

Preventive Effects of Resistance Training on Hemodynamics and Kidney Mitochondrial Bioenergetic Function in Ovariectomized Rats

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results
2.1. Body Parameters, Body Mass and Tissue Mass
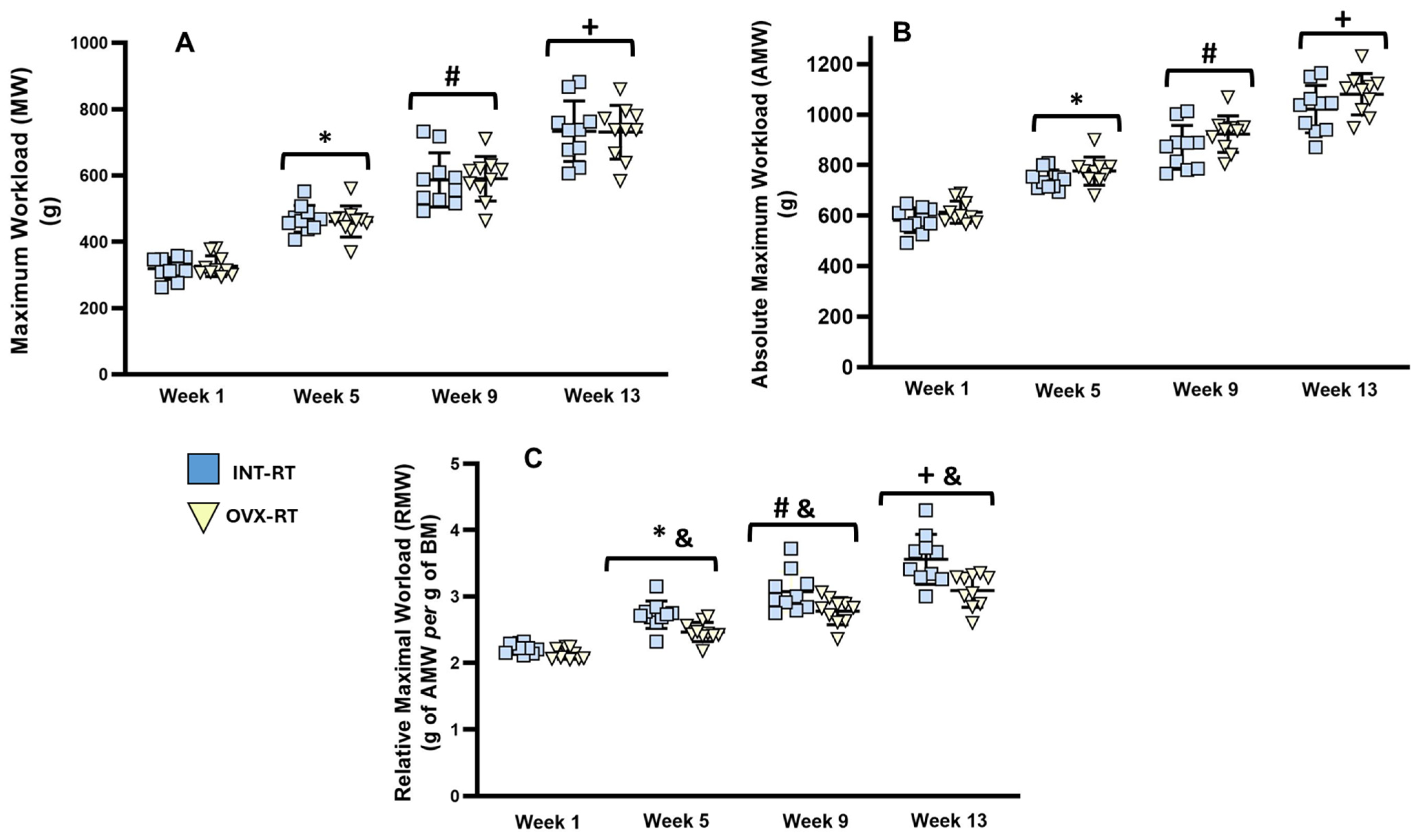
2.2. Maximal Workload
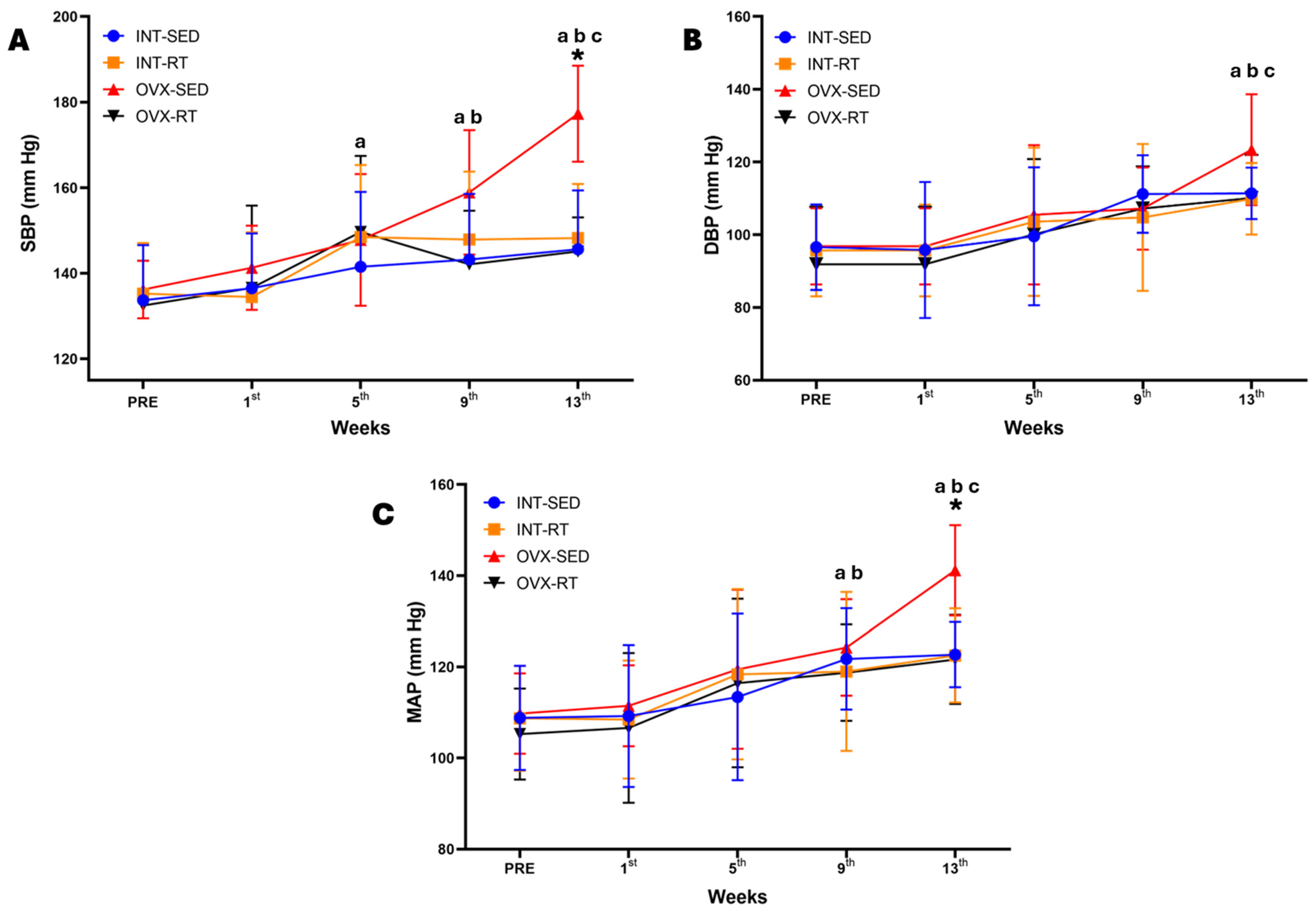
2.3. Hemodynamic Parameters: Systolic Blood Pressure, Diastolic Blood Pressure, Mean Arterial Pressure, Heart Rate, and Double Product
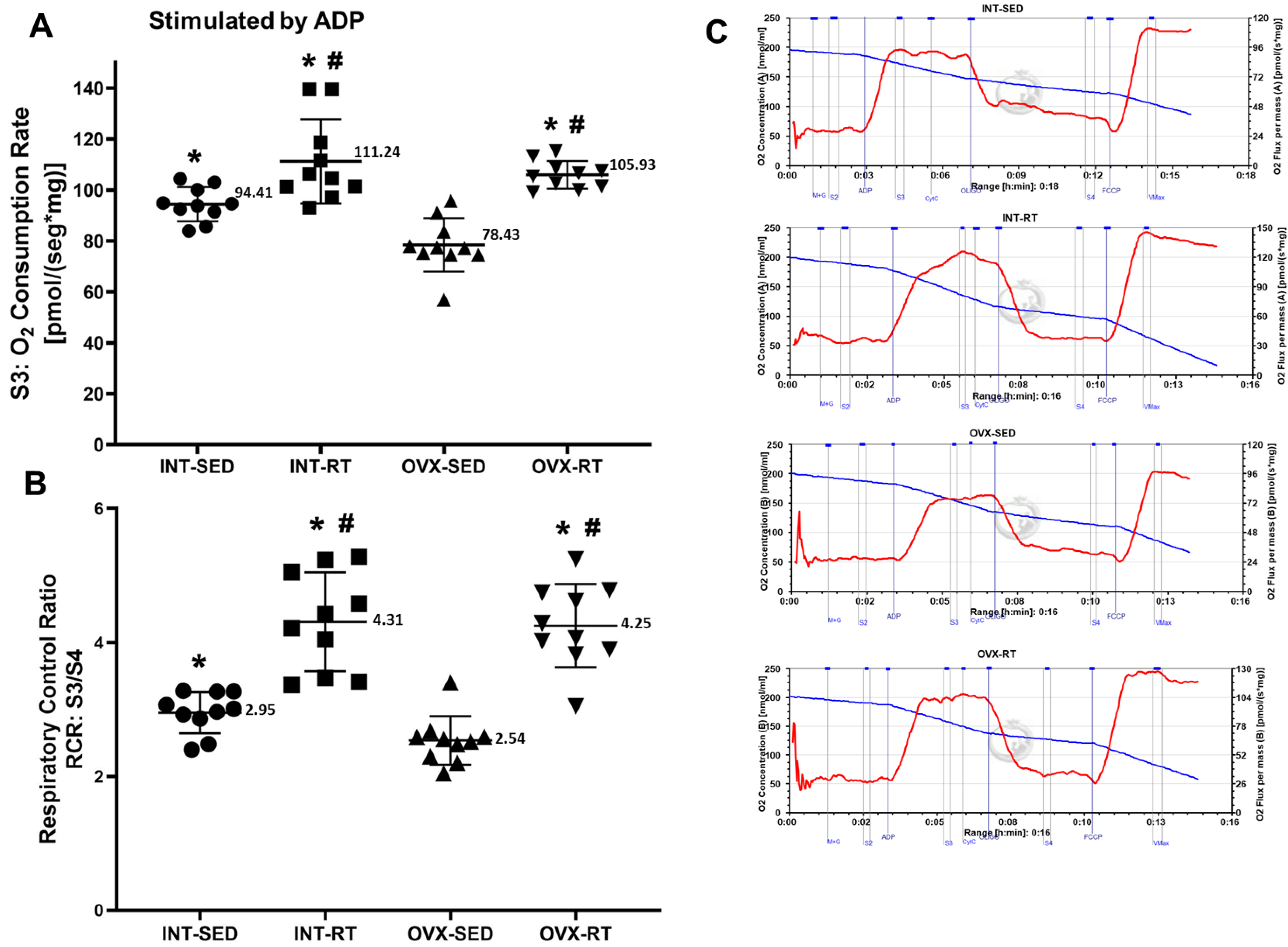
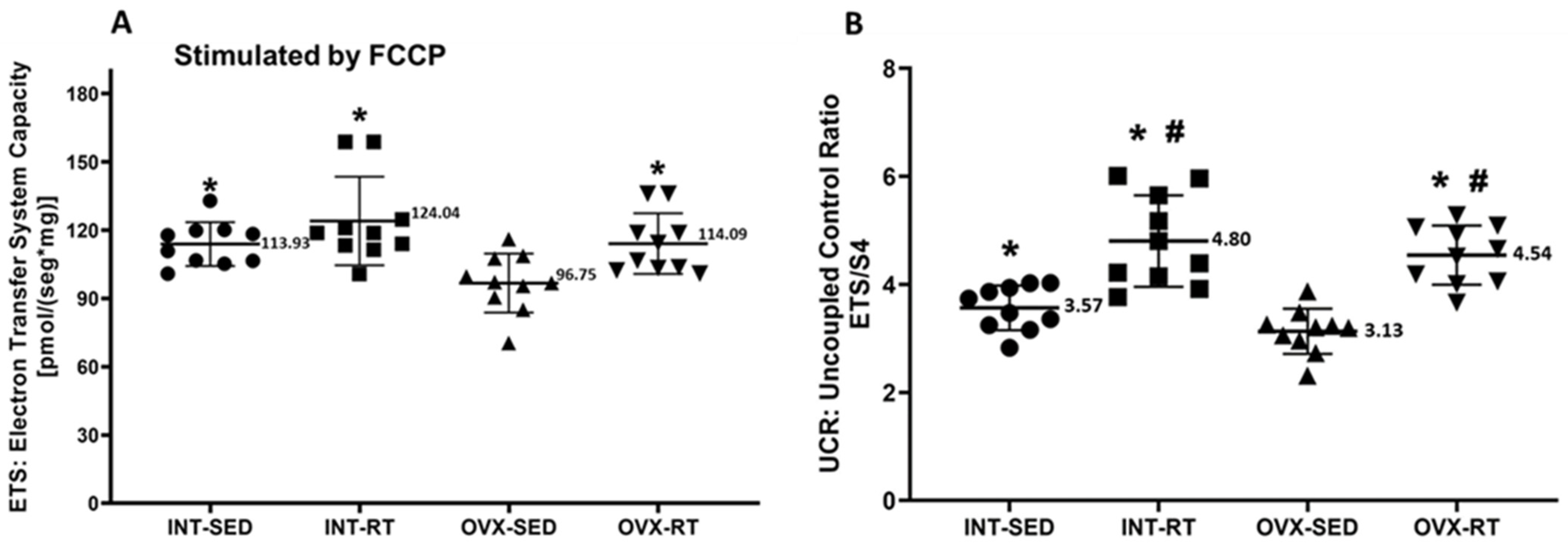
2.4. Mitochondrial Respiratory Function
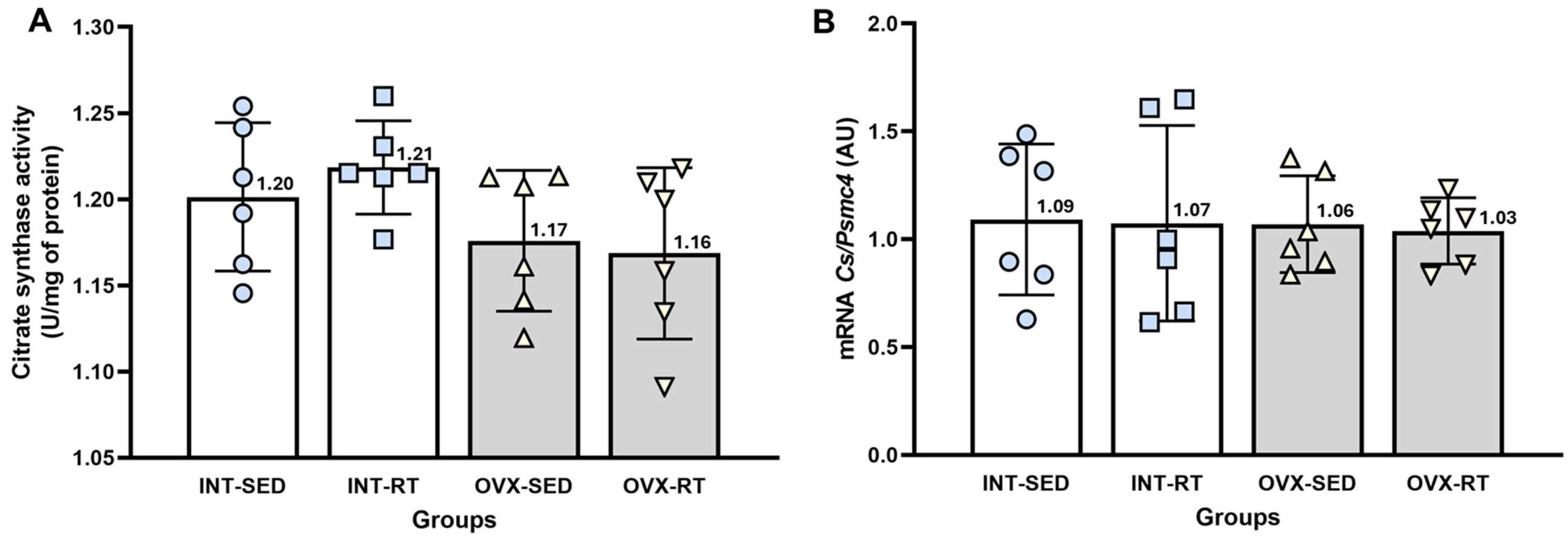
2.5. Citrate Synthase Activity and mRNA Gene Expression of Citrate Synthase
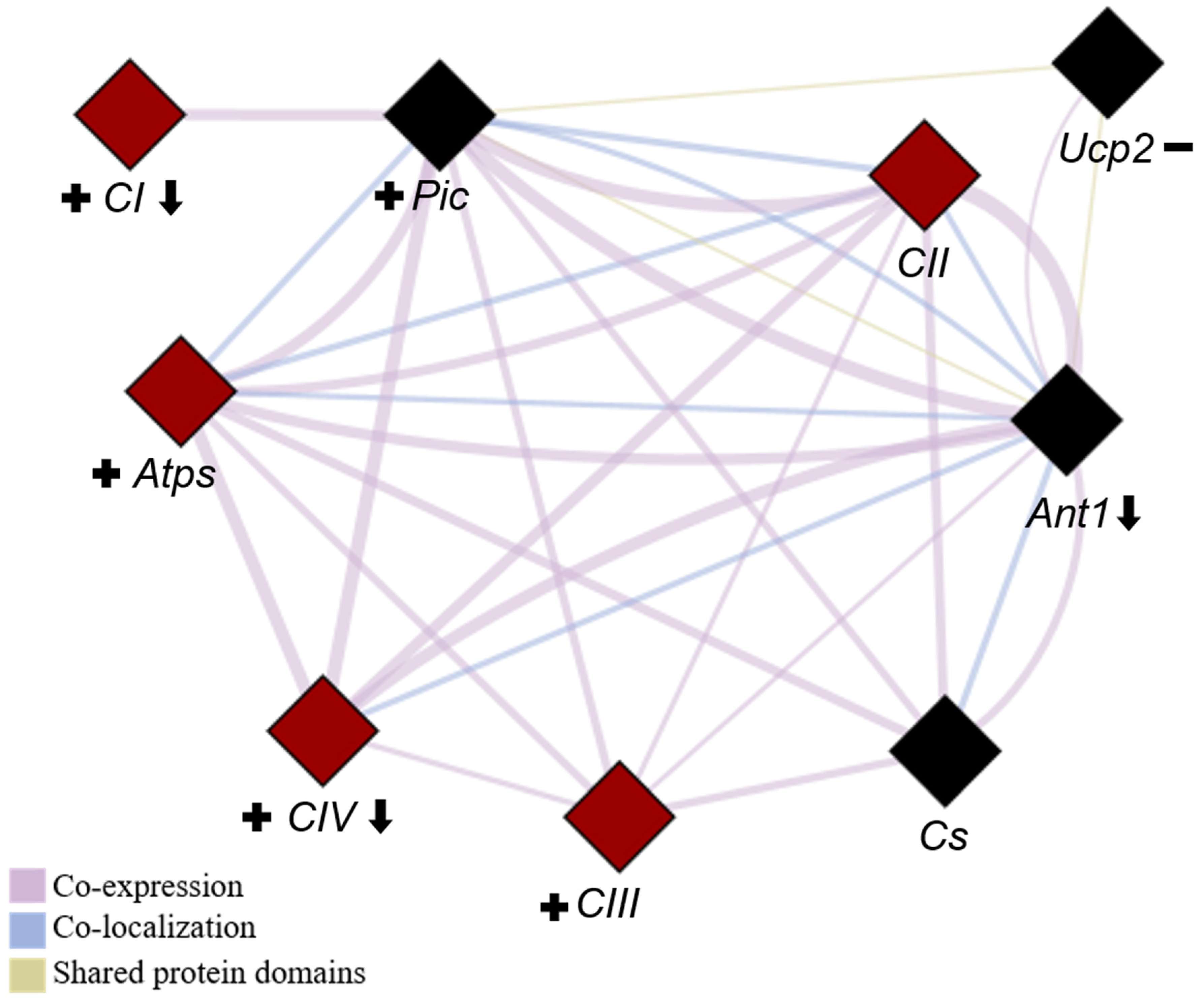
2.6. Quantification of mRNA Gene Expression Related to Mitochondrial ETC and OXPHOS
2.6.1. Gene Expression of ETC Protein Complexes (I to IV)
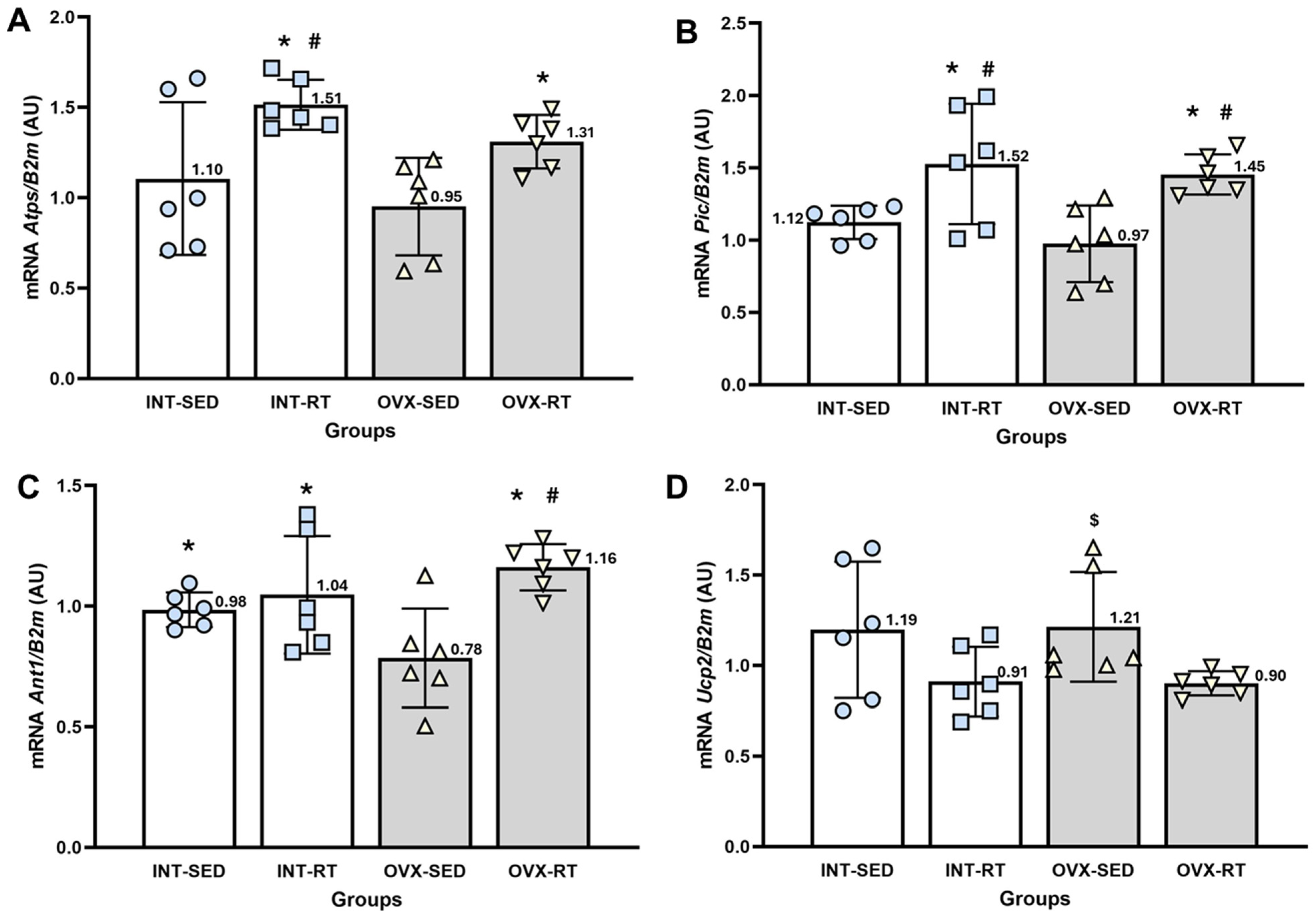
2.6.2. Gene Expression of the Phosphorylation System (Atps, Pic, and Ant1) and Ucp2
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Animals
4.3. Experimental Groups
4.4. Ovariectomy (OVX)
4.5. Resistance Training (RT) Protocol
4.6. Blood Pressure and Heart Rate Measurements
4.7. Euthanasia and Tissue Dissection
4.8. Analysis of Mitochondrial Respiratory Function (High-Resolution Respirometry)
4.8.1. Tissue Preparation, Mechanical and Chemical Permeabilization
4.8.2. Measurement of Oxygen Consumption
4.9. Citrate Synthase Activity
4.10. Gene Expression
4.10.1. Extraction, Purification, Quantification, and Integrity of Total RNA
4.10.2. Reverse Transcription to Complementary DNA (cDNA)
4.10.3. Real-Time Polymerase Chain Reaction (qPCR)
4.10.4. Quality Control, Primer Concentration, and Efficiency
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, H.D. Menopause. Lancet 2008, 371, 760–770. [Google Scholar] [CrossRef]
- Vellanki, K.; Hou, S. Menopause in CKD. Am. J. Kidney Dis. 2018, 71, 710–719. [Google Scholar] [CrossRef]
- Suzuki, H.; Kondo, K. Chronic kidney disease in postmenopausal women. Hypertens. Res. 2012, 35, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Mosca, L.; Appel, L.J.; Benjamin, E.J.; Berra, K.; Chandra-Strobos, N.; Fabunmi, R.P.; Grady, D.; Haan, C.K.; Hayes, S.N.; Judelson, D.R.; et al. Evidence based guidelines for cardiovascular disease prevention in women. Arter. Thromb. Vasc. Biol. 2004, 24, 29–50. [Google Scholar] [CrossRef]
- Rosano, G.M.; Vitale, C.; Marazzi, G.; Volterrani, M. Menopause and cardiovascular disease: The evidence. Climatereic 2007, 10, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.C. Effect of early menopause on bone mineral density and fractures. Menopause 2007, 14, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Honigberg, M.C.; Zekavat, S.M.; Aragam, K.; Finneran, P.; Klarin, D.; Bhatt, D.L.; Januzzi, J.L., Jr.; Scott, N.S.; Natarajan, P. Association of premature natural and surgical menopause with incident cardiovascular disease. JAMA 2019, 322, 2411–2421. [Google Scholar] [CrossRef]
- Anagnostis, P.; Christou, K.; Artzouchaltzi, A.M.; Gkekas, N.K.; Kosmidou, N.; Siolos, P.; Paschou, S.A.; Potoupnis, M.; Kenanidis, E.; Tsiridis, E.; et al. Early menopause and premature ovarian insufficiency are associated with increased risk of type 2 diabetes: A systematic review and meta-analysis. Eur. J. Endocrinol. 2019, 180, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.B.; Ramesh, S. Sex hormones in women with kidney disease. Nephrol. Dial. Transplant. 2016, 31, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Qian, D.; Wang, Z.; Cheng, Y.; Luo, R.; Ge, S.W.; Xu, G. Early Menopause May Associate with a Higher Risk of CKD and All-Cause Mortality in Postmenopausal Women: An Analysis of NHANES, 1999–2014. Front. Med. 2022, 9, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D.R. Global Prevalence of Chronic Kidney Disease—A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [PubMed]
- van der Velde, M.; Matsushita, K.; Coresh, J.; Astor, B.C.; Woodward, M.; Levey, A.S.; Jong, P.E.; Gansevoort, R.T. Lower estimated glomerular filtration rate and higher albuminuria are associated with all-cause and cardiovascular mortality. A collaborative meta-analysis of high risk population cohorts. Kidney Int. 2011, 79, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Yerram, P.; Karuparthi, P.R.; Hesemann, L.; Horst, J.; Whaley-Connell, A. Chronic kidney disease and cardiovascular risk. J. Am. Soc. Hypertens. 2007, 1, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Tedla, F.M.; Brar, A.; Browne, R.; Brown, C. Hypertension in Chronic KidneyDisease: Navigating the Evidence. Int. J. Hypertens. 2011, 2011, 132405. [Google Scholar] [CrossRef] [PubMed]
- Burnier, M.; Damianaki, A. Hypertension as Cardiovascular Risk Factor in Chronic Kidney Disease. Circ. Res. 2023, 132, 1050–1106. [Google Scholar] [CrossRef]
- Ritz, E. Hypertension and kidney disease. Clin. Nephrol. 2010, 74 (Suppl. S1), S39–S43. [Google Scholar] [CrossRef]
- Mahmoodi, B.K.; Matsushita, K.; Woodward, M.; Blankestijn, P.J.; Cirillo, M.; Ohkubo, T.; Rossing, P.; Sarnak, M.J.; Stengel, B.; Yamagishiet, K.; et al. Associations of kidney disease measures with mortality and end-stage renal disease in individuals with and without hypertension: A meta-analysis. Lancet 2012, 380, 1649–1661. [Google Scholar] [CrossRef]
- Koebele, S.V.; Bimonte-Nelson, H.A. Modeling menopause: The utility of rodents in translational behavioral endocrinology research. Maturitas 2016, 87, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Medina-Contreras, J.M.L.; Villalobos-Molina, R.; Zarain-Herzberg, A.; Balderas-Villalobos, J. Ovariectomized rodents as a menopausal metabolic syndrome model. Mol. Cell. Biochem. 2020, 475, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, M.; Fang, G.; Tang, Y.; Chen, Z.; Liu, X. Hypocholesterolemic effect of capsaicinoids by increased bile acids excretion in ovariectomized rats. Mol. Nutr. Food Res. 2013, 57, 1080–1088. [Google Scholar] [CrossRef]
- Sivasinprasasn, S.; Sa-Nguanmoo, P.; Pratchayasakul, W.; Kumfu, S.; Chattipakorn, S.C.; Chattipakorn, N. Obese-insulin resistance accelerates and aggravates cardiometabolic disorders and cardiac mitochondrial dysfunction in estrogen-deprived female rats. Age 2015, 37, 28. [Google Scholar] [CrossRef] [PubMed]
- Lino, A.D.S.; Vianna, D.; Oishi, J.C.; Souza, M.V.C.; Ruffoni, L.D.; Marin, C.T.; Avó, L.R.S.; Perez, S.E.A.; Rodrigues, G.J.; Tirapegui, J.; et al. Resistance training and caloric restriction prevent systolic blood pressure rise by improving the nitric oxide effect on smooth muscle and morphological changes in the aorta of ovariectomized rats. PLoS ONE 2018, 13, e0201843. [Google Scholar] [CrossRef]
- Hassan, H.A.; Abdel-Wahhab, M.A. Effect of soybean oil on atherogenic metabolic risks associated with estrogen deficiency in ovariectomized rats: Dietary soybean oil modulates atherogenic risks in ovariectomized rats. J. Physiol. Biochem. 2012, 68, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Shiguemoto, G.E.; Prestes, J.; Leite, R.D.; Pereira, G.B.; Pontes, C.L.S.; D’Ávila, F.V.; Botero, J.P.; Baldissera, V.; Nonaka, K.O.; Selistre-de-Araújo, H.S.; et al. Effects of resistance training on matrix metalloproteinase-2 activity and biomechanical and physical properties of bone in ovariectomized and intact rats. Scand. J. Med. Sci. Sports 2012, 22, 607–617. [Google Scholar] [CrossRef]
- Yousefzadeh, N.; Jeddi, S.; Zarkesh, M.; Norouzirad, R.; Kashfi, K.; Ghasemi, A. Protective effects of long-term nitrate administration against ovariectomy-induced kidney dysfunction in rats. Pharmacol. Rep. 2023, 75, 979–994. [Google Scholar] [CrossRef]
- Ikeda, M.; Swide, T.; Vayl, A.; Lahm, T.; Anderson, S.; Hutchens, M.P. Estrogen administered after cardiac arrest and cardiopulmonary resuscitation ameliorates acute kidney injury in a sex-and age-specific manner. Crit. Care 2015, 19, 332. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.S.; Sheikh-Hamad, D. Mitochondrial dysfunction in acute kidney injury and sex-specific implications. Med. Res. Arch. 2019, 7, 1–15. [Google Scholar] [CrossRef]
- Patil, N.K.; Parajuli, N.; MacMillan-Crow, L.A.; Mayeux, P.R. Inactivation of renal mitochondrial respiratory complexes and manganese superoxide dismutase during sepsis: Mitochondria-targeted antioxidant mitigates injury. Am. J. Physiol. Ren. Physiol. 2014, 306, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Galvan, D.L.; Green, N.H.; Danesh, F.R. The hallmarks of mitochondrial dysfunction in chronic kidney disease. Kidney Int. 2017, 92, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Granata, S.; Zaza, G.; Simone, S.; Villani, G.; Latorre, D.; Pontrelli, P.; Carella, M.; Schena, F.P.; Grandaliano, G.; Pertosa, G. Mitochondrial dysregulation and oxidative stress in patients with chronic kidney disease. BMC Genom. 2009, 10, 388. [Google Scholar] [CrossRef] [PubMed]
- Che, R.; Yuan, Y.; Huang, S.; Zhang, A. Mitochondrial dysfunction in the pathophysiology of renal diseases. Am. J. Physiol. Ren. Physiol. 2014, 306, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Dhoopun, A.R.; Yuan, Y.; Huang, S.; Zhu, C.; Ding, G.; Liu, B.; Yang, T.; Zhang, A. Mitochondrial dysfunction is an early event in aldosterone-induced podocyte injury. Am. J. Physiol. Ren. Physiol. 2013, 305, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Ventura-Clapier, R.; Piquereau, J.; Veksler, V.; Garnier, A. Estrogens, Estrogen Receptors Effects on Cardiac and Skeletal Muscle Mitochondria. Front. Endocrinol. 2019, 10, 557. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Luo, M.; Wang, R.; Ye, J.; Wang, X. Mitochondria in sex-hormone induced disorder of energy metabolism in males and females. Front. Endocrinol. 2021, 12, 749451. [Google Scholar] [CrossRef]
- Lin, Y.; Lee, S. Cardiovascular Benefits of Exercise Training in Postmenopausal Hypertension. Int. J. Mol. Sci. 2018, 19, 2523. [Google Scholar] [CrossRef] [PubMed]
- Yuk, J.S. Relationship between menopausal hormone therapy and breast cancer: A nationwide population-based cohort study. Int. J. Gynecol. Obstet. 2024, 166, 735–744. [Google Scholar] [CrossRef]
- Manson, J.; Chlebowski, R.; Stefanick, M.; Aragaki, A.; Rossouw, J.; Prentice, R.; Anderson, G.L.; Howard, B.V.; Thomson, C.A.; Lacroix, A.Z.; et al. Menopausal hormone therapy and health outcomes during the intervention and extended post stopping phases of the Women’s Health Initiative randomized trials. JAMA 2013, 310, 1353–1368. [Google Scholar] [CrossRef]
- Manson, J.; Aragaki, K.; Rossouw, J.; Anderson, G.; Prentice, R.; LaCroix, A.; Chlebowski, R.T.; Howard, B.V.; Thomson, C.A.; Margolis, K.L.; et al. Menopausal hormone therapy and long term all-cause and cause-specific mortality: The Women’s Health Initiative randomized trials. JAMA 2017, 318, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Stojanovska, L.; Apostolopoulos, V.; Polman, R.; Borkoles, E. To exercise, or, not to exercise during menopause and beyond. Maturitas 2014, 77, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Grindler, N.M.; Santoro, N.F. Menopause and exercise. Menopause J. N. Am. Menopause Soc. 2015, 22, 1351–1358. [Google Scholar] [CrossRef]
- Figueroa, A.; Park, S.Y.; Seo, D.Y.; Sanchez-Gonzalez, M.A.; Baek, Y.H. Combined resistance and endurance exercise training improves arterial stiffness, blood pressure, and muscle strength in postmenopausal women. Menopause 2011, 18, 980–984. [Google Scholar] [CrossRef]
- Marin, C.T.; Lino, A.D.S.; Avelar, I.D.S.; Barbosa, M.R.; Scarlato, G.C.G.; Cavalini, D.F.; Tamanini, F.; Alexandrino, A.V.; Vercesi, A.E.; Shiguemoto, G.E. Resistance training prevents dynamics and mitochondrial respiratory dysfunction in vastus lateralis muscle of ovariectomized rats. Exp. Gerontol. 2023, 173, 112081. [Google Scholar] [CrossRef]
- Moinuddin, I.; Leehey, D.J. A Comparison of Aerobic Exercise and Resistance Training in Patients with and without Chronic Kidney Disease. Adv. Chronic Kidney Dis. 2008, 15, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Malheiro, L.F.L.; Fernandes, M.M.; Oliveira, C.A.; Barcelos, I.S.; Fernandes, A.J.V.; Silva, B.S.; Ávila, J.S.; Soares, T.J.; Amaral, L.S.B. Renoprotective mechanisms of exercise training against acute and chronic renal diseases—A perspective based on experimental studies. Life Sci. 2024, 346, 122628. [Google Scholar] [CrossRef] [PubMed]
- Leite, A.B.; Lima, H.N.; Flores, C.O.; Oliveira, C.A.; Cunha, L.E.C.; Neves, J.L.; Correia, T.M.L.; de Melo, F.F.; Oliveira, M.V.; de Magalhães, A.C.M.; et al. High-intensity interval training is more effective than continuous training to reduce inflammation markers in female rats with cisplatin nephrotoxicity. Life Sci. 2021, 266, 118880. [Google Scholar] [CrossRef]
- Amaral, L.S.B.; Silva, F.A.; Correia, V.B.; Andrade, C.E.; Dutra, B.A.; Oliveira, M.V.; de Magalhães, A.C.; Volpini, R.A.; Seguro, A.C.; Coimbra, T.M.; et al. Beneficial effects of previous exercise training on renal changes in streptozotocin-induced diabetic female rats. Exp. Biol. Med. 2017, 241, 437–445. [Google Scholar] [CrossRef]
- Zeynali, F.; Nematbakhsh, M.; Mojtahedi, H.; Poorshahnazari, A.; Talebi, A.; Pezeshki, Z.; Mazaheri, S.; Moslemi, F. Protective role of aerobic exercise against cisplatin-induced nephrotoxicity in rats. Asian. J. Sports Med. 2015, 6, e24901. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Bergamaschi, C.T.; Araújo, R.C.; Mouro, M.G.; Rosa, T.S.; Higa, E.M.S. Effects of training and nitric oxide on diabetic nephropathy progression in type I diabetic rats. Exp. Biol. Med. 2011, 236, 1180–1187. [Google Scholar] [CrossRef]
- Yamakoshi, S.; Nakamura, T.; Mori, N.; Suda, C.; Kohzuki, M.; Ito, O. Effects of exercise training on renal interstitial fibrosis and renin-angiotensin system in rats with chronic renal failure. J. Hypertens. 2021, 39, 143–152. [Google Scholar] [CrossRef]
- Almeida, A.A.; Correia, T.M.L.; Pires, R.A.; Silva, D.A.; Coqueiro, R.S.; Machado, M.; de Magalhães, A.; Queiroz, R.; Soares, T.; Pereira, R. Nephroprotective effect of exercise training in cisplatin-induced renal damage in mice: Influence of training protocol. Braz. J. Med. Biol. Res. 2022, 55, e12116. [Google Scholar] [CrossRef]
- Oliveira, C.; Mercês, É.; Portela, F.; Benedictis, J.; Benedictis, L.; Silva, A.; Campanati, J.d.A.G.; Freire de Melo, F.; Oliveira, M.V.; Mendes de Magalhães, A.C.; et al. Benefits of high-intensity interval training compared to continuous training to reduce apoptotic markers in female rats with cisplatin nephrotoxicity—Possible modulatory role of IL-11. Apoptosis 2023, 28, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Bayod, S.; Del Valle, J.; Lalanza, J.; Sanchez-Roige, S.; de Luxan-Delgado, B.; Coto-Montes, A.; Canudas, A.; Camins, A.; Escorihuela, R.; Pallàs, M. Long-term physical exercise induces changes in sirtuin 1 pathway and oxidative parameters in adult rat tissues. Exp. Gerontol. 2012, 47, 925–935. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wang, Y.; Zhang, M.Z.; You, L.; Davis, L.S.; Fan, H.; Yang, H.-C.; Fogo, A.B.; Zent, R.; Harris, R.C.; et al. Sirt1 activation protects the mouse renal medulla from oxidative injury. J. Clin. Investig. 2010, 120, 1056–1068. [Google Scholar] [CrossRef]
- Dong, Y.J.; Liu, N.; Xiao, Z.; Sun, T.; Wu, S.H.; Sun, W.X.; Xu, Z.-G.; Yuan, H. Renal protective effect of sirtuin 1. J. Diabetes Res. 2014, 2014, 843786. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. Estrogenic control of mitochondrial function. Redox Biol. 2020, 31, 101435. [Google Scholar] [CrossRef] [PubMed]
- Anagnostis, P.; Theocharis, P.; Lallas, K.; Konstantis, G.; Mastrogiannis, K.; Bosdou, J.K.; Lambrinoudaki, I.; Stevenson, J.C.; Goulis, D. Early menopause is associated with increased risk of arterial hypertension: A systematic review and meta-analysis. Maturitas 2020, 135, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef]
- Park, S.Y.; Gifford, J.R.; Andtabacha, R.H.I.; Trinity, J.D.; Hyngstrom, J.R.; Garten, R.S.; Diakos, N.A.; Ives, S.J.; Dela, F.; Larsen, S.; et al. Cardiac, skeletal, and smooth muscle mitochondrial respiration: Are all mitochondria created equal? Am. J. Physiol. Heart Circ. Physiol. 2014, 307, 346–352. [Google Scholar] [CrossRef]
- Gnaiger, E. Mitochondrial Pathways and Respiratory Control. An Introduction to OXPHOS Analysis, 4th ed., Mitochondr Physiol Network 17.18. OROBOROS MiPNet Publications, Innsbruck. 2014. Available online: https://wiki.oroboros.at/images/f/fc/Gnaiger_2014_Mitochondr_Physiol_Network_MitoPathways.pdf (accessed on 22 December 2024).
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle: New perspectives of mitochondrial physiology. Int. J. Biochem. Cell Biol. 2009, 41, 1837–1845. [Google Scholar] [CrossRef]
- Ruas, S.R.; Siqueira-Campos, E.S.; Rodrigues-Silva, E.; Castilho, R.F. High glycolytic activity of tumor cells leads to underestimation of electron transport system capacity when mitochondrial ATP synthase is inhibited. Sci. Rep. 2018, 8, 17383. [Google Scholar] [CrossRef]
- Jedlička, J.; Grundmanová, M.; Švíglerová, J.; Tůma, Z.; Nalos, L.; Rajdl, D.; Štengl, M.; Kuncová, J. Mitochondrial Dysfunction in Kidney Cortex and Medulla of Subtotally Nephrectomized Rats. Physiol. Res. 2022, 71, 219–226. [Google Scholar] [CrossRef]
- Porter, C.; Reidy, P.T.; Bhattarai, N.; Sidossis, L.S.; Rasmussen, B.B. Resistance exercise training alters mitochondrial function in human skeletal muscle. Med. Sci. Sports Exerc. 2015, 47, 1922–1931. [Google Scholar] [CrossRef]
- Jacobs, R.A.; Lundby, C. Mitochondria express enhanced quality as well as quantity in association with aerobic fitness across recreationally active individuals up to elite athletes. J. Appl. Physiol. 2013, 114, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Hütter, E.; Unterluggauer, H.; Garedew, A.; Jansen-Dürr, P.; Gnaiger, E. High-resolution respirometry–a modern tool in aging research. Exp. Gerontol. 2006, 41, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Divakaruni, A.S.; Brand, M.D. The regulation and physiology of mitochondrial proton leak. Physiology 2011, 26, 192–205. [Google Scholar] [CrossRef]
- Griffiths, K.K.; Wang, A.; Wang, L.; Tracey, M.; Kleiner, G.; Quinzii, C.M.; Sun, L.; Yang, G.; Perez-Zoghbi, J.F.; Licznerski, P.; et al. Inefficient Thermogenic Mitochondrial Respiration Due to Futile Proton Leak in a Mouse Model of Fragile X Syndrome. FASEB J. 2020, 34, 7404–7426. [Google Scholar] [CrossRef]
- Sreedhar, A.; Zhao, Y. Uncoupling Protein 2 and Metabolic Diseases. Mitochondrion 2017, 34, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Friederich, M.; Fasching, A.; Hansell, P.; Nordquist, L.; Palm, F. Diabetes-induced up-regulation of uncoupling protein-2 results in increased mitochondrial uncoupling in kidney proximal tubular cells. Biochim. Biophys. Acta. 2008, 1777, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 2017, 13, 620–646. [Google Scholar] [CrossRef]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Junyi, Z.; Britta, S.; Wiebke, L.; Heymsfield, S.B.; Manfred, M. Specific metabolic rates of major organs and tissues across adulthood: Evaluation by mechanistic model of resting energy expenditure. Am. J. Clin. Nutr. 2010, 92, 1369–1377. [Google Scholar] [CrossRef]
- Sharma, K. Mitochondrial dysfunction in the diabetic kidney. Adv. Exp. Med. Biol. 2017, 982, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. Mitochondrial hormesis and diabetic complications. Diabetes 2015, 64, 663–672. [Google Scholar] [CrossRef]
- Szeto, H.H. Pharmacologic approaches to improve mitochondrial function in AKI and CKD. J. Am. Soc. Nephrol. 2017, 28, 2856–2865. [Google Scholar] [CrossRef]
- Eirin, A.; Lerman, A.; Lerman, L.O. Enhancing mitochondrial health to treat hypertension. Curr. Hypertens. Rep. 2018, 20, 89. [Google Scholar] [CrossRef]
- McCrimmon, A.; Domondon, M.; Sultanova, R.F.; Ilatovskaya, D.V.; Stadler, K. Comprehensive assessment of mitochondrial respiratory function in freshly isolated nephron segments. Am. J. Physiol. Ren. Physiol. 2020, 318, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, J.; Zhang, Y.; Zhou, J. Focus on Mitochondrial Respiratory Chain: Potential Therapeutic Target for Chronic Renal Failure. Int. J. Mol. Sci. 2024, 25, 949. [Google Scholar] [CrossRef]
- Singh, P. Reprogramming of Energy Metabolism in Kidney Disease. Nephron 2023, 147, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Hu, J.; Chen, Z.; Feng, J.; Yang, X.; Liang, W.; Ding, G. Transition of acute kidney injury to chronic kidney disease: Role of metabolic reprogramming. Metab. Clin. Exp. 2022, 131, 155–194. [Google Scholar] [CrossRef]
- Barrientos, A. In vivo and in organello assessment of OXPHOS activities. Methods 2002, 26, 307–316. [Google Scholar] [CrossRef]
- Yubero, D.; Adin, A.; Montero, R.; Jou, C.; Jiménez-Mallebrera, C.; García-Cazorla, A.; Nascimento, A.; O’Callaghan, M.M.; Montoya, J.; Gortet, L.; et al. A statistical algorithm showing coenzyme Q10 and citrate synthase as biomarkers for mitochondrial respiratory chain enzyme activities. Sci. Rep. 2016, 6, 15. [Google Scholar] [CrossRef]
- Frankish, B.P.; Najdovska, P.; Xu, H.; Wette, S.G.; Murphy, R.M. Effects of voluntary wheel running on mitochondrial content and dynamics in rat skeletal muscle. J. Muscle Res. Cell Motil. 2021, 42, 67–76. [Google Scholar] [CrossRef]
- Ullian, M.E.; Gantt, B.J.; Ford, A.K.; Tholanikunnel, B.G.; Spicer, E.K.; Fitzgibbon, W.R. Potential importance of glomerular citrate synthase activity in remnant nephropathy. Kidney Int. 2003, 63, 156–164. [Google Scholar] [CrossRef]
- Mapuskar, K.A.; Wen, H.; Holanda, D.G.; Rastogi, P.; Steinbach, E.; Han, R.; Coleman, M.C.; Attanasio, M.; Riley, D.; Spitz, D.R.; et al. Persistent increase in mitochondrial superoxide mediates cisplatin-induced chronic kidney disease. Redox Biol. 2019, 20, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Dines, V.A.; Garovic, V.D. Menopause and chronic kidney disease. Nat. Rev. Nephrol. 2023, 20, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; Ballew, S.H.; Wang, A.Y.; Kalyesubula, R.; Schaeffner, E.; Agarwal, R. Epidemiology and risk of cardiovascular disease in populations with chronic kidney disease. Nat. Rev. Nephrol. 2022, 18, 696–707. [Google Scholar] [CrossRef]
- Rashid, A.; Jamil, A.; Khan, Z.; Shakoor, M.; Kamal, U.; Khan, I.; Akram, A.; Shahabi, M.; Yamani, N.; Ali, S.; et al. Trends in mortality related to kidney failure and diabetes mellitus in the United States: A 1999–2020 analysis. J. Nephrol. 2024, 37, 1833–1843. [Google Scholar] [CrossRef]
- Damianaki, A.; Polychronopoulou, E.; Wuerzner, G.; Burnier, M. New Aspects in the Management of Hypertension in Patients with Chronic Kidney Disease not on Renal Replacement Therapy. High Blood Press. Cardiovasc. Prev. 2022, 29, 125–135. [Google Scholar] [CrossRef]
- Dutta, S.R.; Singh, P.; Malik, K.U. Ovariectomy Via 12/15-lipoxygenase Augments Angiotensin II-Induced Hypertension and Its Pathogenesis in Female Mice. Hypertension 2023, 80, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Hong, Y.; Zhou, M.C.; Huang, H.L.; Shyu, W.C.; Chen, J.S.; Ting, H.; Cheng, Y.J.; Yang, A.L.; Lee, S.D. Exercise training attenuates cardiac inflammation and fibrosis in hypertensive ovariectomized rats. J. Appl. Physiol. 2020, 128, 1033–1104. [Google Scholar] [CrossRef]
- Shimojo, G.L.; Palma, R.K.; Brito, J.O.; Sanches, I.C.; Irigoyen, M.C.; De Angelis, K. Dynamic resistance training decreases sympathetic tone in hypertensive ovariectomized rats. Braz. J. Med. Biol. Res. 2015, 48, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Da Palma, R.K.; Moraes-Silva, I.C.; Dias, S.D.; Shimojo, G.L.; Conti, F.F.; Bernardes, N.; Barboza, C.A.; Sanches, I.C.; Araujo, A.S.; Irigoyen, M.C.; et al. Resistance or aerobic training decreases blood pressure and improves cardiovascular autonomic control and oxidative stress in hypertensive menopausal rats. J. Appl. Physiol. 2016, 121, 1032–1038. [Google Scholar] [CrossRef]
- Son, W.M.; Sung, K.D.; Cho, J.M.; Park, S.Y. Combined exercise reduces arterial stiffness, blood pressure, and blood markers for cardiovascular risk in postmenopausal women with hypertension. Menopause 2017, 24, 262–268. [Google Scholar] [CrossRef]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Himmelfarb, C.D.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2018, 71, 1269–1324. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Silva, M.P.S.F.; Dias, D.S.; Bernardes, N.; Irigoyen, M.C.; De Angelis, K. Concurrent exercise training induces additional benefits to hydrochlorothiazide: Evidence for an improvement of autonomic control and oxidative stress in a model of hypertension and postmenopause. PLoS ONE 2023, 18, e0289715. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Thomas, R.L.; Campbell, M.D.; Prior, S.L.; Bracken, R.M.; Churm, R. Effects of exercise training on metabolic syndrome risk factors in post-menopausal women: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2023, 42, 337–351. [Google Scholar] [CrossRef]
- Luan, X.; Tian, X.; Zhang, H.; Huang, R.; Li, N.; Chen, P.; Wang, R. Exercise as a prescription for patients with various diseases. J. Sport Health Sci. 2019, 8, 422–441. [Google Scholar] [CrossRef] [PubMed]
- Mallamaci, F.; Pisano, A.; Triepi, G. Physical activity in chronic kidney disease and the EXerCise Introduction to Enhance trial. Nephrol. Dial. Transplant. 2020, 35, 18–22. [Google Scholar] [CrossRef]
- Wilkinson, T.J.; McAdams-DeMarcob, M.; Bennett, P.N.; Wilund, K. Advances in exercise therapy in predialysis chronic kidney disease, hemodialysis, peritoneal dialysis, and kidney transplantation. Curr. Opin. Nephrol. Hypertens. 2020, 29, 471–479. [Google Scholar] [CrossRef]
- Villanego, F.; Naranjo, J.; Vigara, L.A.; Cazorla, J.M.; Montero, M.E.; García, T.; Torrado, J.; Mazuecos, A. Impacto del ejercicio físico en pacientes con enfermedad renal crónica: Revisión sistemática y metaanálisis. Nefrologia 2020, 40, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Sprick, J.D.; DaCosta, D.R.; Mammino, K.; Nocera, J.R.; Park, J. Exercise modulates sympathetic and vascular function in chronic kidney disease. Clin. Med. 2023, 8, 164–221. [Google Scholar] [CrossRef]
- Corrêa, H.L.; Neves, R.V.P.; Deus, L.A.; Maia, B.C.H.; Maya, A.T.; Tzanno-Martins, C.; Souza, M.K.; Silva, J.A.B.; Haro, A.S.; Costa, F.; et al. Low-load resistance training with blood flow restriction prevent renal function decline: The role of the redox balance, angiotensin 1–7 and vasopressin. Physiol. Behav. 2021, 230, 113–295. [Google Scholar] [CrossRef]
- Saud, A.; Luiz, R.S.; Leite, A.P.O.; Muller, C.R.; Visona, I.; Reinecke, N.; Silva, W.H.; Gloria, M.A.; Razvickas, C.V.; Casarini, D.E.; et al. Resistance exercise training ameliorates chronic kidney disease outcomes in a 5/6 nephrectomy model. Life Sci. 2021, 275, 119–362. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Grundy, D. Principles and standards for reporting animal experiments in The Journal of Physiology and Experimental Physiology. J. Physiol. 2015, 593, 2547–2549. [Google Scholar] [CrossRef] [PubMed]
- Horcajada, M.N.; Habauzit, V.; Trzeciakiewicz, A.; Morand, C.; Gil-Izquierdo, A.; Mardon, J.; Lebecque, P.; Davicco, M.J.; Chee, W.S.S.; Coxam, V.; et al. Hesperidin inhibits ovariectomized-induced osteopenia and shows differential effects on bone mass and strength in young and adult intact rats. J. Appl. Physiol. 2008, 104, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Souza, S.B.C.; Flues, K.; Paulini, J. Role of exercise training in cardiovascular autonomic dysfunction and mortality in diabetic ovariectomized rats. Hypertension 2007, 50, 786–791. [Google Scholar] [CrossRef]
- Brinton, R.D. Minireview: Translational animal models of human menopause: Challenges and emerging opportunities. Endocrinology 2012, 153, 3571–3578. [Google Scholar] [CrossRef] [PubMed]
- Hornberger, T.A., Jr.; Farrar, R.P. Physiological hypertrophy of the FHL muscle following 8 weeks of progressive resistance exercise in the rat. Can. J. Appl. Physiol. 2004, 29, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Whitesall, S.; Zhang, Y.; Beibel, M.; D’ Alecy, L.; DiPetrillo, K. Validation of Volume—Pressure Recording Tail-Cuff Blood Pressure Measurements. Am. J. Hypertens. 2008, 21, 1288–1291. [Google Scholar] [CrossRef]
- Letellier, T.; Malgat, M.; Coquet, M.; Moretto, B.; Parrot-Roulaud, F.; Mazat, J.P. Mitochondrial Myopathy Studies on Permeabilized Muscle Fibers 1. Pediatr. Res. 1992, 32, 17–22. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Veksler, V.; Gellerich, F.N.; Saks, V.; Margreiter, R.; Kunz, W.S. Analysis of mitochondrial function in situ in permeabilized muscle fibers, tissues and cells. Nat. Protoc. 2008, 3, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Tepp, K.; Shevchuk, I.; Chekulayev, V.; Timohhina, N.; Kuznetsov, A.V.; Guzun, R.; Saks, V.; Kaambre, T. High efficiency of energy flux controls within mitochondrial interactosome in cardiac intracellular energetic units. Biochim. Biophys. Acta 2011, 1807, 1540–1561. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Williams, G.R. Respiratory enzymes in oxidative phosphorylation I. Kinetics of oxygen utilization. J. Biol. Chem. 1955, 217, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, D.; Garland, P.B. Citrate synthase from rat liver: [EC 4.1. 3.7 Citrate oxaloacetage-lyase (CoA-acetylating)]. Methods Enzymol. 1969, 13, 11–16. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Livak, N.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 5, 402–408. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial BM (g) | Final BM (g) | Left Kidney (g) | Uterine Mass (g) | Tibia Length (cm) | U/T Ratio | |
|---|---|---|---|---|---|---|
| INT-SED | 280.16 ± 4.21 | 314.17 ± 3.75 * | 1.090 ± 0.30 | 0.570 ± 0.40 *$ | 3.95 ± 0.004 | 0.144 ± 0.03 *$ |
| INT-RT | 260.22 ± 5.34 | 291.68 ± 6.83 *$# | 0.950 ± 0.16 | 0.680 ± 0.30 *$ | 3.96 ± 0.007 | 0.171 ± 0.02 *$ |
| OVX-SED | 270.95 ± 5.51 | 346.67 ± 5.56 | 0.957 ± 0.16 | 0.140 ± 0.13 | 3.94 ± 0.003 | 0.035 ± 0.02 |
| OVX-RT | 274.79 ± 4.89 | 341.10 ± 10.09 | 1.005 ± 0.21 | 0.150 ± 0.10 | 3.95 ± 0.005 | 0.038 ± 0.01 |
| EXPERIMENTAL GROUPS | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| INT-SED | INT-RT | OVX-SED | OVX-RT | |||||||||
| Time | SBP | HR | DP | SBP | HR | DP | SBP | HR | DP | SBP | HR | DP |
| PrOp | 133.7 ± 4.06 | 490.33 ± 18.42 | 66,414.5 ± 3316.93 | 135.22 ± 3.95 | 440.71 ± 23.19 | 61,234.29 ± 4731.12 | 136.2 ± 2.12 | 474.8 ± 7.85 | 64,713.2 ± 2530.21 | 132.44 ± 3.5 | 466.62 ± 18.99 | 62,945.25 ± 3449 |
| W1 | 136.5 ± 4.05 | 446 ± 12.37 | 61,103.2 ± 3001.87 | 134.44 ± 5.04 | 453.75 ± 20.35 c | 58,320.89 ± 2968.64 | 145.3 ± 6.43 | 453.44 ± 17.94 | 66,469.78 ± 4332.45 | 136.55 ± 6.42 | 434.77 ± 17.24 | 60,291.38 ± 2993.11 |
| W5 | 141.5 ± 5.54 | 438.2 ± 12.41 | 62,307.7 ± 3661.55 | 148.44 ± 5.29 | 398.88 ± 7.23 | 59,421.56 ± 2934.23 | 147.8 ± 4.86 | 408.5 ± 13.65 | 60,118.8 ± 2028.86 | 149.66 ± 5.92 | 372.11 ± 40.68 | 55,940.89 ± 6619.52 |
| W9 | 143.2 ± 4.85 | 416.9 ± 7.42 c | 59,868.7 * ± 2734.73 | 147.88 ± 5.29 | 395.11 ± 9.26 | 58,717.56 * ± 3161.71 | 158.9 ± 4.59 | 436.55 ± 13.35 a,b,c | 70,406.44 ± 2398.45 c | 142.11 ± 4.16 | 405.55 ± 10.27 | 57,919.56 * ± 3037.74 |
| W13 | 145.6 ± 4.35 *,a | 401.7 ± 16.3 a,b,c,d | 58,788.5 ± 3648.36 *,a | 148.22 ± 4.21 *,a,b | 387.66 ± 9.2 a,b | 57,565.44 ± 2460.72 * | 177.3 ± 3.54 a,b,c,d | 424.7 ± 14.79 | 73,034 ± 3266.77 c | 145.11 ± 2.64 * | 405.44 ± 16.89 | 58,689.11 ± 2222.66 * |
| Gene | Primer Sequence | Concentration | Efficiency (%) |
|---|---|---|---|
| B2m | F: CGAGACCGATGTATATGCTTGC | 100 nM | 102.99 |
| R: CCGGATCTGGAGTTAAACTGG | 100 nM | ||
| Psmc4 | F: TCGAGAAAGCATACAAGACCG | 150 nM | 102.63 |
| R: TCCTGGGTAAAGAGAAAACTAGC | 150 nM |
| Gene | Primer Sequence | Concentration | Efficiency (%) |
|---|---|---|---|
| CI (Ndufb4) | F: CGGCTTAAACGGGAGTATCTG | 150 nM | 100.83 |
| R: AAAGTGAGTTCTTGGGAGTGG | 150 nM | ||
| CII (Sdha) | F: TGTAAGAACATCAGAGCTGCG | 150 nM | 96.18 |
| R: CCCCTGTCAAACGTCTTCAG | 150 nM | ||
| CIII (Uqcrc1) | F: CCTTCAACATCTCCTACTCTGAG | 150 nM | 99.70 |
| R: TTTTGCCCCGAGTCACC | 150 nM | ||
| CIV (Cox4i1) | F: TTCGCTGAGATGAACAAGGG | 150 nM | 94.40 |
| R: GATCAAAGGTATGAGGGATGGG | 150 nM | ||
| Atps (Atp5f1a) | F: ATGTGGGCTTGTCTGTGTC | 150 nM | 98.07 |
| R: AGCATCCAGATCAGAACCAAAC | 150 nM | ||
| Pic (Slc25a3) | F: TCTACTTCAGGCTCCCTCG | 150 nM | 103.73 |
| R: TTCCTTTGCACTTTCAACACTG | 150 nM | ||
| Ant1 (Slc25a4) | F: TTTCAGTGTCTCTGTGCAGG | 150 nM | 104.27 |
| R: GTCACACTCTGGGCAATCAT | 150 nM | ||
| Cs | F: AAGGAAAGGCTAAGAACCCC | 150 nM | 95.37 |
| R: ATTCATCTCCGTCATGCCATAG | 300 nM | ||
| Ucp2 | F: GCCCCGAACCTTCTACAAG | 300 nM | 96.48 |
| R: ATTCATAGGCAGCCATCAGG | 300 nM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Queiroz, A.L.F.; Garcia, C.B.; Silva, J.P.M.O.; Cavalini, D.F.A.; Alexandrino, A.V.; Cunha, A.F.; Vercesi, A.E.; Castilho, R.F.; Shiguemoto, G.E. Preventive Effects of Resistance Training on Hemodynamics and Kidney Mitochondrial Bioenergetic Function in Ovariectomized Rats. Int. J. Mol. Sci. 2025, 26, 266. https://doi.org/10.3390/ijms26010266
Queiroz ALF, Garcia CB, Silva JPMO, Cavalini DFA, Alexandrino AV, Cunha AF, Vercesi AE, Castilho RF, Shiguemoto GE. Preventive Effects of Resistance Training on Hemodynamics and Kidney Mitochondrial Bioenergetic Function in Ovariectomized Rats. International Journal of Molecular Sciences. 2025; 26(1):266. https://doi.org/10.3390/ijms26010266
Chicago/Turabian StyleQueiroz, Anne L. F., Christopher B. Garcia, João P. M. O. Silva, Diego F. A. Cavalini, André V. Alexandrino, Anderson F. Cunha, Anibal E. Vercesi, Roger F. Castilho, and Gilberto E. Shiguemoto. 2025. "Preventive Effects of Resistance Training on Hemodynamics and Kidney Mitochondrial Bioenergetic Function in Ovariectomized Rats" International Journal of Molecular Sciences 26, no. 1: 266. https://doi.org/10.3390/ijms26010266
APA StyleQueiroz, A. L. F., Garcia, C. B., Silva, J. P. M. O., Cavalini, D. F. A., Alexandrino, A. V., Cunha, A. F., Vercesi, A. E., Castilho, R. F., & Shiguemoto, G. E. (2025). Preventive Effects of Resistance Training on Hemodynamics and Kidney Mitochondrial Bioenergetic Function in Ovariectomized Rats. International Journal of Molecular Sciences, 26(1), 266. https://doi.org/10.3390/ijms26010266