
Arsenic Impairs Wound Healing Processes in Dermal Fibroblasts and Mice
 , ,
, ,
Abstract
1. Introduction
2. Results
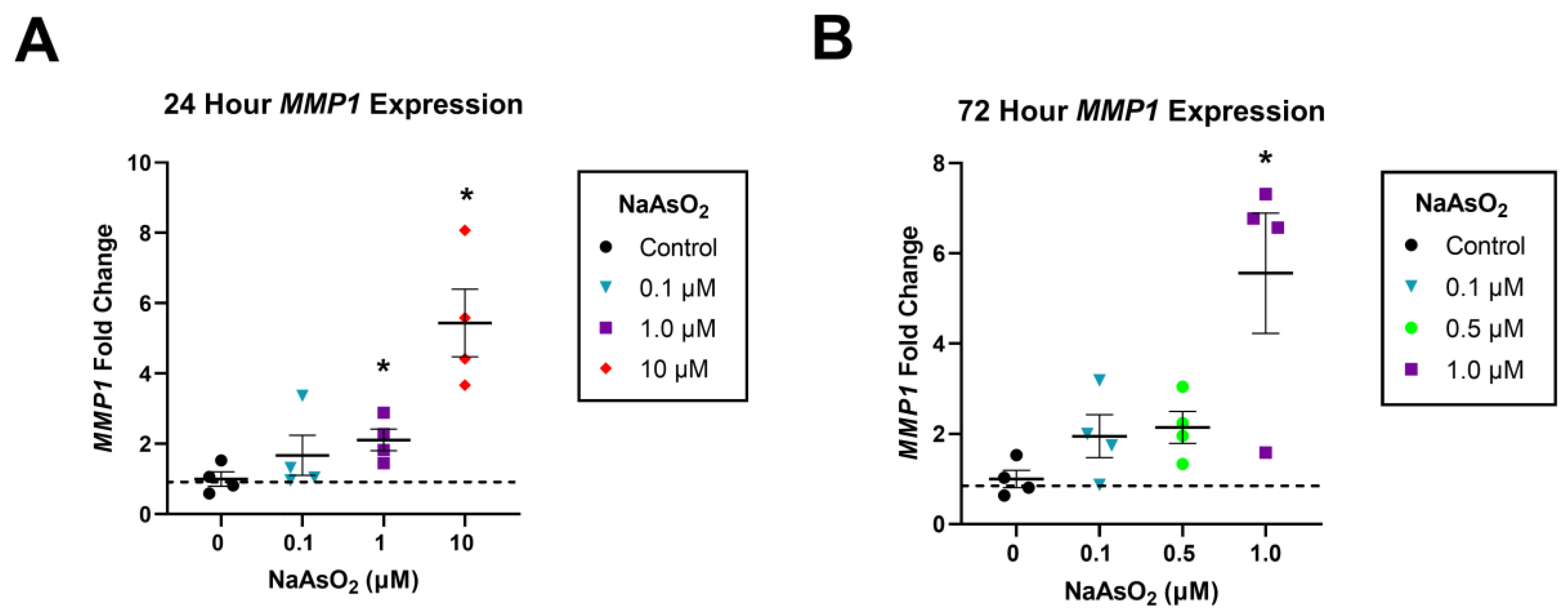
2.1. NaAsO2 Exposure Impairs Scratch Closure and Upregulates MMP1 in Cultured Dermal Fibroblasts
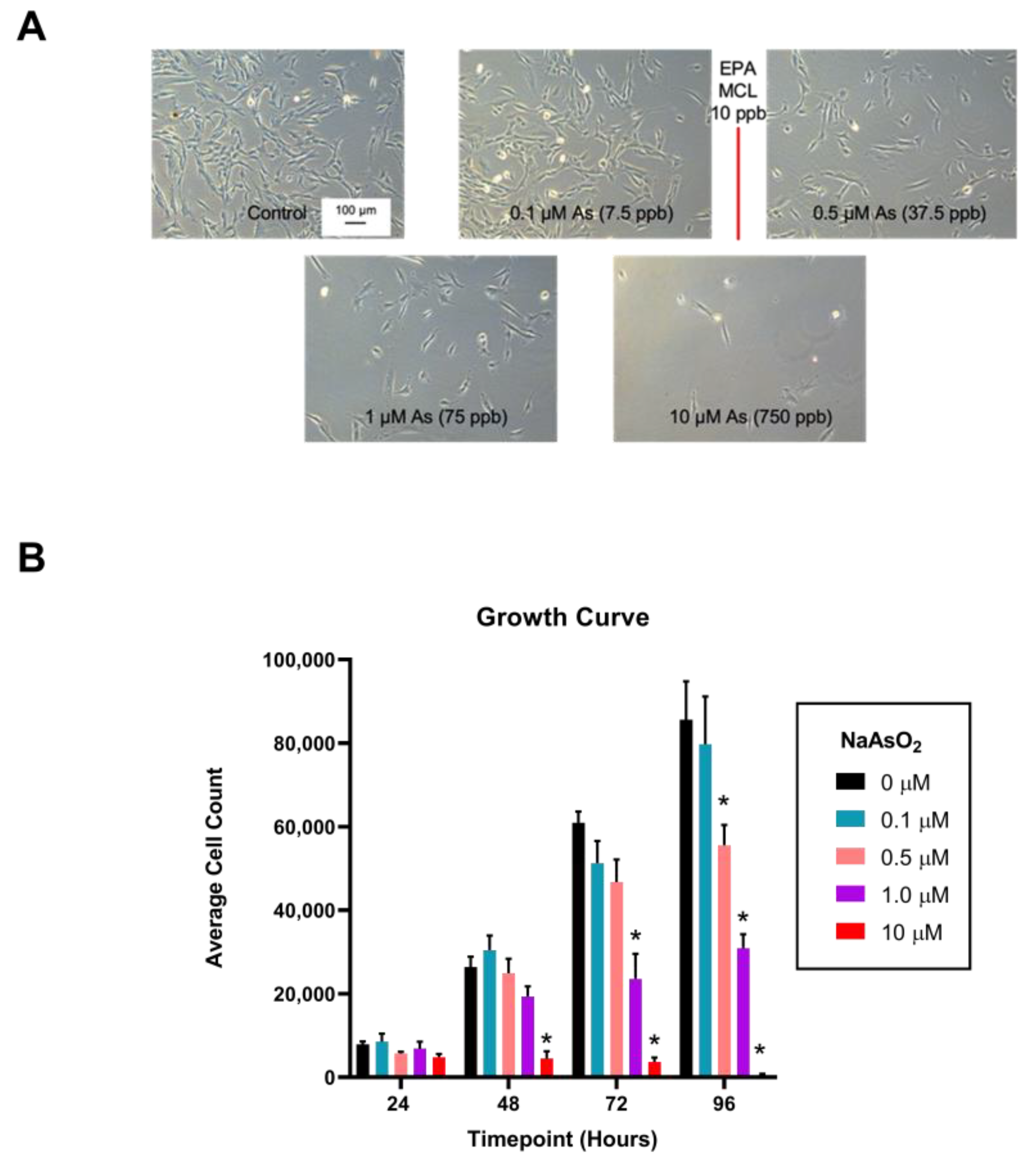
2.2. Exposure to NaAsO2 in Dermal Fibroblasts Alters Cell Morphology and Slows Proliferation
2.3. NaAsO2 Exposure for 24 h at the Time of Scratch Reduces Cell Metabolism and Alters DNA Quantity
2.4. Exposure to NaAsO2 for 72 h Reduces Cell Metabolism and Alters DNA Quantity as a Measure of Viability
2.5. Reduction in Metabolic Output Relative to Viable DNA Content after 24 and 72 h Exposure to NaAsO2
2.6. Wound Healing Processes in Female Mice Are Negatively Affected by NaAsO2-Exposure
2.7. Altered Gene Expression in Mouse Wound Biopsies due to NaAsO2-Exposure
3. Discussion
4. Materials and Methods
4.1. Arsenic Preparation
4.2. General Cell Culture Conditions
4.3. Exposure for 24 h and 72 h Cell Scratch Assay
4.4. Quantitative Polymerase Chain Reaction (qPCR) in Human Dermal Fibroblasts
4.5. Cell Proliferation and Viability Assays
4.6. Growth Curve Assay
4.7. Full-Thickness Wound Creation and Image Analysis
4.8. Tissue Collection and Quantitative Polymerase Chain Reaction (qPCR) in Mouse Wound Biopsies
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayotte, J.D.; Medalie, L.; Qi, S.L.; Backer, L.C.; Nolan, B.T. Estimating the High-Arsenic Domestic-Well Population in the Conterminous United States. Environ. Sci. Technol. 2017, 51, 12443–12454. [Google Scholar] [CrossRef] [PubMed]
- Abdul, K.S.; Jayasinghe, S.S.; Chandana, E.P.; Jayasumana, C.; De Silva, P.M. Arsenic and human health effects: A review. Environ. Toxicol. Pharmacol. 2015, 40, 828–846. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.; Tran, H.C.; Nguyen, T.C.; Pham, H.V.; Schertenleib, R.; Giger, W. Arsenic contamination of groundwater and drinking water in Vietnam: A human health threat. Environ. Sci. Technol. 2001, 35, 2621–2626. [Google Scholar] [CrossRef] [PubMed]
- George, C.M.; Sima, L.; Arias, M.H.; Mihalic, J.; Cabrera, L.Z.; Danz, D.; Checkley, W.; Gilman, R.H. Arsenic exposure in drinking water: An unrecognized health threat in Peru. Bull. World Health Organ. 2014, 92, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.H.; Lingas, E.O.; Rahman, M. Contamination of drinking-water by arsenic in Bangladesh: A public health emergency. Bull. World Health Organ. 2000, 78, 1093–1103. [Google Scholar] [PubMed]
- World Health Organization. Arsenic in Drinking-Water: Background Document for Development of WHO Guidelines for Drinking-Water Quality; World Health Organization: Geneva, Switzerland, 2003.
- Environmental Protection, A. National primary drinking water regulations: Long Term 1 Enhanced Surface Water Treatment Rule. Final rule. Fed. Regist. 2002, 67, 1811–1844. [Google Scholar]
- Beamer, P.I.; Klimecki, W.T.; Loh, M.; Van Horne, Y.O.; Sugeng, A.J.; Lothrop, N.; Billheimer, D.; Guerra, S.; Lantz, R.C.; Canales, R.A.; et al. Association of Children’s Urinary CC16 Levels with Arsenic Concentrations in Multiple Environmental Media. Int. J. Environ. Res. Public Health 2016, 13, 521. [Google Scholar] [CrossRef]
- Foust, R.D., Jr.; Mohapatra, P.; Compton-O’Brien, A.-M.; Reifel, J. Groundwater arsenic in the Verde Valley in central Arizona, USA. Appl. Geochem. 2004, 19, 251–255. [Google Scholar] [CrossRef]
- Robertson, F.N. Arsenic in ground-water under oxidizing conditions, south-west United States. Environ. Geochem. Health 1989, 11, 171–185. [Google Scholar] [CrossRef]
- ADEQ. The Status of Water Quality in Arizona: Arizona’s Integrated 305(b) Assessment and 303(d) Listing Report. In Ground Water Quality; Chapter VII; Arizona Department of Environmental Quality: Tucson, AZ, USA, 2004. [Google Scholar]
- Jones, M.C.; Credo, J.M.; Ingram, J.C.; Baldwin, J.A.; Trotter, R.T., Jr.; Propper, C.R. Arsenic Concentrations in Ground and Surface Waters across Arizona Including Native Lands. J. Contemp. Water Res. Educ. 2020, 169, 44–60. [Google Scholar] [CrossRef]
- Roh, T.; Lynch, C.F.; Weyer, P.; Wang, K.; Kelly, K.M.; Ludewig, G. Low-level arsenic exposure from drinking water is associated with prostate cancer in Iowa. Environ. Res. 2017, 159, 338–343. [Google Scholar] [CrossRef]
- Ettinger, A.S.; Zota, A.R.; Amarasiriwardena, C.J.; Hopkins, M.R.; Schwartz, J.; Hu, H.; Wright, R.O. Maternal arsenic exposure and impaired glucose tolerance during pregnancy. Environ. Health Perspect. 2009, 117, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- James, K.A.; Marshall, J.A.; Hokanson, J.E.; Meliker, J.R.; Zerbe, G.O.; Byers, T.E. A case-cohort study examining lifetime exposure to inorganic arsenic in drinking water and diabetes mellitus. Environ. Res. 2013, 123, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Maull, E.A.; Ahsan, H.; Edwards, J.; Longnecker, M.P.; Navas-Acien, A.; Pi, J.; Silbergeld, E.K.; Styblo, M.; Tseng, C.H.; Thayer, K.A.; et al. Evaluation of the association between arsenic and diabetes: A National Toxicology Program workshop review. Environ. Health Perspect. 2012, 120, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
- Abhyankar, L.N.; Jones, M.R.; Guallar, E.; Navas-Acien, A. Arsenic exposure and hypertension: A systematic review. Environ. Health Perspect. 2012, 120, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Balakumar, P.; Kaur, J. Arsenic exposure and cardiovascular disorders: An overview. Cardiovasc. Toxicol. 2009, 9, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Karagas, M.R. Arsenic and cardiovascular disease: New evidence from the United States. Ann. Intern. Med. 2013, 159, 713–714. [Google Scholar] [CrossRef] [PubMed]
- Dahan, D.; Jude, B.A.; Lamendella, R.; Keesing, F.; Perron, G.G. Exposure to Arsenic Alters the Microbiome of Larval Zebrafish. Front. Microbiol. 2018, 9, 1323. [Google Scholar] [CrossRef]
- Gentry, P.R.; McDonald, T.B.; Sullivan, D.E.; Shipp, A.M.; Yager, J.W.; Clewell, H.J., 3rd. Analysis of genomic dose-response information on arsenic to inform key events in a mode of action for carcinogenicity. Environ. Mol. Mutagen. 2010, 51, 1–14. [Google Scholar] [CrossRef]
- Chen, Y.; Graziano, J.H.; Parvez, F.; Hussain, I.; Momotaj, H.; van Geen, A.; Howe, G.R.; Ahsan, H. Modification of risk of arsenic-induced skin lesions by sunlight exposure, smoking, and occupational exposures in Bangladesh. Epidemiology 2006, 17, 459–467. [Google Scholar] [CrossRef]
- Li, Y.; Jing, D.; Xiao, Y.; Huang, X.; Shen, M. Patient-Reported Outcomes of Arsenic-Related Skin Lesions in China. Biomed. Res. Int. 2020, 2020, 6195975. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Zhang, H.; Tao, S. A review of arsenic exposure and lung cancer. Toxicol. Res. 2019, 8, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Chayapong, J.; Madhyastha, H.; Madhyastha, R.; Nurrahmah, Q.I.; Nakajima, Y.; Choijookhuu, N.; Hishikawa, Y.; Maruyama, M. Arsenic trioxide induces ROS activity and DNA damage, leading to G0/G1 extension in skin fibroblasts through the ATM-ATR-associated Chk pathway. Environ. Sci. Pollut. Res. Int. 2017, 24, 5316–5325. [Google Scholar] [CrossRef] [PubMed]
- Pinto, B.I.; Lujan, O.R.; Ramos, S.A.; Propper, C.R.; Kellar, R.S. Estrogen Mitigates the Negative Effects of Arsenic Contamination in an In Vitro Wound Model. Appl. In Vitro Toxicol. 2018, 4, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Madhyastha, H.; Madhyastha, R.; Choijookhuu, N.; Hishikawa, Y.; Pengjam, Y.; Nakajima, Y.; Maruyama, M. Arsenic acid inhibits proliferation of skin fibroblasts, and increases cellular senescence through ROS mediated MST1-FOXO signaling pathway. J. Toxicol. Sci. 2016, 41, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Horng, H.C.; Chang, W.H.; Yeh, C.C.; Huang, B.S.; Chang, C.P.; Chen, Y.J.; Tsui, K.H.; Wang, P.H. Estrogen Effects on Wound Healing. Int. J. Mol. Sci. 2017, 18, 2325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Fu, Y.; Ju, J.; Wan, D.; Su, H.; Wang, Z.; Rui, H.; Jin, Q.; Le, Y.; Hou, R. Estradiol inhibits fMLP-induced neutrophil migration and superoxide production by upregulating MKP-2 and dephosphorylating ERK. Int. Immunopharmacol. 2019, 75, 105787. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mukherjee, T.K.; Guptasarma, P. Arsenic and 17-beta-estradiol bind to each other and neutralize each other’s signaling effects. Biochem. Biophys. Res. Commun. 2016, 477, 575–580. [Google Scholar] [CrossRef]
- Davey, J.C.; Nomikos, A.P.; Wungjiranirun, M.; Sherman, J.R.; Ingram, L.; Batki, C.; Lariviere, J.P.; Hamilton, J.W. Arsenic as an endocrine disruptor: Arsenic disrupts retinoic acid receptor-and thyroid hormone receptor-mediated gene regulation and thyroid hormone-mediated amphibian tail metamorphosis. Environ. Health Perspect. 2008, 116, 165–172. [Google Scholar] [CrossRef]
- Wall, I.B.; Moseley, R.; Baird, D.M.; Kipling, D.; Giles, P.; Laffafian, I.; Price, P.E.; Thomas, D.W.; Stephens, P. Fibroblast dysfunction is a key factor in the non-healing of chronic venous leg ulcers. J. Investig. Dermatol. 2008, 128, 2526–2549. [Google Scholar] [CrossRef]
- Maione, A.G.; Smith, A.; Kashpur, O.; Yanez, V.; Knight, E.; Mooney, D.J.; Veves, A.; Tomic-Canic, M.; Garlick, J.A. Altered ECM deposition by diabetic foot ulcer-derived fibroblasts implicates fibronectin in chronic wound repair. Wound Repair. Regen. 2016, 24, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Li, Y.; Liu, L.; Qi, K.; Zhang, C.; Chen, Y.; Fang, J. Arsenic trioxide induces cell cycle arrest and affects Trk receptor expression in human neuroblastoma SK-N-SH cells. Biol. Res. 2018, 51, 18. [Google Scholar] [CrossRef] [PubMed]
- Delnomdedieu, M.; Basti, M.M.; Otvos, J.D.; Thomas, D.J. Transfer of arsenite from glutathione to dithiols: A model of interaction. Chem. Res. Toxicol. 1993, 6, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Huang, S.; Martin, S.; Wise, J.P., Sr. Arsenic is cytotoxic and genotoxic to primary human lung cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 760, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Dodson, M.; de la Vega, M.R.; Harder, B.; Castro-Portuguez, R.; Rodrigues, S.D.; Wong, P.K.; Chapman, E.; Zhang, D.D. Low-level arsenic causes proteotoxic stress and not oxidative stress. Toxicol. Appl. Pharmacol. 2018, 341, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lou, B.; Wu, R.; Wang, G.; Lu, C.; Wang, H.; Pi, J.; Xu, Y. The Role of Reactive Oxygen Species in Arsenic Toxicity. Biomolecules 2020, 10, 240. [Google Scholar]
- Liu, S.; Wang, F.; Liu, J.; Jin, P.; Wang, X.; Yang, L.; Xi, S. ATF2 partly mediated the expressions of proliferative factors and inhibited pro-inflammatory factors’ secretion in arsenite-treated human uroepithelial cells. Toxicol. Res. 2017, 6, 468–476. [Google Scholar] [CrossRef]
- Pilcher, B.K.; Dumin, J.A.; Sudbeck, B.D.; Krane, S.M.; Welgus, H.G.; Parks, W.C. The activity of collagenase-1 is required for keratinocyte migration on a type I collagen matrix. J. Cell Biol. 1997, 137, 1445–1457. [Google Scholar] [CrossRef]
- Medina, A.; Scott, P.G.; Ghahary, A.; Tredget, E.E. Pathophysiology of chronic nonhealing wounds. J. Burn. Care Rehabil. 2005, 26, 306–319. [Google Scholar] [CrossRef]
- Berman, B.; Bieley, H.C. Keloids. J. Am. Acad. Dermatol. 1995, 33, 117–123. [Google Scholar] [CrossRef]
- Barone, E.J.; Yager, D.R.; Pozez, A.L.; Olutoye, O.O.; Crossland, M.C.; Diegelmann, R.F.; Cohen, K.I. Interleukin 1 alpha and collagenase activity are elevated in chronic wounds. Plast. Reconstr. Surg. 1998, 102, 1028–1029. [Google Scholar] [CrossRef]
- Lobmann, R.; Ambrosch, A.; Schultz, G.; Waldmann, K.; Schiweck, S.; Lehnert, H. Expression of matrix-metalloproteinases and their inhibitors in the wounds of diabetic and non-diabetic patients. Diabetologia 2002, 45, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Muhetaer, M.; Yang, M.; Xia, R.; Lai, Y.; Wu, J. Gender difference in arsenic biotransformation is an important metabolic basis for arsenic toxicity. BMC Pharmacol. Toxicol. 2022, 23, 15. [Google Scholar] [CrossRef] [PubMed]
- Tordoff, M.G.; Bachmanov, A.A.; Reed, D.R. Forty mouse strain survey of water and sodium intake. Physiol. Behav. 2007, 91, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Niethammer, P.; Grabher, C.; Look, A.T.; Mitchison, T.J. A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature 2009, 459, 996–999. [Google Scholar] [CrossRef] [PubMed]
- Savill, J.; Dransfield, I.; Gregory, C.; Haslett, C. A blast from the past: Clearance of apoptotic cells regulates immune responses. Nat. Rev. Immunol. 2002, 2, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Adachi, A.; Honda, T.; Egawa, G.; Kanameishi, S.; Takimoto, R.; Miyake, T.; Hossain, M.R.; Komine, M.; Ohtsuki, M.; Gunzer, M.; et al. Estradiol suppresses psoriatic inflammation in mice by regulating neutrophil and macrophage functions. J. Allergy Clin. Immunol. 2022, 150, 909–919.e8. [Google Scholar] [CrossRef]
- Davey, J.C.; Bodwell, J.E.; Gosse, J.A.; Hamilton, J.W. Arsenic as an endocrine disruptor: Effects of arsenic on estrogen receptor-mediated gene expression in vivo and in cell culture. Toxicol. Sci. 2007, 98, 75–86. [Google Scholar] [CrossRef]
- Chatterjee, A.; Chatterji, U. Arsenic abrogates the estrogen-signaling pathway in the rat uterus. Reprod. Biol. Endocrinol. 2010, 8, 80. [Google Scholar] [CrossRef]
- Treas, J.N.; Tyagi, T.; Singh, K.P. Effects of chronic exposure to arsenic and estrogen on epigenetic regulatory genes expression and epigenetic code in human prostate epithelial cells. PLoS ONE 2012, 7, e43880. [Google Scholar] [CrossRef]
- Kitchin, K.T.; Wallace, K. Arsenite binding to synthetic peptides based on the Zn finger region and the estrogen binding region of the human estrogen receptor-alpha. Toxicol. Appl. Pharmacol. 2005, 206, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, R.; Sato, S.; Okada, M.; Lee, L.; Tamura, I.; Jozaki, K.; Kajimura, T.; Asada, H.; Yamagata, Y.; Tamura, H.; et al. Tissue-Specific Expression of Estrogen Receptor 1 Is Regulated by DNA Methylation in a T-DMR. Mol. Endocrinol. 2016, 30, 335–347. [Google Scholar] [CrossRef]
- Thornton, M.J.; Taylor, A.H.; Mulligan, K.; Al-Azzawi, F.; Lyon, C.C.; O’Driscoll, J.; Messenger, A.G. The distribution of estrogen receptor beta is distinct to that of estrogen receptor alpha and the androgen receptor in human skin and the pilosebaceous unit. J. Investig. Dermatol. Symp. Proc. 2003, 8, 100–103. [Google Scholar] [CrossRef]
- Campbell, L.; Emmerson, E.; Davies, F.; Gilliver, S.C.; Krust, A.; Chambon, P.; Ashcroft, G.S.; Hardman, M.J. Estrogen promotes cutaneous wound healing via estrogen receptor beta independent of its antiinflammatory activities. J. Exp. Med. 2010, 207, 1825–1833. [Google Scholar] [CrossRef]
- Meza, M.M.; Yu, L.; Rodriguez, Y.Y.; Guild, M.; Thompson, D.; Gandolfi, A.J.; Klimecki, W.T. Developmentally restricted genetic determinants of human arsenic metabolism: Association between urinary methylated arsenic and CYT19 polymorphisms in children. Environ. Health Perspect. 2005, 113, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.C.; Douillet, C.; Su, M.; Zhou, K.; Wu, T.; Chen, W.; Galanko, J.A.; Drobna, Z.; Saunders, R.J.; Martin, E.; et al. Metabolomic profiles of arsenic (+3 oxidation state) methyltransferase knockout mice: Effect of sex and arsenic exposure. Arch. Toxicol. 2017, 91, 189–202. [Google Scholar] [CrossRef]
- Park, D.; Propper, C.R.; Wang, G.; Salanga, M.C. Synonymous single nucleotide polymorphism in arsenic (+3) methyltransferase of the Western mosquitofish (Gambusia affinis) and its gene expression among field populations. Ecotoxicology 2021, 30, 711–718. [Google Scholar] [CrossRef]
- Thomas, D.J.; Li, J.; Waters, S.B.; Xing, W.; Adair, B.M.; Drobna, Z.; Devesa, V.; Styblo, M. Arsenic (+3 oxidation state) methyltransferase and the methylation of arsenicals. Exp. Biol. Med. 2007, 232, 3–13. [Google Scholar]
- Drobna, Z.; Naranmandura, H.; Kubachka, K.M.; Edwards, B.C.; Herbin-Davis, K.; Styblo, M.; Le, X.C.; Creed, J.T.; Maeda, N.; Hughes, M.F.; et al. Disruption of the arsenic (+3 oxidation state) methyltransferase gene in the mouse alters the phenotype for methylation of arsenic and affects distribution and retention of orally administered arsenate. Chem. Res. Toxicol. 2009, 22, 1713–1720. [Google Scholar] [CrossRef]
- Valenzuela, O.L.; Drobna, Z.; Hernandez-Castellanos, E.; Sanchez-Pena, L.C.; Garcia-Vargas, G.G.; Borja-Aburto, V.H.; Styblo, M.; Del Razo, L.M. Association of AS3MT polymorphisms and the risk of premalignant arsenic skin lesions. Toxicol. Appl. Pharmacol. 2009, 239, 200–207. [Google Scholar] [CrossRef]
- Nemeti, B.; Csanaky, I.; Gregus, Z. Effect of an inactivator of glyceraldehyde-3-phosphate dehydrogenase, a fortuitous arsenate reductase, on disposition of arsenate in rats. Toxicol. Sci. 2006, 90, 49–60. [Google Scholar] [CrossRef][Green Version]
- Barton, P.J.; Birks, E.J.; Felkin, L.E.; Cullen, M.E.; Koban, M.U.; Yacoub, M.H. Increased expression of extracellular matrix regulators TIMP1 and MMP1 in deteriorating heart failure. J. Heart Lung Transplant. 2003, 22, 738–744. [Google Scholar] [CrossRef]
- Vaalamo, M.; Weckroth, M.; Puolakkainen, P.; Kere, J.; Saarinen, P.; Lauharanta, J.; Saarialho-Kere, U.K. Patterns of matrix metalloproteinase and TIMP-1 expression in chronic and normally healing human cutaneous wounds. Br. J. Dermatol. 1996, 135, 52–59. [Google Scholar] [CrossRef]
- Masson-Meyers, D.S.; Andrade, T.A.M.; Caetano, G.F.; Guimaraes, F.R.; Leite, M.N.; Leite, S.N.; Frade, M.A.C. Experimental models and methods for cutaneous wound healing assessment. Int. J. Exp. Pathol. 2020, 101, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Podgorski, J.; Berg, M. Global threat of arsenic in groundwater. Science 2020, 368, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Hoover, J.; Gonzales, M.; Shuey, C.; Barney, Y.; Lewis, J. Elevated Arsenic and Uranium Concentrations in Unregulated Water Sources on the Navajo Nation, USA. Expo Health 2017, 9, 113–124. [Google Scholar] [CrossRef]
- Ingram, J.C.; Jones, L.; Credo, J.; Rock, T. Uranium and arsenic unregulated water issues on Navajo lands. J. Vac. Sci. Technol. A 2020, 38, 031003. [Google Scholar] [CrossRef] [PubMed]
- Ahangarpour, A.; Zeidooni, L.; Samimi, A.; Alboghobeish, S.; Khorsandi, L.S.; Moradi, M. Chronic exposure to arsenic and high fat diet additively induced cardiotoxicity in male mice. Res. Pharm. Sci. 2018, 13, 47–56. [Google Scholar] [PubMed]
- Machula, H.; Ensley, B.; Kellar, R. Electrospun Tropoelastin for Delivery of Therapeutic Adipose-Derived Stem Cells to Full-Thickness Dermal Wounds. Adv. Wound Care 2014, 3, 367–375. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Transcript | Gene Abbreviation | Primer Sequence | Annealing Temperature (°C) | Amplicon Length |
|---|---|---|---|---|
| Human | ||||
| Glyceraldehyde 3-phosphate dehydrogenase | GAPDH | F-CTCCAAAATCAAGTGGGGCGA | 60.89 | 70 |
| R-CATGGTGGTGAAGACGCCAG | 61.3 | |||
| Succinate dehydrogenase complex subunit A | SDHA | F-TGGCCCTGAGAAAGATCACG | 59.75 | 193 |
| R-GACCTGCCCCTTGTAGTTGG | 60.32 | |||
| Matrix metalloproteinase 1 | MMP1 | F-GGCCACAAAGTTGATGCAGTT | 59.93 | 137 |
| R-TTCCTGCAGTTGAACCAGCTA | 59.58 | |||
| Mouse | ||||
| Glyceraldehyde 3-phosphate dehydrogenase | GAPDH | F-CATCACTGCCACCCAGAAGACTG | 63.28 | 153 |
| R-ATGCCAGTGAGCTTCCCGTTCAG | 65.15 | |||
| Succinate dehydrogenase complex subunit A | SDHA | F-GAGATACGCACCTGTTGCCAAG | 61.82 | 113 |
| R-GGTAGACGTGATCTTTCTCAGGG | 60.18 | |||
| Matrix metalloproteinase 1a | MMP1a | F-AGGAAGGCGATATTGTGCTCTCC | 62.37 | 98 |
| R-TGGCTGGAAAGTGTGTGAGCAAGC | 63.85 | |||
| TIMP metallopeptidase inhibitor 1 | TIMP1 | F-TCTTGGTTCCCTGGCGTACTCT | 63.08 | 131 |
| R-GTGAGTGTCACTCTCCAGTTTGC | 61.59 | |||
| Arsenic 3-methyltransferase | AS3MT | F-TCCACGTTTGGTCACTGCCGAT | 64.71 | 100 |
| R-GAAGAGGCGAAATGTGGCAGAC | 62.07 | |||
| G-Protein coupled estrogen receptor 1 | GPER1 | F-GCCACATAGTCAACCTTGCAGC | 62.34 | 113 |
| R-CGTCTTCTGCTCCACATAGAGC | 60.8 | |||
| Estrogen receptor 1 | ESR1 | F-TCTGCCAAGGAGACTCGCTACT | 62.86 | 153 |
| R-GGTGCATTGGTTTGTAGCTGGAC | 62.46 | |||
| Estrogen receptor 2 | ESR2 | F-GGTCCTGTGAAGGATGTAAGGC | 60.68 | 139 |
| R-TAACACTTGCGAAGTCGGCAGG | 63.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dresler, S.R.; Pinto, B.I.; Salanga, M.C.; Propper, C.R.; Berry, S.R.; Kellar, R.S. Arsenic Impairs Wound Healing Processes in Dermal Fibroblasts and Mice. Int. J. Mol. Sci. 2024, 25, 2161. https://doi.org/10.3390/ijms25042161
Dresler SR, Pinto BI, Salanga MC, Propper CR, Berry SR, Kellar RS. Arsenic Impairs Wound Healing Processes in Dermal Fibroblasts and Mice. International Journal of Molecular Sciences. 2024; 25(4):2161. https://doi.org/10.3390/ijms25042161
Chicago/Turabian StyleDresler, Sara R., Bronson I. Pinto, Matthew C. Salanga, Catherine R. Propper, Savannah R. Berry, and Robert S. Kellar. 2024. "Arsenic Impairs Wound Healing Processes in Dermal Fibroblasts and Mice" International Journal of Molecular Sciences 25, no. 4: 2161. https://doi.org/10.3390/ijms25042161
APA StyleDresler, S. R., Pinto, B. I., Salanga, M. C., Propper, C. R., Berry, S. R., & Kellar, R. S. (2024). Arsenic Impairs Wound Healing Processes in Dermal Fibroblasts and Mice. International Journal of Molecular Sciences, 25(4), 2161. https://doi.org/10.3390/ijms25042161