
The Potential of Natural Carotenoids-Containing Sericin of the Domestic Silkworm Bombyx mori
 and
and
Abstract
1. Introduction
2. Results
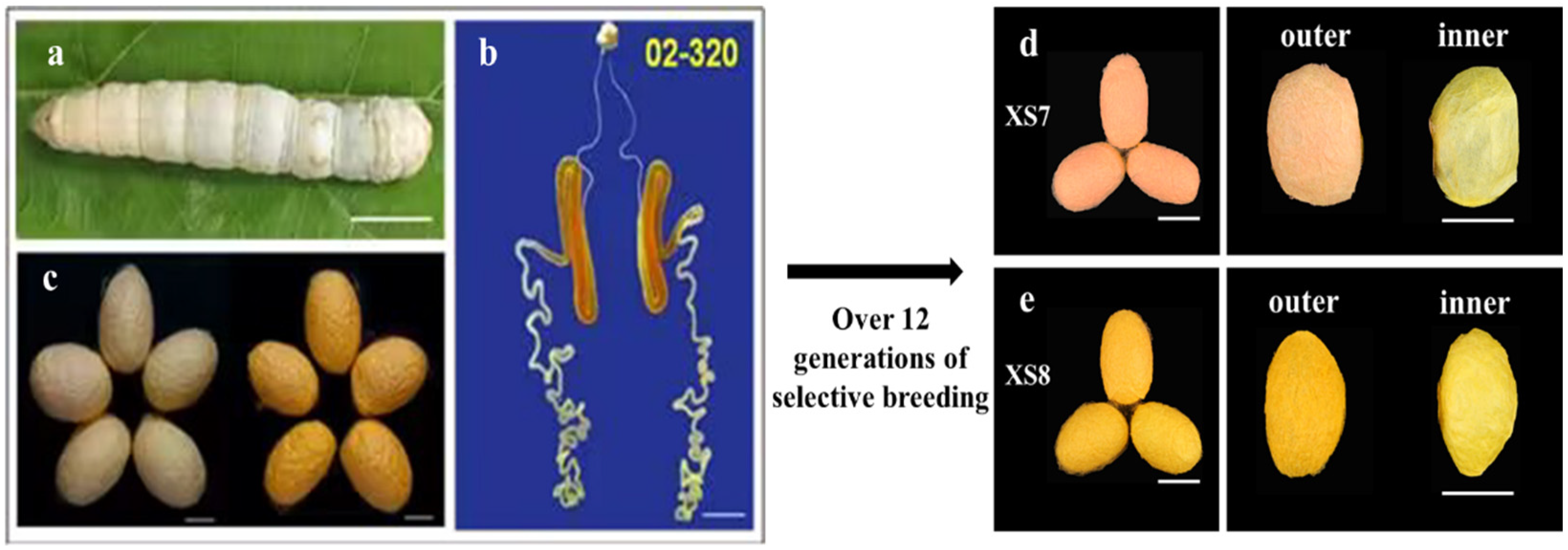
2.1. Breeding of Silkworm Strains XS7 and XS8
2.2. Differences in Cocoon Color between XS7 and XS8
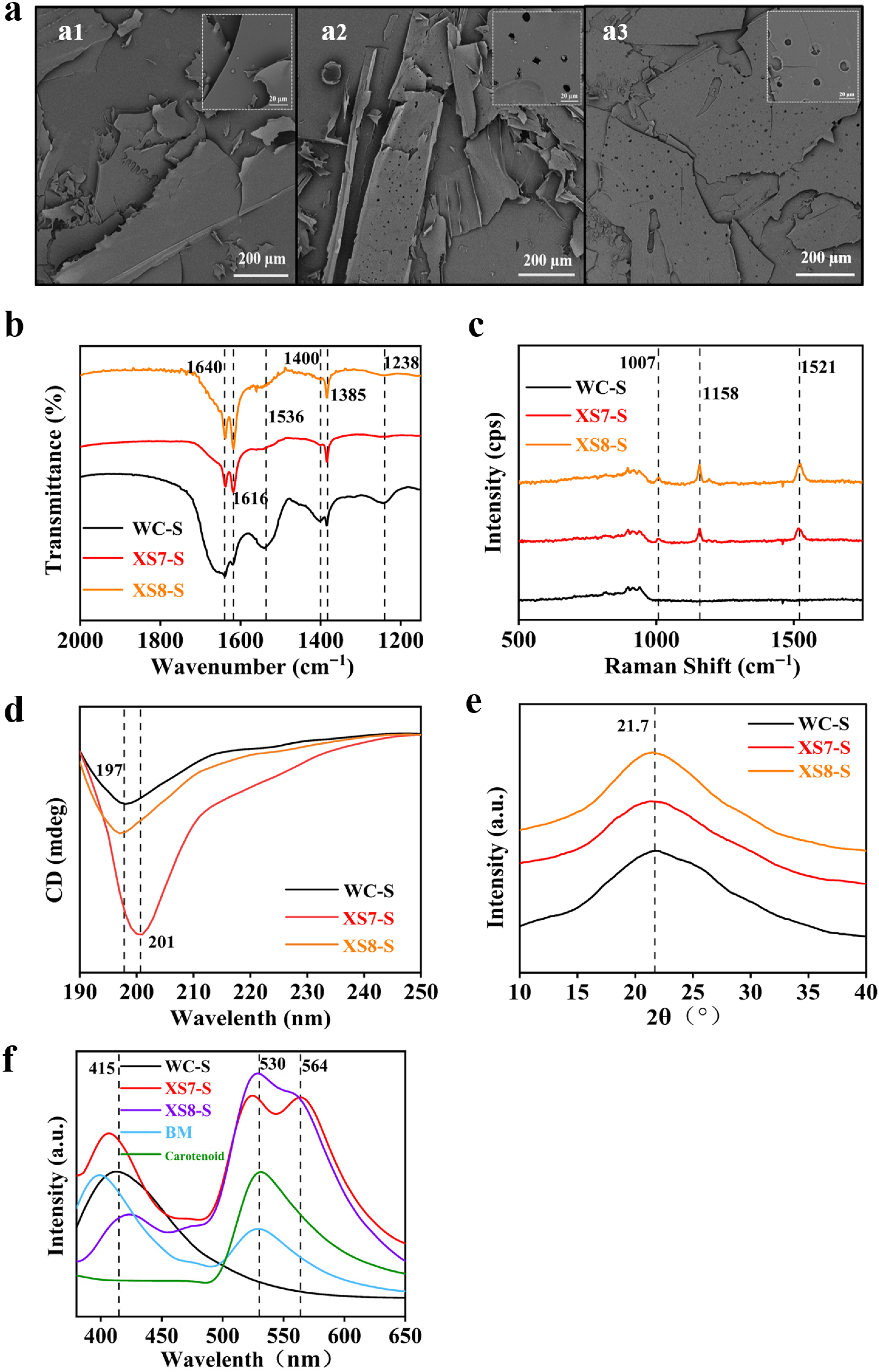
2.3. Comparison of Sericin from XS7, XS8 and Qiubai
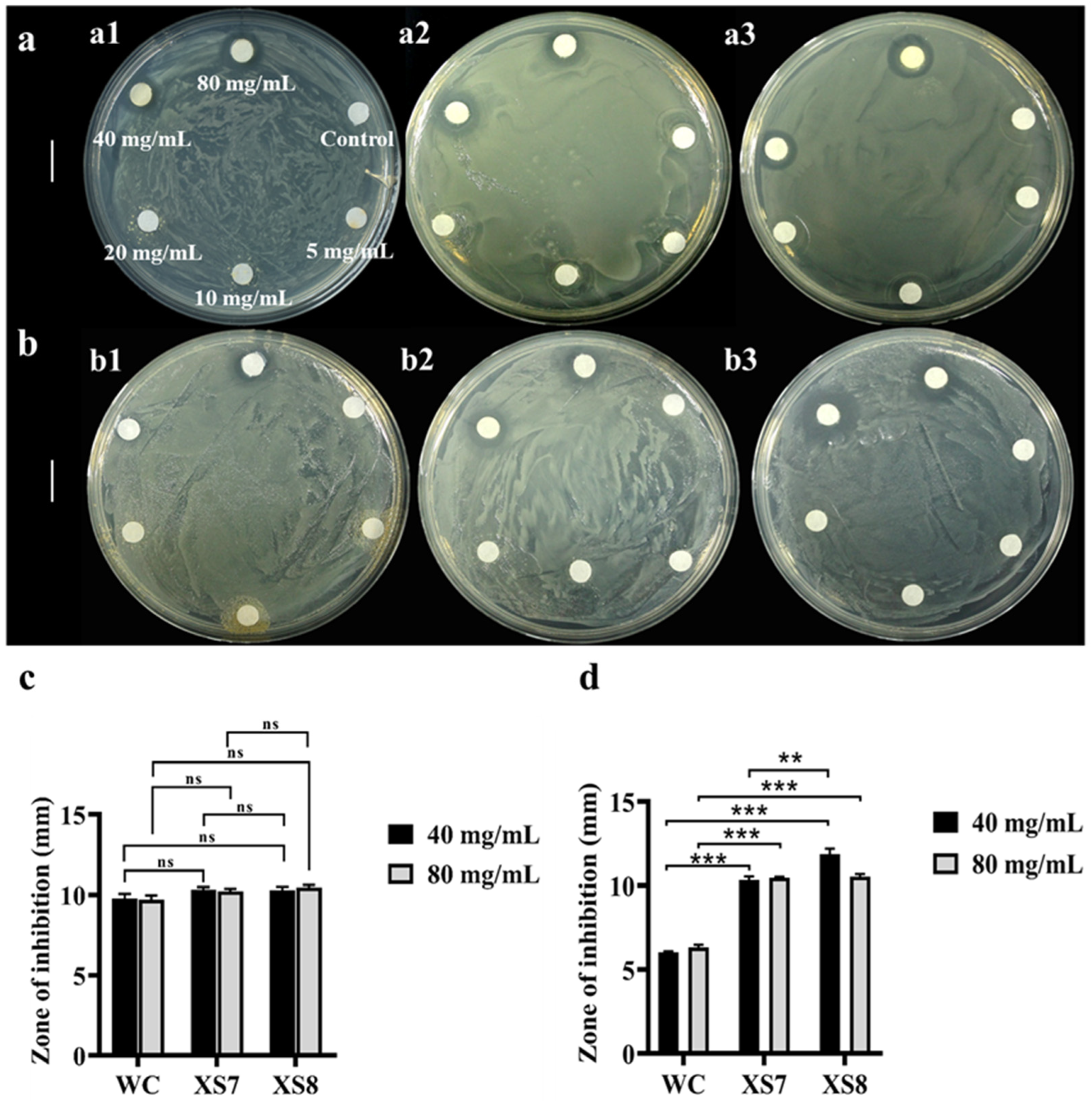
2.4. Comparison of Antimicrobial Effects of Sericin from XS7, XS8 and Qiubai
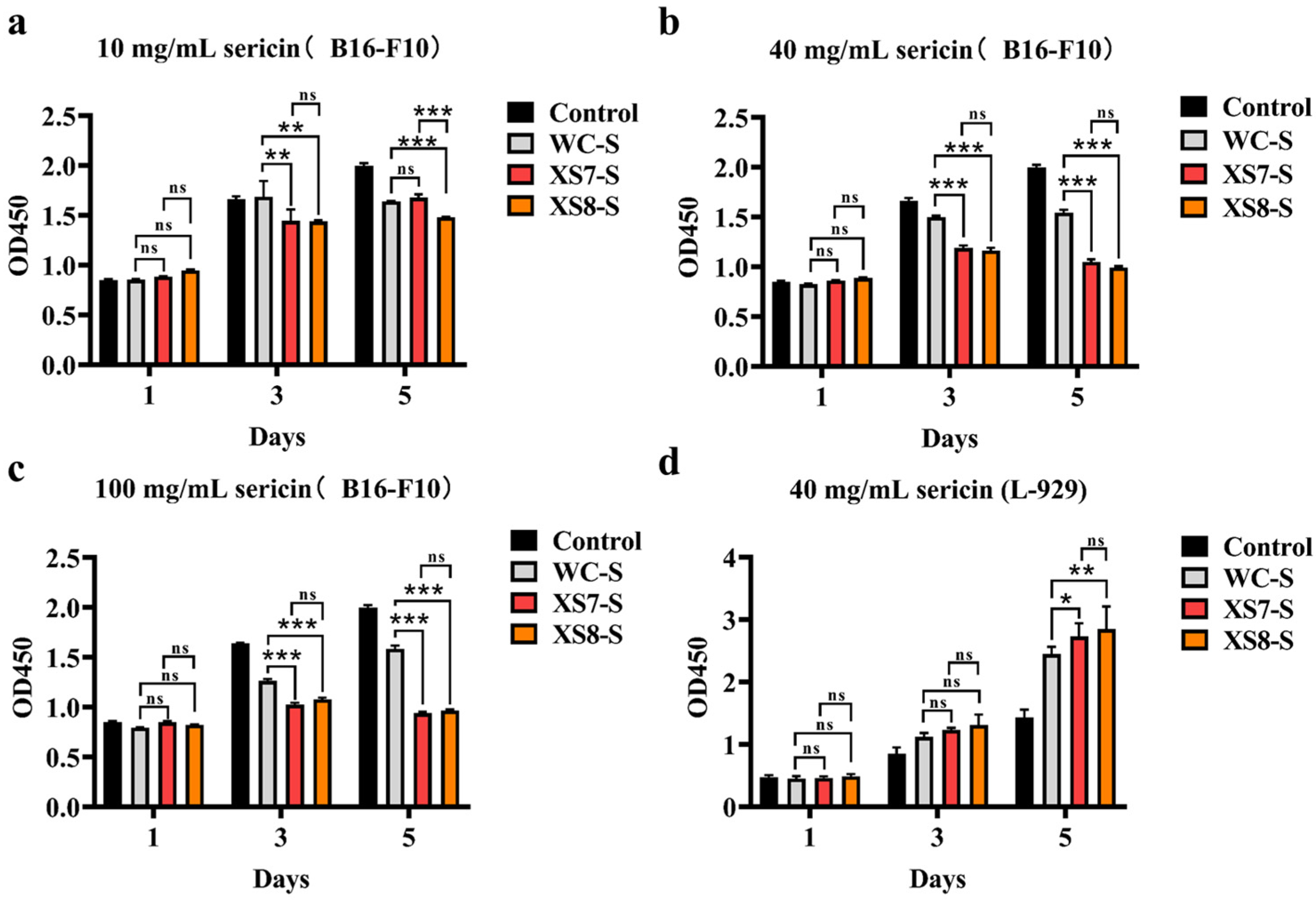
2.5. Anticancer Capability and Biocompatibility of Sericin from XS7, XS8, and Qiubai
2.6. Comparison of Antioxidant Properties of Sericin from XS7, XS8, and Qiubai
3. Discussion
4. Materials and Methods
4.1. Silkworms
4.2. Morphological and Molecular Comparison of the Silk Glands of XS7 and XS8 Silkworms
4.3. Carotenoid Extraction and Determination
4.4. Extraction of Sericin from Cocoons
4.5. Characterization of Sericin
4.6. Antimicrobial Effect Investigation
4.7. Anti-Tumor Activity and Cytocompatibility of Sericin
4.8. Antioxidant Effects of Sericin
4.9. Statistics Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ghosh, S.; Pati, F. Decellularized extracellular matrix and silk fibroin-based hybrid biomaterials: A comprehensive review on fabrication techniques and tissue-specific applications. Int. J. Biol. Macromol. 2023, 253, 127410. [Google Scholar] [CrossRef]
- Xing, T.; He, A.; Huang, Z.; Luo, Y.; Zhang, Y.; Wang, M.; Shi, Z.; Ke, G.; Bai, J.; Zhao, S.; et al. Silk-based flexible electronics and smart wearable Textiles: Progress and beyond. Chem. Eng. J. 2023, 474, 145534. [Google Scholar] [CrossRef]
- Asakura, T.; Williamson, M.P. A review on the structure of Bombyx mori silk fibroin fiber studied using solid-state NMR: An antipolar lamella with an 8-residue repeat. Int. J. Biol. Macromol. 2023, 245, 125537. [Google Scholar] [CrossRef]
- Saad, M.; El-Samad, L.M.; Gomaa, R.A.; Augustyniak, M.; Hassan, M.A. A comprehensive review of recent advances in silk sericin: Extraction approaches, structure, biochemical characterization, and biomedical applications. Int. J. Biol. Macromol. 2023, 250, 126067. [Google Scholar] [CrossRef]
- Yang, C.; Yao, L.; Zhang, L. Silk sericin-based biomaterials shine in food and pharmaceutical industries. Smart Mater. Med. 2023, 4, 447–459. [Google Scholar] [CrossRef]
- Hu, D.; Li, T.; Liang, W.; Wang, Y.; Feng, M.; Sun, J. Silk sericin as building blocks of bioactive materials for advanced therapeutics. J. Control. Release 2023, 353, 303–316. [Google Scholar] [CrossRef]
- Liu, J.; Shi, L.; Deng, Y.; Zou, M.; Cai, B.; Song, Y.; Wang, Z.; Wang, L. Silk sericin-based materials for biomedical applications. Biomaterials 2022, 287, 121638. [Google Scholar] [CrossRef]
- Kumar Dan, A.; Aamna, B.; De, S.; Pereira-Silva, M.; Sahu, R.; Cláudia Paiva-Santos, A.; Parida, S. Sericin nanoparticles: Future nanocarrier for target-specific delivery of chemotherapeutic drugs. J. Mol. Liq. 2022, 368, 120717. [Google Scholar] [CrossRef]
- Zhang, J.; Ding, Z.; Du, W.; Wang, X.; Guan, Y. Carotenoids act on coloration and increase immunity and antioxidant activity in the novel “Yongzhang Golden turtle” strain of Pelodiscus sinensis. Aquaculture 2023, 563, 738871. [Google Scholar] [CrossRef]
- Gebregziabher, B.S.; Gebremeskel, H.; Debesa, B.; Ayalneh, D.; Mitiku, T.; Wendwessen, T.; Habtemariam, E.; Nur, S.; Getachew, T. Carotenoids: Dietary sources, health functions, biofortification, marketing trend and affecting factors—A review. J. Agric. Food Res. 2023, 14, 100834. [Google Scholar] [CrossRef]
- Razz, S.A. Comprehensive overview of microalgae-derived carotenoids and their applications in diverse industries. Algal Res. 2024, 78, 103422. [Google Scholar] [CrossRef]
- Islam, F.; Khan, J.; Zehravi, M.; Das, R.; Haque, M.A.; Banu, A.; Parwaiz, S.; Nainu, F.; Nafady, M.H.; Shahriar, S.M.S.; et al. Synergistic effects of carotenoids: Therapeutic benefits on human health. Process Biochem. 2024, 136, 254–272. [Google Scholar] [CrossRef]
- Rehman, A.; Tong, Q.; Jafari, S.M.; Assadpour, E.; Shehzad, Q.; Aadil, R.M.; Iqbal, M.W.; Rashed, M.M.A.; Mushtaq, B.S.; Ashraf, W. Carotenoid-loaded nanocarriers: A comprehensive review. Adv. Colloid Interface Sci. 2020, 275, 102048. [Google Scholar] [CrossRef]
- Ma, M.; Hussain, M.; Dong, S.; Zhou, W. Characterization of the pigment in naturally yellow-colored domestic silk. Dye. Pigment. 2016, 124, 6–11. [Google Scholar] [CrossRef]
- Tsuchida, K.; Sakudoh, T. Recent progress in molecular genetic studies on the carotenoid transport system using cocoon-color mutants of the silkworm. Arch. Biochem. Biophys. 2015, 572, 151–157. [Google Scholar] [CrossRef]
- Vaishnav, S.R.; Singh, S.A. Chapter 14—Sericin, a by-product of the silk industry: Extraction and applications. In Value-Addition in Agri-Food Industry Waste Through Enzyme Technology; Academic Press: Cambridge, MA, USA, 2023; pp. 199–208. [Google Scholar]
- Sakudoh, T.; Kuwazaki, S.; Iizuka, T.; Narukawa, J.; Yamamoto, K.; Uchino, K.; Sezutsu, H.; Banno, Y.; Tsuchida, K. CD36 homolog divergence is responsible for the selectivity of carotenoid species migration to the silk gland of the silkworm Bombyx mori. J. Lipid Res. 2013, 54, 482–495. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, X.-X.; Zhang, R.; Feng, J.-Y.; Thakur, K.; Zhang, J.-G.; Wei, Z.-J. Anti-hardening effect and mechanism of silkworm sericin peptide in high protein nutrition bars during early storage. Food Chem. 2023, 407, 135168. [Google Scholar] [CrossRef]
- Bascou, R.; Hardouin, J.; Ben Mlouka, M.A.; Guénin, E.; Nesterenko, A. Detailed investigation on new chemical-free methods for silk sericin extraction. Mater. Today Commun. 2022, 33, 104491. [Google Scholar] [CrossRef]
- Zhang, L.; Hao, M.; Yao, L.; Xing, C.; Wen, Q.; Zhang, Z.; Yu, J.; Wang, J.; Xing, D.; Zheng, T.; et al. Sericin “hairpin structure”-based multifunctional anthocyanin nanoencapsulation for remodeling ROS-dependent cutaneous wound healing. Chem. Eng. J. 2023, 475, 145863. [Google Scholar] [CrossRef]
- Castrillón Martínez, D.C.; Zuluaga, C.L.; Restrepo-Osorio, A.; Álvarez-López, C. Characterization of sericin obtained from cocoons and silk yarns. Procedia Eng. 2017, 200, 377–383. [Google Scholar] [CrossRef]
- Rocha, L.K.H.; Favaro, L.I.L.; Rios, A.C.; Silva, E.C.; Silva, W.F.; Stigliani, T.P.; Guilger, M.; Lima, R.; Oliveira, J.M.; Aranha, N.; et al. Sericin from Bombyx mori cocoons. Part I: Extraction and physicochemical-biological characterization for biopharmaceutical applications. Process Biochem. 2017, 61, 163–177. [Google Scholar] [CrossRef]
- Portarena, S.; Anselmi, C.; Leonardi, L.; Proietti, S.; Bizzarri, A.R.; Brugnoli, E.; Baldacchini, C. Lutein/β-carotene ratio in extra virgin olive oil: An easy and rapid quantification method by Raman spectroscopy. Food Chem. 2023, 404, 134748. [Google Scholar] [CrossRef]
- Oliveira, V.E.D.; Castro, H.V.; Edwards, H.G.M.; Oliveira, L.F.C.D. Carotenes and carotenoids in natural biological samples: A Raman spectroscopic analysis. J. Raman Spectrosc. 2010, 41, 642–650. [Google Scholar] [CrossRef]
- Guo, K.; Zhang, X.; Zhao, D.; Qin, L.; Jiang, W.; Hu, W.; Liu, X.; Xia, Q.; Dong, Z.; Zhao, P. Identification and characterization of sericin5 reveals non-cocoon silk sericin components with high β-sheet content and adhesive strength. Acta Biomater. 2022, 150, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Bhagath Singh, G.V.P.; Subramaniam, K.V.L. Quantitative XRD study of amorphous phase in alkali activated low calcium siliceous fly ash. Constr. Build. Mater. 2016, 124, 139–147. [Google Scholar] [CrossRef]
- Shitole, M.; Dugam, S.; Tade, R.; Nangare, S. Pharmaceutical applications of silk sericin. Ann. Pharm. Fr. 2020, 78, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yang, W.; Pan, Q.; Zeng, Q.; Yan, C.; Bai, X.; Liu, Y.; Zhang, L.; Li, B. Effects of long-term blue light irradiation on carotenoid biosynthesis and antioxidant activities in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Food Res. Int. 2023, 174, 113661. [Google Scholar] [CrossRef] [PubMed]
- Khamhaengpol, A.; Siri, S. Composite Electrospun Scaffold Derived from Recombinant Fibroin of Weaver Ant (Oecophylla smaragdina) as Cell-Substratum. Appl. Biochem. Biotechnol. 2017, 183, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Von Lintig, J.; Moon, J.; Babino, D. Molecular components affecting ocular carotenoid and retinoid homeostasis. Prog. Retin. Eye Res. 2021, 80, 100864. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zhu, R.; Wu, D.; Su, H.; Liu, Z.; Chen, Z. How hydrogen bonding and π–π interactions synergistically facilitate mephedrone adsorption by bio-sorbent: An in-depth microscopic scale interpretation. Environ. Pollut. 2024, 342, 123044. [Google Scholar] [CrossRef]
- Song, K.; Wang, Y.; Dong, W.; Li, Z.; Xia, Q.; Zhu, P.; He, H. Decoding silkworm spinning programmed by pH and metal ions. Sci. Bull. 2023, 69, 792–802. [Google Scholar] [CrossRef]
- Mumtaz, S.; Ali, S.; Pervaiz, A.; Qureshi, M.Z.; Kanwal, K.; Saleem, T. Apoptotic and antiproliferative effects of silk protein sericin conjugated-AgNO3 nanoparticles in human breast cancer cells. Saudi J. Biol. Sci. 2023, 30, 103551. [Google Scholar] [CrossRef]
- Shalini, T.; Pavithraa, G.; Rakkesh, R.A.; Balakumar, S. Unravelling the nature-inspired silk sericin—Calcium phosphate hybrid nanocomposites: A promising sustainable biomaterial for hard tissue regeneration applications. Surf. Interfaces 2023, 42, 103396. [Google Scholar] [CrossRef]
- Scoville, A.G.; Barnett, L.L.; Bodbyl-Roels, S.; Kelly, J.K.; Hileman, L.C. Differential regulation of a MYB transcription factor is correlated with transgenerational epigenetic inheritance of trichome density in Mimulus guttatus. New Phytol. 2011, 191, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Menezes Silva, J.V.; Silva Santos, A.; Araujo Pereira, G.; Campos Chisté, R. Ultrasound-assisted extraction using ethanol efficiently extracted carotenoids from peels of peach palm fruits (Bactris gasipaes Kunth) without altering qualitative carotenoid profile. Heliyon 2023, 9, e14933. [Google Scholar] [CrossRef] [PubMed]
- Suo, A.; Fan, G.; Wu, C.; Li, T.; Cong, K. Green extraction of carotenoids from apricot flesh by ultrasound assisted corn oil extraction: Optimization, identification, and application. Food Chem. 2023, 420, 136096. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Luo, L.J.; Yang, C.J.; Lai, J.Y. Highly Retina-Permeating and Long-Acting Resveratrol/Metformin Nanotherapeutics for Enhanced Treatment of Macular Degeneration. ACS Nano 2022, 17, 168–183. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | β-Sheet (%) | Random Coil (%) | α-Helix (%) | β-Turn (%) |
|---|---|---|---|---|
| White cocoon sericin | 28.16 | 16.80 | 12.12 | 42.92 |
| XS7 sericin | 63.87 | 11.40 | 8.74 | 15.99 |
| XS8 sericin | 58.58 | 14.54 | 10.47 | 14.78 |
| Name | Sequence | Length (bp) |
|---|---|---|
| BmSCRB15-F | TGGAATACCACGGCAATAAGAC | 22 |
| BmSCRB15-R | ATGGGCAAACCATAAAAGCAAG | 22 |
| BmCameo2-F | ACAAGCACTCGTTCTATTTCGC | 22 |
| BmCameo2-R | TCATACAATGTGATGTGGTCGC | 22 |
| Bmrpl3-F | TTCGTACTGGCTCTTCTCGT | 20 |
| Bmrpl3-R | CAAAGTTGATAGCAATTCCCT | 21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Zhang, Q.; Zhou, H.; Zhang, B.; Yu, M.; Wang, Y.; Liu, Y.; Chai, C. The Potential of Natural Carotenoids-Containing Sericin of the Domestic Silkworm Bombyx mori. Int. J. Mol. Sci. 2024, 25, 3688. https://doi.org/10.3390/ijms25073688
Liu S, Zhang Q, Zhou H, Zhang B, Yu M, Wang Y, Liu Y, Chai C. The Potential of Natural Carotenoids-Containing Sericin of the Domestic Silkworm Bombyx mori. International Journal of Molecular Sciences. 2024; 25(7):3688. https://doi.org/10.3390/ijms25073688
Chicago/Turabian StyleLiu, Sirui, Qing Zhang, Hanyue Zhou, Bo Zhang, Ming Yu, Yonglong Wang, Yanqun Liu, and Chunli Chai. 2024. "The Potential of Natural Carotenoids-Containing Sericin of the Domestic Silkworm Bombyx mori" International Journal of Molecular Sciences 25, no. 7: 3688. https://doi.org/10.3390/ijms25073688
APA StyleLiu, S., Zhang, Q., Zhou, H., Zhang, B., Yu, M., Wang, Y., Liu, Y., & Chai, C. (2024). The Potential of Natural Carotenoids-Containing Sericin of the Domestic Silkworm Bombyx mori. International Journal of Molecular Sciences, 25(7), 3688. https://doi.org/10.3390/ijms25073688