

Exploring the Molecular Tumor Microenvironment and Translational Biomarkers in Brain Metastases of Non-Small-Cell Lung Cancer


Abstract
1. Introduction
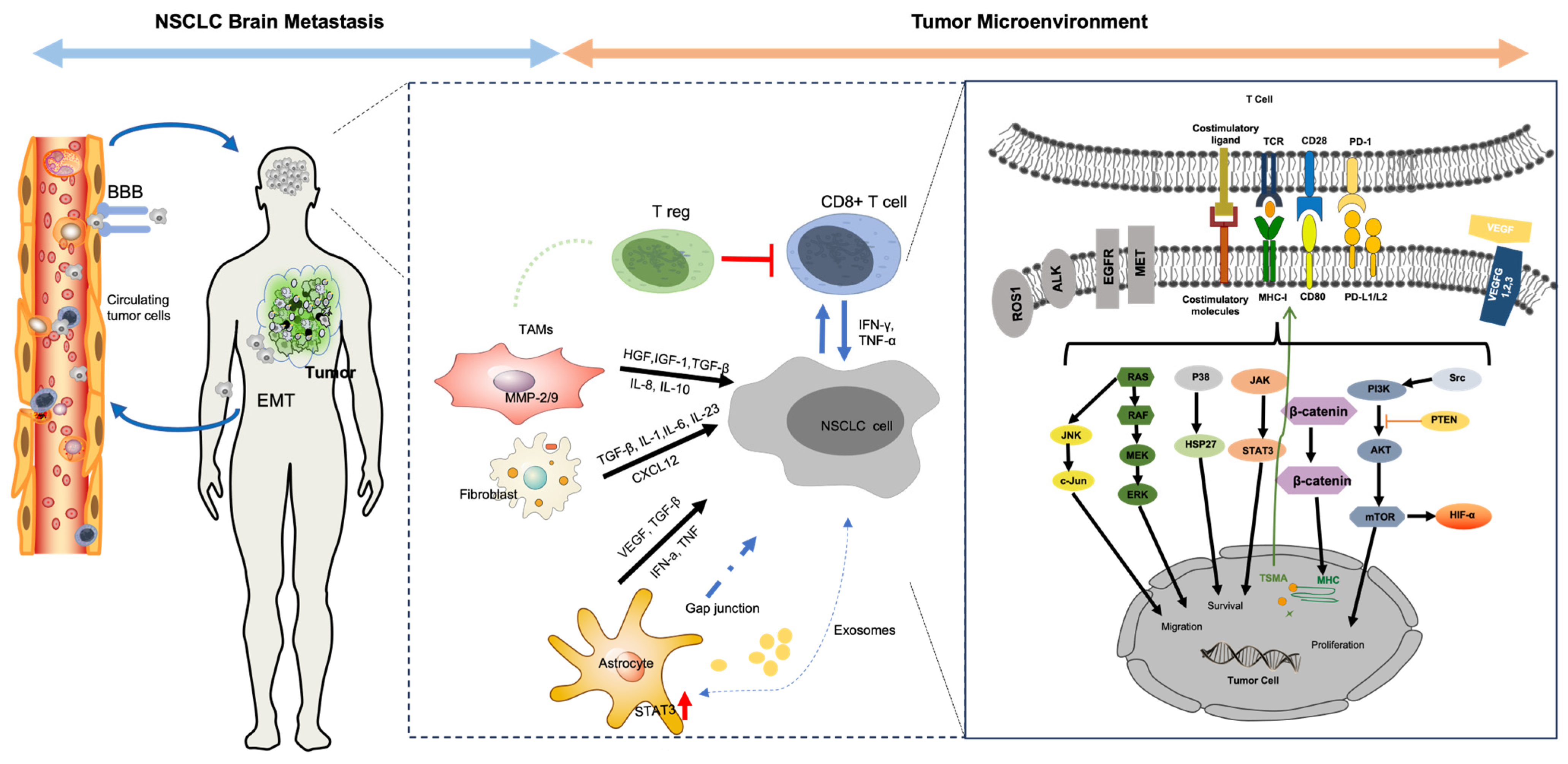
2. Tumor Microenvironment in Brain Metastases of NSCLC
2.1. Overview of TME in Brain Metastases
2.2. Cellular Components
2.2.1. T Cells
2.2.2. Cancer-Associated Fibroblasts
2.2.3. Tumor-Associated Macrophages
2.2.4. Astrocytes
2.3. Molecular Landscape of Brain Metastases
3. Translational Biomarkers in NSCLC Brain Metastases
3.1. Tissue-Based Molecular Biomarkers
3.2. Liquid Biopsy Biomarkers: Circulating Tumor Cells and ctDNA
3.3. Exosomes and Non-Code RNA
3.4. Imaging Biomarkers
3.5. Immune Cells and Serological Biomarkers
3.6. Microbiota
4. Conclusions and Future Therapeutic Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schabath, M.B.; Cote, M.L. Cancer Progress and Priorities: Lung Cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1563–1579. [Google Scholar] [CrossRef]
- El Rassy, E.; Botticella, A.; Kattan, J.; Le Pechoux, C.; Besse, B.; Hendriks, L. Non-small cell lung cancer brain metastases and the immune system: From brain metastases development to treatment. Cancer Treat. Rev. 2018, 68, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Achrol, A.S.; Rennert, R.C.; Anders, C.; Soffietti, R.; Ahluwalia, M.S.; Nayak, L.; Peters, S.; Arvold, N.D.; Harsh, G.R.; Steeg, P.S.; et al. Brain metastases. Nat. Rev. Dis. Primers 2019, 5, 5. [Google Scholar] [CrossRef]
- Getman, V.; Devyatko, E.; Dunkler, D.; Eckersberger, F.; End, A.; Klepetko, W.; Marta, G.; Mueller, M.R. Prognosis of patients with non-small cell lung cancer with isolated brain metastases undergoing combined surgical treatment. Eur. J. Cardiothorac. Surg. 2004, 25, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Soffietti, R.; Ahluwalia, M.; Lin, N.; Ruda, R. Management of brain metastases according to molecular subtypes. Nat. Rev. Neurol. 2020, 16, 557–574. [Google Scholar] [CrossRef] [PubMed]
- Sperduto, P.W.; Yang, T.J.; Beal, K.; Pan, H.; Brown, P.D.; Bangdiwala, A.; Shanley, R.; Yeh, N.; Gaspar, L.E.; Braunstein, S.; et al. Estimating Survival in Patients with Lung Cancer and Brain Metastases: An Update of the Graded Prognostic Assessment for Lung Cancer Using Molecular Markers (Lung-molGPA). JAMA Oncol. 2017, 3, 827–831. [Google Scholar] [CrossRef]
- Lin, A.; Wei, T.; Meng, H.; Luo, P.; Zhang, J. Role of the dynamic tumor microenvironment in controversies regarding immune checkpoint inhibitors for the treatment of non-small cell lung cancer (NSCLC) with EGFR mutations. Mol. Cancer 2019, 18, 139. [Google Scholar] [CrossRef]
- Altorki, N.K.; Markowitz, G.J.; Gao, D.; Port, J.L.; Saxena, A.; Stiles, B.; McGraw, T.; Mittal, V. The lung microenvironment: An important regulator of tumour growth and metastasis. Nat. Rev. Cancer 2019, 19, 9–31. [Google Scholar] [CrossRef]
- Neophytou, C.M.; Panagi, M.; Stylianopoulos, T.; Papageorgis, P. The Role of Tumor Microenvironment in Cancer Metastasis: Molecular Mechanisms and Therapeutic Opportunities. Cancers 2021, 13, 53. [Google Scholar] [CrossRef]
- Schulz, M.; Salamero-Boix, A.; Niesel, K.; Alekseeva, T.; Sevenich, L. Microenvironmental Regulation of Tumor Progression and Therapeutic Response in Brain Metastasis. Front. Immunol. 2019, 10, 1713. [Google Scholar] [CrossRef]
- Mo, F.; Pellerino, A.; Soffietti, R.; Ruda, R. Blood-Brain Barrier in Brain Tumors: Biology and Clinical Relevance. Int. J. Mol. Sci. 2021, 22, 12654. [Google Scholar] [CrossRef]
- Lin, X.; DeAngelis, L.M. Treatment of Brain Metastases. J. Clin. Oncol. 2015, 33, 3475–3484. [Google Scholar] [CrossRef]
- Srinivasan, E.S.; Deshpande, K.; Neman, J.; Winkler, F.; Khasraw, M. The microenvironment of brain metastases from solid tumors. Neurooncol. Adv. 2021, 3, v121–v132. [Google Scholar] [CrossRef]
- Guan, Z.; Lan, H.; Cai, X.; Zhang, Y.; Liang, A.; Li, J. Blood-Brain Barrier, Cell Junctions, and Tumor Microenvironment in Brain Metastases, the Biological Prospects and Dilemma in Therapies. Front. Cell Dev. Biol. 2021, 9, 722917. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, R.; Wa, Y.; Ding, S.; Yang, Y.; Liao, J.; Tong, L.; Xiao, G. Tumor Immune Microenvironment and Immunotherapy in Brain Metastasis From Non-Small Cell Lung Cancer. Front. Immunol. 2022, 13, 829451. [Google Scholar] [CrossRef]
- Wilhelm, I.; Molnar, J.; Fazakas, C.; Hasko, J.; Krizbai, I.A. Role of the blood-brain barrier in the formation of brain metastases. Int. J. Mol. Sci. 2013, 14, 1383–1411. [Google Scholar] [CrossRef]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Caffarel, M.M.; Braza, M.S. Microglia and metastases to the central nervous system: Victim, ravager, or something else? J. Exp. Clin. Cancer Res. 2022, 41, 327. [Google Scholar] [CrossRef] [PubMed]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, D.; Okimoto, T.; Shukuya, T.; Onagi, H.; Hayashi, T.; Sinicropi-Yao, S.L.; Amann, J.M.; Nakatsura, T.; Kitano, S.; Carbone, D.P. Comparison of Tumor Microenvironments between Primary Tumors and Brain Metastases in Patients with NSCLC. JTO Clin. Res. Rep. 2021, 2, 100230. [Google Scholar] [CrossRef] [PubMed]
- Porciello, N.; Franzese, O.; D’Ambrosio, L.; Palermo, B.; Nistico, P. T-cell repertoire diversity: Friend or foe for protective antitumor response? J. Exp. Clin. Cancer Res. 2022, 41, 356. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed]
- Henke, E.; Nandigama, R.; Ergun, S. Extracellular Matrix in the Tumor Microenvironment and Its Impact on Cancer Therapy. Front. Mol. Biosci. 2019, 6, 160. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Li, L.; Tanzhu, G.; Liu, Z.; Gao, X.; Wan, X.; Xiao, D.; Chen, L.; Xia, X.; Zhou, R. Heterogeneity of tumor immune microenvironment of EGFR/ALK-positive tumors versus EGFR/ALK-negative tumors in resected brain metastases from lung adenocarcinoma. J. Immunother. Cancer 2023, 11, e006243. [Google Scholar] [CrossRef] [PubMed]
- Vilarino, N.; Bruna, J.; Bosch-Barrera, J.; Valiente, M.; Nadal, E. Immunotherapy in NSCLC patients with brain metastases. Understanding brain tumor microenvironment and dissecting outcomes from immune checkpoint blockade in the clinic. Cancer Treat. Rev. 2020, 89, 102067. [Google Scholar] [CrossRef]
- Kudo, Y.; Haymaker, C.; Zhang, J.; Reuben, A.; Duose, D.Y.; Fujimoto, J.; Roy-Chowdhuri, S.; Solis Soto, L.M.; Dejima, H.; Parra, E.R.; et al. Suppressed immune microenvironment and repertoire in brain metastases from patients with resected non-small-cell lung cancer. Ann. Oncol. 2019, 30, 1521–1530. [Google Scholar] [CrossRef]
- Song, S.G.; Kim, S.; Koh, J.; Yim, J.; Han, B.; Kim, Y.A.; Jeon, Y.K.; Chung, D.H. Comparative analysis of the tumor immune-microenvironment of primary and brain metastases of non-small-cell lung cancer reveals organ-specific and EGFR mutation-dependent unique immune landscape. Cancer Immunol. Immunother. 2021, 70, 2035–2048. [Google Scholar] [CrossRef]
- Zhang, Q.; Abdo, R.; Iosef, C.; Kaneko, T.; Cecchini, M.; Han, V.K.; Li, S.S. The spatial transcriptomic landscape of non-small cell lung cancer brain metastasis. Nat. Commun. 2022, 13, 5983. [Google Scholar] [CrossRef]
- Karimi, E.; Yu, M.W.; Maritan, S.M.; Perus, L.J.M.; Rezanejad, M.; Sorin, M.; Dankner, M.; Fallah, P.; Dore, S.; Zuo, D.; et al. Single-cell spatial immune landscapes of primary and metastatic brain tumours. Nature 2023, 614, 555–563. [Google Scholar] [CrossRef]
- Huppert, L.A.; Green, M.D.; Kim, L.; Chow, C.; Leyfman, Y.; Daud, A.I.; Lee, J.C. Tissue-specific Tregs in cancer metastasis: Opportunities for precision immunotherapy. Cell Mol. Immunol. 2022, 19, 33–45. [Google Scholar] [CrossRef]
- Li, Y.D.; Lamano, J.B.; Lamano, J.B.; Quaggin-Smith, J.; Veliceasa, D.; Kaur, G.; Biyashev, D.; Unruh, D.; Bloch, O. Tumor-induced peripheral immunosuppression promotes brain metastasis in patients with non-small cell lung cancer. Cancer Immunol. Immunother. 2019, 68, 1501–1513. [Google Scholar] [CrossRef]
- Wainwright, D.A.; Sengupta, S.; Han, Y.; Lesniak, M.S. Thymus-derived rather than tumor-induced regulatory T cells predominate in brain tumors. Neuro Oncol. 2011, 13, 1308–1323. [Google Scholar] [CrossRef]
- Tanaka, A.; Sakaguchi, S. Regulatory T cells in cancer immunotherapy. Cell Res. 2017, 27, 109–118. [Google Scholar] [CrossRef]
- Dykema, A.G.; Zhang, J.; Cheung, L.S.; Connor, S.; Zhang, B.; Zeng, Z.; Cherry, C.M.; Li, T.; Caushi, J.X.; Nishimoto, M.; et al. Lung tumor-infiltrating T(reg) have divergent transcriptional profiles and function linked to checkpoint blockade response. Sci. Immunol. 2023, 8, eadg1487. [Google Scholar] [CrossRef]
- Wischnewski, V.; Maas, R.R.; Aruffo, P.G.; Soukup, K.; Galletti, G.; Kornete, M.; Galland, S.; Fournier, N.; Lilja, J.; Wirapati, P.; et al. Phenotypic diversity of T cells in human primary and metastatic brain tumors revealed by multiomic interrogation. Nat. Cancer 2023, 4, 908–924. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, J.; Huang, H.; Ye, M.; Li, X.; Wu, R.; Liu, H.; Song, Y. Metastasis-associated fibroblasts: An emerging target for metastatic cancer. Biomark. Res. 2021, 9, 47. [Google Scholar] [CrossRef]
- Liu, W.; Powell, C.A.; Wang, Q. Tumor microenvironment in lung cancer-derived brain metastasis. Chin. Med. J. (Engl.) 2022, 135, 1781–1791. [Google Scholar] [CrossRef]
- Zhang, H.; Yue, X.; Chen, Z.; Liu, C.; Wu, W.; Zhang, N.; Liu, Z.; Yang, L.; Jiang, Q.; Cheng, Q.; et al. Define cancer-associated fibroblasts (CAFs) in the tumor microenvironment: New opportunities in cancer immunotherapy and advances in clinical trials. Mol. Cancer 2023, 22, 159. [Google Scholar] [CrossRef]
- Ghorani, E.; Swanton, C.; Quezada, S.A. Cancer cell-intrinsic mechanisms driving acquired immune tolerance. Immunity 2023, 56, 2270–2295. [Google Scholar] [CrossRef]
- Hu, H.; Piotrowska, Z.; Hare, P.J.; Chen, H.; Mulvey, H.E.; Mayfield, A.; Noeen, S.; Kattermann, K.; Greenberg, M.; Williams, A.; et al. Three subtypes of lung cancer fibroblasts define distinct therapeutic paradigms. Cancer Cell 2021, 39, 1531–1547.e10. [Google Scholar] [CrossRef]
- Akanda, M.R.; Ahn, E.J.; Kim, Y.J.; Salam, S.M.A.; Noh, M.G.; Kim, S.S.; Jung, T.Y.; Kim, I.Y.; Kim, C.H.; Lee, K.H.; et al. Different Expression and Clinical Implications of Cancer-Associated Fibroblast (CAF) Markers in Brain Metastases. J. Cancer 2023, 14, 464–479. [Google Scholar] [CrossRef]
- Kim, H.J.; Yang, K.; Kim, K.; Lee, Y.J.; Lee, S.; Ahn, S.Y.; Ahn, Y.H.; Kang, J.L. Reprogramming of cancer-associated fibroblasts by apoptotic cancer cells inhibits lung metastasis via Notch1-WISP-1 signaling. Cell Mol. Immunol. 2022, 19, 1373–1391. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, K.; Qiu, L.; Yue, J.; Jiang, H.; Deng, Q.; Zhou, R.; Yin, Z.; Ma, S.; Ke, Y. Cancer-associated fibroblasts facilitate DNA damage repair by promoting the glycolysis in non-small cell lung cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166670. [Google Scholar] [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-associated macrophages in tumor metastasis: Biological roles and clinical therapeutic applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- Xu, W.; Patel, N.; Deng, Y.; Ding, S.; Wang, T.; Zhang, H. Extracellular vesicle-derived LINC00482 induces microglial M2 polarization to facilitate brain metastasis of NSCLC. Cancer Lett. 2023, 561, 216146. [Google Scholar] [CrossRef]
- Li, X.; Chen, Z.; Ni, Y.; Bian, C.; Huang, J.; Chen, L.; Xie, X.; Wang, J. Tumor-associated macrophages secret exosomal miR-155 and miR-196a-5p to promote metastasis of non-small-cell lung cancer. Transl. Lung Cancer Res. 2021, 10, 1338–1354. [Google Scholar] [CrossRef]
- Wei, K.; Ma, Z.; Yang, F.; Zhao, X.; Jiang, W.; Pan, C.; Li, Z.; Pan, X.; He, Z.; Xu, J.; et al. M2 macrophage-derived exosomes promote lung adenocarcinoma progression by delivering miR-942. Cancer Lett. 2022, 526, 205–216. [Google Scholar] [CrossRef]
- Tiong, T.Y.; Chan, M.L.; Wang, C.H.; Yadav, V.K.; Pikatan, N.W.; Fong, I.H.; Yeh, C.T.; Kuo, K.T.; Huang, W.C. Exosomal miR-21 determines lung-to-brain metastasis specificity through the DGKB/ERK axis within the tumor microenvironment. Life Sci. 2023, 329, 121945. [Google Scholar] [CrossRef]
- Jin, Y.; Kang, Y.; Wang, M.; Wu, B.; Su, B.; Yin, H.; Tang, Y.; Li, Q.; Wei, W.; Mei, Q.; et al. Targeting polarized phenotype of microglia via IL6/JAK2/STAT3 signaling to reduce NSCLC brain metastasis. Signal Transduct. Target. Ther. 2022, 7, 52. [Google Scholar] [CrossRef]
- Croci, D.; Santalla Mendez, R.; Temme, S.; Soukup, K.; Fournier, N.; Zomer, A.; Colotti, R.; Wischnewski, V.; Flogel, U.; van Heeswijk, R.B.; et al. Multispectral fluorine-19 MRI enables longitudinal and noninvasive monitoring of tumor-associated macrophages. Sci. Transl. Med. 2022, 14, eabo2952. [Google Scholar] [CrossRef]
- Wasilewski, D.; Priego, N.; Fustero-Torre, C.; Valiente, M. Reactive Astrocytes in Brain Metastasis. Front. Oncol. 2017, 7, 298. [Google Scholar] [CrossRef]
- Adler, O.; Zait, Y.; Cohen, N.; Blazquez, R.; Doron, H.; Monteran, L.; Scharff, Y.; Shami, T.; Mundhe, D.; Glehr, G.; et al. Reciprocal interactions between innate immune cells and astrocytes facilitate neuroinflammation and brain metastasis via lipocalin-2. Nat. Cancer 2023, 4, 401–418. [Google Scholar] [CrossRef]
- Chen, Q.; Boire, A.; Jin, X.; Valiente, M.; Er, E.E.; Lopez-Soto, A.; Jacob, L.; Patwa, R.; Shah, H.; Xu, K.; et al. Carcinoma-astrocyte gap junctions promote brain metastasis by cGAMP transfer. Nature 2016, 533, 493–498. [Google Scholar] [CrossRef]
- Tamai, S.; Ichinose, T.; Tsutsui, T.; Tanaka, S.; Garaeva, F.; Sabit, H.; Nakada, M. Tumor Microenvironment in Glioma Invasion. Brain Sci. 2022, 12, 505. [Google Scholar] [CrossRef]
- Ma, W.; Oliveira-Nunes, M.C.; Xu, K.; Kossenkov, A.; Reiner, B.C.; Crist, R.C.; Hayden, J.; Chen, Q. Type I interferon response in astrocytes promotes brain metastasis by enhancing monocytic myeloid cell recruitment. Nat. Commun. 2023, 14, 2632. [Google Scholar] [CrossRef]
- Ye, L.; Wu, Y.; Zhou, J.; Xie, M.; Zhang, Z.; Su, C. Influence of Exosomes on Astrocytes in the Pre-Metastatic Niche of Lung Cancer Brain Metastases. Biol. Proced. Online 2023, 25, 5. [Google Scholar] [CrossRef]
- Priego, N.; Zhu, L.; Monteiro, C.; Mulders, M.; Wasilewski, D.; Bindeman, W.; Doglio, L.; Martinez, L.; Martinez-Saez, E.; Ramon, Y.C.S.; et al. STAT3 labels a subpopulation of reactive astrocytes required for brain metastasis. Nat. Med. 2018, 24, 1024–1035. [Google Scholar] [CrossRef]
- Pahlavan, Y.; Mohammadi Nasr, M.; Dalir Abdolahinia, E.; Pirdel, Z.; Razi Soofiyani, S.; Siahpoush, S.; Nejati, K. Prominent roles of microRNA-142 in cancer. Pathol. Res. Pract. 2020, 216, 153220. [Google Scholar] [CrossRef]
- Shin, D.Y.; Na, I., II; Kim, C.H.; Park, S.; Baek, H.; Yang, S.H. EGFR mutation and brain metastasis in pulmonary adenocarcinomas. J. Thorac. Oncol. 2014, 9, 195–199. [Google Scholar] [CrossRef]
- Johung, K.L.; Yeh, N.; Desai, N.B.; Williams, T.M.; Lautenschlaeger, T.; Arvold, N.D.; Ning, M.S.; Attia, A.; Lovly, C.M.; Goldberg, S.; et al. Extended Survival and Prognostic Factors for Patients with ALK-Rearranged Non-Small-Cell Lung Cancer and Brain Metastasis. J. Clin. Oncol. 2016, 34, 123–129. [Google Scholar] [CrossRef]
- Huang, R.S.P.; Harries, L.; Decker, B.; Hiemenz, M.C.; Murugesan, K.; Creeden, J.; Tolba, K.; Stabile, L.P.; Ramkissoon, S.H.; Burns, T.F.; et al. Clinicopathologic and Genomic Landscape of Non-Small Cell Lung Cancer Brain Metastases. Oncologist 2022, 27, 839–848. [Google Scholar] [CrossRef]
- Shih, D.J.H.; Nayyar, N.; Bihun, I.; Dagogo-Jack, I.; Gill, C.M.; Aquilanti, E.; Bertalan, M.; Kaplan, A.; D’Andrea, M.R.; Chukwueke, U.; et al. Genomic characterization of human brain metastases identifies drivers of metastatic lung adenocarcinoma. Nat. Genet. 2020, 52, 371–377. [Google Scholar] [CrossRef]
- Paik, P.K.; Shen, R.; Won, H.; Rekhtman, N.; Wang, L.; Sima, C.S.; Arora, A.; Seshan, V.; Ladanyi, M.; Berger, M.F.; et al. Next-Generation Sequencing of Stage IV Squamous Cell Lung Cancers Reveals an Association of PI3K Aberrations and Evidence of Clonal Heterogeneity in Patients with Brain Metastases. Cancer Discov. 2015, 5, 610–621. [Google Scholar] [CrossRef]
- Brastianos, P.K.; Carter, S.L.; Santagata, S.; Cahill, D.P.; Taylor-Weiner, A.; Jones, R.T.; Van Allen, E.M.; Lawrence, M.S.; Horowitz, P.M.; Cibulskis, K.; et al. Genomic Characterization of Brain Metastases Reveals Branched Evolution and Potential Therapeutic Targets. Cancer Discov. 2015, 5, 1164–1177. [Google Scholar] [CrossRef]
- Vidotto, T.; Melo, C.M.; Lautert-Dutra, W.; Chaves, L.P.; Reis, R.B.; Squire, J.A. Pan-cancer genomic analysis shows hemizygous PTEN loss tumors are associated with immune evasion and poor outcome. Sci. Rep. 2023, 13, 5049. [Google Scholar] [CrossRef]
- Wilson, G.D.; Johnson, M.D.; Ahmed, S.; Cardenas, P.Y.; Grills, I.S.; Thibodeau, B.J. Targeted DNA sequencing of non-small cell lung cancer identifies mutations associated with brain metastases. Oncotarget 2018, 9, 25957–25970. [Google Scholar] [CrossRef]
- Wang, H.; Ou, Q.; Li, D.; Qin, T.; Bao, H.; Hou, X.; Wang, K.; Wang, F.; Deng, Q.; Liang, J.; et al. Genes associated with increased brain metastasis risk in non-small cell lung cancer: Comprehensive genomic profiling of 61 resected brain metastases versus primary non-small cell lung cancer (Guangdong Association Study of Thoracic Oncology 1036). Cancer 2019, 125, 3535–3544. [Google Scholar] [CrossRef]
- Fu, F.; Zhang, Y.; Gao, Z.; Zhao, Y.; Wen, Z.; Han, H.; Li, Y.; Chen, H. Development and validation of a five-gene model to predict postoperative brain metastasis in operable lung adenocarcinoma. Int. J. Cancer 2020, 147, 584–592. [Google Scholar] [CrossRef]
- Zhang, Z.; Cui, F.; Zhou, M.; Wu, S.; Zou, Q.; Gao, B. Single-cell RNA Sequencing Analysis Identifies Key Genes in Brain Metastasis from Lung Adenocarcinoma. Curr. Gene Ther. 2021, 21, 338–348. [Google Scholar] [CrossRef]
- Chatterjee, A.; Rodger, E.J.; Eccles, M.R. Epigenetic drivers of tumourigenesis and cancer metastasis. Semin. Cancer Biol. 2018, 51, 149–159. [Google Scholar] [CrossRef]
- Karlow, J.A.; Devarakonda, S.; Xing, X.; Jang, H.S.; Govindan, R.; Watson, M.; Wang, T. Developmental Pathways Are Epigenetically Reprogrammed during Lung Cancer Brain Metastasis. Cancer Res. 2022, 82, 2692–2703. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, P.; Wei, J.; Zhang, X.; Guo, J.; Lin, Y. Recent progress in targeted therapy for non-small cell lung cancer. Front. Pharmacol. 2023, 14, 1125547. [Google Scholar] [CrossRef]
- Ettinger, D.S.; Wood, D.E.; Aisner, D.L.; Akerley, W.; Bauman, J.R.; Bharat, A.; Bruno, D.S.; Chang, J.Y.; Chirieac, L.R.; D’Amico, T.A.; et al. Non-Small Cell Lung Cancer, Version 3.2022, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2022, 20, 497–530. [Google Scholar] [CrossRef]
- Ha, S.Y.; Choi, S.J.; Cho, J.H.; Choi, H.J.; Lee, J.; Jung, K.; Irwin, D.; Liu, X.; Lira, M.E.; Mao, M.; et al. Lung cancer in never-smoker Asian females is driven by oncogenic mutations, most often involving EGFR. Oncotarget 2015, 6, 5465–5474. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Z.; Zhang, G.; Zhang, M.; Zhang, X.; Li, H.; Zheng, X.; Ma, Z. Driver genes as predictive indicators of brain metastasis in patients with advanced NSCLC: EGFR, ALK, and RET gene mutations. Cancer Med. 2020, 9, 487–495. [Google Scholar] [CrossRef]
- Lamberti, G.; Andrini, E.; Sisi, M.; Rizzo, A.; Parisi, C.; Di Federico, A.; Gelsomino, F.; Ardizzoni, A. Beyond EGFR, ALK and ROS1: Current evidence and future perspectives on newly targetable oncogenic drivers in lung adenocarcinoma. Crit. Rev. Oncol. Hematol. 2020, 156, 103119. [Google Scholar] [CrossRef]
- Remon, J.; Steuer, C.E.; Ramalingam, S.S.; Felip, E. Osimertinib and other third-generation EGFR TKI in EGFR-mutant NSCLC patients. Ann. Oncol. 2018, 29, i20–i27. [Google Scholar] [CrossRef]
- Ceddia, S.; Codacci-Pisanelli, G. Treatment of brain metastases in ALK-positive non-small cell lung cancer. Crit. Rev. Oncol. Hematol. 2021, 165, 103400. [Google Scholar] [CrossRef]
- Drilon, A.; Jenkins, C.; Iyer, S.; Schoenfeld, A.; Keddy, C.; Davare, M.A. ROS1-dependent cancers—Biology, diagnostics and therapeutics. Nat. Rev. Clin. Oncol. 2021, 18, 35–55. [Google Scholar] [CrossRef]
- Liu, X.; Li, G.; Zhang, H.; Chang, Q.; Fang, M.; Lu, C.; Tian, P.; Mei, F. Molecular characteristics and prognostic factors of leptomeningeal metastasis in non-small cell lung cancer. Clin. Neurol. Neurosurg. 2023, 225, 107572. [Google Scholar] [CrossRef]
- Chen, M.; Li, H.; Xu, X.; Bao, X.; Xue, L.; Ai, X.; Xu, J.; Xu, M.; Shi, Y.; Zhen, T.; et al. Identification of RAC1 in promoting brain metastasis of lung adenocarcinoma using single-cell transcriptome sequencing. Cell Death Dis. 2023, 14, 330. [Google Scholar] [CrossRef]
- Munsterberg, J.; Loreth, D.; Brylka, L.; Werner, S.; Karbanova, J.; Gandrass, M.; Schneegans, S.; Besler, K.; Hamester, F.; Robador, J.R.; et al. ALCAM contributes to brain metastasis formation in non-small-cell lung cancer through interaction with the vascular endothelium. Neuro Oncol. 2020, 22, 955–966. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, Y.; Yi, G.; Yang, W.; Chen, K.; Tan, X.; Zhang, X.; Xu, Z.; Yang, Z.; Peng, Y. BDNF is a prognostic biomarker involved in immune infiltration of lung adenocarcinoma and is associated with brain metastasis. Immunology 2023, 168, 320–330. [Google Scholar] [CrossRef]
- Li, Z.; Liang, N.; Wang, N.; Jia, Y.; Tian, C. WDR5 is a prognostic biomarker of brain metastasis from non-small cell lung cancer. Front. Oncol. 2022, 12, 1023776. [Google Scholar] [CrossRef]
- Liang, N.; Sun, S.; Li, Z.; Wu, T.; Zhang, C.; Xin, T. CCKAR is a biomarker for prognosis and asynchronous brain metastasis of non-small cell lung cancer. Front. Oncol. 2022, 12, 1098728. [Google Scholar] [CrossRef]
- Zhou, S.; Xie, J.; Huang, Z.; Deng, L.; Wu, L.; Yu, J.; Meng, X. Anti-PD-(L)1 immunotherapy for brain metastases in non-small cell lung cancer: Mechanisms, advances, and challenges. Cancer Lett. 2021, 502, 166–179. [Google Scholar] [CrossRef]
- Reck, M.; Rodriguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csoszi, T.; Fulop, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef]
- Tonse, R.; Rubens, M.; Appel, H.; Tom, M.C.; Hall, M.D.; Odia, Y.; McDermott, M.W.; Ahluwalia, M.S.; Mehta, M.P.; Kotecha, R. Systematic review and meta-analysis of PD-L1 expression discordance between primary tumor and lung cancer brain metastasis. Neurooncol. Adv. 2021, 3, vdab166. [Google Scholar] [CrossRef]
- Takamori, S.; Toyokawa, G.; Okamoto, I.; Takada, K.; Kinoshita, F.; Kozuma, Y.; Matsubara, T.; Haratake, N.; Akamine, T.; Mukae, N.; et al. Clinical Significance of PD-L1 Expression in Brain Metastases from Non-small Cell Lung Cancer. Anticancer Res. 2018, 38, 553–557. [Google Scholar] [CrossRef]
- Zhou, D.; Gong, Z.; Wu, D.; Ma, C.; Hou, L.; Niu, X.; Xu, T. Harnessing immunotherapy for brain metastases: Insights into tumor-brain microenvironment interactions and emerging treatment modalities. J. Hematol. Oncol. 2023, 16, 121. [Google Scholar] [CrossRef]
- Wang, S.; Hu, C.; Xie, F.; Liu, Y. Use of Programmed Death Receptor-1 and/or Programmed Death Ligand 1 Inhibitors for the Treatment of Brain Metastasis of Lung Cancer. Onco Targets Ther. 2020, 13, 667–683. [Google Scholar] [CrossRef]
- Qian, H.; Zhang, Y.; Xu, J.; He, J.; Gao, W. Progress and application of circulating tumor cells in non-small cell lung cancer. Mol. Ther. Oncolytics 2021, 22, 72–84. [Google Scholar] [CrossRef]
- Lin, D.; Shen, L.; Luo, M.; Zhang, K.; Li, J.; Yang, Q.; Zhu, F.; Zhou, D.; Zheng, S.; Chen, Y.; et al. Circulating tumor cells: Biology and clinical significance. Signal Transduct. Target. Ther. 2021, 6, 404. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tang, C.; Liu, Y.; Sun, J.; Mu, X.; Zhang, L.; Dai, B.; Li, X.; Zhuo, H.; et al. Label-Free Isolation and mRNA Detection of Circulating Tumor Cells from Patients with Metastatic Lung Cancer for Disease Diagnosis and Monitoring Therapeutic Efficacy. Anal. Chem. 2015, 87, 11893–11900. [Google Scholar] [CrossRef]
- Hanssen, A.; Riebensahm, C.; Mohme, M.; Joosse, S.A.; Velthaus, J.L.; Berger, L.A.; Bernreuther, C.; Glatzel, M.; Loges, S.; Lamszus, K.; et al. Frequency of Circulating Tumor Cells (CTC) in Patients with Brain Metastases: Implications as a Risk Assessment Marker in Oligo-Metastatic Disease. Cancers 2018, 10, 527. [Google Scholar] [CrossRef]
- Scharpenseel, H.; Hanssen, A.; Loges, S.; Mohme, M.; Bernreuther, C.; Peine, S.; Lamszus, K.; Goy, Y.; Petersen, C.; Westphal, M.; et al. EGFR and HER3 expression in circulating tumor cells and tumor tissue from non-small cell lung cancer patients. Sci. Rep. 2019, 9, 7406. [Google Scholar] [CrossRef]
- Rehman, A.U.; Khan, P.; Maurya, S.K.; Siddiqui, J.A.; Santamaria-Barria, J.A.; Batra, S.K.; Nasser, M.W. Liquid biopsies to occult brain metastasis. Mol. Cancer 2022, 21, 113. [Google Scholar] [CrossRef]
- Li, M.; Hou, X.; Zheng, L.; Ma, Y.; Li, D.; Lv, Y.; Chen, J.; Zheng, W.; Shao, Y.; Mou, Y.; et al. Utilizing phenotypic characteristics of metastatic brain tumors to improve the probability of detecting circulating tumor DNA from cerebrospinal fluid in non-small-cell lung cancer patients: Development and validation of a prediction model in a prospective cohort study. ESMO Open 2022, 7, 100305. [Google Scholar] [CrossRef]
- Aldea, M.; Hendriks, L.; Mezquita, L.; Jovelet, C.; Planchard, D.; Auclin, E.; Remon, J.; Howarth, K.; Benitez, J.C.; Gazzah, A.; et al. Circulating Tumor DNA Analysis for Patients with Oncogene-Addicted NSCLC with Isolated Central Nervous System Progression. J. Thorac. Oncol. 2020, 15, 383–391. [Google Scholar] [CrossRef]
- Lubotzky, A.; Zemmour, H.; Neiman, D.; Gotkine, M.; Loyfer, N.; Piyanzin, S.; Ochana, B.L.; Lehmann-Werman, R.; Cohen, D.; Moss, J.; et al. Liquid biopsy reveals collateral tissue damage in cancer. JCI Insight 2022, 7, e153559. [Google Scholar] [CrossRef]
- Lam, V.K.; Zhang, J.; Wu, C.C.; Tran, H.T.; Li, L.; Diao, L.; Wang, J.; Rinsurongkawong, W.; Raymond, V.M.; Lanman, R.B.; et al. Genotype-Specific Differences in Circulating Tumor DNA Levels in Advanced NSCLC. J. Thorac. Oncol. 2021, 16, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Ye, X.; Hu, Z.; Peng, K.; Song, Y.; Yin, X.; Zhu, G.; Ji, Q.; Peng, Y. EGFR mutation status and its impact on survival of Chinese non-small cell lung cancer patients with brain metastases. Tumour Biol. 2014, 35, 2437–2444. [Google Scholar] [CrossRef] [PubMed]
- Reichert, Z.R.; Morgan, T.M.; Li, G.; Castellanos, E.; Snow, T.; Dall’Olio, F.G.; Madison, R.W.; Fine, A.D.; Oxnard, G.R.; Graf, R.P.; et al. Prognostic value of plasma circulating tumor DNA fraction across four common cancer types: A real-world outcomes study. Ann. Oncol. 2023, 34, 111–120. [Google Scholar] [CrossRef]
- Li, M.; Chen, J.; Zhang, B.; Yu, J.; Wang, N.; Li, D.; Shao, Y.; Zhu, D.; Liang, C.; Ma, Y.; et al. Dynamic monitoring of cerebrospinal fluid circulating tumor DNA to identify unique genetic profiles of brain metastatic tumors and better predict intracranial tumor responses in non-small cell lung cancer patients with brain metastases: A prospective cohort study (GASTO 1028). BMC Med. 2022, 20, 398. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Huang, T.; Wang, Y.; Song, M.M.; Song, T.; Long, G.; Zhang, X.; Li, X.; Zhang, L. Cerebrospinal fluid circulating tumor DNA depicts profiling of brain metastasis in NSCLC. Mol. Oncol. 2023, 17, 810–824. [Google Scholar] [CrossRef]
- Tsakonas, G.; Tadigotla, V.; Chakrabortty, S.K.; Stragliotto, G.; Chan, D.; Lewensohn, R.; Yu, W.; Skog, J.K.; Hydbring, P.; Ekman, S. Cerebrospinal fluid as a liquid biopsy for molecular characterization of brain metastasis in patients with non-small cell lung cancer. Lung Cancer 2023, 182, 107292. [Google Scholar] [CrossRef]
- De Mattos-Arruda, L.; Mayor, R.; Ng, C.K.Y.; Weigelt, B.; Martinez-Ricarte, F.; Torrejon, D.; Oliveira, M.; Arias, A.; Raventos, C.; Tang, J.; et al. Cerebrospinal fluid-derived circulating tumour DNA better represents the genomic alterations of brain tumours than plasma. Nat. Commun. 2015, 6, 8839. [Google Scholar] [CrossRef]
- Aljohani, H.M.; Aittaleb, M.; Furgason, J.M.; Amaya, P.; Deeb, A.; Chalmers, J.J.; Bahassi, E.M. Genetic mutations associated with lung cancer metastasis to the brain. Mutagenesis 2018, 33, 137–145. [Google Scholar] [CrossRef]
- Ma, C.; Yang, X.; Xing, W.; Yu, H.; Si, T.; Guo, Z. Detection of circulating tumor DNA from non-small cell lung cancer brain metastasis in cerebrospinal fluid samples. Thorac. Cancer 2020, 11, 588–593. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Xu, X.; Qian, Z.; Zhang, C.; Niu, Y.; Wang, Z.; Sun, J.; Zhang, X.; Yu, Y. The biological functions and clinical applications of exosomes in lung cancer. Cell Mol. Life Sci. 2019, 76, 4613–4633. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Cheng, Z.; Qin, W.; Jiang, L. Exosomes as a liquid biopsy for lung cancer. Lung Cancer 2018, 116, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.D.; Castanho, M.; Neves, V. Exosomes and Brain Metastases: A Review on Their Role and Potential Applications. Int. J. Mol. Sci. 2021, 22, 10899. [Google Scholar] [CrossRef]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, Y.; Liu, R.; Xie, Y.; Jin, Y.; Wang, M.; Yu, Z.; Wang, W.; Luo, X. Non-small cell lung cancer-derived exosomes promote proliferation, phagocytosis, and secretion of microglia via exosomal microRNA in the metastatic microenvironment. Transl. Oncol. 2023, 27, 101594. [Google Scholar] [CrossRef]
- Li, S.; Qu, Y.; Liu, L.; Zhang, X.; He, Y.; Wang, C.; Guo, Y.; Yuan, L.; Ma, Z.; Bai, H.; et al. Comparative proteomic profiling of plasma exosomes in lung cancer cases of liver and brain metastasis. Cell Biosci. 2023, 13, 180. [Google Scholar] [CrossRef]
- Rodrigues, G.; Hoshino, A.; Kenific, C.M.; Matei, I.R.; Steiner, L.; Freitas, D.; Kim, H.S.; Oxley, P.R.; Scandariato, I.; Casanova-Salas, I.; et al. Tumour exosomal CEMIP protein promotes cancer cell colonization in brain metastasis. Nat. Cell Biol. 2019, 21, 1403–1412. [Google Scholar] [CrossRef]
- Chen, G.Y.; Cheng, J.C.; Chen, Y.F.; Yang, J.C.; Hsu, F.M. Circulating Exosomal Integrin beta3 Is Associated with Intracranial Failure and Survival in Lung Cancer Patients Receiving Cranial Irradiation for Brain Metastases: A Prospective Observational Study. Cancers 2021, 13, 380. [Google Scholar] [CrossRef]
- Xiao, W.; Ma, W.; Wei, S.; Li, Q.; Liu, R.; Carney, R.P.; Yang, K.; Lee, J.; Nyugen, A.; Yoneda, K.Y.; et al. High-affinity peptide ligand LXY30 for targeting alpha3beta1 integrin in non-small cell lung cancer. J. Hematol. Oncol. 2019, 12, 56. [Google Scholar] [CrossRef]
- Chen, L.T.; Xu, S.D.; Xu, H.; Zhang, J.F.; Ning, J.F.; Wang, S.F. MicroRNA-378 is associated with non-small cell lung cancer brain metastasis by promoting cell migration, invasion and tumor angiogenesis. Med. Oncol. 2012, 29, 1673–1680. [Google Scholar] [CrossRef]
- Zhao, C.; Xu, Y.; Zhang, Y.; Tan, W.; Xue, J.; Yang, Z.; Zhang, Y.; Lu, Y.; Hu, X. Downregulation of miR-145 contributes to lung adenocarcinoma cell growth to form brain metastases. Oncol. Rep. 2013, 30, 2027–2034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, S.; Yao, J.; Lowery, F.J.; Zhang, Q.; Huang, W.C.; Li, P.; Li, M.; Wang, X.; Zhang, C.; et al. Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth. Nature 2015, 527, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Deng, S.; Li, L.; Liu, T.; Zhang, T.; Li, J.; Yu, Y.; Xu, Y. TGF-beta1-mediated exosomal lnc-MMP2-2 increases blood-brain barrier permeability via the miRNA-1207-5p/EPB41L5 axis to promote non-small cell lung cancer brain metastasis. Cell Death Dis. 2021, 12, 721. [Google Scholar] [CrossRef]
- Im, J.H.; Kim, T.H.; Lee, K.Y.; Gwak, H.S.; Lin, W.; Park, J.B.; Kim, J.H.; Yoo, B.C.; Park, S.M.; Kwon, J.W.; et al. Exploratory Profiling of Extracellular MicroRNAs in Cerebrospinal Fluid Comparing Leptomeningeal Metastasis with Other Central Nervous System Tumor Statuses. J. Clin. Med. 2021, 10, 4860. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, R.; Cai, Q.; Gao, X.; Tong, F.; Dong, J.; Hu, Y.; Wu, G.; Dong, X. MicroRNA-330-3p promotes brain metastasis and epithelial-mesenchymal transition via GRIA3 in non-small cell lung cancer. Aging 2019, 11, 6734–6761. [Google Scholar] [CrossRef]
- Tsakonas, G.; Koulouris, A.; Kazmierczak, D.; Botling, J.; Ortiz-Villalon, C.; Nord, H.; Lindskog, M.; Sandelin, M.; Micke, P.; Hydbring, P.; et al. Matched Analyses of Brain Metastases versus Primary Non-Small Cell Lung Cancer Reveal a Unique microRNA Signature. Int. J. Mol. Sci. 2022, 24, 193. [Google Scholar] [CrossRef]
- Chen, L.J.; Li, X.Y.; Zhao, Y.Q.; Liu, W.J.; Wu, H.J.; Liu, J.; Mu, X.Q.; Wu, H.B. Down-regulated microRNA-375 expression as a predictive biomarker in non-small cell lung cancer brain metastasis and its prognostic significance. Pathol. Res. Pract. 2017, 213, 882–888. [Google Scholar] [CrossRef]
- Li, C.; Zheng, H.; Xiong, J.; Huang, Y.; Li, H.; Jin, H.; Ai, S.; Wang, Y.; Su, T.; Sun, G.; et al. miR-596-3p suppresses brain metastasis of non-small cell lung cancer by modulating YAP1 and IL-8. Cell Death Dis. 2022, 13, 699. [Google Scholar] [CrossRef]
- Nowakowski, A.; Lahijanian, Z.; Panet-Raymond, V.; Siegel, P.M.; Petrecca, K.; Maleki, F.; Dankner, M. Radiomics as an emerging tool in the management of brain metastases. Neurooncol. Adv. 2022, 4, vdac141. [Google Scholar] [CrossRef]
- Chu, X.; Gong, J.; Yang, X.; Ni, J.; Gu, Y.; Zhu, Z. A “Seed-and-Soil” Radiomics Model Predicts Brain Metastasis Development in Lung Cancer: Implications for Risk-Stratified Prophylactic Cranial Irradiation. Cancers 2023, 15, 307. [Google Scholar] [CrossRef]
- Kniep, H.C.; Madesta, F.; Schneider, T.; Hanning, U.; Schonfeld, M.H.; Schon, G.; Fiehler, J.; Gauer, T.; Werner, R.; Gellissen, S. Radiomics of Brain MRI: Utility in Prediction of Metastatic Tumor Type. Radiology 2019, 290, 479–487. [Google Scholar] [CrossRef]
- Zhou, Y.; Ma, X.L.; Zhang, T.; Wang, J.; Zhang, T.; Tian, R. Use of radiomics based on (18)F-FDG PET/CT and machine learning methods to aid clinical decision-making in the classification of solitary pulmonary lesions: An innovative approach. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 2904–2913. [Google Scholar] [CrossRef]
- Mendoza, D.P.; Piotrowska, Z.; Lennerz, J.K.; Digumarthy, S.R. Role of imaging biomarkers in mutation-driven non-small cell lung cancer. World J. Clin. Oncol. 2020, 11, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, Y.Z.; Li, W.H.; Han, Y.; Li, Q.; Ye, Z. Pretreatment Thoracic CT Radiomic Features to Predict Brain Metastases in Patients with ALK-Rearranged Non-Small Cell Lung Cancer. Front. Genet. 2022, 13, 772090. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Kwon, H.; Yang, J.J.; Park, M.; Cha, Y.J.; Suh, S.H.; Lee, J.M. Contrast-enhanced T1-weighted image radiomics of brain metastases may predict EGFR mutation status in primary lung cancer. Sci. Rep. 2020, 10, 8905. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Zeng, J.; Chen, S.; Lyu, Y.; Toomey, K.A.; Phan, C.T.; Yoneda, K.Y.; Li, T. Small molecule tyrosine kinase inhibitors modulated blood immune cell counts in patients with oncogene-driven NSCLC. Biomark. Res. 2021, 9, 69. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Park, S.; Byun, H.K.; Lee, C.G.; Cho, J.; Hong, M.H.; Kim, H.R.; Cho, B.C.; Kim, S.; Park, J.; et al. Impact of Treatment-Related Lymphopenia on Immunotherapy for Advanced Non-Small Cell Lung Cancer. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Wu, J.; Liu, Y.; Zhou, J.; Wang, W.; Wang, X.; Guo, J.; Wang, Q.; Zhang, X.; Li, D.; et al. Relationship Between Neutrophil-To-Lymphocyte Ratio and Brain Metastasis in Non-Small Cell Lung Cancer Patients. Cancer Control 2022, 29, 10732748221076805. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, L.; Wang, Z.; Xu, Q.; Huang, H.; Wang, H.; Li, X.; Yu, M.; Chen, J.; Lin, F.; et al. Prognostic value of the modified systemic inflammation score in non-small-cell lung cancer with brain metastasis. Cancer Cell Int. 2022, 22, 320. [Google Scholar] [CrossRef]
- Li, W.; Qu, Y.; Wen, F.; Yu, R.; He, X.; Jia, H.; Liu, H.; Yu, H. Prognostic nutritional index and systemic immune-inflammation index are prognostic biomarkers for non-small-cell lung cancer brain metastases. Biomark. Med. 2021, 15, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Wei, S.; Long, S.; Tian, E.C.; McLaughlin, B.; Jaimes, M.; Montoya, D.J.; Viswanath, V.R.; Chien, J.; Zhang, Q.; et al. Dynamic evaluation of blood immune cells predictive of response to immune checkpoint inhibitors in NSCLC by multicolor spectrum flow cytometry. Front. Immunol. 2023, 14, 1206631. [Google Scholar] [CrossRef] [PubMed]
- Woldmar, N.; Schwendenwein, A.; Kuras, M.; Szeitz, B.; Boettiger, K.; Tisza, A.; Laszlo, V.; Reiniger, L.; Bago, A.G.; Szallasi, Z.; et al. Proteomic analysis of brain metastatic lung adenocarcinoma reveals intertumoral heterogeneity and specific alterations associated with the timing of brain metastases. ESMO Open 2023, 8, 100741. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, O.; Saavedra-Perez, D.; Kuri, R.; Aviles-Salas, A.; Martinez, L.; Mendoza-Posada, D.; Castillo, P.; Astorga, A.; Guzman, E.; De la Garza, J. Brain metastasis development and poor survival associated with carcinoembryonic antigen (CEA) level in advanced non-small cell lung cancer: A prospective analysis. BMC Cancer 2009, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Liu, W.; Xu, M.; Qin, H.; Liu, C.; Zhang, R.; Zhou, S.; Li, E.; Liu, Z.; Wang, Q. Cathepsin F and Fibulin-1 as novel diagnostic biomarkers for brain metastasis of non-small cell lung cancer. Br. J. Cancer 2022, 126, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, X.; Wu, H.; Jia, Y.; Liu, J.; Mu, X.; Wu, H.; Zhao, Y. Over-expression of S100B protein as a serum marker of brain metastasis in non-small cell lung cancer and its prognostic value. Pathol. Res. Pract. 2019, 215, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wen, M.; Liu, Y.; Wang, Y.; Jing, P.; Gu, Z.; Jiang, T.; Wang, W. The human microbiome: A promising target for lung cancer treatment. Front. Immunol. 2023, 14, 1091165. [Google Scholar] [CrossRef]
- Lu, H.; Gao, N.L.; Tong, F.; Wang, J.; Li, H.; Zhang, R.; Ma, H.; Yang, N.; Zhang, Y.; Wang, Y.; et al. Alterations of the Human Lung and Gut Microbiomes in Non-Small Cell Lung Carcinomas and Distant Metastasis. Microbiol. Spectr. 2021, 9, e0080221. [Google Scholar] [CrossRef]
- Chow, S.C.; Gowing, S.D.; Cools-Lartigue, J.J.; Chen, C.B.; Berube, J.; Yoon, H.W.; Chan, C.H.; Rousseau, M.C.; Bourdeau, F.; Giannias, B.; et al. Gram negative bacteria increase non-small cell lung cancer metastasis via Toll-like receptor 4 activation and mitogen-activated protein kinase phosphorylation. Int. J. Cancer 2015, 136, 1341–1350. [Google Scholar] [CrossRef]
- Huang, D.; Su, X.; Yuan, M.; Zhang, S.; He, J.; Deng, Q.; Qiu, W.; Dong, H.; Cai, S. The characterization of lung microbiome in lung cancer patients with different clinicopathology. Am. J. Cancer Res. 2019, 9, 2047–2063. [Google Scholar]
- Wu, S.Y.; Watabe, K. The roles of microglia/macrophages in tumor progression of brain cancer and metastatic disease. Front. Biosci. (Landmark. Ed.) 2017, 22, 1805–1829. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, L.; Bin, Y.; Zeng, H.; Wang, J.; Zhang, R.; Tong, F.; Yang, N.; Dong, X. Microbiota-modulated spermine promotes brain metastasis in non-small cell lung cancer by regulating microglia M2 polarization via the STAT3 pathway. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Ernani, V.; Stinchcombe, T.E. Management of Brain Metastases in Non-Small-Cell Lung Cancer. J. Oncol. Pract. 2019, 15, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Brozos-Vazquez, E.M.; Rodriguez-Lopez, C.; Cortegoso-Mosquera, A.; Lopez-Landrove, S.; Muinelo-Romay, L.; Garcia-Gonzalez, J.; Lopez-Lopez, R.; Leon-Mateos, L. Immunotherapy in patients with brain metastasis: Advances and challenges for the treatment and the application of circulating biomarkers. Front. Immunol. 2023, 14, 1221113. [Google Scholar] [CrossRef]
- Wei, Y.; Xu, Y.; Wang, M. Immune checkpoint inhibitors for the treatment of non-small cell lung cancer brain metastases. Chin. Med. J. (Engl.) 2023, 136, 1523–1531. [Google Scholar] [CrossRef]
- Hou, X.; Zhou, C.; Wu, G.; Lin, W.; Xie, Z.; Zhang, H.; Yi, J.; Peng, Z.; Yin, L.; Ma, C.; et al. Efficacy, Safety, and Health-Related Quality of Life with Camrelizumab Plus Pemetrexed and Carboplatin as First-Line Treatment for Advanced Nonsquamous NSCLC with Brain Metastases (CAP-BRAIN): A Multicenter, Open-Label, Single-Arm, Phase 2 Study. J. Thorac. Oncol. 2023, 18, 769–779. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Wakuda, K.; Fukuda, M.; Kenmotsu, H.; Mukae, H.; Ito, K.; Chibana, K.; Inoue, K.; Miura, S.; Tanaka, K.; et al. A Phase II Study of Osimertinib for Radiotherapy-Naive Central Nervous System Metastasis From NSCLC: Results for the T790M Cohort of the OCEAN Study (LOGIK1603/WJOG9116L). J. Thorac. Oncol. 2021, 16, 2121–2132. [Google Scholar] [CrossRef]
- Zhou, R.; Zheng, S.; Huo, L.; Luo, Q.; Wang, D.; Guo, J.; Liu, B.; Zhang, J.; Wang, B.; Qiu, B.; et al. Effect and safety of combining bevacizumab and fractionated stereotactic radiotherapy for brain metastases in patients with non-small cell lung cancer: A phase II trial. J. Clin. Oncol. 2023, 41, e21132. [Google Scholar] [CrossRef]
- Hong, M.H.; Choi, Y.J.; Ahn, H.K.; Keam, B.; Kim, Y.J.; Kim, J.W.; Kim, H.R.; Kang, J.-H. Phase II trial of lazertinib in patients with epidermal growth factor receptor (EGFR) mutation-positive (M+), metastatic non-small cell lung cancer (NSCLC) with asymptomatic or mild symptomatic brain metastases after failure of EGFR tyrosine kinase inhibitor (TKI) (KCSG LU20-15). J. Clin. Oncol. 2023, 41, 9054. [Google Scholar] [CrossRef]
- Nadal, E.; Rodriguez-Abreu, D.; Massuti, B.; Juan-Vidal, O.; Huidobro Vence, G.; Lopez, R.; de Castro Carpeño, J.; Estival, A.; Campelo, R.G.; Sullivan, I.; et al. Updated analysis from the ATEZO-BRAIN trial: Atezolizumab plus carboplatin and pemetrexed in patients with advanced nonsquamous non–small cell lung cancer with untreated brain metastases. J. Clin. Oncol. 2022, 40, 9010. [Google Scholar] [CrossRef]
- Zhang, L.; You, Y.; Liu, X.; Liu, F.; Nie, K.; Ji, Y. Osimertinib combined with bevacizumab as the first-line treatment in non-small cell lung cancer patients with brain metastasis harboring epidermal growth factor receptor mutations. Thorac. Cancer 2023, 14, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yu, Q.; Yang, N.; Liu, L.; Yu, Y.; Wang, Q.; Zhao, Y.; Yang, Y.; Jiang, W.; Tan, L.; et al. Tislelizumab plus chemotherapy in patients with advanced brain metastases of non-squamous non-small cell lung cancer: A multicenter, prospective, open-label phase 2 study. J. Clin. Oncol. 2023, 41, 9080. [Google Scholar] [CrossRef]
- Wu, Y.-L.; Zhou, Q.; Wang, J.; Yu, Y.; Xing, L.; Wang, Y.; Cheng, Y.; Pan, Y.; Fan, Y.; Shi, J.; et al. Randomized phase 3 study of first-line AZD3759 (zorifertinib) versus gefitinib or erlotinib in EGFR-mutant (EGFRm+) non–small-cell lung cancer (NSCLC) with central nervous system (CNS) metastasis. J. Clin. Oncol. 2023, 41, 9001. [Google Scholar] [CrossRef]
- Paik, P.K.; Garassino, M.C.; Le, X.; Thomas, M.; Sakai, H.; Veillon, R.; Smit, E.F.; Mazieres, J.; Cortot, A.B.; Raskin, J.; et al. Long-term outcomes of tepotinib in patients with MET exon 14 skipping NSCLC from the VISION study. J. Clin. Oncol. 2023, 41, 9060. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Liu, Z.; Wang, L.; Yao, Y.; Liu, Y.; Hao, X.Z.; Wang, J.; Xing, P.; Li, J. Efficacy of dacomitinib in patients with EGFR-mutated NSCLC and brain metastases. Thorac. Cancer 2021, 12, 3407–3415. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Park, S.; Lee, S.H.; Ahn, J.S.; Ahn, M.J.; Sun, J.M. Dacomitinib in EGFR-mutant non-small-cell lung cancer with brain metastasis: A single-arm, phase II study. ESMO Open 2023, 8, 102068. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.C.; Han, J.Y.; Kim, S.W.; Lee, K.H.; Cho, E.K.; Lee, Y.G.; Kim, D.W.; Kim, J.H.; Lee, G.W.; Lee, J.S.; et al. A Phase 1/2 Study of Lazertinib 240 mg in Patients with Advanced EGFR T790M-Positive NSCLC After Previous EGFR Tyrosine Kinase Inhibitors. J. Thorac. Oncol. 2022, 17, 558–567. [Google Scholar] [CrossRef]
- Cho, B.C.; Ahn, M.-J.; Kang, J.H.; Soo, R.A.; Reungwetwattana, T.; Yang, J.C.-H.; Cicin, I.; Kim, D.-W.; Wu, Y.-L.; Lu, S.; et al. Lazertinib Versus Gefitinib as First-Line Treatment in Patients with EGFR-Mutated Advanced Non–Small-Cell Lung Cancer: Results From LASER301. J. Clin. Oncol. 2023, 41, 4208–4217. [Google Scholar] [CrossRef]
- Crino, L.; Bronte, G.; Bidoli, P.; Cravero, P.; Minenza, E.; Cortesi, E.; Garassino, M.C.; Proto, C.; Cappuzzo, F.; Grossi, F.; et al. Nivolumab and brain metastases in patients with advanced non-squamous non-small cell lung cancer. Lung Cancer 2019, 129, 35–40. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Clinical Trials Identifier | Drug Class | Drug Name | Phase of Clinical Trial | Number of Patients | Biomarker | Disease | Median OS | PFS | HR | ORR/DCR | Ref |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NCT04211090 | PD-1/L1 inhibitor | Camrelizumab, Pemetrexed, Carboplatin | 2 | 45 | Driven Gene-negative | NSCLC Brain Metastases | 21 months | 7.6 months | NA | 46.7%/34% | [156] |
| NCT03769103 | TKI | Osimertinib and Stereotactic Radiosurgery (SRS) | 2 | 66 | EGFR Mutation | EGFR-Mutated NSCLC With Brain Metastases | 25.2 | 7.1 months | NA | 40.5%/NA | [157] |
| NCT04345146 | Antiangiogenic agents | Fractionated stereotactic radiation therapy (FSRT) combined with Bevacizumab | 2 | 108 | None | Non-Squamous Non-Small-Cell Lung Cancer with Brain Metastases | NA | 15 months | NA | NA/NA | [158] |
| NCT05326425 | TKI | Lazertinib (YH25448) | 2 | 40 | EGFR Mutation | Asymptomatic or Mild Symptomatic Brain Metastases NSCLC | Not reached | Not reached | NA | 57.9%/97.4% | [159] |
| NCT03526900 | PD-1/L1 inhibitor | Atezolizumab in Combination with Carboplatin Plus Pemetrexed | 2 | 40 | EGFR mutation and ALK fusion negative | NSCLC with Asymptomatic Brain Metastasis | 13.6 | 8.9 months | 0.99; 95% CI 0.35 to 2.12 | NA/NA | [160] |
| NCT05104281 | Antiangiogenic agents | Osimertinib oral and bevazizumab intravenously | 3 | 52 | EGFR Mutation | NSCLC Brain Metastases | Not reached | 22 months | NA | 82.7%/96.2% | [161] |
| NCT04507217 | PD-1/L1 inhibitor | Tislelizumab, Carboplatin, Pemetrexed | 2 | 36 | None | Non-Squamous Non-Small-Cell Lung Cancer Brain Metastatic | Not reached | Not reached | NA | 56.7%/96.7% | [162] |
| NCT03653546 | TKI | AZD3759, Erlotinib, Gefitinib | 2, 3 | 439 | EGFR Mutation | NSCLC Brain Metastases | NA | 9.6 months | NA | 68.60% | [163] |
| NCT02864992 | MET tyrosine kinase inhibitor | Tepotinib | 2 | 313 | MET exon 14 skipping mutation | Patients with advanced or metastatic NSCLC with a confirmed MET exon 14 skipping mutation | 19.6 months | 11.2 months | NA | 51.4%/NA | [164] |
| NCT04768491 | TKI | Dacomitinib | Ambispective cohort study in real world | 32 | EGFR Mutation | Advanced NSCLC with EGFR Mutations and Brain Metastasis | NA | Not reached | NA | 87.5%/100% | [165] |
| NCT04339829 | TKI | Dacomitinib | 2 | 30 | EGFR Mutation | EGFR Mutation-Positive NSCLC | NA | 17.5 months | NA | 96.7% | [166] |
| NCT03046992 | TKI | Lazertinib | Phase 1/2 | 78 | EGFR Mutation | EGFR Mutation-Positive NSCLC | Not reached | 11.1 months | NA | 55.30%/NA | [167] |
| NCT04248829 | TKI | Lazertinib | 3 | 393 | EGFR Mutation | EGFR Mutation-Positive NSCLC | Not reached | 20.6 months | 0.45; 95% CI, 0.34 to 0.58 | 76%/NA | [168] |
| CA209966 | PD-1/L1 inhibitor | Nivolumab | NA | 409 | None | Advanced Non-Squamous Non-Small-Cell Lung Cancer | 8.6 months | 3 months | NA | 17%/39% | [169] |
| Clinical Trials Identifier | Status | Phase of Clinical Trial | Estimated Enrollment | First Posted | Estimated Completion Date | Intervention/Treatment | Condition or Disease | Biomarker |
|---|---|---|---|---|---|---|---|---|
| NCT04889066 | Not yet recruiting | 2 | 46 | 2021.5 | 2025.1 | Durvalumab and radiotherapy | NSCLC Brain Metastases | PD-L1 Expression ≥ 1% |
| NCT04978753 | Recruiting | 2 | 54 | 2021.7 | 2023.12 | Anlotinib | NSCLC Brain Metastases | EGFR Mutation |
| NCT05812534 | Not yet recruiting | 2 | 36 | 2023.4 | 2025.6 | Cadonilimab combined with bevacizumab and chemotherapy | NSCLC Brain Metastases | EGFR and ALK negative |
| NCT05012254 | Recruiting | 2 | 71 | 2021.8 | 2026.12 | Platinum-based chemotherapy plus nivolumab and ipilimumab | NSCLC Brain Metastases | N/A |
| NCT05746481 | Not yet recruiting | 2 | 35 | 2023.2 | 2026.12 | Tiragolumab combined with carboplatin, pemetrexed, and atezolizumab | Non-Squamous Non-Small-Cell Lung Cancer (NSCLC) and Untreated Brain Metastases | N/A |
| NCT04824079 | Recruiting | 2 | 30 | 2022.4 | 2024.7 | Keynatinib | EGFR-Mutated NSCLC With Brain Metastases | EGFR Mutation |
| NCT05800223 | Not yet recruiting | 3 | 300 | 2023.4 | 2028.12 | Armatinib combined with stereotactic body radiotherapy (SRT) | EGFR-Mutated NSCLC With Brain Metastases | EGFR Mutation |
| NCT04967417 | Recruiting | 2 | 50 | 2021.7 | 2026.2 | Pemetrexed, carboplatin, pembrolizumab | NSCLC Patients with Asymptomatic Brain Metastases | N/A |
| NCT05840770 | Not yet recruiting | 2 | 34 | 2023.5 | 2029.1 | Cemiplimab | NSCLC Brain Metastases | PD-L1 ≥ 50% |
| NCT03497767 | Recruiting | 2 | 80 | 2018.4 | 2024.3 | Osimertinib and stereotactic radiosurgery (SRS) | NSCLC Brain Metastases | EGFR Mutation |
| NCT04905550 | Recruiting | 2 | 50 | 2021.5 | 2024.8 | Craniocerebral radiotherapy combined with Almonertinib | NSCLC Brain Metastases | EGFR Mutation |
| NCT05522660 | Recruiting | 3 | 190 | 2022.8 | 2026.12 | Immune checkpoint inhibitor-combined stereotactic radiosurgery | NSCLC Brain Metastases | N/A |
| NCT05477615 | Not yet recruiting | 2 | 28 | 2022.7 | 2025.6 | Lazertinib, pemetrexed, carboplatin | NSCLC Brain Metastases | EGFR Mutation |
| NCT05567055 | Not yet recruiting | 2 | 35 | 2022.10 | 2028.5 | Capmatinib | NSCLC with Asymptomatic Brain Metastases | MET amplification or METΔex14 detected on cfDNA |
| NCT04768075 | Not yet recruiting | 3 | 200 | 2021.2 | 2024.4 | Camrelizumab combined with SRT/WBRT and chemotherapy | NSCLC Brain Metastases | Driven Gene-negative |
| NCT05236946 | Recruiting | 3 | 190 | 2022.2 | 2026.7 | TKI combined with SRS/WBRT | NSCLC with Asymptomatic Brain Metastases | EGFR/ALK Mutation |
| NCT05807893 | Not yet recruiting | 2, 3 | 30 | 2023.4 | 2025.3 | Serplulimab combined with bevacizumab and first-line chemotherapy | NSCLC Brain Metastases | Negative Driver Gene |
| NCT04233021 | Active, not recruiting | 2 | 69 | 2020.1 | 2024.2 | Osimertinib | NSCLC Brain Metastases | EGFR Activating Mutation |
| NCT05180422 | Recruiting | 1, 2 | 43 | 2022.1 | 2028.6 | AMG 510 plus MVASI | NSCLC Brain Metastases | KRAS p.G12C Mutant |
| NCT04964960 | Recruiting | 2 | 45 | 2021.7 | 2033.5 | Pembrolizumab and Chemotherapy | NSCLC with Asymptomatic Brain Metastatic | Driven Gene-negative |
| NCT04291092 | Recruiting | 2 | 63 | 2020.3 | 2023.6 | Camrelizumab combined with chemotherapy and local treatment of brain metastases (WBRT, r-knife, SRS, etc.) | NSCLC Brain Metastases | Driven Gene-negative |
| NCT05146219 | Recruiting | 2 | 60 | 2021.12 | 2024.12 | TY-9591 tablets | NSCLC Brain Metastases | EGFR Activating Mutation |
| NCT05317858 | Recruiting | 3 | 20 | 2022.8 | 2024.12 | Pembrolizumab | NSCLC Brain Metastases | Driven Gene-negative |
| NCT04829019 | Recruiting | 2 | 88 | 2021.4 | 2025.10 | Osimertinib and whole-brain irradiation | NSCLC Brain Metastases | EGFR Mutation |
| NCT04789668 | Active, not recruiting | 1, 2 | 10 | 2021.3 | 2025.1 | Bintrafusp alfa and pimasertib | NSCLC Brain Metastases | N/A |
| NCT06020066 | Recruiting | 2 | 202 | 2023.8 | 2029.8 | EGFR-TK inhibitor or SRS | NSCLC Brain Metastases | EGFR L858R, EGFR exon 19 deletion |
| NCT05999357 | Not yet recruiting | 2 | 42 | 2023.8 | 2028.8 | JDQ443 | NSCLC Brain Metastases | KRAS G12C |
| NCT05865990 | Not yet recruiting | 2 | 60 | 2023.5 | 2026.10 | Patritumab deruxtecan (HER3-DXd), | NSCLC Brain Metastases | HER2-Positive |
| NCT05948813 | Recruiting | 2 | 420 | 2023.7 | 2027.12 | TY-9591 | EGFR-Mutated Non-Small-Cell Lung Cancer with Brain Metastases | EGFR-Mutated |
| NCT06047379 | Not yet recruiting | 1, 2 | 134 | 2023.9 | 2026.8 | NEO212 pembrolizumab or nivolumab | NSCLC expressing PD-L1, with no EGFR or ALK genomic tumor aberrations | PD-L1, no EGFR or ALK genomic tumor aberrations |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, J.; Yu, J.-Z.; Liu, C.; Ould Ismail, A.A.O.; Ma, W. Exploring the Molecular Tumor Microenvironment and Translational Biomarkers in Brain Metastases of Non-Small-Cell Lung Cancer. Int. J. Mol. Sci. 2024, 25, 2044. https://doi.org/10.3390/ijms25042044
Wen J, Yu J-Z, Liu C, Ould Ismail AAO, Ma W. Exploring the Molecular Tumor Microenvironment and Translational Biomarkers in Brain Metastases of Non-Small-Cell Lung Cancer. International Journal of Molecular Sciences. 2024; 25(4):2044. https://doi.org/10.3390/ijms25042044
Chicago/Turabian StyleWen, Jiexi, Jie-Zeng Yu, Catherine Liu, A. Aziz O. Ould Ismail, and Weijie Ma. 2024. "Exploring the Molecular Tumor Microenvironment and Translational Biomarkers in Brain Metastases of Non-Small-Cell Lung Cancer" International Journal of Molecular Sciences 25, no. 4: 2044. https://doi.org/10.3390/ijms25042044
APA StyleWen, J., Yu, J.-Z., Liu, C., Ould Ismail, A. A. O., & Ma, W. (2024). Exploring the Molecular Tumor Microenvironment and Translational Biomarkers in Brain Metastases of Non-Small-Cell Lung Cancer. International Journal of Molecular Sciences, 25(4), 2044. https://doi.org/10.3390/ijms25042044