
GLP-1 Receptor Agonists: A New Treatment in Parkinson’s Disease




Abstract
1. Introduction
2. T2DM and PD: Diseases with Overlapping Pathophysiology?
3. GLP-1 and GLP-1 Receptor
4. GLP-1 Receptor Agonists
5. GLP-1 Receptor Agonists in Parkinson’s Disease Treatment
5.1. Evidence from Animal Models: Preclinical Studies
5.2. Evidence from Clinical Studies
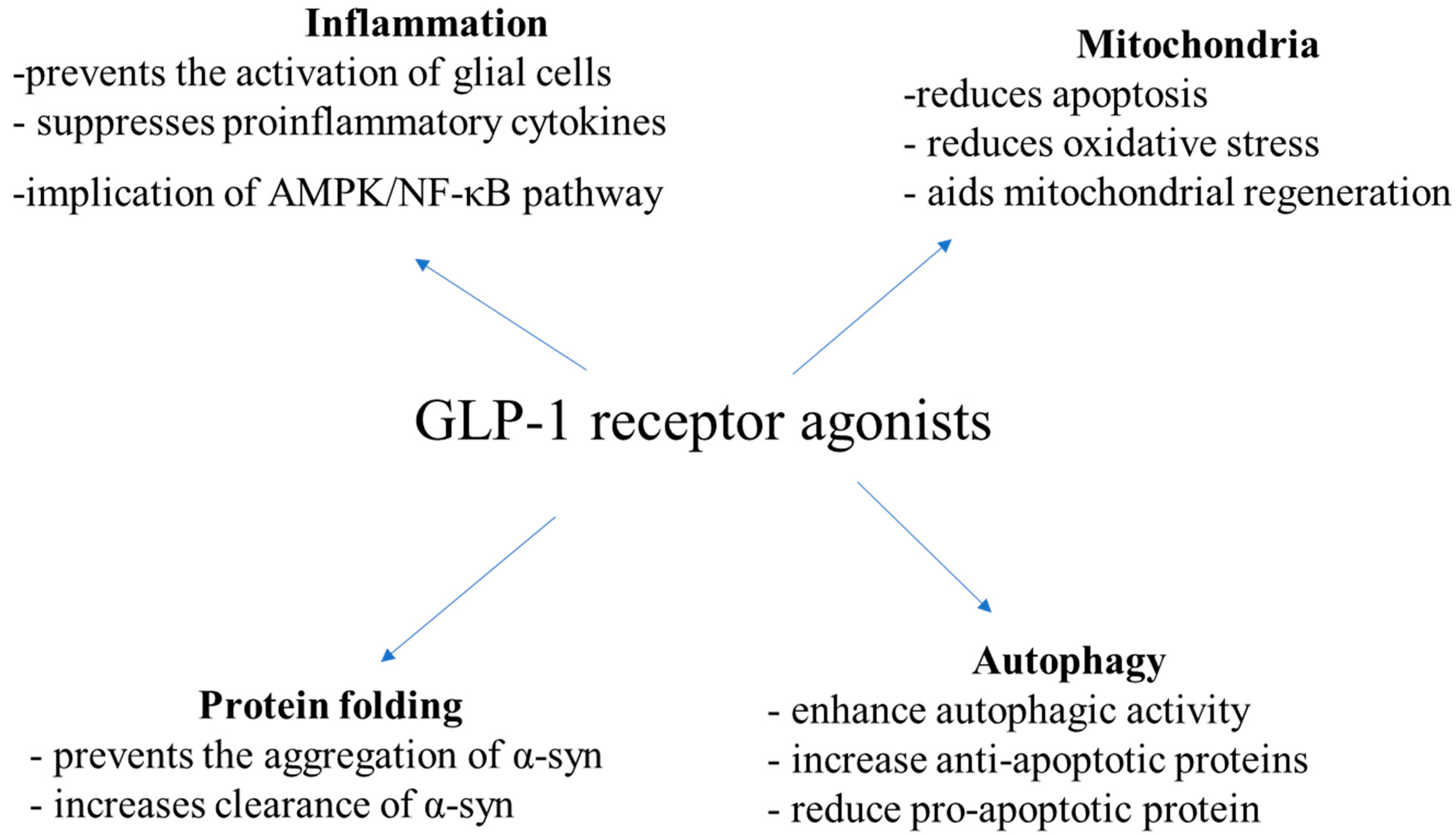
5.3. Mechanism of Action
6. Research Gaps and Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fahn, S. Description of Parkinson’s disease as a clinical syndrome. Ann. N. Y. Acad. Sci. 2003, 991, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kalinderi, K.; Papaliagkas, V.; Fidani, L. The Genetic Landscape of Sleep Disorders in Parkinson’s Disease. Diagnostics 2024, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Kalinderi, K.; Papaliagkas, V.; Fidani, L. Current genetic data on depression and anxiety in Parkinson’s disease patients. Parkinsonism. Relat. Disord. 2024, 118, 105922. [Google Scholar] [CrossRef] [PubMed]
- Kalinderi, K.; Papaliagkas, V.; Fidani, L. Pharmacogenetics and levodopa induced motor complications. Int. J. Neurosci. 2019, 129, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, M.; Moraitou, D.; Tata, D.A.; Kalinderi, K.; Papamitsou, T.; Papaliagkas, V. Alzheimer’s Disease as Type 3 Diabetes: Common Pathophysiological Mechanisms between Alzheimer’s Disease and Type 2 Diabetes. Int. J. Mol. Sci. 2022, 23, 2687. [Google Scholar] [CrossRef] [PubMed]
- Svenningsson, P.; Wirdefeldt, K.; Yin, L.; Fang, F.; Markaki, I.; Efendic, S.; Ludvigsson, J.F. Reduced Incidence of Parkinson’s Disease after Dipeptidyl Peptidase-4 Inhibitors—A Nationwide Case-Control Study. Mov. Disord. 2016, 31, 1422–1423. [Google Scholar] [CrossRef]
- Brauer, R.; Wei, L.; Ma, T.; Athauda, D.; Girges, C.; Vijiaratnam, N.; Auld, G.; Whittlesea, C.; Wong, I.; Foltynie, T. Diabetes Medications and Risk of Parkinson’s Disease: A Cohort Study of Patients with Diabetes. Brain 2020, 143, 3067–3076. [Google Scholar] [CrossRef] [PubMed]
- Barkas, F.; Elisaf, M.; Milionis, H. Protection against Stroke with Glucagon-like Peptide 1 Receptor Agonists: A Systematic Review and Meta-analysis. Eur. J. Neurol. 2019, 26, 559–565. [Google Scholar] [CrossRef]
- Malhotra, K.; Katsanos, A.H.; Lambadiari, V.; Goyal, N.; Palaiodimou, L.; Kosmidou, M.; Krogias, C.; Alexandrov, A.V.; Tsivgoulis, G. GLP-1 Receptor Agonists in Diabetes for Stroke Prevention: A Systematic Review and Meta-Analysis. J. Neurol. 2020, 1, 3. [Google Scholar] [CrossRef]
- Perry, T.; Lahiri, D.K.; Chen, D.; Zhou, J.; Shaw, K.T.Y.; Egan, J.M.; Greig, N.H. A Novel Neurotrophic Property of Glucagon-like Peptide 1: A Promoter of Nerve Growth Factor-Mediated Differentiation in PC12 Cells. J. Pharmacol. Exp. Ther. 2002, 300, 958–966. [Google Scholar] [CrossRef]
- Perry, T.; Haughey, N.J.; Mattson, M.P.; Egan, J.M.; Greig, N.H. Protection and Reversal of Excitotoxic Neuronal Damage by Glucagon-Like Peptide-1 and Exendin-4. J. Pharmacol. Exp. Ther. 2002, 302, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Gilman, C.P.; Perry, T.A.; Furukawa, K.; Grieg, N.H.; Egan, J.M.; Mattson, M.P. Glucagon-like Peptide 1 Modulates Calcium Responses to Glutamate and Membrane Depolarization in Hippocampal Neurons. J. Neurochem. 2003, 87, 1137–1144. [Google Scholar] [CrossRef]
- Perry, T.; Lahiri, D.K.; Sambamurti, K.; Chen, D.; Mattson, M.P.; Egan, J.M.; Greig, N.H. Glucagon-like Peptide-1 Decreases Endogenous Amyloid-Beta Peptide (Abeta) Levels and Protects Hippocampal Neurons from Death Induced by Abeta and Iron. J. Neurosci. Res. 2003, 72, 603–612. [Google Scholar] [CrossRef]
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Komici, K.; Femminella, G.D.; Bencivenga, L.; Rango, G.; Pagano, G. Diabetes mellitus and Parkinson’s disease: A systematic review and meta-analyses. J. Parkinson’s Dis. 2021, 11, 1585–1596. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.W.; Hsieh, T.F.; Li, C.I.; Liu, C.S.; Lin, W.Y.; Chiang, J.H.; Li, T.C.; Lin, C.C. Increased risk of Parkinson disease with diabetes mellitus in a population-based study. Medicine 2017, 96, e5921. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Li, H.; Yan, H.; Zhang, P.; Chang, L.; Li, T. Risk of Parkinson disease in diabetes mellitus: An updated meta-analysis of population- based cohort studies. Medicine 2016, 95, e3549. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Han, K.D.; Kwan, H.; Park, S.E.; Park, Y.G.; Kim, Y.H.; Yoo, S.J.; Rhee, E.J.; Lee, W.Y. Association between glycemic status and the risk of Parkinson disease: A nationwide population-based study. Diabetes Care 2020, 43, 2169–2175. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, F.; Moretti, A.; Villa, R.F. Incretin-based drugs as potential therapy for neurodegenerative diseases: Current status and perspectives. Pharmacol. Ther. 2022, 239, 108277. [Google Scholar] [CrossRef]
- Hong, C.T.; Chen, K.Y.; Wang, W.; Chiu, J.Y.; Wu, D.; Chao, T.Y.; Hu, C.J.; Chau, K.D.; Bamodu, O.A. Insulin resistance promotes Parkinson’s disease through aberrant expression of alfa-Synuclein, mitochondrial dysfunction, and deregulation of the polo-like kinase 2 signaling. Cells 2020, 9, 740. [Google Scholar] [CrossRef]
- Wang, L.; Zhai, Y.Q.; Xu, L.L.; Qiao, C.; Sun, X.L.; Ding, J.H.; Lu, M.; Hu, G. Metabolic inflammation exacerbates dopaminergic neuronal degeneration in response to acute MPTP challenge in type 2 diabetes mice. Exp. Neurol. 2014, 251, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, D.; Clavero, P.; Salas, C.G.; Lamet, I.; Arbizu, J.; Gonzalez-Redondo, R.; Obeso, J.A.; Rodriguez-Oroz, M.C. Posterior parietooccipital hypometabolism may differentiate mild cognitive impairment from dementia in Parkinson’s disease. Eur. J. Nucl. Med. Mol. Imaging 2012, 39, 1767–1777. [Google Scholar] [CrossRef]
- Hosokai, Y.; Nishio, Y.; Hirayama, K.; Takeda, A.; Ishioka, T.; Sawada, Y.; Suzuki, K.; Itoyama, Y.; Takahashi, S.; Fukuda, H.; et al. Distinct patterns of regional cerebral glucose metabolism in Parkinson’s disease with and without mild cognitive impairment. Mov. Disord. 2009, 24, 854–862. [Google Scholar] [CrossRef]
- Barbiero, J.K.; Santiago, R.M.; Persike, D.S.; da Silva Fernandes, M.J.; Tonin, F.S.; da Cunha, C.; Lucio Boschen, S.; Lima, M.M.; Vital, M.A. Neuroprotective effects of peroxisome proliferator-activated receptor alpha and gamma agonists in model of parkinsonism induced by intranigral 1-methyl-4-phenyl-1,2,3,6-tetrahyropyridine. Behav. Brain Res. 2014, 274, 390–399. [Google Scholar] [CrossRef]
- Iravanpour, F.; Dargahi, L.; Rezaei, M.; Haghani, M.; Heidari, R.; Valian, N.; Ahmadiani, A. Intranasal insulin improves mitochondrial function and attenuates motor deficits in a rat 6-OHDA model of Parkinson’s disease. CNS Neurosci. Ther. 2021, 27, 308–319. [Google Scholar] [CrossRef]
- Novak, P.; Pimentel Maldonado, D.A.; Novak, V. Safety and preliminary efficacy of intranasal insulin for cognitive impairment in Parkinson disease and multiple system atrophy: A double-blinded placebo-controlled pilot study. PLoS ONE 2019, 14, e214364. [Google Scholar] [CrossRef] [PubMed]
- Hölscher, C. Brain insulin resistance: Role in neurodegenerative disease and potential for targeting. Expert Opin. Investig. Drugs 2020, 29, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Nauck, M.A. The incretin system: Glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of action and therapeutic application of glucagon-like peptide-1. Cell Metab. 2018, 27, 740–756. [Google Scholar] [CrossRef]
- Pyke, C.; Heller, R.S.; Kirk, R.K.; Orskov, C.; Reedtz-Runge, S.; Kaastrup, P.; Hvelplund, A.; Bardram, L.; Calatayud, D.; Knudsen, L.B. GLP-1 receptor localization in monkey and human tissue: Novel distribution revealed with extensively validated monoclonal antibody. Endocrinology 2014, 155, 1280–1290. [Google Scholar] [CrossRef]
- Kopp, K.O.; Glotfelty, E.J.; Li, Y.; Greig, N.H. Glucagon-like Peptide-1 (GLP-1) Receptor Agonists and Neuroinflammation: Implications for Neurodegenerative Disease Treatment. Pharmacol. Res. 2022, 186, 106550. [Google Scholar] [CrossRef] [PubMed]
- Kalinderi, K.; Goula, V.; Sapountzi, E.; Tsinopoulou, V.R.; Fidani, L. Syndromic and Monogenic Obesity: New Opportunities Due to Genetic-Based Pharmacological Treatment. Children 2024, 11, 153. [Google Scholar] [CrossRef] [PubMed]
- Glotfelty, E.J.; Olson, L.; Karlsson, T.E.; Li, Y.; Greig, N.H. Glucagon-like peptide-1 (GLP-1)-based receptor agonists as a treatment for Parkinson’s disease. Expert Opin. Investig. Drugs 2020, 29, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Salameh, T.S.; Rhea, E.M.; Talbot, K.; Banks, W.A. Brain uptake pharmacokinetics of incretin receptor agonists showing promise as Alzheimer’s and Parkinson’s disease therapeutics. Biochem. Pharmacol. 2020, 180, 114187. [Google Scholar] [CrossRef] [PubMed]
- Rhea, E.M.; Babin, A.; Thomas, P.; Omer, M.; Weaver, R.; Hansen, K.; Banks, W.A.; Talbot, K. Brain uptake pharmacokinetics of albiglutide, dulaglutide, tirzepatide, and DA5-CH in the search for new treatments of Alzheimer’s and Parkinson’s diseases. Tissue Barriers 2023, 2292461, Online ahead of print. [Google Scholar] [CrossRef]
- Jalewa, J.; Sharma, M.K.; Holscher, C. Novel incretin analogues improve autophagy and protect from mitochondrial stress induced by rotenone in SH-SY5Y cells. J. Neurochem. 2016, 139, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Kishi, T.; Lee, C.E.; Choi, B.J.; Fang, H.; Hollenberg, A.N.; Drucker, D.J.; Elmquist, J.K. Glucagon-like peptide-1-responsive catecholamine neurons in the area postrema link peripheral glucagon-like peptide-1 with central autonomic control sites. J. Neurosci. 2003, 23, 2939–2946. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.D.; Yi, L.X.; Wang, D.Q.; Lim, T.M.; Tan, E.K. Role of dopamine in the pathophysiology of Parkinson’s disease. Transl. Neurodegener. 2023, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Elbassuoni, E.A.; Ahmed, R.F. Mechanism of the neuroprotective effect of GLP-1 in a rat model of Parkinson’s with pre-existing diabetes. Neurochem. Int. 2019, 131, 104583. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Perry, T.; Kindy, M.S.; Harvey, B.K.; Tweedie, D.; Holloway, H.W.; Powers, K.; Shen, H.; Egan, J.M.; Sambamurti, K.; et al. GLP-1 receptor stimulation preserves primary cortical and dopaminergic neurons in cellular and rodent models of stroke and Parkinsonism. Proc. Natl. Acad. Sci. USA 2009, 106, 1285–1290. [Google Scholar] [CrossRef]
- Harkavyi, A.; Abuirmeileh, A.; Lever, R.; Kingsbury, A.E.; Biggs, C.S.; Whitton, P.S. Glucagon-like peptide 1 receptor stimulation reverses key deficits in distinct rodent models of Parkinson’s disease. J. Neuroinflamm. 2008, 5, 19. [Google Scholar] [CrossRef]
- Aksoy, D.; Solmaz, V.; Cavusoglu, T.; Meral, A.; Ates, U.; Erbas, O. Neuroprotective Effects of Eexenatide in a Rotenone-Induced Rat Model of Parkinson’s Disease. Am. J. Med. Sci. 2017, 354, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Chen, Y.H.; Liang, K.C.; Jheng, Y.S.; Jhao, J.J.; Su, M.T.; Lee-Chen, G.J.; Hsieh-Li, H.M. Exendin-4 Protected against Cognitive Dysfunction in Hyperglycemic Mice Receiving an Intrahippocampal Lipopolysaccharide Injection. PLoS ONE 2012, 7, e39656. [Google Scholar] [CrossRef]
- Bu, L.L.; Liu, Y.Q.; Shen, Y.; Fan, Y.; Yu, W.B.; Jiang, D.L.; Tang, Y.L.; Yang, Y.J.; Wu, P.; Zuo, C.T.; et al. Neuroprotection of Exendin-4 by Enhanced Autophagy in a Parkinsonian Rat Model of alpha-Synucleinopathy. Neurotherapeutics 2021, 18, 962–978. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.P.; Kam, T.I.; Panicker, N.; Kim, S.; Oh, Y.; Park, J.S.; Kwon, S.H.; Park, Y.J.; Karuppagounder, S.S.; Park, H.; et al. Block of A1 astrocyte conversion by microglia is neuroprotective in models of Parkinson’s disease. Nat. Med. 2018, 24, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; McLean, B.A.; Baggio, L.L.; Koehler, J.A.; Hammoud, R.; Rittig, N.; Yabut, J.M.; Seeley, R.J.; Brown, T.J.; Drucker, D.J. Central glucagon-like peptide 1 receptor activation inhibits Toll-like receptor agonist-induced inflammation. Cell Metab. 2024, 36, 130–143.e5. [Google Scholar] [CrossRef]
- Lv, M.; Xue, G.; Cheng, H.; Meng, P.; Lian, X.; Hölscher, C.; Li, D. The GLP-1/GIP dual-receptor agonist DA5-CH inhibits the NF-kappaB inflammatory pathway in the MPTP mouse model of Parkinson’s disease more effectively than the GLP-1 single-receptor agonist NLY01. Brain Behav. 2021, 11, e2231. [Google Scholar] [CrossRef]
- Chen, S.C.; Yu, S.J.; Li, Y.Z.; Lecca, D.; Glotfelty, E.; Kim, H.K.; Choi, H.I.; Hoffer, B.J.; Greig, N.H.; Kim, D.S.; et al. Post-treatment with PT302, a long-acting Exendin-4 sustained release GLP-1R agonist in Parkinson disease formulation, reduces dopaminergic neurodegeneration in a 6-Hydroxydopamine rat model of Parkinson’s Disease. Sci. Rep. 2018, 8, 10722. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.T.; Chen, Y.H.; Wang, V.; Huang, E.Y.; Ma, K.H.; Greig, N.H.; Jung, J.; Choi, H.I.; Olson, L.; Hoffer, B.J.; et al. PT320, a Sustained-Release GLP-1 Receptor Agonist, Ameliorates L-DOPA-Induced Dyskinesia in a Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 4687. [Google Scholar] [CrossRef]
- Liu, W.; Jalewa, J.; Sharma, M.; Li, G.; Li, L.; Holscher, C. Neuroprotective Effects of Lixisenatide and Liraglutide in the 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine Mouse Model of Parkinson’s Disease. Neuroscience 2015, 303, 42–50. [Google Scholar] [CrossRef]
- Hansen, H.H.; Fabricius, K.; Barkholt, P.; Mikkelsen, J.D.; Jelsing, J.; Pyke, C.; Knudsen, L.B.; Vrang, N. Characterization of liraglutide, a glucagon-like peptide-1 (GLP-1) receptor agonist, in rat partial and full nigral 6-hydroxydopamine lesion models of Parkinson’s disease. Brain Res. 2016, 1646, 354–365. [Google Scholar] [CrossRef]
- Yu, H.Y.; Sun, T.; Wang, Z.; Li, H.; Xu, D.; An, J.; Wen, L.L.; Li, J.Y.; Li, W.; Feng, J. Exendin-4 and linagliptin attenuate neuroinflammation in a mouse model of Parkinson’s disease. Neural. Regen. Res. 2023, 18, 1818–1826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.P.; Zhang, L.Y.; Li, L.; Holscher, C. Semaglutide is Neuroprotective and Reduces alpha-Synuclein Levels in the Chronic MPTP Mouse Model of Parkinson’s Disease. J. Parkinson’s Dis. 2019, 9, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.P.; Zhang, L.Y.; Li, L.; Holscher, C. Neuroprotective effects of the novel GLP-1 long acting analogue semaglutide in the MPTP Parkinson’s disease mouse model. Neuropeptides 2018, 71, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Chen, Y.M.; Li, L.; Holscher, C. Neuroprotective effects of (Val8)GLP-1-Glu-PAL in the MPTP Parkinson’s disease mouse model. Behav. Brain Res. 2015, 293, 107–113. [Google Scholar] [CrossRef]
- Badawi, G.A.; Abd, E.L.; Fattah, M.A.; Zaki, H.F.; El Sayed, M.I. Sitagliptin and liraglutide reversed nigrostriatal GLP-1R agonist in Parkinson disease degeneration of rodent brain in rotenone-induced Parkinson’s disease. Inflammopharmacology 2017, 25, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Badawi, G.A.; Abd El Fattah, M.A.; Zaki, H.F.; El Sayed, M.I. Sitagliptin and Liraglutide Modulate L-dopa Effect and Attenuate Dyskinetic Movements in Rotenone-Lesioned Rats. Neurotox. Res. 2019, 35, 635–653. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.J.; Li, D.F.; Feng, P.; Li, L.; Xue, G.F.; Li, G.; Hölscher, C. A novel dual GLP-1 and GIP incretin receptor agonist is neuroprotective in a mouse model of Parkinson’s disease by reducing chronic inflammation in the brain. Neuroreport 2016, 27, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Xue, G.F.; Lijun, C.; Feng, P.; Li, D.; Li, L.; Li, G.; Hölscher, C. A novel dual GLP-1 and GIP receptor agonist is neuroprotective in the MPTP mouse model of Parkinson’s disease by increasing expression of BNDF. Brain Res. 2016, 1634, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jalewa, J.; Sharma, M.K.; Gengler, S.; Hölscher, C. A novel GLP-1/GIP dual receptor agonist protects from 6-OHDA lesion in a rat model of Parkinson’s disease. Neuropharmacology 2017, 117, 238–248. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Jin, Q.Q.; Holscher, C.; Li, L. Glucagon-like peptide-1/glucose-dependent insulinotropic polypeptide dual receptor agonist DA-CH5 is superior to exendin-4 in protecting neurons in the 6-hydroxydopamine rat Parkinson model. Neural Regen. Res. 2021, 16, 1660–1670. [Google Scholar]
- Yuan, Z.Y.; Li, D.F.; Feng, P.; Xue, G.; Ji, C.; Li, G.; Hölscher, C. A novel GLP-1/GIP dual agonist is more effective than liraglutide in reducing inflammation and enhancing GDNF release in the MPTP mouse model of Parkinson’s disease. Eur. J. Pharmacol. 2017, 812, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, C.; Zhang, Z.; Zhang, Z.; Jin, Q.Q.; Li, L.; Hölscher, C. DA5-CH and Semaglutide Protect against Neurodegeneration and Reduce α-Synuclein Levels in the 6-OHDA Parkinson’s Disease Rat Model. Parkinson’s Dis. 2022, 2022, 1428817. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tu, L.; Gu, R.; Yang, X.L.; Liu, X.J.; Zhang, G.P.; Wang, Q.; Ren, Y.P.; Wang, B.J.; Tian, J.Y. Neuroprotection of GLP-1/GIP receptor agonist via inhibition of mitochondrial stress by AKT/JNK pathway in a Parkinson’s disease model. Life Sci. 2020, 256, 117824. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, M.; Li, Z.; Ling, Y.; Zhai, L.; Yuan, Y.; Ma, H.; Hao, L.; Li, Z.; Zhang, Z.; et al. A Dual GLP-1/GIP Receptor Agonist Is More Effective than Liraglutide in the A53T Mouse Model of Parkinson’s Disease. Parkinson’s Dis. 2023, 2023, 7427136. [Google Scholar] [CrossRef]
- Feng, P.; Zhang, X.J.; Li, D.F.; Ji, C.; Yuan, Z.; Wang, R.; Xue, G.; Li, G.; Hölscher, C. Two novel dual GLP-1/GIP receptor agonists are neuroprotective in the MPTP mouse model of Parkinson’s disease. Neuropharmacology 2018, 133, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Hölscher, C. Novel dual GLP-1/GIP receptor agonists show neuroprotective effects in Alzheimer’s and Parkinson’s disease models. Neuropharmacology 2018, 136, 251–259. [Google Scholar] [CrossRef]
- Aviles-Olmos, I.; Dickson, J.; Kefalopoulou, Z.; Djamshidian, A.; Ell, P.; Soderlund, T.; Whitton, P.; Wyse, R.; Isaacs, T.; Lees, A.; et al. Exenatide and the treatment of patients with Parkinson’s disease. J. Clin. Investig. 2013, 123, 2730–2736. [Google Scholar] [CrossRef] [PubMed]
- Athauda, D.; Maclagan, K.; Skene, S.S.; Bajwa-Joseph, M.; Letchford, D.; Chowdhury, K.; Hibbert, S.; Budnik, N.; Zampedri, L.; Dickson, J.; et al. Exenatide once weekly versus placebo in Parkinson’s disease: A randomised, double-blind, placebo-controlled trial. Lancet 2017, 390, 1664–1675. [Google Scholar] [CrossRef]
- Malatt, C.; Wu, T.; Bresee, C.; Hogg, E.J.; Wertheimer, J.C.; Tan, E.; Pomeroy, H.; Obialisi, G.; Tagliati, M. Liraglutide Improves Non-Motor Function and Activities of Daily Living in Patients with Parkinson’s disease: A Randomized, Double-Blind, Placebo-Controlled Trial. (P9.11-005). Neurology 2022, 98 (18_supplement). [Google Scholar] [CrossRef]
- Meissner, W.G.; Remy, P.; Giordana, C.; Maltete, D.; Damier, P.; Houeto, J.L.; Geny, C.; Hopes, L.; Durif, F.; Defer, G.; et al. Multicenter, randomized, placebo-controlled, double-blind, parallel-group proof-of-concept study of lixisenatide in patients with early Parkinson’s disease (PD): The LIXIPARK trial (NCT03439943). [abstract]. Mov Disord. 2023, 38 (Suppl. S1), S41. [Google Scholar]
- Aviles-Olmos, I.; Dickson, J.; Kefalopoulou, Z.; Djamshidian, A.; Kahan, J.; Ell, P.; Whitton, P.; Wyse, R.; Isaacs, T.; Lees, A.; et al. Motor and Cognitive Advantages Persist 12 Months After Exenatide Exposure in Parkinson’s Disease. J. Parkinson’s Dis. 2014, 4, 337–344. [Google Scholar] [CrossRef]
- Athauda, D.; Maclagan, K.; Budnik, N.; Zampedri, L.; Hibbert, S.; Skene, S.S.; Chowdhury, K.; Aviles-Olmos, I.; Limousin, P.; Foltynie, T. What Effects Might Exenatide have on Non-Motor Symptoms in Parkinson’s Disease: A Post Hoc Analysis. J. Parkinson’s Dis. 2018, 8, 247–258. [Google Scholar] [CrossRef]
- Athauda, D.; Maclagan, K.; Budnik, N.; Zampedri, L.; Hibbert, S.; Aviles-Olmos, I.; Chowdhury, K.; Skene, S.S.; Limousin, P.; Foltynie, T. Post hoc analysis of the Exenatide-PD trial-Factors that predict response. Eur. J. Neurosci. 2019, 49, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Athauda, D.; Gulyani, S.; Karnati, H.K.; Li, Y.; Tweedie, D.; Mustapic, M.; Chawla, S.; Chowdhury, K.; Skene, S.S.; Greig, N.H.; et al. Utility of Neuronal-Derived Exosomes to Examine Molecular Mechanisms that Affect Motor Function in Patients with Parkinson Disease: A Secondary Analysis of the Exenatide-PD Trial. JAMA Neurol. 2019, 76, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Gullo, F.; Ceriani, M.; D’Aloia, A.; Wanke, E.; Constanti, A.; Costa, B.; Lecchi, M. Plant Polyphenols and Exendin-4 Prevent Hyperactivity and TNFalpha Release in LPS-Treated In vitro Neuron/Astrocyte/Microglial Networks. Front. Neurosci. 2017, 11, 500. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Jeon, S.J.; Cho, K.S.; Moon, E.; Sapkota, A.; Jun, H.S.; Ryu, J.H.; Choi, J.W. Activation of Glucagon-Like Peptide-1 Receptor Promotes Neuroprotection in Experimental Autoimmune Encephalomyelitis by Reducing Neuroinflammatory Responses. Mol. Neurobiol. 2018, 55, 3007–3020. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Tang, X.Q.; Liu, H.; Mao, X.F.; Wang, Y.X. Both classic Gs-cAMP/PKA/CREB and alternative GscAMP/PKA/p38 beta/CREB signal pathways mediate exenatide-stimulated expression of M2 microglial markers. J. Neuroimmunol. 2018, 316, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Yoon, G.; Kim, Y.K.; Song, J. Glucagon-like peptide-1 suppresses neuroinflammation and improves neural structure. Pharmacol. Res. 2020, 152, 104615. [Google Scholar] [CrossRef] [PubMed]
- Gea, L.P.; da Rosa, E.D.; Panizzutti, B.S.; de Aguiar, É.Z.; de Oliveira, L.F.; Ferrari, P.; Piato, A.; Gomez, R.; Colombo, R.; Rosa, A.R. Reduction of hippocampal IL-6 levels in LPS-injected rats following acute exendin-4 treatment. Naunyn. Schmiedebergs Arch. Pharmacol. 2020, 393, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Zhang, Y.; Chen, J.; Wu, P.; Dong, Y.; Wang, Y. Neuroprotective effects of liraglutide against inflammation through the AMPK/NF-kappaB pathway in a mouse model of Parkinson’s disease. Metab. Brain Dis. 2022, 37, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, Y.; Li, L.; Holscher, C. Neuroprotective effects of geniposide in the MPTP mouse model of Parkinson’s disease. Eur. J. Pharmacol. 2015, 768, 21–27. [Google Scholar] [CrossRef]
- Nassar, N.N.; Al-Shorbagy, M.Y.; Arab, H.H.; Abdallah, D.M. Saxagliptin: A novel antiparkinsonian approach. Neuropharmacology 2015, 89, 308–317. [Google Scholar] [CrossRef]
- Oh, Y.S.; Jun, H.S. Effects of Glucagon-Like Peptide-1 on Oxidative Stress and Nrf2 Signaling. Int. J. Mol. Sci. 2018, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Dong, Y.; Chen, J.; Guan, T.; Cao, B.; Zhang, Y.; Qi, Y.; Guan, Z.; Wang, Y. Liraglutide Regulates Mitochondrial Quality Control System Through PGC-1alpha in a Mouse Model of Parkinson’s Disease. Neurotox. Res. 2022, 40, 286–297. [Google Scholar] [CrossRef]
- Lin, T.K.; Lin, K.J.; Lin, H.Y.; Lin, K.L.; Lan, M.Y.; Wang, P.W.; Wang, T.J.; Wang, F.S.; Tsai, P.C.; Liou, C.W.; et al. Glucagon-like Peptide-1 Receptor Agonist Ameliorates 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP) Neurotoxicity Through Enhancing Mitophagy Flux and Reducing alpha-Synuclein and Oxidative Stress. Front. Mol. Neurosci. 2021, 14, 697440. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tweedie, D.; Mattson, M.P.; Holloway, H.W.; Greig, N.H. Enhancing the GLP-1 receptor signaling pathway leads to proliferation and neuroprotection in human neuroblastoma cells. J. Neurochem. 2010, 113, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.L.; Zhang, Y.S.; Shi, Z.S.; Lu, D.; Li, T.; Ding, Y.; Ruan, Y.; Xu, A. The Neuroprotection of Liraglutide Against Ischaemia-induced Apoptosis through the Activation of the PI3K/AKT and MAPK Pathways. Sci. Rep. 2016, 6, 26859. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.P.; Scott, S.V.; Kim, J.; Klionsky, D.J. The itinerary of a vesicle component, Aut7p/Cvt5p, terminates in the yeast vacuole via the autophagy/Cvt pathways. J. Biol. Chem. 2000, 275, 5845–5851. [Google Scholar] [CrossRef]
- Liu, D.X.; Zhao, C.S.; Wei, X.N.; Ma, Y.P.; Wu, J.K. Semaglutide Protects against 6-OHDA Toxicity by Enhancing Autophagy and Inhibiting Oxidative Stress. Parkinson’s Dis. 2022, 2022, 6813017. [Google Scholar] [CrossRef]


{kind=link}
| ClinicalTrial.Gov Identifier | Drug | Result | Reference |
|---|---|---|---|
| NCT01174810 | Exendin-4 | Improvement in MDS-UPDRS and Mattis DRS-2 | [68] |
| NCT01971242 | Exendin-4 | Improvement in MDS-UPDRS | [69] |
| NCT02953665 | Liraglutide | Improvement in daily living of PD patients | [70] |
| NCT03439943 | Lixisenatide | Improvement in MDS-UPDRS III | [71] |
| Title | ClinicalTrials.gov ID | Phase | Status | Intervention | Results Overview |
|---|---|---|---|---|---|
| Exenatide Once Weekly Over 2 Years as a Potential Disease Modifying Treatment for Parkinson’s Disease (Exenatide-PD3) | NCT04232969 | Phase 3 | Active, not recruiting | Drug: Exenatide extended release 2 mg (Bydureon) | Result not yet available |
| Effects of Exenatide on Motor Function and the Brain | NCT03456687 | Phase 1 | Completed | Drug: Exenatide | No result available |
| Exenatide Treatment in Parkinson’s Disease | NCT04305002 | Phase 2 | Active, not recruiting | Drug: Exenatide Other: Placebo | No result available |
| SR-Exenatide (PT320) to Evaluate Efficacy and Safety in Patients With Early Parkinson’s Disease | NCT04269642 | Phase 2 | Active, not recruiting | Drug: PT320 2.0 mg Placebo Drug: PT320 2.0 mg Drug: PT320 2.5 mg | Unknown status |
| GLP1R in Parkinson’s Disease (GIPD) | NCT03659682 | Phase 2 | Not yet recruiting | Drug: Semaglutide | No result available |
| A Clinical Study of NLY01 in Patient’s With Early Parkinson’s Disease | NCT04154072 | Phase 2 | Active, not recruiting | Drug: NLY01 Drug: Vehicle | No result available |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalinderi, K.; Papaliagkas, V.; Fidani, L. GLP-1 Receptor Agonists: A New Treatment in Parkinson’s Disease. Int. J. Mol. Sci. 2024, 25, 3812. https://doi.org/10.3390/ijms25073812
Kalinderi K, Papaliagkas V, Fidani L. GLP-1 Receptor Agonists: A New Treatment in Parkinson’s Disease. International Journal of Molecular Sciences. 2024; 25(7):3812. https://doi.org/10.3390/ijms25073812
Chicago/Turabian StyleKalinderi, Kallirhoe, Vasileios Papaliagkas, and Liana Fidani. 2024. "GLP-1 Receptor Agonists: A New Treatment in Parkinson’s Disease" International Journal of Molecular Sciences 25, no. 7: 3812. https://doi.org/10.3390/ijms25073812
APA StyleKalinderi, K., Papaliagkas, V., & Fidani, L. (2024). GLP-1 Receptor Agonists: A New Treatment in Parkinson’s Disease. International Journal of Molecular Sciences, 25(7), 3812. https://doi.org/10.3390/ijms25073812