
LncRNA-Mediated Tissue-Specific Plastic Responses to Salinity Changes in Oysters

Abstract
1. Introduction
2. Results
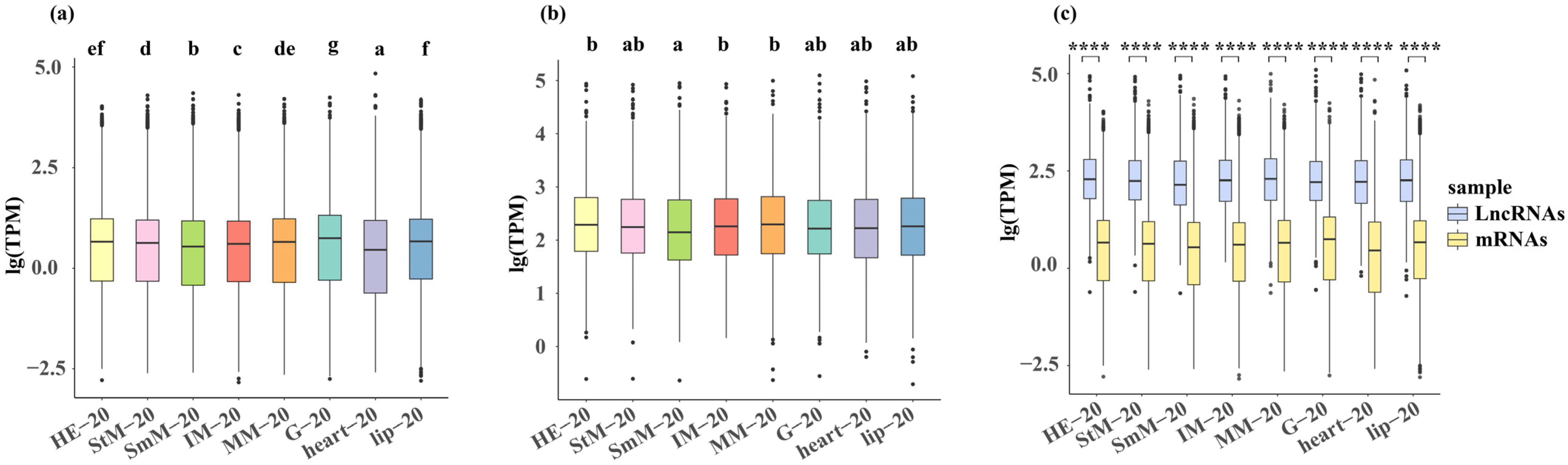
2.1. Differences in the Response to Salinity Among the Eight Tissues
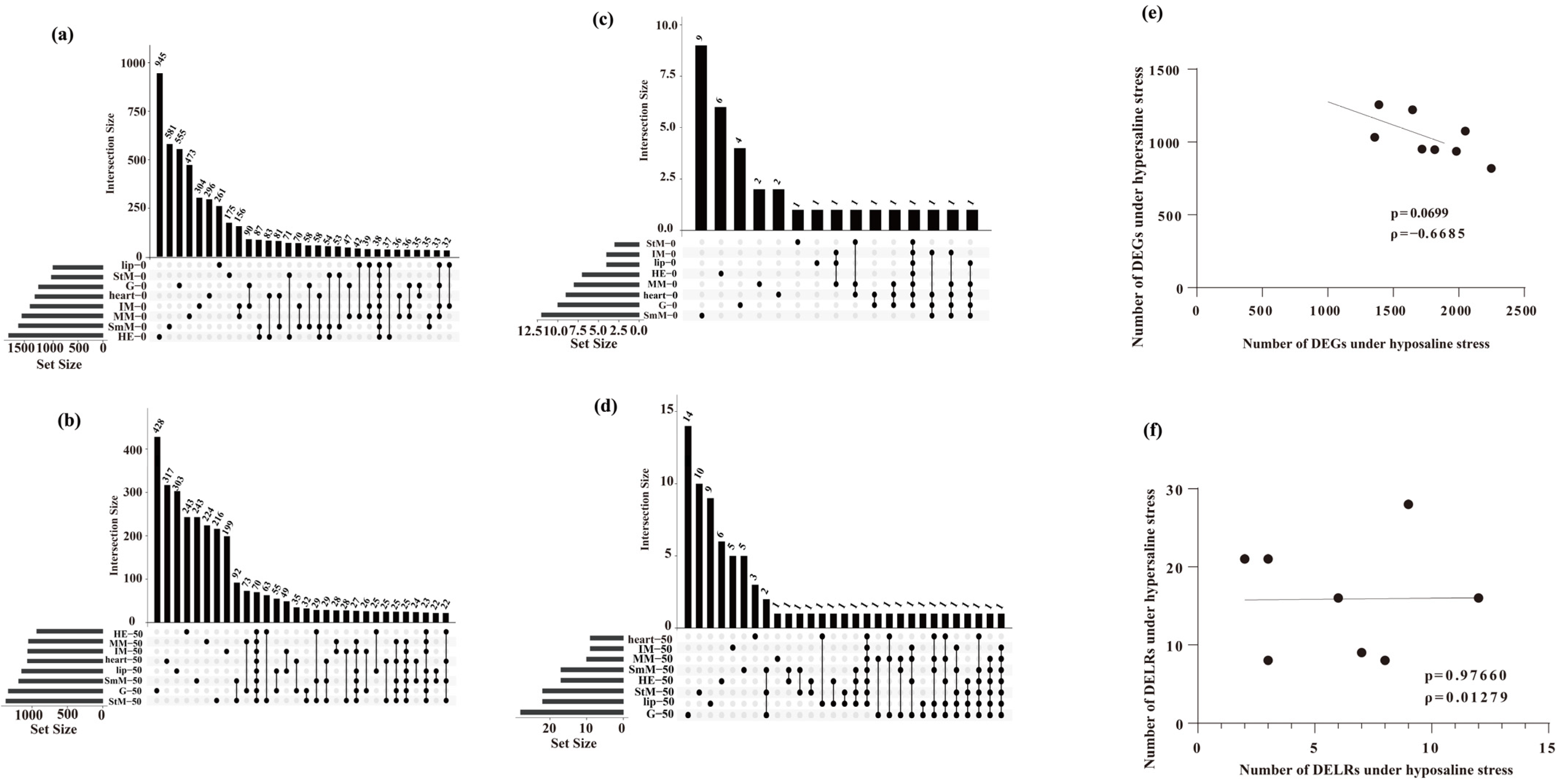
2.2. DEGs and DELRs in Response to Hyper- and Hypo-Saline Stresses
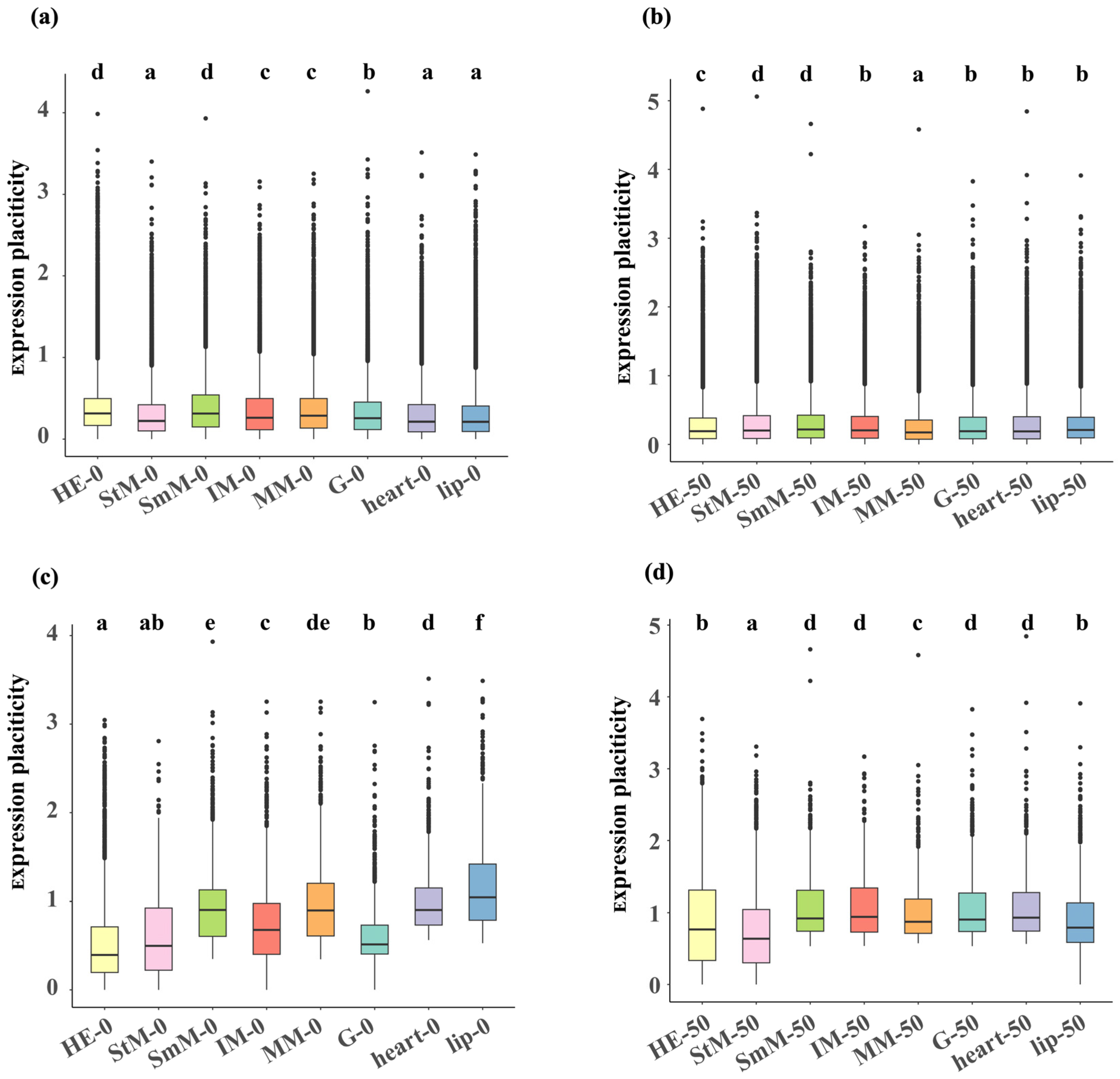
2.3. Transcriptional Changes in Genome-Wide Genes and DEGs
2.4. Expression Patterns of mRNAs and lncRNAs in HE and StM
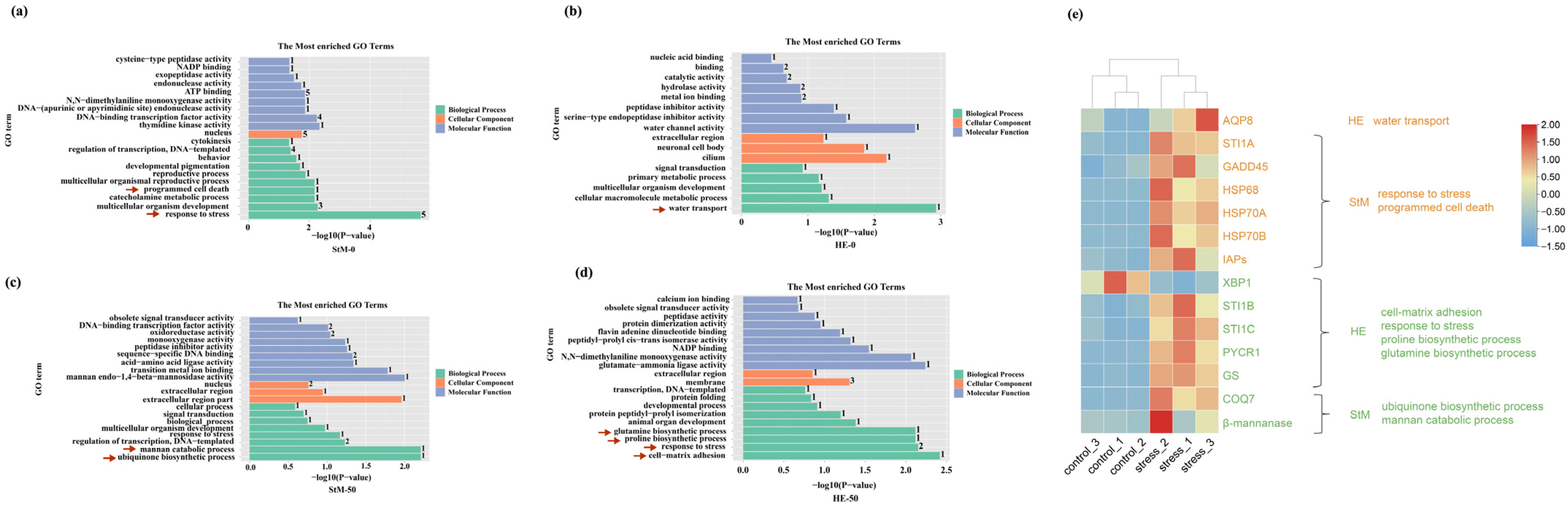
2.5. Biological Processes of HE and StM upon Hypo- and Hyper-Saline Conditions
3. Discussion
4. Materials and Methods
4.1. Oyster Samples
4.2. RNA-Sequencing
4.3. Identification of lncRNAs
4.4. Differential Expression Analyses and Cluster Analysis
4.5. Transcriptional Changes in Genes upon Salinity Stresses
4.6. Construction of lncRNA-mRNA Co-Expression Network and Functional Enrichment Analyses
4.7. High-Throughput Chromosome Conformation Capture (Hi-C) and Assay for Transposase-Accessible Chromatin Sequencing (ATAC-Seq)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McLusky, D.S.; Hagerman, L.; Mitchell, P. Effect of salinity acclimation on osmoregulation in Crangon crangon and Praunus flexuosus. Ophelia 1982, 21, 89–100. [Google Scholar] [CrossRef]
- Gong, J.; Li, Q.; Yu, H.; Liu, S.; Kong, L. Effects of low salinity on hemolymph osmolality and transcriptome of the Iwagaki oyster, Crassostrea nippona. Fish Shellfish Immunol. 2022, 126, 211–216. [Google Scholar] [CrossRef]
- Johnson, C.R.; Banks, S.C.; Barrett, N.S.; Cazassus, F.; Dunstan, P.K.; Edgar, G.J.; Frusher, S.D.; Gardner, C.; Haddon, M.; Helidoniotis, F.; et al. Climate change cascades: Shifts in oceanography, species’ ranges and subtidal marine community dynamics in eastern Tasmania. J. Exp. Mar. Biol. Ecol. 2011, 400, 17–32. [Google Scholar] [CrossRef]
- Philippart, C.J.M.; Anadón, R.; Danovaro, R.; Dippner, J.W.; Drinkwater, K.F.; Hawkins, S.J.; Oguz, T.; O’Sullivan, G.; Reid, P.C. Impacts of climate change on European marine ecosystems: Observations, expectations and indicators. J. Exp. Mar. Biol. Ecol. 2011, 400, 52–69. [Google Scholar] [CrossRef]
- Gienapp, P.; Teplitsky, C.; Alho, J.S.; Mills, J.A.; Merilä, J. Climate change and evolution: Disentangling environmental and genetic responses. Mol. Ecol. 2008, 17, 167–178. [Google Scholar] [CrossRef]
- Kenkel, C.D.; Matz, M.V. Gene expression plasticity as a mechanism of coral adaptation to a variable environment. Nat. Ecol. Evol. 2017, 1, 14. [Google Scholar] [CrossRef]
- Li, L.; Li, A.; Song, K.; Meng, J.; Guo, X.; Li, S.; Li, C.; De Wit, P.; Que, H.; Wu, F.; et al. Divergence and plasticity shape adaptive potential of the Pacific oyster. Nat. Ecol. Evol. 2018, 2, 1751–1760. [Google Scholar] [CrossRef]
- Chevin, L.-M.; Lande, R.; Mace, G.M. Adaptation, Plasticity, and Extinction in a Changing Environment: Towards a Predictive Theory. PLoS Biol. 2010, 8, e1000357. [Google Scholar] [CrossRef]
- Li, A.; Zhao, M.; Zhang, Z.; Wang, C.; Zhang, K.; Zhang, X.; De Wit, P.R.; Wang, W.; Gao, J.; Guo, X.; et al. Genome architecture and selective signals compensatorily shape plastic response to a new environment. Innovation 2023, 4, 100464. [Google Scholar] [CrossRef]
- Li, A.; Li, L.; Zhang, Z.; Li, S.; Wang, W.; Guo, X.; Zhang, G.; Saitou, N. Noncoding variation and transcriptional plasticity promote thermal adaptation in oysters by altering energy metabolism. Mol. Biol. Evol. 2021, 38, 5144–5155. [Google Scholar] [CrossRef]
- Li, A.; Dai, H.; Guo, X.; Zhang, Z.; Zhang, K.; Wang, C.; Wang, X.; Wang, W.; Chen, H.; Li, X.; et al. Genome of the estuarine oyster provides insights into climate impact and adaptive plasticity. Commun. Biol. 2021, 4, 1287. [Google Scholar] [CrossRef]
- Fischer, E.K.; Ghalambor, C.K.; Hoke, K.L. Can a network approach resolve how adaptive vs nonadaptive plasticity impacts evolutionary trajectories? Integr. Comp. Biol. 2016, 56, 877–888. [Google Scholar] [CrossRef]
- Pfennig, D.W.; Wund, M.A.; Snell-Rood, E.C.; Cruickshank, T.; Schlichting, C.D.; Moczek, A.P. Phenotypic plasticity’s impacts on diversification and speciation. Trends Ecol. Evol. 2010, 25, 459–467. [Google Scholar] [CrossRef]
- Wheaton, F.; Hall, S. Research needs for automated oyster shucking. Aquac. Eng. 2007, 37, 67–72. [Google Scholar] [CrossRef]
- Zhang, K.; Li, A.; Qi, H.; Yang, Q.; Du, M.; Wang, X.; Zhang, Z.; Wang, C.; Wang, W.; Zhang, G.; et al. The development of a 30 K SNP genotyping tool targeting genomic regions of temperature and salinity adaptation in estuarine oyster. Aquaculture 2023, 566, 739168. [Google Scholar] [CrossRef]
- She, Z.; Li, L.; Meng, J.; Jia, Z.; Que, H.; Zhang, G. Population resequencing reveals candidate genes associated with salinity adaptation of the Pacific oyster Crassostrea gigas. Sci. Rep. 2018, 8, 8683. [Google Scholar] [CrossRef] [PubMed]
- Dineshram, R.; Chandramouli, K.; Ko, G.W.; Zhang, H.; Qian, P.Y.; Ravasi, T.; Thiyagarajan, V. Quantitative analysis of oyster larval proteome provides new insights into the effects of multiple climate change stressors. Glob. Change Biol. 2016, 22, 2054–2068. [Google Scholar] [CrossRef]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, Q.; Du, M.; Zhang, Z.; Wang, W.; Zhang, G.; Li, A.; Li, L. Genome-wide mapping of regulatory variants for temperature- and salinity-adaptive genes reveals genetic basis of genotype-by-environment interaction in Crassostrea ariakensis. Environ. Res. 2023, 236, 116614. [Google Scholar] [CrossRef]
- Pérez–Velasco, R.; Manzano–Sarabia, M.; Hurtado–Oliva, M.Á. Effect of hypo–and hypersaline stress conditions on physiological, metabolic, and immune responses in the oyster Crassostrea corteziensis (Bivalvia: Ostreidae). Fish Shellfish Immunol. 2022, 120, 252–260. [Google Scholar] [CrossRef]
- Zhao, M.; Li, A.; Zhang, K.; Wang, W.; Zhang, G.; Li, L. The role of the balance between energy production and ammonia detoxification mediated by key amino acids in divergent hypersaline adaptation among Crassostrea oysters. Environ. Res. 2024, 248, 118213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, A.; She, Z.; Wang, X.; Jia, Z.; Wang, W.; Zhang, G.; Li, L. Adaptive divergence and underlying mechanisms in response to salinity gradients between two Crassostrea oysters revealed by phenotypic and transcriptomic analyses. Evol. Appl. 2023, 16, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, L.; Li, A.; Li, Y.; Wang, W.; Zhang, G. Transcriptome and gene coexpression network analyses of two wild populations provides insight into the high-salinity adaptation mechanisms of Crassostrea ariakensis. Mar. Biotechnol. 2019, 21, 596–612. [Google Scholar] [CrossRef]
- Meng, J.; Zhu, Q.; Zhang, L.; Li, C.; Li, L.; She, Z.; Huang, B.; Zhang, G. Genome and transcriptome analyses provide insight into the euryhaline adaptation mechanism of Crassostrea gigas. PLoS ONE 2013, 8, e58563. [Google Scholar] [CrossRef] [PubMed]
- Eierman, L.E.; Hare, M.P. Reef-specific patterns of gene expression plasticity in eastern oysters (Crassostrea virginica). J. Hered. 2016, 107, 90–100. [Google Scholar] [CrossRef]
- Zhao, X.; Yu, H.; Kong, L.; Liu, S.; Li, Q. Comparative transcriptome analysis of two oysters, Crassostrea gigas and Crassostrea hongkongensis provides insights into adaptation to hypo-osmotic conditions. PLoS ONE 2014, 9, e111915. [Google Scholar] [CrossRef]
- Pourmozaffar, S.; Tamadoni Jahromi, S.; Rameshi, H.; Sadeghi, A.; Bagheri, T.; Behzadi, S.; Gozari, M.; Zahedi, M.R.; Abrari Lazarjani, S. The role of salinity in physiological responses of bivalves. Rev. Aquac. 2019, 12, 1548–1566. [Google Scholar] [CrossRef]
- Wang, H.; Guo, X.; Zhang, G.; Zhang, F. Classification of jinjiang oysters Crassostrea rivularis (Gould, 1861) from China, based on morphology and phylogenetic analysis. Aquaculture 2004, 242, 137–155. [Google Scholar] [CrossRef]
- Zhou, M.F.; Standish, K., Jr. A review of published work on Crassostrea ariakensis. J. Shellfish Res. 2003, 22, 1–20. [Google Scholar]
- Jiang, W.; Chen, L. Tissue specificity of gene expression evolves across mammal species. J. Comput. Biol. 2022, 29, 880–891. [Google Scholar] [CrossRef]
- Pan, B.; Wang, Y.; Li, D.; Wang, T.; Du, L. Tissue-specific distribution and bioaccumulation pattern of trace metals in fish species from the heavily sediment-laden Yellow River, China. J. Hazard. Mater. 2022, 425, 128050. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Zhang, Z.; Zhao, X.; Geng, H.; Xue, L.; Liu, D. Spatial distribution and enrichment dynamics of foodborne norovirus in oyster tissues. Foods 2023, 13, 128. [Google Scholar] [CrossRef] [PubMed]
- Barshir, R.; Shwartz, O.; Smoly, I.Y.; Yeger-Lotem, E. Comparative analysis of human tissue interactomes reveals factors leading to tissue-specific manifestation of hereditary diseases. PLoS Comput. Biol. 2014, 10, e1003632. [Google Scholar] [CrossRef]
- Gibcus, J.H.; Dekker, J. The hierarchy of the 3D genome. Mol. Cell 2013, 49, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Hamba, Y.; Kamatani, T.; Miya, F.; Boroevich, K.A.; Tsunoda, T. Topologically associating domain underlies tissue specific expression of long intergenic non-coding RNAs. iScience 2023, 26, 106640. [Google Scholar] [CrossRef]
- Marques, A.C.; Hughes, J.; Graham, B.; Kowalczyk, M.S.; Higgs, D.R.; Ponting, C.P. Chromatin signatures at transcriptional start sites separate two equally populated yet distinct classes of intergenic long noncoding RNAs. Genome Biol. 2013, 14, R131. [Google Scholar] [CrossRef]
- Mantica, F.; Iniguez, L.P.; Marquez, Y.; Permanyer, J.; Torres-Mendez, A.; Cruz, J.; Franch-Marro, X.; Tulenko, F.; Burguera, D.; Bertrand, S.; et al. Evolution of tissue-specific expression of ancestral genes across vertebrates and insects. Nat. Ecol. Evol. 2024, 8, 1140–1153. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, Y.; Zou, C.; Yang, C.; Pan, G.; Ma, L.; Shen, Y. Integrated analysis of long non-coding RNAs and mRNAs reveals the regulatory network of maize seedling root responding to salt stress. BMC Genom. 2022, 23, 50. [Google Scholar] [CrossRef]
- Baruah, P.M.; Krishnatreya, D.B.; Bordoloi, K.S.; Gill, S.S.; Agarwala, N. Genome wide identification and characterization of abiotic stress responsive lncRNAs in Capsicum annuum. Plant Physiol. Biochem. 2021, 162, 221–236. [Google Scholar] [CrossRef]
- de Goede, O.M.; Nachun, D.C.; Ferraro, N.M.; Gloudemans, M.J.; Rao, A.S.; Smail, C.; Eulalio, T.Y.; Aguet, F.; Ng, B.; Xu, J.; et al. Population-scale tissue transcriptomics maps long non-coding RNAs to complex disease. Cell 2021, 184, 2633–2648.e19. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Author correction: Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 159. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, J.A.-O.; Terai, G.; Hamada, M. Computational prediction of lncRNA-mRNA interactionsby integrating tissue specificity in human transcriptome. Biol. Direct 2017, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Flower, C.T.; Chen, L.; Jung, H.J.; Raghuram, V.; Knepper, M.A.; Yang, C.R. An integrative proteogenomics approach reveals peptides encoded by annotated lincRNA in the mouse kidney inner medulla. Physiol. Genom. 2020, 52, 485–491. [Google Scholar] [CrossRef]
- Much, C.; Lasda, E.L.; Pereira, I.T.; Vallery, T.K.; Ramirez, D.; Lewandowski, J.P.; Dowell, R.D.; Smallegan, M.J.; Rinn, J.L. The temporal dynamics of lncRNA Firre-mediated epigenetic and transcriptional regulation. Nat. Commun. 2024, 15, 6821. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, F.; Wang, H.; Teschendorff, A.E.; Xie, F.; He, Y. Pan-cancer characterization of long non-coding RNA and DNA methylation mediated transcriptional dysregulation. eBioMedicine 2021, 68, 103399. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jin, S.; Lin, L.; Yang, Q.; Jiang, H. Changes in the long noncoding RNA expression profile in the development of the embryonic external ear after BMP5 gene mutation. J. Craniofac. Surg. 2023, 34, 1605–1609. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, L.; Zhong, Y.; Cai, M.; Gao, J.; Tan, C.; Han, X.; Guo, R.; Han, L. LncRNA H19 over-expression inhibited Th17 cell differentiation to relieve endometriosis through miR-342-3p/IER3 pathway. Cell Biosci. 2019, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Wang, Q.; Liu, Y.L.; Xiang, Z.; Wang, Q.Q.; Yin, L.; Liu, S.L. LncRNA POU3F3 contributes to dacarbazine resistance of human melanoma through the MiR-650/MGMT axis. Front. Oncol. 2021, 11, 643613. [Google Scholar] [CrossRef]
- Zhu, M.; Dong, Q.; Bing, J.; Songbuerbatu; Zheng, L.; Dorjee, T.; Liu, Q.; Zhou, Y.; Gao, F. Combined lncRNA and mRNA expression profiles identified the lncRNA-miRNA-mRNA modules regulating the cold stress response in Ammopiptanthus nanus. Int. J. Mol. Sci. 2023, 24, 6502. [Google Scholar] [CrossRef]
- Mirdar Mansuri, R.; Azizi, A.H.; Sadri, A.H.; Shobbar, Z.S. Long non-coding RNAs as the regulatory hubs in rice response to salt stress. Sci. Rep. 2022, 12, 21696. [Google Scholar] [CrossRef]
- Chen, L.; Shi, S.; Jiang, N.; Khanzada, H.; Wassan, G.M.; Zhu, C.; Peng, X.; Xu, J.; Chen, Y.; Yu, Q.; et al. Genome-wide analysis of long non-coding RNAs affecting roots development at an early stage in the rice response to cadmium stress. BMC Genom. 2018, 19, 460. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Long, X.; Zhang, X.; Dong, X.; Wang, Z.; Jiang, H.; An, M.; Chen, J.; Gan, L. Identification and characterization of long non-coding RNAs in intestinal immune regulation of largemouth bass, Micropterus salmoides, under acute heat stress. Comp. Biochem. Physiol. Part D Genom. Proteom. 2023, 48, 101132. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Y.; Qi, G. LncRNA-IRAR-mediated regulation of insulin receptor transcripts in Drosophila melanogaster during nutritional stress. Insect Mol. Biol. 2022, 31, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, H.; Li, Q.; Liu, S. Transcription analysis for core networks of lncRNAs-mRNAs: Implication for potential role in sterility of Crassostrea gigas. Biology 2022, 11, 378. [Google Scholar] [CrossRef]
- Feng, D.; Li, Q.; Yu, H.; Kong, L.; Du, S. Transcriptional profiling of long non-coding RNAs in mantle of Crassostrea gigas and their association with shell pigmentation. Sci. Rep. 2018, 8, 1436. [Google Scholar] [CrossRef]
- Song, K. Genome-wide identification of long non-coding RNAs in Crassostrea gigas and their association with heat stress. Mar. Biotechnol. 2022, 24, 744–752. [Google Scholar] [CrossRef]
- Peng, M.; Cardoso, J.C.R.; Pearson, G.; Vm Canário, A.; Power, D.M. Core genes of biomineralization and cis-regulatory long non-coding RNA regulate shell growth in bivalves. J. Adv. Res. 2024, 64, 117–129. [Google Scholar] [CrossRef]
- Zheng, Z.; Xie, B.; Cai, W.; Yang, C.; Du, X. Identification of a long non-coding RNA (LncMSEN2) from pearl oyster and its potential roles in exoskeleton formation and LPS stimulation. Fish Shellfish Immunol. 2020, 103, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; He, Q.; Xie, B.; Xu, Z.; Wang, C.; Yang, C.; Liao, Y.; Zheng, Z. Long non-coding RNA LncMPEG1 responds to multiple environmental stressors by affecting biomineralization in pearl oyster Pinctada fucata martensii. Front. Mar. Sci. 2022, 9, 1014810. [Google Scholar] [CrossRef]
- Kurihara, M.; Otsuka, K.; Matsubara, S.; Shiraishi, A.; Satake, H.; Kimura, A.P. A Testis-Specific Long Non-Coding RNA, lncRNA-Tcam1, Regulates Immune-Related Genes in Mouse Male Germ Cells. Front. Endocrinol. 2017, 8, 229. [Google Scholar] [CrossRef]
- Gong, J.; Li, Q. Comparative transcriptome and WGCNA analysis reveal molecular responses to salinity change in larvae of the iwagaki oyster Crassostrea Nippona. Mar. Biotechnol. 2023, 25, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, C.; Wang, W.; Zhang, Z.; Liu, M.; She, Z.; Jia, Z.; Zhang, G.; Li, L. Molecular and fitness data reveal local adaptation of southern and northern estuarine oysters (Crassostrea ariakensis). Front. Mar. Sci. 2020, 7, 589099. [Google Scholar] [CrossRef]
- List, E.O.; Berryman, D.E.; Jensen, E.A.; Kulkarni, P.; McKenna, S.; Kopchick, J.J. New insights of growth hormone (GH) actions from tissue-specific GH receptor knockouts in mice. Arch. Endocrinol. Metab. 2019, 63, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Widman, N.; Feng, S.; Jacobsen, S.E.; Pellegrini, M. Epigenetic differences between shoots and roots in Arabidopsis reveals tissue-specific regulation. Epigenetics 2014, 9, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Modrek, B.; Lee, C. Genome-wide detection of tissue-specific alternative splicing in the human transcriptome. Nucleic Acids Res. 2002, 30, 3754–3766. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Nong, Q.; Solanki, M.K.; Liang, Q.; Xie, J.; Liu, X.; Li, Y.; Wang, W.; Yang, L.; Li, Y. Differential expression profiles and pathways of genes in sugarcane leaf at elongation stage in response to drought stress. Sci. Rep. 2016, 6, 25698. [Google Scholar] [CrossRef]
- Yao, H.; Liang, Z.; Wang, W.; Niu, C. Integrative analyses of transcriptomes and metabolomes provide insight into salinity adaption in Bangia (Rhodaphyta). Int. J. Biol. Macromol. 2023, 253, 127466. [Google Scholar] [CrossRef]
- Dupont-Prinet, A.; Chatain, B.; Grima, L.; Vandeputte, M.; Claireaux, G.; McKenzie, D.J. Physiological mechanisms underlying a trade-off between growth rate and tolerance of feed deprivation in the European sea bass (Dicentrarchus labrax). J. Exp. Biol. 2010, 213, 1143–1152. [Google Scholar] [CrossRef]
- Wang, C.; Li, A.; Wang, W.; Cong, R.; Wang, L.; Zhang, G.; Li, L. Integrated application of transcriptomics and metabolomics reveals the energy allocation-mediated mechanisms of growth-defense trade-offs in Crassostrea gigas and Crassostrea angulata. Front. Mar. Sci. 2021, 8, 744626. [Google Scholar] [CrossRef]
- Li, A.; Li, L.; Wang, W.; Song, K.; Zhang, G.F. Transcriptomics and fitness data reveal adaptive plasticity of thermal tolerance in oysters inhabiting different tidal zones. Front. Physiol. 2018, 9, 825. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Li, L.; Song, K.; Wang, W.; Zhang, G. Temperature, energy metabolism, and adaptive divergence in two oyster subspecies. Ecol. Evol. 2017, 7, 6151–6162. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Li, L.; Wang, W.; Zhang, G. Evolutionary trade-offs between baseline and plastic gene expression in two congeneric oyster species. Biol. Lett. 2019, 15, 20190202. [Google Scholar] [CrossRef] [PubMed]
- Murren, C.J.; Auld, J.R.; Callahan, H.; Ghalambor, C.K.; Handelsman, C.A.; Heskel, M.A.; Kingsolver, J.G.; Maclean, H.J.; Masel, J.; Maughan, H.; et al. Constraints on the evolution of phenotypic plasticity: Limits and costs of phenotype and plasticity. Heredity 2015, 115, 293–301. [Google Scholar] [CrossRef]
- DeWitt, T.J.; Sih, A.; Wilson, D.S. Costs and limits of phenotypic plasticity. Trends Ecol. Evol. 1998, 13, 77–81. [Google Scholar] [CrossRef]
- Siljestam, M.; Östman, Ö. The combined effects of temporal autocorrelation and the costs of plasticity on the evolution of plasticity. J. Evol. Biol. 2017, 30, 1361–1371. [Google Scholar] [CrossRef]
- Li, Y.; Xu, C.; Li, Q. Physiological and gene expression responses of diploid and triploid Pacific oyster (Crassostrea gigas) to heat acclimation. Aquac. Res. 2022, 53, 6641–6650. [Google Scholar] [CrossRef]
- Ballantyne, J.S.; Berges, J.A. Enzyme activities of gill, hepatopancreas, mantle, and adductor muscle of the oyster (Crassostrea virginica) after changes in diet and salinity. Can. J. Fish. Aquat. Sci. 1991, 48, 1117–1123. [Google Scholar] [CrossRef]
- Tanimoto, S.; Kawakami, K.; Morimoto, S. Changes in the free amino acid content of the shucked oyster Crassostrea gigas stored in salt water at 3 °C. Fish. Aquat. Sci. 2013, 16, 63–69. [Google Scholar] [CrossRef]
- Mendela, T.S.; Isaac, S.R.; Enzor, L.A. Impacts of elevated temperature, decreased salinity and microfibers on the bioenergetics and oxidative stress in eastern oyster, Crassostrea virginica. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2024, 274, 111002. [Google Scholar] [CrossRef]
- Bresnahan, S.T.; Doke, M.A.; Giray, T.; Grozinger, C.M. Tissue-specific transcriptional patterns underlie seasonal phenotypes in honey bees (Apis mellifera). Mol. Ecol. 2022, 31, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Munro, T.; Mattick, J.S. The potential of long noncoding RNA therapies. Trends Pharmacol. Sci. 2022, 43, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Huanca-Mamani, W.; Arias-Carrasco, R.; Cardenas-Ninasivincha, S.; Rojas-Herrera, M.; Sepulveda-Hermosilla, G.; Caris-Maldonado, J.C.; Bastias, E.; Maracaja-Coutinho, V. Long non-coding RNAs responsive to salt and boron stress in the hyper-arid lluteno maize from Atacama Desert. Genes 2018, 9, 170. [Google Scholar] [CrossRef]
- Hu, W.; Wang, G.; Wang, S.; Nie, X.; Wang, C.; Wang, Y.; Zhang, H.; Ji, W. Co-regulation of long non-coding RNAs with allele-specific genes in wheat responding to powdery mildew infection. Agronomy 2020, 10, 896. [Google Scholar] [CrossRef]
- Long, Y.; Wang, X.; Youmans, D.T.; Cech, T.R. How do lncRNAs regulate transcription? Sci. Adv. 2017, 3, eaao2110. [Google Scholar] [CrossRef]
- Groff, A.F.; Barutcu, A.R.; Lewandowski, J.P.; Rinn, J.L. Enhancers in the peril lincRNA locus regulate distant but not local genes. Genome Biol. 2018, 19, 219. [Google Scholar] [CrossRef]
- Wang, C.; Li, A.; Cong, R.; Qi, H.; Wang, W.; Zhang, G.; Li, L.; Wittkopp, P. Cis- and trans-variations of stearoyl-CoA desaturase provide new insights into the mechanisms of diverged pattern of phenotypic plasticity for temperature adaptation in two congeneric oyster species. Mol. Biol. Evol. 2023, 40, msad015. [Google Scholar] [CrossRef]
- Sanford, E.; Kelly, M.W. Local adaptation in marine invertebrates. Annu. Rev. Mar. Sci. 2011, 3, 509–535. [Google Scholar] [CrossRef]
- Somero, G. The physiology of global change: Linking patterns to mechanisms. Annu. Rev. Mar. Sci. 2012, 4, 39–61. [Google Scholar] [CrossRef]
- Zhang, G.; Li, L.; Meng, J.; Qi, H.; Qu, T.; Xu, F.; Zhang, L. Molecular basis for adaptation of oysters to stressful marine intertidal environments. Annu. Rev. Anim. Biosci. 2016, 4, 357–381. [Google Scholar] [CrossRef] [PubMed]
- Tomanek, L. Proteomics to study adaptations in marine organisms to environmental stress. J. Proteom. 2014, 105, 92–106. [Google Scholar] [CrossRef]
- Burford, M.O.; Scarpa, J.; Cook, B.J.; Hare, M.P. Local adaptation of a marine invertebrate with a high dispersal potential: Evidence from a reciprocal transplant experiment of the eastern oyster Crassostrea virginica. Mar. Ecol. Prog. Ser. 2014, 505, 161–175. [Google Scholar] [CrossRef]
- Berger, V.J.; Kharazova, A.D. Mechanisms of salinity adaptations in marine molluscs. Hydrobiologia 1997, 355, 115–126. [Google Scholar] [CrossRef]
- Richardson, C.A.; Collis, S.A.; Ekaratne, K.; Dare, P.; Key, D. The age determination and growth rate of the European fat oyster, Ostrea edulis, in British waters determined from acetate peels ofumbo growth lines. ICES J. Mar. Sci. 1993, 50, 493–500. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth Gk Fau–Shi, W.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.A.-O. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Pertea, G.; Pertea, M.A.-O. GFF Utilities: GffRead and GffCompare. F1000 Res. 2024, 9, 304. [Google Scholar] [CrossRef]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Zhang, J.; Zhou, Z. PLEK: A tool for predicting long non-coding RNAs and messenger RNAs based on an improved k-mer scheme. BMC Bioinform. 2014, 15, 311. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef]
- Bateman, A.; Birney, E.; Durbin, R.; Eddy, S.R.; Howe, K.L.; Sonnhammer, E.L.L. The Pfam Protein Families Database. Nucleic Acids Res. 2000, 28, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Alexa, A.; Rahnenführer, J.; Lengauer, T. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 2006, 22, 1600–1607. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DELRs | DEGs | |
|---|---|---|
| HE-0 | 6 | 767 |
| HE-50 | 16 | 628 |
| StM-0 | 2 | 336 |
| StM-50 | 21 | 987 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Zhao, J.; Li, A.; Zhao, M.; Huo, M.; Deng, J.; Wang, L.; Wang, W.; Zhang, G.; Li, L. LncRNA-Mediated Tissue-Specific Plastic Responses to Salinity Changes in Oysters. Int. J. Mol. Sci. 2025, 26, 4523. https://doi.org/10.3390/ijms26104523
Zhang M, Zhao J, Li A, Zhao M, Huo M, Deng J, Wang L, Wang W, Zhang G, Li L. LncRNA-Mediated Tissue-Specific Plastic Responses to Salinity Changes in Oysters. International Journal of Molecular Sciences. 2025; 26(10):4523. https://doi.org/10.3390/ijms26104523
Chicago/Turabian StyleZhang, Mengshi, Jinlong Zhao, Ao Li, Mingjie Zhao, Meitong Huo, Jinhe Deng, Luping Wang, Wei Wang, Guofan Zhang, and Li Li. 2025. "LncRNA-Mediated Tissue-Specific Plastic Responses to Salinity Changes in Oysters" International Journal of Molecular Sciences 26, no. 10: 4523. https://doi.org/10.3390/ijms26104523
APA StyleZhang, M., Zhao, J., Li, A., Zhao, M., Huo, M., Deng, J., Wang, L., Wang, W., Zhang, G., & Li, L. (2025). LncRNA-Mediated Tissue-Specific Plastic Responses to Salinity Changes in Oysters. International Journal of Molecular Sciences, 26(10), 4523. https://doi.org/10.3390/ijms26104523