
When the Embryo Meets the Endometrium: Identifying the Features Required for Successful Embryo Implantation

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
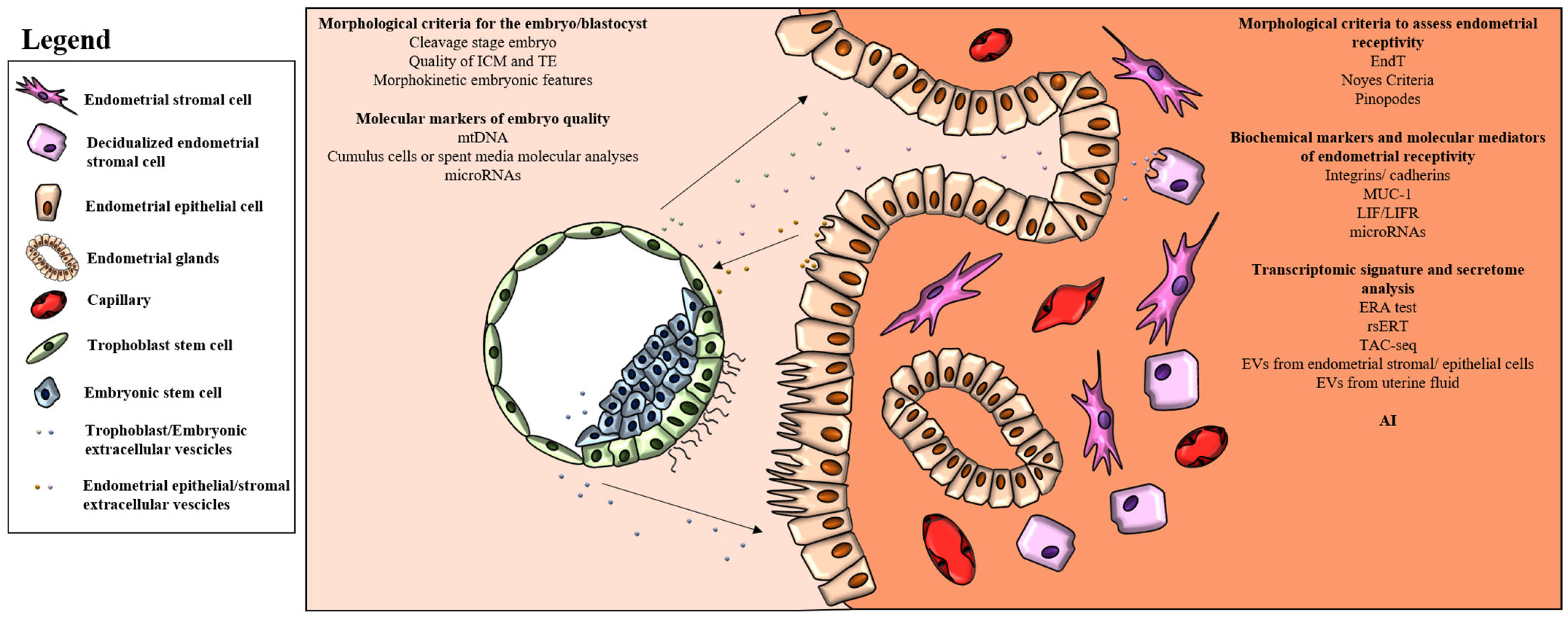
2. Morphological Criteria to Identify the Best Embryo
2.1. Static and Morphological Embryonic Features–Cleavage Stage Embryo
2.2. Static and Morphological Embryonic Features–Blastocyst
2.3. Morphokinetic Embryonic Features
3. Developmental Timing to Identify the Best Blastocyst
4. Molecular Markers of Embryo Quality
4.1. Mitochondrial DNA (mtDNA)
4.2. Cumulus Cells or Spent Media Molecular Analyses
5. Morphological Criteria to Assess Endometrial Receptivity
5.1. Endometrial Thickness (EndT)
5.2. Noyes Criteria
5.3. Pinopodes

{kind=link}
{kind=link}
| Embryo | Criteria | Description | Evidence in Support | Evidence Against |
| Good embryo according to Istanbul consensus | At least 8 blastomeres even sized on Day 3, <10% fragmentation and no signs of multinucleation | [7,33] | [34] | |
| Multinucleated blastomeres | A multinucleation in Day 2 and Day 3 cleavage embryos | [35,36,37,38,39] | ||
| ICM grading | The grading scale for ICM quality of the blastocyst | [52,53,54] | ||
| Expansion stage and TE grading | The grading scale for expansion and TE quality of the blastocyst | [55,56,57] | ||
| Developmental timing | Full blastocyst expansion should be assessed at 116 ± 2 h post-insemination | [58,64,65] | ||
| Endometrium | Endometrial Thickness (EndT) | Optimal thickness for receptive endometrium of about 16–18 mm (evaluated by transvaginal ultrasound) | [124,126,130,131,149] | [122,123,128,129,132] |
| Noyes Criteria | Histological criteria identifying gland mitosis and tortuosity, apical position of secretory vesicles in cells of the glandular epithelium, secreted material in the glandular lumen, stromal edema, pseudo-decidual reaction, and leukocyte infiltration | [136] | [18,22,23,133,134,135] | |
| Pinopodes | Evaluation of density and morphology of plasma membrane protrusions on epithelial cells projecting toward the uterine lumen on days 20–22 of a natural menstrual cycle | [137,138,140,142,143,145] | [146,147] |
6. Biochemical Markers and Molecular Mediators of Endometrial Receptivity
| Embryo | Markers | Local Expression | Samples of Study | Evidence in Support | Evidence Against |
| mitochondrial DNA | Embryo | [67,68] | [70,71,72,73] | ||
| CCND2, CXCR4, GPX3, CTNND1 DHCR7, DVL3, HSPB1, and TRIM28 | Cumulus cells | [77] | |||
| ANG, RGS2, and PLIN2 | Cumulus cells | [78] | |||
| PTGS2, CAMK1D, HAS2, STC1, and EFNB2 | Cumulus cells | [79,80,81,82] | |||
| CTGF, SERPINE2, PCK1, HHIP, HS3ST, and BIRC5 | Cumulus cells | [89] | |||
| miR-20a and miR-30c | Spent embryo culture media | [100] | [101] | ||
| miR-372 and miR-191 | Spent embryo culture media | [96] | |||
| miR-661 | Spent embryo culture media | [97] | |||
| miR-142-3p | Spent embryo culture media | [102] | |||
| Endometrium | αvβ3 | Luminal epithelium | Biopsies from women with unexplained infertility, endometriosis, PCOS | [158,167,168,169,170,171,172] | [173,174,175,176,177,178] |
| MUC-1 | Luminal and glandular epithelium | Biopsies from RIF, RPL, PCOS, endometriosis | [144,152,153,187,188] | ||
| LIF/LIFR | Luminal epithelium and blastocyst | Biopsies from women with unexplained infertility | [150,156,161,162] | ||
| let-7 family, miR-30d, miR-183-5p, miR-192, miR-23a-3p, miR-30a-3p, miR-145, and miR-200c | Epithelial cells | [191,192,193,195,198] | |||
| miR21, miR-96, miR-181a, miR-200 miR-148a, miR-181b, miR-194, and miR-542 | Stromal cells | [199,200,201,202,203,204,205,206] | |||
| miR-6821-5p, miR-483-5p 008, miR-4521, and miR-4421 | Uterine fluid in secretory phase | Biopsies from RIF | [207] | ||
| miR-96-5p, miR-186-5p, miR-628-3p, and miR-183-5p | Uterine fluid insecretory phase | Biopsies from healthy women | [207] |
7. Transcriptomic Signature and Secretome Analysis
8. Artificial Intelligence in ART
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ART | assisted reproduction technology |
| IVF | in vitro fertilization |
| PGT | preimplantation genetic testing |
| RIF | recurrent implantation failure |
| WOI | window of implantation |
| LH | luteinizing hormone |
| ERA | endometrial receptivity assay |
| ICM | inner cell mass |
| TE | trophectoderm |
| TLT | time-lapse technology |
| tPNf | time of pronuclear fading |
| tEB | time for expansion blastocyst |
| tM | time to morula |
| tB | time to blastocyst |
| mtDNA | mitochondrial DNA |
| CCs | cumulus cells |
| CCND2 | cyclin D2 |
| CXCR4 | CXC chemokine receptor 4 |
| GPX3 | glutathione peroxidase 3 |
| CTNND1 | catenin delta-1 |
| DHCR7 | 7-dehydrocholesterol reductase |
| DVL3 | disheveled dsh homologue 3 |
| HSPB1 | heatshock 27 kDa protein 1 |
| TRIM28 | tripartite motif-containing 28 |
| ANG | angiogenin |
| RGS2 | regulator of G-protein signaling 2 |
| PLIN2 | perilipin 2 |
| PTGS2 | Prostaglandin-endoperoxide synthase 2 |
| CAMK1D | calcium/calmodulin-dependent protein kinase 1D |
| HAS2 | hyaluronic acid synthase 2 |
| STC1 | stanniocalcin-1 |
| EFNB2 | ephrinB2 |
| NFIB | nuclear factor 1 b |
| BCL2L11 | BCL2 like 11 |
| PCK1 | phosphoenolpyruvate carboxykinase 1 |
| VCAN | versican |
| GREM1 | gremlin 1 |
| PFKP | phosphofructokinase, platelet |
| STC2 | stanniocalcin-2 |
| FGF12 | fibroblast growth factor 12 |
| GPR137B | G-protein–coupled receptor 13b |
| SLC2A9 | Solute carrier family 2 (facilitated glucose transporter), member 9 |
| ARID1B | AT-rich interactive domain 1B (SWI1-like) |
| NR2F6 | Nuclear receptor subfamily 2, group F, member 6 |
| ZNF132 | Zinc finger protein 132 |
| FAM36A | Family with sequence similarity 36, member A |
| ZNF93 | Zinc finger protein 93 |
| RHBDL2 | Rhomboid, veinlike 2 (Drosophila) |
| DNAJC15 | DnaJ (Hsp40) homologue, subfamily C, member 15 |
| MTUS1 | Microtubule-associated tumor suppressor 1 |
| NUP133 | Nucleoporin 133kDa |
| DEGs | uncover differentially expressed genes |
| CTGF | Connective tissue growth factor |
| HHIP | Hedgehog interacting protein |
| HS3ST | Heparan sulfate glucosamine 3-O-sulfotransferase 1 |
| BIRC5 | Baculoviral IAP Repeat-Containing 5 |
| SCM | spent embryo culture media |
| cfDNA | cell-free DNA |
| miRNA | microRNA |
| SET | single embryo transfers |
| EV | extracellular vesicles |
| ICSI | intracytoplasmic sperm injection |
| EndT | Endometrial Thickness |
| E2 | estrogen |
| P4 | progesterone |
| pFET | personalized frozen embryo transfer |
| MUC-1 | Mucin-1 |
| LIF1 | leukemia inhibitory factor |
| LIFR | LIF Receptor |
| EGF | epidermal growth factor |
| HB-EGF | heparin-binding EGF |
| PCOS | polycystic ovary syndrome |
| TNF-α | tumor necrosis factor α |
| RPL | recurrent pregnancy loss |
| rsERT | RNA-Seq-based endometrial receptivity test |
| TAC-seq | Targeted Allele Counting by sequencing |
| hESC | endometrial stromal cells |
| ROS | reactive oxygen species |
| AI | artificial intelligence |
| FSH | follicle-stimulating hormone |
References
- Macklon, N.S.; Geraedts, J.P.M.; Fauser, B.C.J.M. Conception to Ongoing Pregnancy: The “black Box” of Early Pregnancy Loss. Hum. Reprod. Update 2002, 8, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Maziotis, E.; Kalampokas, T.; Giannelou, P.; Grigoriadis, S.; Rapani, A.; Anifantakis, M.; Kotsifaki, A.; Pantou, A.; Triantafyllidou, O.; Tzanakaki, D.; et al. Commercially Available Molecular Approaches to Evaluate Endometrial Receptivity: A Systematic Review and Critical Analysis of the Literature. Diagnostics 2022, 12, 2611. [Google Scholar] [CrossRef]
- Galati, G.; Reschini, M.; Mensi, L.; Di Dio, C.; Somigliana, E.; Muzii, L. The Impact of Difficult Embryo Transfer on the Success of IVF: A Systematic Review and Meta-Analysis. Sci. Rep. 2023, 13, 22188. [Google Scholar] [CrossRef]
- Mani, S.; Mainigi, M. Embryo Culture Conditions and the Epigenome. Semin. Reprod. Med. 2018, 36, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Simopoulou, M.; Sfakianoudis, K.; Rapani, A.; Giannelou, P.; Anifandis, G.; Bolaris, S.; Pantou, A.; Lambropoulou, M.; Pappas, A.; Deligeoroglou, E.; et al. Considerations Regarding Embryo Culture Conditions: From Media to Epigenetics. In Vivo 2018, 32, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, K.; Agerholm, I.E.; Ingerslev, H.J. Time-Lapse Monitoring as a Tool for Clinical Embryo Assessment. Hum. Reprod. 2012, 27, 1277–1285. [Google Scholar] [CrossRef]
- Gardner, D.K.; Balaban, B. Assessment of Human Embryo Development Using Morphological Criteria in an Era of Time-Lapse, Algorithms and “OMICS”: Is Looking Good Still Important? Mol. Hum. Reprod. 2016, 22, 704–718. [Google Scholar] [CrossRef]
- Cimadomo, D.; Rienzi, L.; Capalbo, A.; Rubio, C.; Innocenti, F.; García-Pascual, C.M.; Ubaldi, F.M.; Handyside, A. The Dawn of the Future: 30 Years from the First Biopsy of a Human Embryo. The Detailed History of an Ongoing Revolution. Hum. Reprod. Update 2020, 26, 453–473. [Google Scholar] [CrossRef]
- Kontopoulos, G.; Simopoulou, M.; Zervomanolakis, I.; Prokopakis, T.; Dimitropoulos, K.; Dedoulis, E.; Grigorakis, S.; Agapitou, K.; Nikitos, E.; Rapani, A.; et al. Cleavage Stage versus Blastocyst Stage Embryo Transfer in Oocyte Donation Cycles. Medicina 2019, 55, 293. [Google Scholar] [CrossRef]
- Simopoulou, M.; Sfakianoudis, K.; Tsioulou, P.; Rapani, A.; Maziotis, E.; Giannelou, P.; Grigoriadis, S.; Pantou, A.; Nikolettos, K.; Vlahos, N.; et al. Should the Flexibility Enabled by Performing a Day-4 Embryo Transfer Remain as a Valid Option in the IVF Laboratory? A Systematic Review and Network Meta-Analysis. J. Assist. Reprod. Genet. 2019, 36, 1049–1061. [Google Scholar] [CrossRef]
- Li, Y.-X.; Wang, J.; Sun, T.-Z.; Lv, M.-Q.; Ge, P.; Li, H.-N.; Zhou, D.-X. Pregnancy Outcomes after Day 5 versus Day 6 Blastocyst-Stage Embryo Transfer: A Systematic Review and Meta-Analysis. J. Obstet. Gynaecol. Res. 2020, 46, 595–605. [Google Scholar] [CrossRef]
- Coughlan, C.; Ledger, W.; Wang, Q.; Liu, F.; Demirol, A.; Gurgan, T.; Cutting, R.; Ong, K.; Sallam, H.; Li, T.C. Recurrent Implantation Failure: Definition and Management. Reprod. Biomed. Online 2014, 28, 14–38. [Google Scholar] [CrossRef]
- Macklon, N.S.; Brosens, J.J. The Human Endometrium as a Sensor of Embryo Quality. Biol. Reprod. 2014, 91, 98. [Google Scholar] [CrossRef] [PubMed]
- Psychoyos, A. Uterine Receptivity for Nidation. Ann. N. Y. Acad. Sci. 1986, 476, 36–42. [Google Scholar] [CrossRef]
- Ma, W.; Song, H.; Das, S.K.; Paria, B.C.; Dey, S.K. Estrogen Is a Critical Determinant That Specifies the Duration of the Window of Uterine Receptivity for Implantation. Proc. Natl. Acad. Sci. USA 2003, 100, 2963–2968. [Google Scholar] [CrossRef] [PubMed]
- Navot, D.; Scott, R.T.; Droesch, K.; Veeck, L.L.; Liu, H.C.; Rosenwaks, Z. The Window of Embryo Transfer and the Efficiency of Human Conception In Vitro. Fertil. Steril. 1991, 55, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Donaghay, M.; Lessey, B.A. Uterine Receptivity: Alterations Associated with Benign Gynecological Disease. Semin. Reprod. Med. 2007, 25, 461–475. [Google Scholar] [CrossRef]
- Cohen, A.M.; Ye, X.Y.; Colgan, T.J.; Greenblatt, E.M.; Chan, C. Comparing Endometrial Receptivity Array to Histologic Dating of the Endometrium in Women with a History of Implantation Failure. Syst. Biol. Reprod. Med. 2020, 66, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Järvelä, I.Y.; Sladkevicius, P.; Kelly, S.; Ojha, K.; Campbell, S.; Nargund, G. Evaluation of Endometrial Receptivity during In-Vitro Fertilization Using Three-Dimensional Power Doppler Ultrasound. Ultrasound Obstet. Gynecol. 2005, 26, 765–769. [Google Scholar] [CrossRef]
- Bonilla-Musoles, F.; Raga, F.; Osborne, N.G.; Castillo, J.C.; Bonilla, F. Endometrial Receptivity: Evaluation with Ultrasound. Ultrasound Q. 2013, 29, 3–20. [Google Scholar] [CrossRef]
- Noyes, R.W.; Hertig, A.T.; Rock, J. Dating the Endometrial Biopsy. Am. J. Obst. Gynecol. 1975, 122, 262–263. [Google Scholar] [CrossRef]
- Smith, S.; Hosid, S.; Scott, L. Endometrial Biopsy Dating. Interobserver Variation and Its Impact on Clinical Practice. J. Reprod. Med. 1995, 40, 1–3. [Google Scholar]
- Murray, M.J.; Meyer, W.R.; Lessey, B.A.; Zaino, R.J.; Novotny, D.B.; Fritz, M.A. Endometrial Dating Revisited: A Randomized Systematic Study of Secretory Phase Histologic Characteristics in Normally Cycling Fertile Women. Fertil. Steril. 2002, 78, S67. [Google Scholar] [CrossRef]
- Díaz-Gimeno, P.; Horcajadas, J.A.; Martínez-Conejero, J.A.; Esteban, F.J.; Alamá, P.; Pellicer, A.; Simón, C. A Genomic Diagnostic Tool for Human Endometrial Receptivity Based on the Transcriptomic Signature. Fertil. Steril. 2011, 95, 50–60.e15. [Google Scholar] [CrossRef]
- Díaz-Gimeno, P.; Ruiz-Alonso, M.; Blesa, D.; Bosch, N.; Martínez-Conejero, J.A.; Alamá, P.; Garrido, N.; Pellicer, A.; Simón, C. The Accuracy and Reproducibility of the Endometrial Receptivity Array Is Superior to Histology as a Diagnostic Method for Endometrial Receptivity. Fertil. Steril. 2013, 99, 508–517. [Google Scholar] [CrossRef]
- Garrido-Gómez, T.; Ruiz-Alonso, M.; Blesa, D.; Diaz-Gimeno, P.; Vilella, F.; Simón, C. Profiling the Gene Signature of Endometrial Receptivity: Clinical Results. Fertil. Steril. 2013, 99, 1078–1085. [Google Scholar] [CrossRef]
- Ruiz-Alonso, M.; Blesa, D.; Díaz-Gimeno, P.; Gómez, E.; Fernández-Sánchez, M.; Carranza, F.; Carrera, J.; Vilella, F.; Pellicer, A.; Simón, C. The Endometrial Receptivity Array for Diagnosis and Personalized Embryo Transfer as a Treatment for Patients with Repeated Implantation Failure. Fertil. Steril. 2013, 100, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Alonso, M.; Galindo, N.; Pellicer, A.; Simon, C. What a Difference Two Days Make: “Personalized” Embryo Transfer (pET) Paradigm: A Case Report and Pilot Study. Hum. Reprod. 2014, 29, 1244–1247. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Koizumi, M.; Doshida, M.; Toya, M.; Sagara, E.; Oka, N.; Nakajo, Y.; Aono, N.; Igarashi, H.; Kyono, K. Efficacy of the Endometrial Receptivity Array for Repeated Implantation Failure in Japan: A Retrospective, Two-Centers Study. Reprod. Med. Biol. 2017, 16, 290–296. [Google Scholar] [CrossRef]
- Bassil, R.; Casper, R.; Samara, N.; Hsieh, T.-B.; Barzilay, E.; Orvieto, R.; Haas, J. Does the Endometrial Receptivity Array Really Provide Personalized Embryo Transfer? J. Assist. Reprod. Genet. 2018, 35, 1301–1305. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Kan, A.; Hitkari, J.; Taylor, B.; Tallon, N.; Warraich, G.; Yuzpe, A.; Nakhuda, G. The Role of the Endometrial Receptivity Array (ERA) in Patients Who Have Failed Euploid Embryo Transfers. J. Assist. Reprod. Genet. 2018, 35, 683–692. [Google Scholar] [CrossRef]
- Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology. The Istanbul Consensus Workshop on Embryo Assessment: Proceedings of an Expert Meeting. Hum. Reprod. 2011, 26, 1270–1283. [Google Scholar] [CrossRef]
- Racowsky, C.; Stern, J.E.; Gibbons, W.E.; Behr, B.; Pomeroy, K.O.; Biggers, J.D. National Collection of Embryo Morphology Data into Society for Assisted Reproductive Technology Clinic Outcomes Reporting System: Associations among Day 3 Cell Number, Fragmentation and Blastomere Asymmetry, and Live Birth Rate. Fertil. Steril. 2011, 95, 1985–1989. [Google Scholar] [CrossRef]
- Pons, M.C.; Carrasco, B.; Parriego, M.; Boada, M.; González-Foruria, I.; Garcia, S.; Coroleu, B.; Barri, P.N.; Veiga, A. Deconstructing the Myth of Poor Prognosis for Fast-Cleaving Embryos on Day 3. Is It Time to Change the Consensus? J. Assist. Reprod. Genet. 2019, 36, 2299–2305. [Google Scholar] [CrossRef]
- Van Royen, E.; Mangelschots, K.; Vercruyssen, M.; De Neubourg, D.; Valkenburg, M.; Ryckaert, G.; Gerris, J. Multinucleation in Cleavage Stage Embryos. Hum. Reprod. 2003, 18, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Meriano, J.; Clark, C.; Cadesky, K.; Laskin, C.A. Binucleated and Micronucleated Blastomeres in Embryos Derived from Human Assisted Reproduction Cycles. Reprod. Biomed. Online 2004, 9, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Yakin, K.; Balaban, B.; Urman, B. Impact of the Presence of One or More Multinucleated Blastomeres on the Developmental Potential of the Embryo to the Blastocyst Stage. Fertil. Steril. 2005, 83, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Ambroggio, J.; Gindoff, P.R.; Dayal, M.B.; Khaldi, R.; Peak, D.; Frankfurter, D.; Dubey, A.K. Multinucleation of a Sibling Blastomere on Day 2 Suggests Unsuitability for Embryo Transfer in IVF-Preimplantation Genetic Screening Cycles. Fertil. Steril. 2011, 96, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Balakier, H.; Sojecki, A.; Motamedi, G.; Librach, C. Impact of Multinucleated Blastomeres on Embryo Developmental Competence, Morphokinetics, and Aneuploidy. Fertil. Steril. 2016, 106, 608–614.e2. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Schoolcraft, W.B. Culture and Transfer of Human Blastocysts. Curr. Opin. Obstet. Gynecol. 1999, 11, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Schoolcraft, W.B.; Gardner, D.K.; Lane, M.; Schlenker, T.; Hamilton, F.; Meldrum, D.R. Blastocyst Culture and Transfer: Analysis of Results and Parameters Affecting Outcome in Two In Vitro Fertilization Programs. Fertil. Steril. 1999, 72, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Lane, M.; Stevens, J.; Schlenker, T.; Schoolcraft, W.B. Blastocyst Score Affects Implantation and Pregnancy Outcome: Towards a Single Blastocyst Transfer. Fertil. Steril. 2000, 73, 1155–1158. [Google Scholar] [CrossRef] [PubMed]
- Alfarawati, S.; Fragouli, E.; Colls, P.; Stevens, J.; Gutiérrez-Mateo, C.; Schoolcraft, W.B.; Katz-Jaffe, M.G.; Wells, D. The Relationship between Blastocyst Morphology, Chromosomal Abnormality, and Embryo Gender. Fertil. Steril. 2011, 95, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, A.; Rienzi, L.; Cimadomo, D.; Maggiulli, R.; Elliott, T.; Wright, G.; Nagy, Z.P.; Ubaldi, F.M. Correlation between Standard Blastocyst Morphology, Euploidy and Implantation: An Observational Study in Two Centers Involving 956 Screened Blastocysts. Hum. Reprod. 2014, 29, 1173–1181. [Google Scholar] [CrossRef]
- Fragouli, E.; Alfarawati, S.; Spath, K.; Wells, D. Morphological and Cytogenetic Assessment of Cleavage and Blastocyst Stage Embryos. Mol. Hum. Reprod. 2014, 20, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Minasi, M.G.; Colasante, A.; Riccio, T.; Ruberti, A.; Casciani, V.; Scarselli, F.; Spinella, F.; Fiorentino, F.; Varricchio, M.T.; Greco, E. Correlation between Aneuploidy, Standard Morphology Evaluation and Morphokinetic Development in 1730 Biopsied Blastocysts: A Consecutive Case Series Study. Hum. Reprod. 2016, 31, 2245–2254. [Google Scholar] [CrossRef] [PubMed]
- Barash, O.O.; Ivani, K.A.; Willman, S.P.; Rosenbluth, E.M.; Wachs, D.S.; Hinckley, M.D.; Pittenger Reid, S.; Weckstein, L.N. Association between Growth Dynamics, Morphological Parameters, the Chromosomal Status of the Blastocysts, and Clinical Outcomes in IVF PGS Cycles with Single Embryo Transfer. J. Assist. Reprod. Genet. 2017, 34, 1007–1016. [Google Scholar] [CrossRef]
- Guzman, L.; Nuñez, D.; López, R.; Inoue, N.; Portella, J.; Vizcarra, F.; Noriega-Portella, L.; Noriega-Hoces, L.; Munné, S. The Number of Biopsied Trophectoderm Cells May Affect Pregnancy Outcomes. J. Assist. Reprod. Genet. 2019, 36, 145–151. [Google Scholar] [CrossRef]
- Hernandez-Nieto, C.; Lee, J.A.; Duke, M.; Stein, D.E.; Copperman, A.B.; Sandler, B.; Mukherjee, T. Direct Correlation between Β-HCG Levels and Trophectoderm Morphology Quality in Single Euploid Embryo Transfer Cycles. Fertil. Steril. 2019, 112, e49. [Google Scholar] [CrossRef]
- Kim, M.K.; Park, J.K.; Jeon, Y.; Choe, S.-A.; Lee, H.J.; Kim, J.; Chang, E.M.; Kim, J.W.; Lyu, S.W.; Kim, J.Y.; et al. Correlation between Morphologic Grading and Euploidy Rates of Blastocysts, and Clinical Outcomes in In Vitro Fertilization Preimplantation Genetic Screening. J. Korean Med. Sci. 2019, 34, e27. [Google Scholar] [CrossRef]
- Viñals Gonzalez, X.; Odia, R.; Naja, R.; Serhal, P.; Saab, W.; Seshadri, S.; Ben-Nagi, J. Euploid Blastocysts Implant Irrespective of Their Morphology after NGS-(PGT-A) Testing in Advanced Maternal Age Patients. J. Assist. Reprod. Genet. 2019, 36, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.S.; Harris, D.C.; Daneshmand, S.T.; Shapiro, B.S. Quantitative Grading of a Human Blastocyst: Optimal Inner Cell Mass Size and Shape. Fertil. Steril. 2001, 76, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Boynukalin, F.K.; Gultomruk, M.; Cavkaytar, S.; Turgut, E.; Findikli, N.; Serdarogullari, M.; Coban, O.; Yarkiner, Z.; Rubio, C.; Bahceci, M. Parameters Impacting the Live Birth Rate per Transfer after Frozen Single Euploid Blastocyst Transfer. PLoS ONE 2020, 15, e0227619. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Jin, L.; Zheng, Y.; Yang, P.; Huang, B.; Dong, X. The Morphology of Inner Cell Mass Is the Strongest Predictor of Live Birth after a Frozen-Thawed Single Embryo Transfer. Front. Endocrinol. 2021, 12, 621221. [Google Scholar] [CrossRef] [PubMed]
- Ahlström, A.; Westin, C.; Reismer, E.; Wikland, M.; Hardarson, T. Trophectoderm Morphology: An Important Parameter for Predicting Live Birth after Single Blastocyst Transfer. Hum. Reprod. 2011, 26, 3289–3296. [Google Scholar] [CrossRef] [PubMed]
- Honnma, H.; Baba, T.; Sasaki, M.; Hashiba, Y.; Ohno, H.; Fukunaga, T.; Endo, T.; Saito, T.; Asada, Y. Trophectoderm Morphology Significantly Affects the Rates of Ongoing Pregnancy and Miscarriage in Frozen-Thawed Single-Blastocyst Transfer Cycle In Vitro Fertilization. Fertil. Steril. 2012, 98, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, J.; Wu, X.; Cao, S.; Zhou, L.; Wang, Y.; Chen, X.; Lu, J.; Zhao, C.; Chen, M.; et al. Trophectoderm Morphology Predicts Outcomes of Pregnancy in Vitrified-Warmed Single-Blastocyst Transfer Cycle in a Chinese Population. J. Assist. Reprod. Genet. 2014, 31, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Cimadomo, D.; Rienzi, L.; Conforti, A.; Forman, E.; Canosa, S.; Innocenti, F.; Poli, M.; Hynes, J.; Gemmell, L.; Vaiarelli, A.; et al. Opening the Black Box: Why Do Euploid Blastocysts Fail to Implant? A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2023, 29, 570–633. [Google Scholar] [CrossRef]
- Meseguer, M.; Herrero, J.; Tejera, A.; Hilligsøe, K.M.; Ramsing, N.B.; Remohí, J. The Use of Morphokinetics as a Predictor of Embryo Implantation. Hum. Reprod. 2011, 26, 2658–2671. [Google Scholar] [CrossRef]
- ESHRE Working Group on Time-Lapse Technology; Apter, S.; Ebner, T.; Freour, T.; Guns, Y.; Kovacic, B.; Le Clef, N.; Marques, M.; Meseguer, M.; Montjean, D.; et al. Good Practice Recommendations for the Use of Time-Lapse Technology. Hum. Reprod. Open 2020, 2020, hoaa008. [Google Scholar] [CrossRef]
- Rienzi, L.; Cimadomo, D.; Delgado, A.; Minasi, M.G.; Fabozzi, G.; Gallego, R.D.; Stoppa, M.; Bellver, J.; Giancani, A.; Esbert, M.; et al. Time of Morulation and Trophectoderm Quality Are Predictors of a Live Birth after Euploid Blastocyst Transfer: A Multicenter Study. Fertil. Steril. 2019, 112, 1080–1093.e1. [Google Scholar] [CrossRef]
- Bamford, T.; Barrie, A.; Montgomery, S.; Dhillon-Smith, R.; Campbell, A.; Easter, C.; Coomarasamy, A. Morphological and Morphokinetic Associations with Aneuploidy: A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2022, 28, 656–686. [Google Scholar] [CrossRef] [PubMed]
- McQueen, D.B.; Mazur, J.; Kimelman, D.; Confino, R.; Robins, J.C.; Bernardi, L.A.; Yeh, C.; Zhang, J.; Pavone, M.E. Can Embryo Morphokinetic Parameters Predict Euploid Pregnancy Loss? Fertil. Steril. 2021, 115, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, M.; Pocate-Cheriet, K.; Finet de Bantel, A.; Grzegorczyk-Martin, V.; Amar Hoffet, A.; Arbo, E.; Poulain, M.; Santulli, P. Day 5 versus Day 6 Blastocyst Transfers: A Systematic Review and Meta-Analysis of Clinical Outcomes. Hum. Reprod. 2019, 34, 1948–1964. [Google Scholar] [CrossRef] [PubMed]
- Corti, L.; Cermisoni, G.C.; Alteri, A.; Pagliardini, L.; Ambrosini, G.; Andrisani, A.; Papaleo, E.; Viganò, P.; Noventa, M. Clinical Outcomes Deriving from Transfer of Blastocysts Developed in Day 7: A Systematic Review and Meta-Analysis of Frozen-Thawed IVF Cycles. Reprod. Sci. 2022, 29, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Fragouli, E.; Wells, D. Mitochondrial DNA Assessment to Determine Oocyte and Embryo Viability. Semin. Reprod. Med. 2015, 33, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Fragouli, E.; Spath, K.; Alfarawati, S.; Kaper, F.; Craig, A.; Michel, C.-E.; Kokocinski, F.; Cohen, J.; Munne, S.; Wells, D. Altered Levels of Mitochondrial DNA Are Associated with Female Age, Aneuploidy, and Provide an Independent Measure of Embryonic Implantation Potential. PLoS Genet. 2015, 11, e1005241. [Google Scholar] [CrossRef] [PubMed]
- Diez-Juan, A.; Rubio, C.; Marin, C.; Martinez, S.; Al-Asmar, N.; Riboldi, M.; Díaz-Gimeno, P.; Valbuena, D.; Simón, C. Mitochondrial DNA Content as a Viability Score in Human Euploid Embryos: Less Is Better. Fertil. Steril. 2015, 104, 534–541.e1. [Google Scholar] [CrossRef]
- Podolak, A.; Woclawek-Potocka, I.; Lukaszuk, K. The Role of Mitochondria in Human Fertility and Early Embryo Development: What Can We Learn for Clinical Application of Assessing and Improving Mitochondrial DNA? Cells 2022, 11, 797. [Google Scholar] [CrossRef]
- Victor, A.R.; Brake, A.J.; Tyndall, J.C.; Griffin, D.K.; Zouves, C.G.; Barnes, F.L.; Viotti, M. Accurate Quantitation of Mitochondrial DNA Reveals Uniform Levels in Human Blastocysts Irrespective of Ploidy, Age, or Implantation Potential. Fertil. Steril. 2017, 107, 34–42.e3. [Google Scholar] [CrossRef]
- Treff, N.R.; Zhan, Y.; Tao, X.; Olcha, M.; Han, M.; Rajchel, J.; Morrison, L.; Morin, S.J.; Scott, R.T. Levels of Trophectoderm Mitochondrial DNA Do Not Predict the Reproductive Potential of Sibling Embryos. Hum. Reprod. 2017, 32, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Podolak, A.; Liss, J.; Kiewisz, J.; Pukszta, S.; Cybulska, C.; Rychlowski, M.; Lukaszuk, A.; Jakiel, G.; Lukaszuk, K. Mitochondrial DNA Copy Number in Cleavage Stage Human Embryos-Impact on Infertility Outcome. Curr. Issues Mol. Biol. 2022, 44, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Lukaszuk, K.; Podolak, A. Does Trophectoderm Mitochondrial DNA Content Affect Embryo Developmental and Implantation Potential? Int. J. Mol. Sci. 2022, 23, 5976. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.T.; Sun, L.; Zhan, Y.; Marin, D.; Tao, X.; Seli, E. Mitochondrial DNA Content Is Not Predictive of Reproductive Competence in Euploid Blastocysts. Reprod. Biomed. Online 2020, 41, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Robert, C. Nurturing the Egg: The Essential Connection between Cumulus Cells and the Oocyte. Reprod. Fertil. Dev. 2021, 34, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Richani, D.; Dunning, K.R.; Thompson, J.G.; Gilchrist, R.B. Metabolic Co-Dependence of the Oocyte and Cumulus Cells: Essential Role in Determining Oocyte Developmental Competence. Hum. Reprod. Update 2021, 27, 27–47. [Google Scholar] [CrossRef] [PubMed]
- van Montfoort, A.P.A.; Geraedts, J.P.M.; Dumoulin, J.C.M.; Stassen, A.P.M.; Evers, J.L.H.; Ayoubi, T.A.Y. Differential Gene Expression in Cumulus Cells as a Prognostic Indicator of Embryo Viability: A Microarray Analysis. Mol. Hum. Reprod. 2008, 14, 157–168. [Google Scholar] [CrossRef]
- Feuerstein, P.; Puard, V.; Chevalier, C.; Teusan, R.; Cadoret, V.; Guerif, F.; Houlgatte, R.; Royere, D. Genomic Assessment of Human Cumulus Cell Marker Genes as Predictors of Oocyte Developmental Competence: Impact of Various Experimental Factors. PLoS ONE 2012, 7, e40449. [Google Scholar] [CrossRef]
- Scarica, C.; Cimadomo, D.; Dovere, L.; Giancani, A.; Stoppa, M.; Capalbo, A.; Ubaldi, F.M.; Rienzi, L.; Canipari, R. An Integrated Investigation of Oocyte Developmental Competence: Expression of Key Genes in Human Cumulus Cells, Morphokinetics of Early Divisions, Blastulation, and Euploidy. J. Assist. Reprod. Genet. 2019, 36, 875–887. [Google Scholar] [CrossRef]
- McKenzie, L.J.; Pangas, S.A.; Carson, S.A.; Kovanci, E.; Cisneros, P.; Buster, J.E.; Amato, P.; Matzuk, M.M. Human Cumulus Granulosa Cell Gene Expression: A Predictor of Fertilization and Embryo Selection in Women Undergoing IVF. Hum. Reprod. 2004, 19, 2869–2874. [Google Scholar] [CrossRef]
- Cillo, F.; Brevini, T.A.L.; Antonini, S.; Paffoni, A.; Ragni, G.; Gandolfi, F. Association between Human Oocyte Developmental Competence and Expression Levels of Some Cumulus Genes. Reproduction 2007, 134, 645–650. [Google Scholar] [CrossRef]
- Wathlet, S.; Adriaenssens, T.; Segers, I.; Verheyen, G.; Janssens, R.; Coucke, W.; Devroey, P.; Smitz, J. New Candidate Genes to Predict Pregnancy Outcome in Single Embryo Transfer Cycles When Using Cumulus Cell Gene Expression. Fertil. Steril. 2012, 98, 432–439.e4. [Google Scholar] [CrossRef] [PubMed]
- Assou, S.; Haouzi, D.; Mahmoud, K.; Aouacheria, A.; Guillemin, Y.; Pantesco, V.; Rème, T.; Dechaud, H.; De Vos, J.; Hamamah, S. A Non-Invasive Test for Assessing Embryo Potential by Gene Expression Profiles of Human Cumulus Cells: A Proof of Concept Study. Mol. Hum. Reprod. 2008, 14, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, K.M.; Feil, D.K.; Dunning, K.R.; Lane, M.; Russell, D.L. Human Cumulus Cell Gene Expression as a Biomarker of Pregnancy Outcome after Single Embryo Transfer. Fertil. Steril. 2011, 96, 47–52.e2. [Google Scholar] [CrossRef] [PubMed]
- Iager, A.E.; Kocabas, A.M.; Otu, H.H.; Ruppel, P.; Langerveld, A.; Schnarr, P.; Suarez, M.; Jarrett, J.C.; Conaghan, J.; Rosa, G.J.M.; et al. Identification of a Novel Gene Set in Human Cumulus Cells Predictive of an Oocyte’s Pregnancy Potential. Fertil. Steril. 2013, 99, 745–752.e6. [Google Scholar] [CrossRef] [PubMed]
- Parks, J.C.; Patton, A.L.; McCallie, B.R.; Griffin, D.K.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Corona Cell RNA Sequencing from Individual Oocytes Revealed Transcripts and Pathways Linked to Euploid Oocyte Competence and Live Birth. Reprod. Biomed. Online 2016, 32, 518–526. [Google Scholar] [CrossRef]
- Green, K.A.; Franasiak, J.M.; Werner, M.D.; Tao, X.; Landis, J.N.; Scott, R.T.; Treff, N.R. Cumulus Cell Transcriptome Profiling Is Not Predictive of Live Birth after In Vitro Fertilization: A Paired Analysis of Euploid Sibling Blastocysts. Fertil. Steril. 2018, 109, 460–466.e2. [Google Scholar] [CrossRef]
- Papler, T.B.; Bokal, E.V.; Maver, A.; Kopitar, A.N.; Lovrečić, L. Transcriptomic Analysis and Meta-Analysis of Human Granulosa and Cumulus Cells. PLoS ONE 2015, 10, e0136473. [Google Scholar] [CrossRef]
- Sachs, M.K.; Makieva, S.; Velasco Gil, A.; Xie, M.; Ille, F.; Salvadori, V.; Schmidhauser, M.; Saenz-de-Juano, M.D.; Ulbrich, S.E.; Leeners, B. Transcriptomic Signature of Luteinized Cumulus Cells of Oocytes Developing to Live Birth after Women Received Intracytoplasmic Sperm Injection. F&S Sci. 2024, 5, 24–38. [Google Scholar] [CrossRef]
- Katz-Jaffe, M.G.; McReynolds, S.; Gardner, D.K.; Schoolcraft, W.B. The Role of Proteomics in Defining the Human Embryonic Secretome. Mol. Hum. Reprod. 2009, 15, 271–277. [Google Scholar] [CrossRef]
- Capalbo, A.; Romanelli, V.; Patassini, C.; Poli, M.; Girardi, L.; Giancani, A.; Stoppa, M.; Cimadomo, D.; Ubaldi, F.M.; Rienzi, L. Diagnostic Efficacy of Blastocoel Fluid and Spent Media as Sources of DNA for Preimplantation Genetic Testing in Standard Clinical Conditions. Fertil. Steril. 2018, 110, 870–879.e5. [Google Scholar] [CrossRef]
- Ho, J.R.; Arrach, N.; Rhodes-Long, K.; Ahmady, A.; Ingles, S.; Chung, K.; Bendikson, K.A.; Paulson, R.J.; McGinnis, L.K. Pushing the Limits of Detection: Investigation of Cell-Free DNA for Aneuploidy Screening in Embryos. Fertil. Steril. 2018, 110, 467–475.e2. [Google Scholar] [CrossRef] [PubMed]
- Kuznyetsov, V.; Madjunkova, S.; Antes, R.; Abramov, R.; Motamedi, G.; Ibarrientos, Z.; Librach, C. Evaluation of a Novel Non-Invasive Preimplantation Genetic Screening Approach. PLoS ONE 2018, 13, e0197262. [Google Scholar] [CrossRef] [PubMed]
- Vera-Rodriguez, M.; Diez-Juan, A.; Jimenez-Almazan, J.; Martinez, S.; Navarro, R.; Peinado, V.; Mercader, A.; Meseguer, M.; Blesa, D.; Moreno, I.; et al. Origin and Composition of Cell-Free DNA in Spent Medium from Human Embryo Culture during Preimplantation Development. Hum. Reprod. 2018, 33, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Belandres, D.; Shamonki, M.; Arrach, N. Current Status of Spent Embryo Media Research for Preimplantation Genetic Testing. J. Assist. Reprod. Genet. 2019, 36, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Rosenbluth, E.M.; Shelton, D.N.; Wells, L.M.; Sparks, A.E.T.; Van Voorhis, B.J. Human Embryos Secrete microRNAs into Culture Media—A Potential Biomarker for Implantation. Fertil. Steril. 2014, 101, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Cuman, C.; Van Sinderen, M.; Gantier, M.P.; Rainczuk, K.; Sorby, K.; Rombauts, L.; Osianlis, T.; Dimitriadis, E. Human Blastocyst Secreted microRNA Regulate Endometrial Epithelial Cell Adhesion. EBioMedicine 2015, 2, 1528–1535. [Google Scholar] [CrossRef] [PubMed]
- Kropp, J.; Khatib, H. Characterization of microRNA in Bovine In Vitro Culture Media Associated with Embryo Quality and Development. J. Dairy Sci. 2015, 98, 6552–6563. [Google Scholar] [CrossRef]
- Mutia, K.; Wiweko, B.; Abinawanto, A.; Dwiranti, A.; Bowolaksono, A. microRNAs as A Biomarker to Predict Embryo Quality Assessment in In Vitro Fertilization. Int. J. Fertil. Steril. 2023, 17, 85–91. [Google Scholar] [CrossRef]
- Capalbo, A.; Ubaldi, F.M.; Cimadomo, D.; Noli, L.; Khalaf, Y.; Farcomeni, A.; Ilic, D.; Rienzi, L. MicroRNAs in Spent Blastocyst Culture Medium Are Derived from Trophectoderm Cells and Can Be Explored for Human Embryo Reproductive Competence Assessment. Fertil. Steril. 2016, 105, 225–235.e3. [Google Scholar] [CrossRef]
- Cimadomo, D.; Rienzi, L.; Giancani, A.; Alviggi, E.; Dusi, L.; Canipari, R.; Noli, L.; Ilic, D.; Khalaf, Y.; Ubaldi, F.M.; et al. Definition and Validation of a Custom Protocol to Detect miRNAs in the Spent Media after Blastocyst Culture: Searching for Biomarkers of Implantation. Hum. Reprod. 2019, 34, 1746–1761. [Google Scholar] [CrossRef]
- Borges, E., Jr.; Setti, A.S.; Braga, D.P.A.F.; Geraldo, M.V.; de Cássia S Figueira, R.; Iaconelli, A., Jr. miR-142-3p as a Biomarker of Blastocyst Implantation Failure—A Pilot Study. JBRA Assist. Reprod. 2016, 20, 200–205. [Google Scholar] [CrossRef]
- Leaver, M.; Wells, D. Non-Invasive Preimplantation Genetic Testing (niPGT): The next Revolution in Reproductive Genetics? Hum. Reprod. Update 2020, 26, 16–42. [Google Scholar] [CrossRef]
- Rubio, C.; Rienzi, L.; Navarro-Sánchez, L.; Cimadomo, D.; García-Pascual, C.M.; Albricci, L.; Soscia, D.; Valbuena, D.; Capalbo, A.; Ubaldi, F.; et al. Embryonic Cell-Free DNA versus Trophectoderm Biopsy for Aneuploidy Testing: Concordance Rate and Clinical Implications. Fertil. Steril. 2019, 112, 510–519. [Google Scholar] [CrossRef]
- Yeung, Q.S.Y.; Zhang, Y.X.; Chung, J.P.W.; Lui, W.T.; Kwok, Y.K.Y.; Gui, B.; Kong, G.W.S.; Cao, Y.; Li, T.C.; Choy, K.W. A Prospective Study of Non-Invasive Preimplantation Genetic Testing for Aneuploidies (NiPGT-A) Using next-Generation Sequencing (NGS) on Spent Culture Media (SCM). J. Assist. Reprod. Genet. 2019, 36, 1609–1621. [Google Scholar] [CrossRef]
- Magli, M.C.; Albanese, C.; Crippa, A.; Tabanelli, C.; Ferraretti, A.P.; Gianaroli, L. Deoxyribonucleic Acid Detection in Blastocoelic Fluid: A New Predictor of Embryo Ploidy and Viable Pregnancy. Fertil. Steril. 2019, 111, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Pavani, K.C.; Hendrix, A.; Van Den Broeck, W.; Couck, L.; Szymanska, K.; Lin, X.; De Koster, J.; Van Soom, A.; Leemans, B. Isolation and Characterization of Functionally Active Extracellular Vesicles from Culture Medium Conditioned by Bovine Embryos In Vitro. Int. J. Mol. Sci. 2018, 20, 38. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, E.; Vago, R.; Sanchez, A.M.; Podini, P.; Zarovni, N.; Murdica, V.; Rizzo, R.; Bortolotti, D.; Candiani, M.; Viganò, P. Secretome of In Vitro Cultured Human Embryos Contains Extracellular Vesicles That Are Uptaken by the Maternal Side. Sci. Rep. 2017, 7, 5210. [Google Scholar] [CrossRef] [PubMed]
- Saadeldin, I.M.; Kim, S.J.; Choi, Y.B.; Lee, B.C. Improvement of Cloned Embryos Development by Co-Culturing with Parthenotes: A Possible Role of Exosomes/Microvesicles for Embryos Paracrine Communication. Cell. Reprogram. 2014, 16, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Pavani, K.C.; Alminana, C.; Wydooghe, E.; Catteeuw, M.; Ramírez, M.A.; Mermillod, P.; Rizos, D.; Van Soom, A. Emerging Role of Extracellular Vesicles in Communication of Preimplantation Embryos In Vitro. Reprod. Fertil. Dev. 2016, 29, 66–83. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef] [PubMed]
- Vlaeminck-Guillem, V. Extracellular Vesicles in Prostate Cancer Carcinogenesis, Diagnosis, and Management. Front. Oncol. 2018, 8, 222. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Beckers, E.; Mc Cafferty, S.; Gansemans, Y.; Joanna Szymańska, K.; Chaitanya Pavani, K.; Catani, J.P.; Van Nieuwerburgh, F.; Deforce, D.; De Sutter, P.; et al. Bovine Embryo-Secreted microRNA-30c Is a Potential Non-Invasive Biomarker for Hampered Preimplantation Developmental Competence. Front. Genet. 2019, 10, 315. [Google Scholar] [CrossRef] [PubMed]
- Pavani, K.C.; Meese, T.; Pascottini, O.B.; Guan, X.; Lin, X.; Peelman, L.; Hamacher, J.; Van Nieuwerburgh, F.; Deforce, D.; Boel, A.; et al. Hatching Is Modulated by microRNA-378a-3p Derived from Extracellular Vesicles Secreted by Blastocysts. Proc. Natl. Acad. Sci. USA 2022, 119, e2122708119. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, K.; Nõmm, M.; Lättekivi, F.; Ord, J.; Ressaissi, Y.; Godakumara, K.; Reshi, Q.U.A.; Viil, J.; Jääger, K.; Velthut-Meikas, A.; et al. Oviduct as a Sensor of Embryo Quality: Deciphering the Extracellular Vesicle (EV)-Mediated Embryo-Maternal Dialogue. J. Mol. Med. 2021, 99, 685–697. [Google Scholar] [CrossRef]
- Godakumara, K.; Ord, J.; Lättekivi, F.; Dissanayake, K.; Viil, J.; Boggavarapu, N.R.; Faridani, O.R.; Jääger, K.; Velthut-Meikas, A.; Jaakma, Ü.; et al. Trophoblast Derived Extracellular Vesicles Specifically Alter the Transcriptome of Endometrial Cells and May Constitute a Critical Component of Embryo-Maternal Communication. Reprod. Biol. Endocrinol. 2021, 19, 115. [Google Scholar] [CrossRef]
- Es-Haghi, M.; Godakumara, K.; Häling, A.; Lättekivi, F.; Lavrits, A.; Viil, J.; Andronowska, A.; Nafee, T.; James, V.; Jaakma, Ü.; et al. Specific Trophoblast Transcripts Transferred by Extracellular Vesicles Affect Gene Expression in Endometrial Epithelial Cells and May Have a Role in Embryo-Maternal Crosstalk. Cell Commun. Signal. 2019, 17, 146. [Google Scholar] [CrossRef]
- Ochoa-Bernal, M.A.; Fazleabas, A.T. Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human Primates. Int. J. Mol. Sci. 2020, 21, 1973. [Google Scholar] [CrossRef]
- Mazur, E.C.; Large, M.J.; DeMayo, F.J. Chapter 24—Human Oviduct and Endometrium: Changes over the Menstrual Cycle. In Knobil and Neill’s Physiology of Reproduction, 4th ed.; Plant, T.M., Zeleznik, A.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 1077–1097. ISBN 978-0-12-397175-3. [Google Scholar]
- Giri, S.K.; Nayak, B.L.; Mohapatra, J. Thickened Endometrium: When to Intervene? A Clinical Conundrum. J. Obstet. Gynaecol. India 2021, 71, 216–225. [Google Scholar] [CrossRef] [PubMed]
- De Geyter, C.; Schmitter, M.; De Geyter, M.; Nieschlag, E.; Holzgreve, W.; Schneider, H.P. Prospective Evaluation of the Ultrasound Appearance of the Endometrium in a Cohort of 1186 Infertile Women. Fertil. Steril. 2000, 73, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Schild, R.L.; Knobloch, C.; Dorn, C.; Fimmers, R.; van der Ven, H.; Hansmann, M. Endometrial Receptivity in an In Vitro Fertilization Program as Assessed by Spiral Artery Blood Flow, Endometrial Thickness, Endometrial Volume, and Uterine Artery Blood Flow. Fertil. Steril. 2001, 75, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Ng, E.H.Y.; Chan, C.C.W.; Tang, O.S.; Yeung, W.S.B.; Ho, P.C. The Role of Endometrial and Subendometrial Blood Flows Measured by Three-Dimensional Power Doppler Ultrasound in the Prediction of Pregnancy during IVF Treatment. Hum. Reprod. 2006, 21, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-L.; Wu, F.-R.; Luo, C.; Chen, X.; Shi, X.-Y.; Zheng, H.-Y.; Ni, Y.-P. Combined Analysis of Endometrial Thickness and Pattern in Predicting Outcome of In Vitro Fertilization and Embryo Transfer: A Retrospective Cohort Study. Reprod. Biol. Endocrinol. 2010, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Qiao, J.; Li, R.; Zhen, X.; Liu, Z. Role of Endometrial Blood Flow Assessment with Color Doppler Energy in Predicting Pregnancy Outcome of IVF-ET Cycles. Reprod. Biol. Endocrinol. 2010, 8, 122. [Google Scholar] [CrossRef] [PubMed]
- Kuć, P.; Kuczyńska, A.; Topczewska, M.; Tadejko, P.; Kuczyński, W. The Dynamics of Endometrial Growth and the Triple Layer Appearance in Three Different Controlled Ovarian Hyperstimulation Protocols and Their Influence on IVF Outcomes. Gynecol. Endocrinol. 2011, 27, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, Q.; Li, Y. The Effect of Endometrial Thickness and Pattern Measured by Ultrasonography on Pregnancy Outcomes during IVF-ET Cycles. Reprod. Biol. Endocrinol. 2012, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Kasius, A.; Smit, J.G.; Torrance, H.L.; Eijkemans, M.J.C.; Mol, B.W.; Opmeer, B.C.; Broekmans, F.J.M. Endometrial Thickness and Pregnancy Rates after IVF: A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2014, 20, 530–541. [Google Scholar] [CrossRef]
- Griesinger, G.; Trevisan, S.; Cometti, B. Endometrial Thickness on the Day of Embryo Transfer Is a Poor Predictor of IVF Treatment Outcome. Hum. Reprod. Open 2018, 2018, hox031. [Google Scholar] [CrossRef]
- Liu, K.E.; Hartman, M.; Hartman, A.; Luo, Z.-C.; Mahutte, N. The Impact of a Thin Endometrial Lining on Fresh and Frozen-Thaw IVF Outcomes: An Analysis of over 40,000 Embryo Transfers. Hum. Reprod. 2018, 33, 1883–1888. [Google Scholar] [CrossRef]
- Liao, S.; Wang, R.; Hu, C.; Pan, W.; Pan, W.; Yu, D.; Jin, L. Analysis of Endometrial Thickness Patterns and Pregnancy Outcomes Considering 12,991 Fresh IVF Cycles. BMC Med. Inform. Decis. Mak. 2021, 21, 176. [Google Scholar] [CrossRef]
- Cruz, F.; Bellver, J. Live Birth after Embryo Transfer in an Unresponsive Thin Endometrium. Gynecol. Endocrinol. 2014, 30, 481–484. [Google Scholar] [CrossRef]
- Acosta, A.A.; Elberger, L.; Borghi, M.; Calamera, J.C.; Chemes, H.; Doncel, G.F.; Kliman, H.; Lema, B.; Lustig, L.; Papier, S. Endometrial Dating and Determination of the Window of Implantation in Healthy Fertile Women. Fertil. Steril. 2000, 73, 788–798. [Google Scholar] [CrossRef]
- Murray, M.J.; Meyer, W.R.; Zaino, R.J.; Lessey, B.A.; Novotny, D.B.; Ireland, K.; Zeng, D.; Fritz, M.A. A Critical Analysis of the Accuracy, Reproducibility, and Clinical Utility of Histologic Endometrial Dating in Fertile Women. Fertil. Steril. 2004, 81, 1333–1343. [Google Scholar] [CrossRef]
- Alfer, J.; Fattahi, A.; Bleisinger, N.; Krieg, J.; Behrens, R.; Dittrich, R.; Beckmann, M.W.; Hartmann, A.; Classen-Linke, I.; Popovici, R.M. Endometrial Dating Method Detects Individual Maturation Sequences During the Secretory Phase. In Vivo 2020, 34, 1951–1963. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.F.; Liao, J.N.; Fan, X.X.; Hu, Y.B.; Gan, R.; Lu, G.; Lin, G.; Gong, F. Clinical Value of Histologic Endometrial Dating for Personalized Frozen-Thawed Embryo Transfer in Patients with Repeated Implantation Failure in Natural Cycles. BMC Pregnancy Childbirth 2020, 20, 527. [Google Scholar] [CrossRef] [PubMed]
- Aghajanova, L.; Stavreus-Evers, A.; Nikas, Y.; Hovatta, O.; Landgren, B.-M. Coexpression of Pinopodes and Leukemia Inhibitory Factor, as Well as Its Receptor, in Human Endometrium. Fertil. Steril. 2003, 79, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Nikas, G. Pinopodes as Markers of Endometrial Receptivity in Clinical Practice. Hum. Reprod. 1999, 14, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Johannisson, E.; Nilsson, L. Scanning Electron Microscopic Study of the Human Endometrium. Fertil. Steril. 1972, 23, 613–625. [Google Scholar] [CrossRef]
- Stavreus-Evers, A.; Nikas, G.; Sahlin, L.; Eriksson, H.; Landgren, B.-M. Formation of Pinopodes in Human Endometrium Is Associated with the Concentrations of Progesterone and Progesterone Receptors. Fertil. Steril. 2001, 76, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Usadi, R.S.; Murray, M.J.; Bagnell, R.C.; Fritz, M.A.; Kowalik, A.I.; Meyer, W.R.; Lessey, B.A. Temporal and Morphologic Characteristics of Pinopod Expression across the Secretory Phase of the Endometrial Cycle in Normally Cycling Women with Proven Fertility. Fertil. Steril. 2003, 79, 970–974. [Google Scholar] [CrossRef]
- Jin, X.Y.; Zhao, L.J.; Luo, D.H.; Liu, L.; Dai, Y.D.; Hu, X.X.; Wang, Y.Y.; Lin, X.; Hong, F.; Li, T.C.; et al. Pinopode Score around the Time of Implantation Is Predictive of Successful Implantation Following Frozen Embryo Transfer in Hormone Replacement Cycles. Hum. Reprod. 2017, 32, 2394–2403. [Google Scholar] [CrossRef]
- Qiong, Z.; Jie, H.; Yonggang, W.; Bin, X.; Jing, Z.; Yanping, L. Clinical Validation of Pinopode as a Marker of Endometrial Receptivity: A Randomized Controlled Trial. Fertil. Steril. 2017, 108, 513–517.e2. [Google Scholar] [CrossRef]
- Wu, F.; Mao, D.; Liu, Y.; Chen, X.; Xu, H.; Li, T.C.; Wang, C.C. Localization of Mucin 1 in Endometrial Luminal Epithelium and Its Expression in Women with Reproductive Failure during Implantation Window. J. Mol. Histol. 2019, 50, 563–572. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, S.; Di Nicuolo, F.; Papi, M.; Castellani, R.; Palmieri, V.; Masciullo, V.; Arena, V.; Tersigni, C.; Bernabei, M.; Pontecorvi, A.; et al. Expression of Pinopodes in the Endometrium from Recurrent Pregnancy Loss Women. Role of Thrombomodulin and Ezrin. J. Clin. Med. 2020, 9, 2634. [Google Scholar] [CrossRef] [PubMed]
- Quinn, C.; Ryan, E.; Claessens, E.A.; Greenblatt, E.; Hawrylyshyn, P.; Cruickshank, B.; Hannam, T.; Dunk, C.; Casper, R.F. The Presence of Pinopodes in the Human Endometrium Does Not Delineate the Implantation Window. Fertil. Steril. 2007, 87, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Quinn, C.E.; Casper, R.F. Pinopodes: A Questionable Role in Endometrial Receptivity. Hum. Reprod. Update 2009, 15, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Matson, B.C.; Pierce, S.L.; Espenschied, S.T.; Holle, E.; Sweatt, I.H.; Davis, E.S.; Tarran, R.; Young, S.L.; Kohout, T.A.; van Duin, M.; et al. Adrenomedullin Improves Fertility and Promotes Pinopodes and Cell Junctions in the Peri-Implantation Endometrium. Biol. Reprod. 2017, 97, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lv, S.; Mao, W.; Pei, M.; Yang, X. Assessment of Endometrial Receptivity during Implantation Window in Women with Unexplained Infertility. Gynecol. Endocrinol. 2020, 36, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Charnock-Jones, D.S.; Sharkey, A.M.; Fenwick, P.; Smith, S.K. Leukaemia Inhibitory Factor mRNA Concentration Peaks in Human Endometrium at the Time of Implantation and the Blastocyst Contains mRNA for the Receptor at This Time. J. Reprod. Fertil. 1994, 101, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Hey, N.A.; Graham, R.A.; Seif, M.W.; Aplin, J.D. The Polymorphic Epithelial Mucin MUC1 in Human Endometrium Is Regulated with Maximal Expression in the Implantation Phase. J. Clin. Endocrinol. Metab. 1994, 78, 337–342. [Google Scholar] [CrossRef]
- Hey, N.A.; Li, T.C.; Devine, P.L.; Graham, R.A.; Saravelos, H.; Aplin, J.D. MUC1 in Secretory Phase Endometrium: Expression in Precisely Dated Biopsies and Flushings from Normal and Recurrent Miscarriage Patients. Hum. Reprod. 1995, 10, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.D.; Hey, N.A.; Li, T.C. MUC1 as a Cell Surface and Secretory Component of Endometrial Epithelium: Reduced Levels in Recurrent Miscarriage. Am. J. Reprod. Immunol. 1996, 35, 261–266. [Google Scholar] [CrossRef]
- Fujimoto, J.; Ichigo, S.; Hori, M.; Tamaya, T. Alteration of E-Cadherin, α- and β-Catenin mRNA Expression in Human Uterine Endometrium during the Menstrual Cycle. Gynecol. Endocrinol. 1996, 10, 187–191. [Google Scholar] [CrossRef]
- Vogiagis, D.; Marsh, M.M.; Fry, R.C.; Salamonsen, L.A. Leukaemia Inhibitory Factor in Human Endometrium throughout the Menstrual Cycle. J. Endocrinol. 1996, 148, 95–102. [Google Scholar] [CrossRef]
- Laird, S.M.; Tuckerman, E.M.; Dalton, C.F.; Dunphy, B.C.; Li, T.C.; Zhang, X. The Production of Leukaemia Inhibitory Factor by Human Endometrium: Presence in Uterine Flushings and Production by Cells in Culture. Hum. Reprod. 1997, 12, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.D.; DeSouza, M.M.; Regisford, E.G. Mucin and Proteoglycan Functions in Embryo Implantation. Bioessays 1998, 20, 577–583. [Google Scholar] [CrossRef]
- Apparao, K.B.; Murray, M.J.; Fritz, M.A.; Meyer, W.R.; Chambers, A.F.; Truong, P.R.; Lessey, B.A. Osteopontin and Its Receptor αvβ3 Integrin Are Coexpressed in the Human Endometrium during the Menstrual Cycle but Regulated Differentially. J. Clin. Endocrinol. Metab. 2001, 86, 4991–5000. [Google Scholar] [CrossRef] [PubMed]
- Lessey, B.A. Adhesion Molecules and Implantation. J. Reprod. Immunol. 2002, 55, 101–112. [Google Scholar] [CrossRef]
- Achache, H.; Revel, A. Endometrial Receptivity Markers, the Journey to Successful Embryo Implantation. Hum. Reprod. Update 2006, 12, 731–746. [Google Scholar] [CrossRef]
- Xu, B.; Sun, X.; Li, L.; Wu, L.; Zhang, A.; Feng, Y. Pinopodes, Leukemia Inhibitory Factor, Integrin-Β3, and Mucin-1 Expression in the Peri-Implantation Endometrium of Women with Unexplained Recurrent Pregnancy Loss. Fertil. Steril. 2012, 98, 389–395. [Google Scholar] [CrossRef]
- Rosario, G.X.; Stewart, C.L. The Multifaceted Actions of Leukaemia Inhibitory Factor in Mediating Uterine Receptivity and Embryo Implantation. Am. J. Reprod. Immunol. 2016, 75, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Massimiani, M.; Lacconi, V.; La Civita, F.; Ticconi, C.; Rago, R.; Campagnolo, L. Molecular Signaling Regulating Endometrium-Blastocyst Crosstalk. Int. J. Mol. Sci. 2019, 21, 23. [Google Scholar] [CrossRef] [PubMed]
- Somkuti, S.G.; Yuan, L.; Fritz, M.A.; Lessey, B.A. Epidermal Growth Factor and Sex Steroids Dynamically Regulate a Marker of Endometrial Receptivity in Ishikawa Cells. J. Clin. Endocrinol. Metab. 1997, 82, 2192–2197. [Google Scholar] [CrossRef]
- Lessey, B.A. Two Pathways of Progesterone Action in the Human Endometrium: Implications for Implantation and Contraception. Steroids 2003, 68, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.-W.; Park, M.-J.; Kim, H.S.; Choi, H.-J.; Ha, K.-T. Integrin αVβ3 and αVβ5 Are Required for Leukemia Inhibitory Factor-Mediated the Adhesion of Trophoblast Cells to the Endometrial Cells. Biochem. Biophys. Res. Commun. 2016, 469, 936–940. [Google Scholar] [CrossRef]
- Revel, A.; Koler, M.; Prus, D.; Tsafrir, A.; Laufer, N.; Reich, R. Implementation of Integrin Β3 Level as Predictor of Implantation in an IVF Program. Fertil. Steril. 2005, 84, S144. [Google Scholar] [CrossRef]
- Lessey, B.A.; Castelbaum, A.J.; Sawin, S.W.; Sun, J. Integrins as Markers of Uterine Receptivity in Women with Primary Unexplained Infertility. Fertil. Steril. 1995, 63, 535–542. [Google Scholar] [CrossRef]
- Tei, C.; Maruyama, T.; Kuji, N.; Miyazaki, T.; Mikami, M.; Yoshimura, Y. Reduced Expression of αvβ3 Integrin in the Endometrium of Unexplained Infertility Patients with Recurrent IVF-ET Failures: Improvement by Danazol Treatment. J. Assist. Reprod. Genet. 2003, 20, 13–20. [Google Scholar] [CrossRef]
- Boroujerdnia, M.G.; Nikbakht, R. Beta3 Integrin Expression within Uterine Endometrium and Its Relationship with Unexplained Infertility. Pak. J. Biol. Sci. 2008, 11, 2495–2499. [Google Scholar] [CrossRef][Green Version]
- Elnaggar, A.; Farag, A.H.; Gaber, M.E.; Hafeez, M.A.; Ali, M.S.; Atef, A.M. AlphaVBeta3 Integrin Expression within Uterine Endometrium in Unexplained Infertility: A Prospective Cohort Study. BMC Womens Health 2017, 17, 90. [Google Scholar] [CrossRef]
- Lessey, B.A.; Castelbaum, A.J.; Sawin, S.W.; Buck, C.A.; Schinnar, R.; Bilker, W.; Strom, B.L. Aberrant Integrin Expression in the Endometrium of Women with Endometriosis. J. Clin. Endocrinol. Metab. 1994, 79, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Creus, M.; Ordi, J.; Fábregues, F.; Casamitjana, R.; Ferrer, B.; Coll, E.; Vanrell, J.A.; Balasch, J. αvβ3 Integrin Expression and Pinopod Formation in Normal and Out-of-Phase Endometria of Fertile and Infertile Women. Hum. Reprod. 2002, 17, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.; Thomson, A.; Wood, S.; Kingsland, C.; Vince, G.; Lewis-Jones, I. Endometrial Integrin Expression in Women Undergoing In Vitro Fertilization and the Association with Subsequent Treatment Outcome. Fertil. Steril. 2003, 80, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Ceydeli, N.; Kaleli, S.; Calay, Z.; Erel, C.T.; Akbas, F.; Ertungealp, E. Difference in αvβ3 Integrin Expression in Endometrial Stromal Cell in Subgroups of Women with Unexplained Infertility. Eur. J. Obstet. Gynecol. Reprod. Biol. 2006, 126, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Casals, G.; Ordi, J.; Creus, M.; Fábregues, F.; Casamitjana, R.; Quinto, L.; Campo, E.; Balasch, J. Osteopontin and αvβ3 Integrin Expression in the Endometrium of Infertile and Fertile Women. Reprod. Biomed. Online 2008, 16, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Casals, G.; Ordi, J.; Creus, M.; Fábregues, F.; Carmona, F.; Casamitjana, R.; Balasch, J. Expression Pattern of Osteopontin and αvβ3 Integrin during the Implantation Window in Infertile Patients with Early Stages of Endometriosis. Hum. Reprod. 2012, 27, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, C.; Sinagra, M.; Ledger, W.; Li, T.C.; Laird, S. Endometrial Integrin Expression in Women with Recurrent Implantation Failure after In Vitro Fertilization and Its Relationship to Pregnancy Outcome. Fertil. Steril. 2013, 100, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S.; Liu, H.C.; Davis, O.K.; Rosenwaks, Z. Tumor Necrosis Factor Is Present in Maternal Sera and Embryo Culture Fluids during In Vitro Fertilization. J. Reprod. Immunol. 1991, 19, 85–93. [Google Scholar] [CrossRef]
- Bischof, P.; Meisser, A.; Campana, A. Mechanisms of Endometrial Control of Trophoblast Invasion. J. Reprod. Fertil. Suppl. 2000, 55, 65–71. [Google Scholar]
- Meseguer, M.; Aplin, J.D.; Caballero-Campo, P.; O’Connor, J.E.; Martín, J.C.; Remohí, J.; Pellicer, A.; Simón, C. Human Endometrial Mucin MUC1 Is Up-Regulated by Progesterone and Down-Regulated In Vitro by the Human Blastocyst. Biol. Reprod. 2001, 64, 590–601. [Google Scholar] [CrossRef]
- Thathiah, A.; Blobel, C.P.; Carson, D.D. Tumor Necrosis Factor-α Converting Enzyme/ADAM 17 Mediates MUC1 Shedding. J. Biol. Chem. 2003, 278, 3386–3394. [Google Scholar] [CrossRef]
- Francis, L.W.; Yao, S.N.; Powell, L.C.; Griffiths, S.; Berquand, A.; Piasecki, T.; Howe, W.; Gazze, A.S.; Farach-Carson, M.C.; Constantinou, P.; et al. Highly Glycosylated MUC1 Mediates High Affinity L-Selectin Binding at the Human Endometrial Surface. J. Nanobiotechnol. 2021, 19, 50. [Google Scholar] [CrossRef] [PubMed]
- Surveyor, G.A.; Gendler, S.J.; Pemberton, L.; Das, S.K.; Chakraborty, I.; Julian, J.; Pimental, R.A.; Wegner, C.C.; Dey, S.K.; Carson, D.D. Expression and Steroid Hormonal Control of Muc-1 in the Mouse Uterus. Endocrinology 1995, 136, 3639–3647. [Google Scholar] [CrossRef] [PubMed]
- DeSouza, M.M.; Mani, S.K.; Julian, J.; Carson, D.D. Reduction of Mucin-1 Expression during the Receptive Phase in the Rat Uterus. Biol. Reprod. 1998, 58, 1503–1507. [Google Scholar] [CrossRef] [PubMed]
- Dharmaraj, N.; Gendler, S.J.; Carson, D.D. Expression of Human MUC1 During Early Pregnancy in the Human MUC1 Transgenic Mouse Model. Biol. Reprod. 2009, 81, 1182. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chen, X.; Liu, Y.; Liang, B.; Xu, H.; Li, T.C.; Wang, C.C. Decreased MUC1 in Endometrium Is an Independent Receptivity Marker in Recurrent Implantation Failure during Implantation Window. Reprod. Biol. Endocrinol. 2018, 16, 60. [Google Scholar] [CrossRef] [PubMed]
- Margarit, L.; Taylor, A.; Roberts, M.H.; Hopkins, L.; Davies, C.; Brenton, A.G.; Conlan, R.S.; Bunkheila, A.; Joels, L.; White, J.O.; et al. MUC1 as a Discriminator between Endometrium from Fertile and Infertile Patients with PCOS and Endometriosis. J. Clin. Endocrinol. Metab. 2010, 95, 5320–5329. [Google Scholar] [CrossRef]
- Altmäe, S.; Martinez-Conejero, J.A.; Esteban, F.J.; Ruiz-Alonso, M.; Stavreus-Evers, A.; Horcajadas, J.A.; Salumets, A. MicroRNAs miR-30b, miR-30d, and miR-494 Regulate Human Endometrial Receptivity. Reprod. Sci. 2013, 20, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Altmäe, S.; Koel, M.; Võsa, U.; Adler, P.; Suhorutšenko, M.; Laisk-Podar, T.; Kukushkina, V.; Saare, M.; Velthut-Meikas, A.; Krjutškov, K.; et al. Meta-Signature of Human Endometrial Receptivity: A Meta-Analysis and Validation Study of Transcriptomic Biomarkers. Sci. Rep. 2017, 7, 10077. [Google Scholar] [CrossRef]
- Li, Q.; Liu, W.; Chiu, P.C.N.; Yeung, W.S.B. Mir-Let-7a/g Enhances Uterine Receptivity via Suppressing Wnt/β-Catenin under the Modulation of Ovarian Hormones. Reprod. Sci. 2020, 27, 1164–1174. [Google Scholar] [CrossRef]
- Moreno-Moya, J.M.; Vilella, F.; Martínez, S.; Pellicer, A.; Simón, C. The Transcriptomic and Proteomic Effects of Ectopic Overexpression of miR-30d in Human Endometrial Epithelial Cells. Mol. Hum. Reprod. 2014, 20, 550–566. [Google Scholar] [CrossRef]
- Akbar, R.; Ullah, K.; Rahman, T.U.; Cheng, Y.; Pang, H.-Y.; Jin, L.-Y.; Wang, Q.-J.; Huang, H.-F.; Sheng, J.-Z. miR-183-5p Regulates Uterine Receptivity and Enhances Embryo Implantation. J. Mol. Endocrinol. 2020, 64, 43–52. [Google Scholar] [CrossRef]
- Liang, J.; Cao, D.; Zhang, X.; Liu, L.; Tan, Q.; Shi, S.; Chen, K.; Liang, J.; Wang, Z. miR-192-5p Suppresses Uterine Receptivity Formation through Impeding Epithelial Transformation during Embryo Implantation. Theriogenology 2020, 157, 360–371. [Google Scholar] [CrossRef]
- Huang, K.; Chen, G.; Fan, W.; Hu, L. miR-23a-3p Increases Endometrial Receptivity via CUL3 during Embryo Implantation. J. Mol. Endocrinol. 2020, 65, 35–44. [Google Scholar] [CrossRef]
- Li, L.; Gou, J.; Yi, T.; Li, Z. MicroRNA-30a-3p Regulates Epithelial-Mesenchymal Transition to Affect Embryo Implantation by Targeting Snai2. Biol. Reprod. 2019, 100, 1171–1179. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, H.; Li, W.; Bao, H.; Qu, Q.; Ma, D. Up-Regulation of miR-145 May Contribute to Repeated Implantation Failure after IVF-Embryo Transfer by Targeting PAI-1. Reprod. Biomed. Online 2020, 40, 627–636. [Google Scholar] [CrossRef]
- Zheng, Q.; Zhang, D.; Yang, Y.; Cui, X.; Sun, J.; Liang, C.; Qin, H.; Yang, X.; Liu, S.; Yan, Q. MicroRNA-200c Impairs Uterine Receptivity Formation by Targeting FUT4 and A1,3-Fucosylation. Cell Death Differ. 2017, 24, 2161–2172. [Google Scholar] [CrossRef]
- Yan, Q.; Yan, G.; Zhang, C.; Wang, Z.; Huang, C.; Wang, J.; Zhou, J.; Liu, Y.; Ding, L.; Zhang, Q.; et al. miR-21 Reverses Impaired Decidualization through Modulation of KLF12 and NR4A1 Expression in Human Endometrial Stromal Cells. Biol. Reprod. 2019, 100, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xie, Y.; Wu, M.; Geng, Y.; Li, R.; Xu, L.; Liu, X.; Pan, Y. Expression of Mmu-miR-96 in the Endometrium during Early Pregnancy and Its Regulatory Effects on Stromal Cell Apoptosis via Bcl2. Mol. Med. Rep. 2017, 15, 1547–1554. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, H.; Jiang, Y.; Xue, B.; Diao, Z.; Ding, L.; Zhen, X.; Sun, H.; Yan, G.; Hu, Y. MicroRNA-181a Is Involved in the Regulation of Human Endometrial Stromal Cell Decidualization by Inhibiting Krüppel-like Factor 12. Reprod. Biol. Endocrinol. 2015, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.T.; Mainigi, M.A.; Word, R.A.; Kraus, W.L.; Mendelson, C.R. miR-200 Regulates Endometrial Development during Early Pregnancy. Mol. Endocrinol. 2016, 30, 977–987. [Google Scholar] [CrossRef]
- Zhang, Q.; Ni, T.; Dang, Y.; Ding, L.; Jiang, J.; Li, J.; Xia, M.; Yu, N.; Ma, J.; Yan, J.; et al. MiR-148a-3p May Contribute to Flawed Decidualization in Recurrent Implantation Failure by Modulating HOXC8. J. Assist. Reprod. Genet. 2020, 37, 2535–2544. [Google Scholar] [CrossRef]
- Graham, A.; Holbert, J.; Nothnick, W.B. miR-181b-5p Modulates Cell Migratory Proteins, Tissue Inhibitor of Metalloproteinase 3, and Annexin A2 During In Vitro Decidualization in a Human Endometrial Stromal Cell Line. Reprod. Sci. 2017, 24, 1264–1274. [Google Scholar] [CrossRef]
- Pei, T.; Liu, C.; Liu, T.; Xiao, L.; Luo, B.; Tan, J.; Li, X.; Zhou, G.; Duan, C.; Huang, W. miR-194-3p Represses the Progesterone Receptor and Decidualization in Eutopic Endometrium from Women with Endometriosis. Endocrinology 2018, 159, 2554–2562. [Google Scholar] [CrossRef]
- Tochigi, H.; Kajihara, T.; Mizuno, Y.; Mizuno, Y.; Tamaru, S.; Kamei, Y.; Okazaki, Y.; Brosens, J.J.; Ishihara, O. Loss of miR-542-3p Enhances IGFBP-1 Expression in Decidualizing Human Endometrial Stromal Cells. Sci. Rep. 2017, 7, 40001. [Google Scholar] [CrossRef] [PubMed]
- von Grothusen, C.; Frisendahl, C.; Modhukur, V.; Lalitkumar, P.G.; Peters, M.; Faridani, O.R.; Salumets, A.; Boggavarapu, N.R.; Gemzell-Danielsson, K. Uterine Fluid microRNAs Are Dysregulated in Women with Recurrent Implantation Failure. Hum. Reprod. 2022, 37, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Koot, Y.E.M.; van Hooff, S.R.; Boomsma, C.M.; van Leenen, D.; Groot Koerkamp, M.J.A.; Goddijn, M.; Eijkemans, M.J.C.; Fauser, B.C.J.M.; Holstege, F.C.P.; Macklon, N.S. An Endometrial Gene Expression Signature Accurately Predicts Recurrent Implantation Failure after IVF. Sci. Rep. 2016, 6, 19411. [Google Scholar] [CrossRef]
- Doyle, N.; Jahandideh, S.; Hill, M.J.; Widra, E.A.; Levy, M.; Devine, K. Effect of Timing by Endometrial Receptivity Testing vs Standard Timing of Frozen Embryo Transfer on Live Birth in Patients Undergoing In Vitro Fertilization: A Randomized Clinical Trial. JAMA 2022, 328, 2117–2125. [Google Scholar] [CrossRef]
- Doyle, N.; Combs, J.C.; Jahandideh, S.; Wilkinson, V.; Devine, K.; O’Brien, J.E. Live Birth after Transfer of a Single Euploid Vitrified-Warmed Blastocyst According to Standard Timing versus Timing as Recommended by Endometrial Receptivity Analysis. Fertil. Steril. 2022, 118, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Enciso, M.; Carrascosa, J.P.; Sarasa, J.; Martínez-Ortiz, P.A.; Munné, S.; Horcajadas, J.A.; Aizpurua, J. Development of a New Comprehensive and Reliable Endometrial Receptivity Map (ER Map/ER Grade) Based on RT-qPCR Gene Expression Analysis. Hum. Reprod. 2018, 33, 220–228. [Google Scholar] [CrossRef]
- Enciso, M.; Aizpurua, J.; Rodríguez-Estrada, B.; Jurado, I.; Ferrández-Rives, M.; Rodríguez, E.; Pérez-Larrea, E.; Climent, A.B.; Marron, K.; Sarasa, J. The Precise Determination of the Window of Implantation Significantly Improves ART Outcomes. Sci. Rep. 2021, 11, 13420. [Google Scholar] [CrossRef] [PubMed]
- He, A.; Zou, Y.; Wan, C.; Zhao, J.; Zhang, Q.; Yao, Z.; Tian, F.; Wu, H.; Huang, X.; Fu, J.; et al. The Role of Transcriptomic Biomarkers of Endometrial Receptivity in Personalized Embryo Transfer for Patients with Repeated Implantation Failure. J. Transl. Med. 2021, 19, 176. [Google Scholar] [CrossRef] [PubMed]
- Meltsov, A.; Saare, M.; Teder, H.; Paluoja, P.; Arffman, R.K.; Piltonen, T.; Laudanski, P.; Wielgoś, M.; Gianaroli, L.; Koel, M.; et al. Targeted Gene Expression Profiling for Accurate Endometrial Receptivity Testing. Sci. Rep. 2023, 13, 13959. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, R.; Wang, R.; Huang, H.-X.; Zhong, K. Local Injury to the Endometrium in Controlled Ovarian Hyperstimulation Cycles Improves Implantation Rates. Fertil. Steril. 2008, 89, 1166–1176. [Google Scholar] [CrossRef] [PubMed]
- Mackens, S.; Santos-Ribeiro, S.; Racca, A.; Daneels, D.; Koch, A.; Essahib, W.; Verpoest, W.; Bourgain, C.; Van Riet, I.; Tournaye, H.; et al. The Proliferative Phase Endometrium in IVF/ICSI: An in-Cycle Molecular Analysis Predictive of the Outcome Following Fresh Embryo Transfer. Hum. Reprod. 2020, 35, 130–144. [Google Scholar] [CrossRef]
- Teklenburg, G.; Salker, M.; Molokhia, M.; Lavery, S.; Trew, G.; Aojanepong, T.; Mardon, H.J.; Lokugamage, A.U.; Rai, R.; Landles, C.; et al. Natural Selection of Human Embryos: Decidualizing Endometrial Stromal Cells Serve as Sensors of Embryo Quality upon Implantation. PLoS ONE 2010, 5, e10258. [Google Scholar] [CrossRef]
- Weimar, C.H.E.; Macklon, N.S.; Post Uiterweer, E.D.; Brosens, J.J.; Gellersen, B. The Motile and Invasive Capacity of Human Endometrial Stromal Cells: Implications for Normal and Impaired Reproductive Function. Hum. Reprod. Update 2013, 19, 542–557. [Google Scholar] [CrossRef]
- Peter Durairaj, R.R.; Aberkane, A.; Polanski, L.; Maruyama, Y.; Baumgarten, M.; Lucas, E.S.; Quenby, S.; Chan, J.K.Y.; Raine-Fenning, N.; Brosens, J.J.; et al. Deregulation of the Endometrial Stromal Cell Secretome Precedes Embryo Implantation Failure. Mol. Hum. Reprod. 2017, 23, 478–487. [Google Scholar] [CrossRef]
- Gurung, S.; Greening, D.W.; Catt, S.; Salamonsen, L.; Evans, J. Exosomes and Soluble Secretome from Hormone-Treated Endometrial Epithelial Cells Direct Embryo Implantation. Mol. Hum. Reprod. 2020, 26, 510–520. [Google Scholar] [CrossRef]
- Casado-Vela, J.; Rodriguez-Suarez, E.; Iloro, I.; Ametzazurra, A.; Alkorta, N.; García-Velasco, J.A.; Matorras, R.; Prieto, B.; González, S.; Nagore, D.; et al. Comprehensive Proteomic Analysis of Human Endometrial Fluid Aspirate. J. Proteome Res. 2009, 8, 4622–4632. [Google Scholar] [CrossRef]
- Hannan, N.J.; Stephens, A.N.; Rainczuk, A.; Hincks, C.; Rombauts, L.J.F.; Salamonsen, L.A. 2D-DiGE Analysis of the Human Endometrial Secretome Reveals Differences between Receptive and Nonreceptive States in Fertile and Infertile Women. J. Proteome Res. 2010, 9, 6256–6264. [Google Scholar] [CrossRef]
- Bielfeld, A.P.; Pour, S.J.; Poschmann, G.; Stühler, K.; Krüssel, J.-S.; Baston-Büst, D.M. A Proteome Approach Reveals Differences between Fertile Women and Patients with Repeated Implantation Failure on Endometrial Level—Does hCG Render the Endometrium of RIF Patients? Int. J. Mol. Sci. 2019, 20, 425. [Google Scholar] [CrossRef]
- Rai, A.; Poh, Q.H.; Fatmous, M.; Fang, H.; Gurung, S.; Vollenhoven, B.; Salamonsen, L.A.; Greening, D.W. Proteomic Profiling of Human Uterine Extracellular Vesicles Reveal Dynamic Regulation of Key Players of Embryo Implantation and Fertility during Menstrual Cycle. Proteomics 2021, 21, 2000211. [Google Scholar] [CrossRef]
- Santonocito, M.; Vento, M.; Guglielmino, M.R.; Battaglia, R.; Wahlgren, J.; Ragusa, M.; Barbagallo, D.; Borzì, P.; Rizzari, S.; Maugeri, M.; et al. Molecular Characterization of Exosomes and Their microRNA Cargo in Human Follicular Fluid: Bioinformatic Analysis Reveals That Exosomal microRNAs Control Pathways Involved in Follicular Maturation. Fertil. Steril. 2014, 102, 1751–1761.e1. [Google Scholar] [CrossRef]
- Bridi, A.; Perecin, F.; Silveira, J.C. da Extracellular Vesicles Mediated Early Embryo-Maternal Interactions. Int. J. Mol. Sci. 2020, 21, 1163. [Google Scholar] [CrossRef]
- Murdica, V.; Giacomini, E.; Makieva, S.; Zarovni, N.; Candiani, M.; Salonia, A.; Vago, R.; Viganò, P. In Vitro Cultured Human Endometrial Cells Release Extracellular Vesicles That Can Be Uptaken by Spermatozoa. Sci. Rep. 2020, 10, 8856. [Google Scholar] [CrossRef]
- Jauniaux, E.; Poston, L.; Burton, G.J. Placental-Related Diseases of Pregnancy: Involvement of Oxidative Stress and Implications in Human Evolution. Hum. Reprod. Update 2006, 12, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Hirota, Y.; Acar, N.; Tranguch, S.; Burnum, K.E.; Xie, H.; Kodama, A.; Osuga, Y.; Ustunel, I.; Friedman, D.B.; Caprioli, R.M.; et al. Uterine FK506-Binding Protein 52 (FKBP52)-Peroxiredoxin-6 (PRDX6) Signaling Protects Pregnancy from Overt Oxidative Stress. Proc. Natl. Acad. Sci. USA 2010, 107, 15577–15582. [Google Scholar] [CrossRef] [PubMed]
- Esteves, S.C.; Yarali, H.; Ubaldi, F.M.; Carvalho, J.F.; Bento, F.C.; Vaiarelli, A.; Cimadomo, D.; Özbek, İ.Y.; Polat, M.; Bozdag, G.; et al. Validation of ART Calculator for Predicting the Number of Metaphase II Oocytes Required for Obtaining at Least One Euploid Blastocyst for Transfer in Couples Undergoing In Vitro Fertilization/Intracytoplasmic Sperm Injection. Front. Endocrinol. 2020, 10, 917. [Google Scholar] [CrossRef]
- Raimundo, J.; Cabrita, P. Artificial Intelligence at Assisted Reproductive Technology. Procedia Comput. Sci. 2021, 181, 442–447. [Google Scholar] [CrossRef]
- Fanton, M.; Nutting, V.; Solano, F.; Maeder-York, P.; Hariton, E.; Barash, O.; Weckstein, L.; Sakkas, D.; Copperman, A.B.; Loewke, K. An Interpretable Machine Learning Model for Predicting the Optimal Day of Trigger during Ovarian Stimulation. Fertil. Steril. 2022, 118, 101–108. [Google Scholar] [CrossRef]
- Bulletti, F.M.; Berrettini, M.; Sciorio, R.; Bulletti, C. Artificial Intelligence Algorithms for Optimizing Assisted Reproductive Technology Programs: A Systematic Review. Glob. Transl. Med. 2023, 2, 0308. [Google Scholar] [CrossRef]
- Correia, K.F.B.; Missmer, S.A.; Weinerman, R.; Ginsburg, E.S.; Rossi, B.V. Development of a Model to Estimate the Optimal Number of Oocytes to Attempt to Fertilize during Assisted Reproductive Technology Treatment. JAMA Netw. Open 2023, 6, e2249395. [Google Scholar] [CrossRef]
- Wakimoto, Y.; Fukui, A.; Kojima, T.; Hasegawa, A.; Shigeta, M.; Shibahara, H. Application of Computer-Aided Sperm Analysis (CASA) for Detecting Sperm-Immobilizing Antibody. Am. J. Reprod. Immunol. 2018, 79, e12814. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, P.; Kazemi, E.; Zhan, Q.; Malmsten, J.E.; Toschi, M.; Zisimopoulos, P.; Sigaras, A.; Lavery, S.; Cooper, L.A.D.; Hickman, C.; et al. Deep Learning Enables Robust Assessment and Selection of Human Blastocysts after In Vitro Fertilization. NPJ Digit. Med. 2019, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Pan, W.; Jin, L.; Li, Y.; Geng, Y.; Gao, C.; Chen, G.; Wang, H.; Ma, D.; Liao, S. Artificial Intelligence in Reproductive Medicine. Reproduction 2019, 158, R139–R154. [Google Scholar] [CrossRef]
- Bormann, C.L.; Thirumalaraju, P.; Kanakasabapathy, M.K.; Kandula, H.; Souter, I.; Dimitriadis, I.; Gupta, R.; Pooniwala, R.; Shafiee, H. Consistency and Objectivity of Automated Embryo Assessments Using Deep Neural Networks. Fertil. Steril. 2020, 113, 781–787.e1. [Google Scholar] [CrossRef]
- Cimadomo, D.; Chiappetta, V.; Innocenti, F.; Saturno, G.; Taggi, M.; Marconetto, A.; Casciani, V.; Albricci, L.; Maggiulli, R.; Coticchio, G.; et al. Towards Automation in IVF: Pre-Clinical Validation of a Deep Learning-Based Embryo Grading System during PGT-A Cycles. J. Clin. Med. 2023, 12, 1806. [Google Scholar] [CrossRef] [PubMed]
- Coticchio, G.; Barrie, A.; Lagalla, C.; Borini, A.; Fishel, S.; Griffin, D.; Campbell, A. Plasticity of the Human Preimplantation Embryo: Developmental Dogmas, Variations on Themes and Self-Correction. Hum. Reprod. Update 2021, 27, 848–865. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.R.; Al-Insaif, S.; Hossain, M.I.; Kamruzzaman, J. A Machine Learning Approach for Prediction of Pregnancy Outcome Following IVF Treatment. Neural Comput. Appl. 2020, 32, 2283–2297. [Google Scholar] [CrossRef]
- Singhal, N.; Mukherjee, S.; Perrey, C. Automated Assessment of Endometrium from Transvaginal Ultrasound Using Deep Learned Snake. In Proceedings of the 2017 IEEE 14th International Symposium on Biomedical Imaging (ISBI 2017), Melbourne, VIC, Australia, 18–21 April 2017; pp. 283–286. [Google Scholar]
- Hu, S.-Y.; Xu, H.; Li, Q.; Telfer, B.A.; Brattain, L.J.; Samir, A.E. Deep Learning-Based Automatic Endometrium Segmentation and Thickness Measurement for 2D Transvaginal Ultrasound. In Proceedings of the 2019 41st Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Berlin, Germany, 23–27 July 2019; Volume 2019, pp. 993–997. [Google Scholar] [CrossRef]


| Assay | Outcome | Samples of Study | References |
|---|---|---|---|
| Endometrial ReceptivityAssay (ERA) | Identification of 238 genes to classify the endometrium as non-receptive, pre-receptive, receptive, or post-receptive | Biopsies from healthy fertile women, RIF, hydrosalpinx, and sub-fertile | [25,26,27,29,30,31,33,209,210] |
| High throughput RT-qPCR | Identification of 40 genes to classify the endometrium as receptive or non-receptive | Biopsies from fertile and sub-fertile patients | [211,212] |
| RNA-Seq-based endometrial receptivity test (rsERT) | Identification of 175 predictive genes to identify the receptive endometrium | Biopsies from RIF | [213] |
| Targeted Allele Countingby sequencing (TAC-seq) | Identification of 68 endometrial receptivity genes to identify pre-receptive, receptive, and post-receptive endometrium | Biopsies from healthy volunteers and RIF | [214] |
| Transcriptome analysis (gene chip analysis) | Identification of 218 genes in proliferative phase to perform fresh embryo transfer in the same cycle | Biopsies from women undergoing fresh IVF-ET cycles | [215] |
| Secretome analysis | Identification of 45 cytokines in media from in vitro culture of decidualized hESC | Biopsies from fertile patients and RIF | [216] |
| Secretome analysis | Identification of the exosome fraction able to stimulate adhesion and outgrowth of trophoblast spheroids | ECC1 epithelial cell line | [220] |
| Secretome analysis | Identification of proteins, secreted factors, nucleic acid, and sEV and exosomes differentially expressed between proliferative and secretory phases of endometrial sample | Uterine fluid and/or trophoblast cell line | [221,222,223,224,225,226,227] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacconi, V.; Massimiani, M.; Carriero, I.; Bianco, C.; Ticconi, C.; Pavone, V.; Alteri, A.; Muzii, L.; Rago, R.; Pisaturo, V.; et al. When the Embryo Meets the Endometrium: Identifying the Features Required for Successful Embryo Implantation. Int. J. Mol. Sci. 2024, 25, 2834. https://doi.org/10.3390/ijms25052834
Lacconi V, Massimiani M, Carriero I, Bianco C, Ticconi C, Pavone V, Alteri A, Muzii L, Rago R, Pisaturo V, et al. When the Embryo Meets the Endometrium: Identifying the Features Required for Successful Embryo Implantation. International Journal of Molecular Sciences. 2024; 25(5):2834. https://doi.org/10.3390/ijms25052834
Chicago/Turabian StyleLacconi, Valentina, Micol Massimiani, Ilenia Carriero, Claudia Bianco, Carlo Ticconi, Valentina Pavone, Alessandra Alteri, Ludovico Muzii, Rocco Rago, Valerio Pisaturo, and et al. 2024. "When the Embryo Meets the Endometrium: Identifying the Features Required for Successful Embryo Implantation" International Journal of Molecular Sciences 25, no. 5: 2834. https://doi.org/10.3390/ijms25052834
APA StyleLacconi, V., Massimiani, M., Carriero, I., Bianco, C., Ticconi, C., Pavone, V., Alteri, A., Muzii, L., Rago, R., Pisaturo, V., & Campagnolo, L. (2024). When the Embryo Meets the Endometrium: Identifying the Features Required for Successful Embryo Implantation. International Journal of Molecular Sciences, 25(5), 2834. https://doi.org/10.3390/ijms25052834