel. Mol. Cancer Ther. 2024, 23, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nat. Protoc. 2013, 8, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Diaz, A.; Ortiz-Soto, G.; Suarez-Arroyo, I.J.; Zayas-Santiago, A.; Martinez Montemayor, M.M. Ganoderma lucidum Extract Reduces the Motility of Breast Cancer Cells Mediated by the RAC–Lamellipodin Axis. Nutrients 2019, 11, 1116. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Montemayor, M.M.; Acevedo, R.R.; Otero-Franqui, E.; Cubano, L.A.; Dharmawardhane, S.F. Ganoderma lucidum (Reishi) inhibits cancer cell growth and expression of key molecules in inflammatory breast cancer. Nutr. Cancer 2011, 63, 1085–1094. [Google Scholar] [CrossRef]
- Suarez-Arroyo, I.J.; Rosario-Acevedo, R.; Aguilar-Perez, A.; Clemente, P.L.; Cubano, L.A.; Serrano, J.; Schneider, R.J.; Martinez-Montemayor, M.M. Anti-tumor effects of Ganoderma lucidum (reishi) in inflammatory breast cancer in in vivo and in vitro models. PLoS ONE 2013, 8, e57431. [Google Scholar] [CrossRef]
- Suarez-Arroyo, I.J.; Rios-Fuller, T.J.; Feliz-Mosquea, Y.R.; Lacourt-Ventura, M.; Leal-Alviarez, D.J.; Maldonado-Martinez, G.; Cubano, L.A.; Martinez-Montemayor, M.M. Ganoderma lucidum Combined with the EGFR Tyrosine Kinase Inhibitor, Erlotinib Synergize to Reduce Inflammatory Breast Cancer Progression. J. Cancer 2016, 7, 500–511. [Google Scholar] [CrossRef]
- Suárez-Arroyo, I.J.; Acevedo-Díaz, A.; Ríos-Fuller, T.J.; Ortiz-Soto, G.; Vallejo-Calzada, R.; Reyes-Chea, J.; Maldonado-Martínez, G.; Schneider, R.J.; Martínez-Montemayor, M.M. Ganoderma lucidum enhances carboplatin chemotherapy effect by inhibiting the DNA damage response pathway and stemness. Am. J. Cancer Res. 2022, 12, 1282–1294. [Google Scholar]
- Ortiz-Soto, G.; Babilonia-Díaz, N.S.; Lacourt-Ventura, M.Y.; Rivera-Rodríguez, D.M.; Quiñones-Rodríguez, J.I.; Colón-Vargas, M.; Almodóvar-Rivera, I.; Ferrer-Torres, L.E.; Suárez-Arroyo, I.J.; Martínez-Montemayor, M.M. Metadherin Regulates Inflammatory Breast Cancer Invasion and Metastasis. Int. J. Mol. Sci. 2023, 24, 4694. [Google Scholar] [CrossRef]
- Reyes-Ramos, A.M.; Ramos-Cruz, K.P.; Rodriguez-Merced, N.J.; Martinez-Montemayor, M.M.; Franqui-Rios, N.D.; Rios-Grant, J.P.; Flores, A.; Maldonado-Martinez, G.; Torres-Garcia, W.; Domenech, M. Mesenchymal Cells Support the Oncogenicity and Therapeutic Response of the Hedgehog Pathway in Triple-Negative Breast Cancer. Cancers 2019, 11, 1522. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocachica-Adorno, A.L.; Aponte-Ramos, A.Y.; Rivera-Fuentes, P.S.; Espinosa-Ponce, N.P.; Arroyo-Cruz, L.V.; Ling, T.; Pérez-Ríos, N.; Rivas-Tumanyan, S.; Almodóvar-Rivera, I.A.; Barreto-Gamarra, C.; et al. Ergosterol Peroxide Disrupts Triple-Negative Breast Cancer Mitochondrial Function and Inhibits Tumor Growth and Metastasis. Int. J. Mol. Sci. 2025, 26, 4588. https://doi.org/10.3390/ijms26104588
Bocachica-Adorno AL, Aponte-Ramos AY, Rivera-Fuentes PS, Espinosa-Ponce NP, Arroyo-Cruz LV, Ling T, Pérez-Ríos N, Rivas-Tumanyan S, Almodóvar-Rivera IA, Barreto-Gamarra C, et al. Ergosterol Peroxide Disrupts Triple-Negative Breast Cancer Mitochondrial Function and Inhibits Tumor Growth and Metastasis. International Journal of Molecular Sciences. 2025; 26(10):4588. https://doi.org/10.3390/ijms26104588
Chicago/Turabian StyleBocachica-Adorno, Aliyah L., Adriana Y. Aponte-Ramos, Paola S. Rivera-Fuentes, Natalia P. Espinosa-Ponce, Luz V. Arroyo-Cruz, Taotao Ling, Naydi Pérez-Ríos, Sona Rivas-Tumanyan, Israel A. Almodóvar-Rivera, Carlos Barreto-Gamarra, and et al. 2025. "Ergosterol Peroxide Disrupts Triple-Negative Breast Cancer Mitochondrial Function and Inhibits Tumor Growth and Metastasis" International Journal of Molecular Sciences 26, no. 10: 4588. https://doi.org/10.3390/ijms26104588
APA StyleBocachica-Adorno, A. L., Aponte-Ramos, A. Y., Rivera-Fuentes, P. S., Espinosa-Ponce, N. P., Arroyo-Cruz, L. V., Ling, T., Pérez-Ríos, N., Rivas-Tumanyan, S., Almodóvar-Rivera, I. A., Barreto-Gamarra, C., Domenech-García, M., Rivas, F., & Martínez-Montemayor, M. M. (2025). Ergosterol Peroxide Disrupts Triple-Negative Breast Cancer Mitochondrial Function and Inhibits Tumor Growth and Metastasis. International Journal of Molecular Sciences, 26(10), 4588. https://doi.org/10.3390/ijms26104588