
Engineered Chimera Protein Constructs to Facilitate the Production of Heterologous Transmembrane Proteins in E. coli



Abstract
1. Introduction
2. Fusion Strategies to Produce Heterologous Transmembrane Proteins in E. coli
2.1. Fusion Proteins Aid the Insertion and Folding of Heterologous TMPs in the E. coli Plasma Membrane
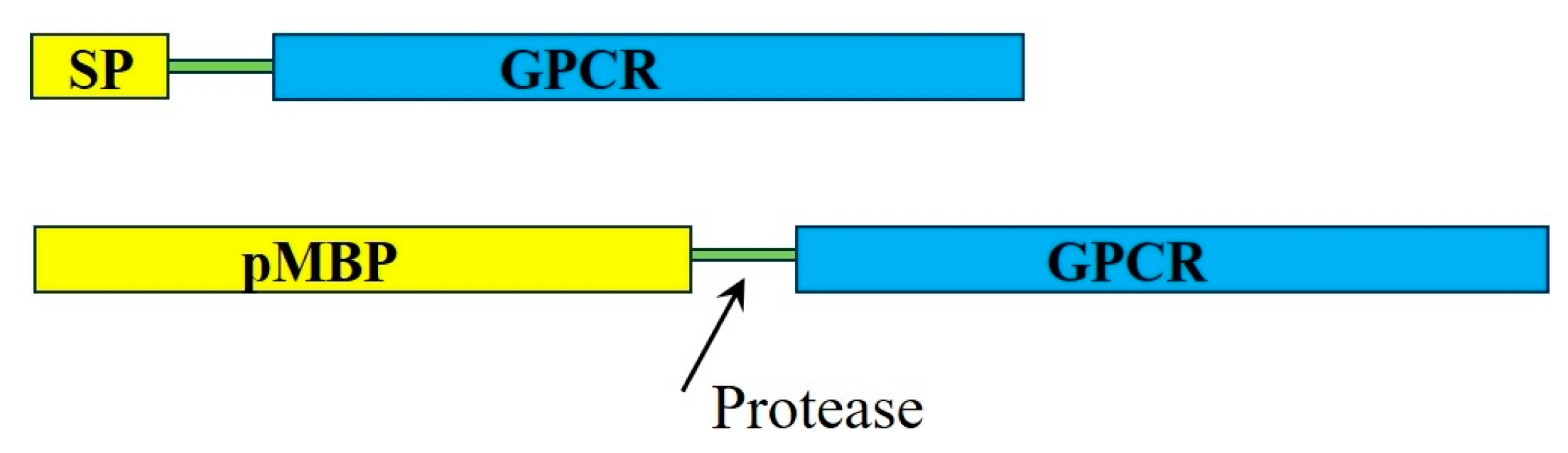
2.1.1. Signal Peptides and Precursor Maltose Binding Protein Fusion Strategies
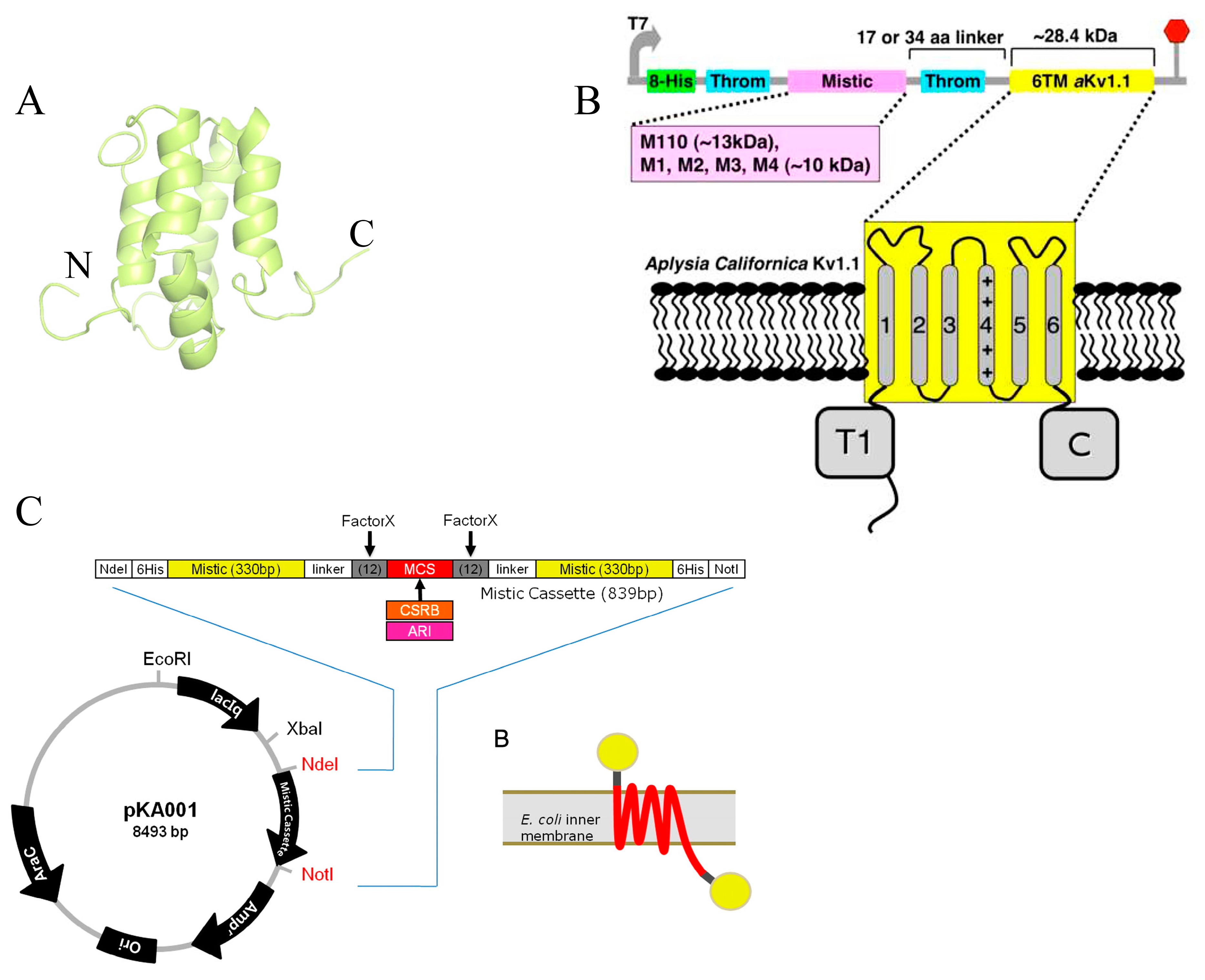
2.1.2. Mistic Protein Fusion Strategies
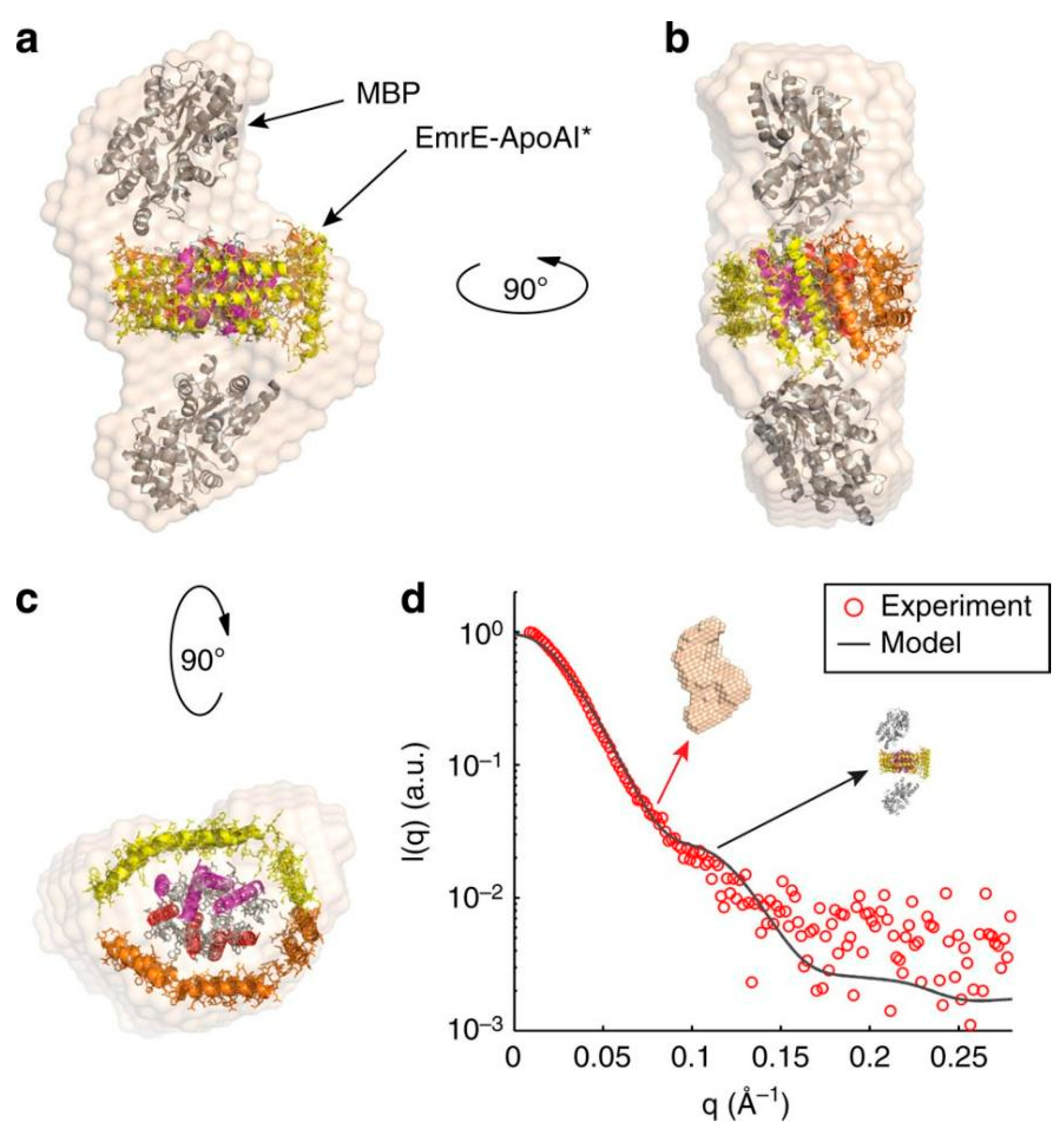
2.1.3. Apolipoprotein A-I Fusion Strategy
2.2. Fusion Proteins Aid the Production of Heterologous TMPs in Soluble Form in E. coli
2.2.1. Mature (without Signal Peptide) Maltose-Binding Protein Fusion Strategies
2.2.2. Apolipoprotein A-I Strategies to Produce Soluble TMPs
2.2.3. Other Protein Design Strategies to Produce and Stabilize Soluble TMPs
3. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cournia, Z.; Allen, T.W.; Andricioaei, I.; Antonny, B.; Baum, D.; Brannigan, G.; Buchete, N.-V.; Deckman, J.T.; Delemotte, L.; Del Val, C.; et al. Membrane Protein Structure, Function, and Dynamics: A Perspective from Experiments and Theory. J. Membr. Biol. 2015, 248, 611–640. [Google Scholar] [CrossRef]
- Kim, S.; Jeon, T.J.; Oberai, A.; Yang, D.; Schmidt, J.J.; Bowie, J.U. Transmembrane glycine zippers: Physiological and pathological roles in membrane proteins. Proc. Natl. Acad. Sci. USA 2005, 102, 14278–14283. [Google Scholar] [CrossRef]
- Robertson, J.L. The lipid bilayer membrane and its protein constituents. J. Gen. Physiol. 2018, 150, 1472–1483. [Google Scholar] [CrossRef]
- Marinko, J.T.; Huang, H.; Penn, W.D.; Capra, J.A.; Schlebach, J.P.; Sanders, C.R. Folding and Misfolding of Human Membrane Proteins in Health and Disease: From Single Molecules to Cellular Proteostasis. Chem. Rev. 2019, 119, 5537–5606. [Google Scholar] [CrossRef] [PubMed]
- Wallin, E.; von Heijne, G. Genome-wide analysis of integral membrane proteins from eubacterial, archaean, and eukaryotic organisms. Protein Sci. 1998, 7, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Flynn, A.D. Drugging Membrane Protein Interactions. Annu. Rev. Biomed. Eng. 2016, 18, 51–76. [Google Scholar] [CrossRef] [PubMed]
- Majeed, S.; Ahmad, A.B.; Sehar, U.; Georgieva, E.R. Lipid Membrane Mimetics in Functional and Structural Studies of Integral Membrane Proteins. Membranes 2021, 11, 685. [Google Scholar] [CrossRef] [PubMed]
- Errasti-Murugarren, E.; Bartoccioni, P.; Palacin, M. Membrane Protein Stabilization Strategies for Structural and Functional Studies. Membranes 2021, 11, 155. [Google Scholar] [CrossRef]
- Hu, J.; Qin, H.; Li, C.; Sharma, M.; Cross, T.A.; Gao, F.P. Structural biology of transmembrane domains: Efficient production and characterization of transmembrane peptides by NMR. Protein Sci. 2007, 16, 2153–2165. [Google Scholar] [CrossRef]
- Gulezian, E.; Crivello, C.; Bednenko, J.; Zafra, C.; Zhang, Y.; Colussi, P.; Hussain, S. Membrane protein production and formulation for drug discovery. Trends Pharmacol. Sci. 2021, 42, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Kesidis, A.; Depping, P.; Lodé, A.; Vaitsopoulou, A.; Bill, R.M.; Goddard, A.D.; Rothnie, A.J. Expression of Eukaryotic Membrane Proteins in Eukaryotic and Prokaryotic Hosts. Available online: https://publications.aston.ac.uk/id/eprint/41666/1/Kesidis_et_al_2020_accepted_version.pdf (accessed on 9 November 2023).
- Bernaudat, F.; Frelet-Barrand, A.; Pochon, N.; Dementin, S.; Hivin, P.; Boutigny, S.; Rioux, J.B.; Salvi, D.; Seigneurin-Berny, D.; Richaud, P.; et al. Heterologous expression of membrane proteins: Choosing the appropriate host. PLoS ONE 2011, 6, e29191. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef]
- Hattab, G.; Warschawski, D.E.; Moncoq, K.; Miroux, B. Escherichia coli as host for membrane protein structure determination: A global analysis. Sci. Rep. 2015, 5, 12097. [Google Scholar] [CrossRef]
- Mathieu, K.; Javed, W.; Vallet, S.; Lesterlin, C.; Candusso, M.P.; Ding, F.; Xu, X.N.; Ebel, C.; Jault, J.-M.; Orelle, C. Functionality of membrane proteins overexpressed and purified from E. coli is highly dependent upon the strain. Sci. Rep. 2019, 9, 2654. [Google Scholar] [CrossRef]
- Kleiner-Grote, G.R.; Risse, J.M.; Friehs, K. Secretion of recombinant proteins from E. coli. Eng. Life Sci. 2018, 18, 532–550. [Google Scholar] [CrossRef]
- Francis, D.M.; Page, R. Strategies to optimize protein expression in E. coli. Curr. Protoc. Protein Sci. 2010, 61, 5–24. [Google Scholar] [CrossRef]
- Korepanova, A.; Moore, J.D.; Nguyen, H.B.; Hua, Y.; Cross, T.A.; Gao, F. Expression of membrane proteins from Mycobacterium tuberculosis in Escherichia coli as fusions with maltose binding protein. Protein Expr. Purif. 2007, 53, 24–30. [Google Scholar] [CrossRef]
- Dvir, H.; Choe, S. Bacterial expression of a eukaryotic membrane protein in fusion to various Mistic orthologs. Protein Expr. Purif. 2009, 68, 28–33. [Google Scholar] [CrossRef]
- Rogl, H.; Kosemund, K.; Kuhlbrandt, W.; Collinson, I. Refolding of Escherichia coli produced membrane protein inclusion bodies immobilised by nickel chelating chromatography. FEBS Lett. 1998, 432, 21–26. [Google Scholar] [CrossRef]
- Lee, K.A.; Lee, S.S.; Kim, S.Y.; Choi, A.R.; Lee, J.H.; Jung, K.H. Mistic-fused expression of algal rhodopsins in Escherichia coli and its photochemical properties. Biochim. Biophys. Acta 2015, 1850, 1694–1703. [Google Scholar] [CrossRef]
- Reuten, R.; Nikodemus, D.; Oliveira, M.B.; Patel, T.R.; Brachvogel, B.; Breloy, I.; Stetefeld, J.; Koch, M. Maltose-binding protein (MBP), a secretion-enhancing tag for mammalian protein expression systems. PLoS ONE 2016, 11, e0152386. [Google Scholar] [CrossRef]
- Majeed, S.; Adetuyi, O.; Borbat, P.P.; Islam, M.M.; Ishola, O.; Zhao, B.; Georgieva, E.R. Insights into the oligomeric structure of the HIV-1 Vpu protein. J. Struct. Biol. 2023, 215, 107943. [Google Scholar] [CrossRef]
- Majeed, S.; Dang, L.; Islam, M.M.; Ishola, O.; Borbat, P.P.; Ludtke, S.J.; Georgieva, E.R. HIV-1 Vpu protein forms stable oligomers in aqueous solution via its transmembrane domain self-association. Sci. Rep. 2023, 13, 14691. [Google Scholar] [CrossRef] [PubMed]
- Mizrachi, D.; Chen, Y.; Liu, J.; Peng, H.M.; Ke, A.; Pollack, L.; Turner, R.J.; Auchus, R.J.; DeLisa, M.P. Making water-soluble integral membrane proteins in vivo using an amphipathic protein fusion strategy. Nat. Commun. 2015, 6, 6826. [Google Scholar] [CrossRef] [PubMed]
- Bertin, B.; Freissmuth, M.; Breyer, R.M.; Schütz, W.; Strosberg, A.D.; Marullo, S. Functional expression of the human serotonin 5-HT1A receptor in Escherichia coli. Ligand binding properties and interaction with recombinant G protein alpha-subunits. J. Biol. Chem. 1992, 267, 8200–8206. [Google Scholar] [CrossRef] [PubMed]
- Grisshammer, R.; Duckworth, R.; Henderson, R. Expression of a rat neurotensin receptor in Escherichia coli. Biochem. J. 1993, 295, 571–576. [Google Scholar] [CrossRef]
- Grisshammer, R.; Little, J.; Aharony, D. Expression of rat NK-2 (neurokinin A) receptor in E. coli. Recept. Channels 1994, 2, 295–302. [Google Scholar] [PubMed]
- Furukawa, H.; Haga, T. Expression of functional M2 muscarinic acetylcholine receptor in Escherichia coli. J. Biochem. 2000, 127, 151–161. [Google Scholar] [CrossRef]
- Yeliseev, A.A.; Wong, K.K.; Soubias, O.; Gawrisch, K. Expression of human peripheral cannabinoid receptor for structural studies. Protein Sci. 2005, 14, 2638–2653. [Google Scholar] [CrossRef] [PubMed]
- Ishola, O.; Ogunbowale, A.; Islam, M.M.; Hadadianpour, E.; Majeed, S.; Adetuyi, O.; Georgieva, E.R. Protein engineering, production, reconstitution in lipid nanoparticles, and initial characterization of the Mycobacterium tuberculosis EfpA drug exporter. bioRxiv 2023. [Google Scholar] [CrossRef]
- Eliseev, R.; Alexandrov, A.; Gunter, T. High-yield expression and purification of p18 form of Bax as an MBP-fusion protein. Protein Expr. Purif. 2004, 35, 206–209. [Google Scholar] [CrossRef]
- Lei, X.; Ahn, K.; Zhu, L.; Ubarretxena-Belandia, I.; Li, Y.-M. Soluble oligomers of the intramembrane serine protease YqgP are catalytically active in the absence of detergents. Biochemistry 2008, 47, 11920–11929. [Google Scholar] [CrossRef]
- Fiermonte, G.; Walker, J.E.; Palmieri, F. Abundant bacterial expression and reconstitution of an intrinsic membrane-transport protein from bovine mitochondria. Biochem. J. 1993, 294 Pt 1, 293–299. [Google Scholar] [CrossRef]
- Jappelli, R.; Perrin, M.H.; Lewis, K.A.; Vaughan, J.M.; Tzitzilonis, C.; Rivier, J.E.; Vale, W.W.; Riek, R. Expression and functional characterization of membrane-integrated mammalian corticotropin releasing factor receptors 1 and 2 in Escherichia coli. PLoS ONE 2014, 9, e84013. [Google Scholar] [CrossRef]
- Tucker, J.; Grisshammer, R. Purification of a rat neurotensin receptor expressed in Escherichia coli. Biochem. J. 1996, 317 Pt 3, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Abiko, L.A.; Rogowski, M.; Gautier, A.; Schertler, G.; Grzesiek, S. Efficient production of a functional G protein-coupled receptor in E. coli for structural studies. J. Biomol. NMR 2021, 75, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Egloff, P.; Hillenbrand, M.; Klenk, C.; Batyuk, A.; Heine, P.; Balada, S.; Schlinkmann, K.M.; Scott, D.J.; Schutz, M.; Pluckthun, A. Structure of signaling-competent neurotensin receptor 1 obtained by directed evolution in Escherichia coli. Proc. Natl. Acad. Sci. USA 2014, 111, E655–E662. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.J.; Williams, L.M.; Abdul-Ridha, A.; Gunatilaka, A.; Vaid, T.M.; Kocan, M.; Whitehead, A.R.; Griffin, M.D.W.; Bathgate, R.A.D.; Scott, D.J.; et al. Probing the correlation between ligand efficacy and conformational diversity at the alpha(1A)-adrenoreceptor reveals allosteric coupling of its microswitches. J. Biol. Chem. 2020, 295, 7404–7417. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Deluigi, M.; Pantić, M.; Vacca, S.; Baumann, C.; Scott, D.J.; Pluckthun, A.; Zerbe, O. Optimizing the alpha(1B)-adrenergic receptor for solution NMR studies. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183354. [Google Scholar] [CrossRef]
- Waugh, D.S. An overview of enzymatic reagents for the removal of affinity tags. Protein Expr. Purif. 2011, 80, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.; Feng, R.; Tong, Q.; Zhang, Y.; Xie, X.-Q. Mistic and TarCF as fusion protein partners for functional expression of the cannabinoid receptor 2 in Escherichia coli. Protein Expr. Purif. 2012, 83, 128–134. [Google Scholar] [CrossRef]
- Roosild, T.P.; Greenwald, J.; Vega, M.; Castronovo, S.; Riek, R.; Choe, S. NMR structure of Mistic, a membrane-integrating protein for membrane protein expression. Science 2005, 307, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Dvir, H.; Lundberg, M.E.; Maji, S.K.; Riek, R.; Choe, S. Mistic: Cellular localization, solution behavior, polymerization, and fibril formation. Protein Sci. 2009, 18, 1564–1570. [Google Scholar] [CrossRef] [PubMed]
- Marino, J.; Bordag, N.; Keller, S.; Zerbe, O. Mistic’s membrane association and its assistance in overexpression of a human GPCR are independent processes. Protein Sci. 2015, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Marino, J.; Geertsma, E.R.; Zerbe, O. Topogenesis of heterologously expressed fragments of the human Y4 GPCR. Biochim. Biophys. Acta 2012, 1818, 3055–3063. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.S.; Astrinidis, S.A.; Eisenhardt, N.; Sieverding, C.; Redolfi, J.; Lorenz, M.; Weberruss, M.; Moreno-Andres, D.; Antonin, W. MISTIC-fusion proteins as antigens for high quality membrane protein antibodies. Sci. Rep. 2017, 7, 41519. [Google Scholar] [CrossRef] [PubMed]
- Blain, K.Y.; Kwiatkowski, W.; Choe, S. The functionally active Mistic-fused histidine kinase receptor, EnvZ. Biochemistry 2010, 49, 9089–9095. [Google Scholar] [CrossRef]
- Curtiss, L.K.; Valenta, D.T.; Hime, N.J.; Rye, K.A. What is so special about apolipoprotein AI in reverse cholesterol transport? Arterioscler. Thromb. Vasc. Biol. 2006, 26, 12–19. [Google Scholar] [CrossRef]
- Denisov, I.G.; Grinkova, Y.V.; Lazarides, A.A.; Sligar, S.G. Directed self-assembly of monodisperse phospholipid bilayer Nanodiscs with controlled size. J. Am. Chem. Soc. 2004, 126, 3477–3487. [Google Scholar] [CrossRef]
- Denisov, I.G.; Sligar, S.G. Nanodiscs for structural and functional studies of membrane proteins. Nat. Struct. Mol. Biol. 2016, 23, 481–486. [Google Scholar] [CrossRef]
- Denisov, I.G.; Sligar, S.G. Nanodiscs in Membrane Biochemistry and Biophysics. Chem. Rev. 2017, 117, 4669–4713. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.O.; Forte, T.M.; Oda, M.N. Optimized bacterial expression of human apolipoprotein A-I. Protein Expr. Purif. 2003, 27, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, H.; Krieger, J.; Olszewski, J.D.; von Heijne, G.; Prestwich, G.D.; Breer, H. Expression of an olfactory receptor in Escherichia coli: Purification, reconstitution, and ligand binding. Biochemistry 1996, 35, 16077–16084. [Google Scholar] [CrossRef] [PubMed]
- Leviatan, S.; Sawada, K.; Moriyama, Y.; Nelson, N. Combinatorial method for overexpression of membrane proteins in Escherichia coli. J. Biol. Chem. 2010, 285, 23548–23556. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Qin, H.; Gao, F.P.; Cross, T.A. A systematic assessment of mature MBP in membrane protein production: Overexpression, membrane targeting and purification. Protein Expr. Purif. 2011, 80, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Gouaux, J.E. Overexpression of bacterio-opsin in Escherichia coli as a water-soluble fusion to maltose binding protein: Efficient regeneration of the fusion protein and selective cleavage with trypsin. Protein Sci. 1996, 5, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Duong-Ly, K.C.; Gabelli, S.B. Affinity Purification of a Recombinant Protein Expressed as a Fusion with the Maltose-Binding Protein (MBP) Tag. Methods Enzymol. 2015, 559, 17–26. [Google Scholar]
- Sun, P.; Tropea, J.E.; Waugh, D.S. Enhancing the solubility of recombinant proteins in Escherichia coli by using hexahistidine-tagged maltose-binding protein as a fusion partner. In Heterologous Gene Expression in E. coli: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2011; pp. 259–274. [Google Scholar]
- Lebendiker, M.; Danieli, T. Purification of proteins fused to maltose-binding protein. In Protein Chromatography: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2011; pp. 281–293. [Google Scholar]
- Nallamsetty, S.; Waugh, D.S. Mutations that alter the equilibrium between open and closed conformations of Escherichia coli maltose-binding protein impede its ability to enhance the solubility of passenger proteins. Biochem. Biophys. Res. Commun. 2007, 364, 639–644. [Google Scholar] [CrossRef]
- Raran-Kurussi, S.; Waugh, D.S. The ability to enhance the solubility of its fusion partners is an intrinsic property of maltose-binding protein but their folding is either spontaneous or chaperone-mediated. PLoS ONE 2012, 7, e49589. [Google Scholar] [CrossRef]
- Raran-Kurussi, S.; Keefe, K.; Waugh, D.S. Positional effects of fusion partners on the yield and solubility of MBP fusion proteins. Protein Expr. Purif. 2015, 110, 159–164. [Google Scholar] [CrossRef]
- Luckey, M. Membrane Structural Biology: With Biochemical and Biophysical Foundations; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Smalinskaitė, L.; Hegde, R.S. The biogenesis of multipass membrane proteins. Cold Spring Harb. Perspect. Biol. 2023, 15, a041251. [Google Scholar] [CrossRef]
- Brown, M.S.; Ye, J.; Rawson, R.B.; Goldstein, J.L. Regulated intramembrane proteolysis: A control mechanism conserved from bacteria to humans. Cell 2000, 100, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, M.S.; Kopan, R. Intramembrane proteolysis: Theme and variations. Science 2004, 305, 1119–1123. [Google Scholar] [CrossRef]
- Georgieva, E.R. Protein Conformational Dynamics upon Association with the Surfaces of Lipid Membranes and Engineered Nanoparticles: Insights from Electron Paramagnetic Resonance Spectroscopy. Molecules 2020, 25, 5393. [Google Scholar] [CrossRef]
- Franke, D.; Svergun, D.I. DAMMIF, a program for rapid ab-initio shape determination in small-angle scattering. J. Appl. Crystallogr. 2009, 42, 342–346. [Google Scholar] [CrossRef]
- Liu, S.; Li, S.; Krezel, A.M.; Li, W. Stabilization and structure determination of integral membrane proteins by termini restraining. Nat. Protoc. 2022, 17, 540–565. [Google Scholar] [CrossRef] [PubMed]
- Andrews, D.A.; Xie, M.; Hughes, V.; Wilce, M.C.; Roujeinikova, A. Design, purification and characterization of a soluble variant of the integral membrane protein MotB for structural studies. J. R. Soc. Interface 2013, 10, 20120717. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Imada, K.; Namba, K. Molecular motors of the bacterial flagella. Curr. Opin. Struct. Biol. 2008, 18, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Schafmeister, C.E.; Miercke, L.J.; Stroud, R.M. Structure at 2.5 A of a designed peptide that maintains solubility of membrane proteins. Science 1993, 262, 734–738. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fusion Tag | Produced TMP | Benefit for Structural and/or Functional Studies | Protein Yield |
|---|---|---|---|
| MBP signal peptide/entire MBP | Serotonin 5-HT1A [27], neurotensin receptor [28], NK-2 (neurokinin A) [29], M2 muscarinic acetyl choline receptor [30], and peripheral cannabinoid receptor [31] | It promotes the proper folding and insertion of the recombinant fusion protein into the plasma membrane. It supports the application of functional assays in the study of the activities of the transmembrane protein. | Yeliseev et al. (2005) showed that the small scale purification yielded 1–2 mg of recombinant receptor/1 L of culture [31] |
| Mistic protein | aKv1.1 channel [20], and eukaryotic type I rhodopsin [22] | It promotes high expression yield of heterologous TMPs as well as facilitating the expression of functional proteins with both N-terminus inside or N-terminus outside. | The yield of Mistic-aKv1.1 was approximately 2 mg/L culture [20] The Mistic-ARI yield amounted to roughly 0.12 mg/L culture [22] |
| Apolipoprotein AI | Mtb-EfpA [32], EmrE transporter [26], human cyt b5 [26], HSD17β3 [26], GluA2 [26], DsbB [26], CLDN1 [26], CLDN3 [26], S5ɑR1 [26], S5ɑR2 [26], NRC-1bR [26], OmpX [26], and VDAC1 [26] | The tertiary conformation of the TMP–lipid–apoAI forms a discoidal nanoparticle stabilized by a double belt of apoAI It increases the solubilization of TMPs with high levels of expression and supports the functional study of the protein (e.g., ligand binding and protein–protein interaction). | Mizrachi et al. (2015) showed that the diverse range of IMPs yielded approximately 5–10 mg/1 L of culture [26] |
| mMBP without signal peptide | Vpu [24], p18 [33], and Yqgp protease [34] | It is useful as a purification affinity tag when in combination with polyhistidine tag for Ni-affinity purification. It is a natural fusion tag that is a solubility enhancer. | The approximate yield of MBP-p18 reached around 20mg/L culture [33] The yield of MBP-Yqgp protease was 2 mg/1 L of culture [34] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogunbowale, A.; Georgieva, E.R. Engineered Chimera Protein Constructs to Facilitate the Production of Heterologous Transmembrane Proteins in E. coli. Int. J. Mol. Sci. 2024, 25, 2354. https://doi.org/10.3390/ijms25042354
Ogunbowale A, Georgieva ER. Engineered Chimera Protein Constructs to Facilitate the Production of Heterologous Transmembrane Proteins in E. coli. International Journal of Molecular Sciences. 2024; 25(4):2354. https://doi.org/10.3390/ijms25042354
Chicago/Turabian StyleOgunbowale, Adeyemi, and Elka R. Georgieva. 2024. "Engineered Chimera Protein Constructs to Facilitate the Production of Heterologous Transmembrane Proteins in E. coli" International Journal of Molecular Sciences 25, no. 4: 2354. https://doi.org/10.3390/ijms25042354
APA StyleOgunbowale, A., & Georgieva, E. R. (2024). Engineered Chimera Protein Constructs to Facilitate the Production of Heterologous Transmembrane Proteins in E. coli. International Journal of Molecular Sciences, 25(4), 2354. https://doi.org/10.3390/ijms25042354