
Mechanisms and Physiological Roles of Polymorphisms in Gestational Diabetes Mellitus

Abstract
1. Introduction
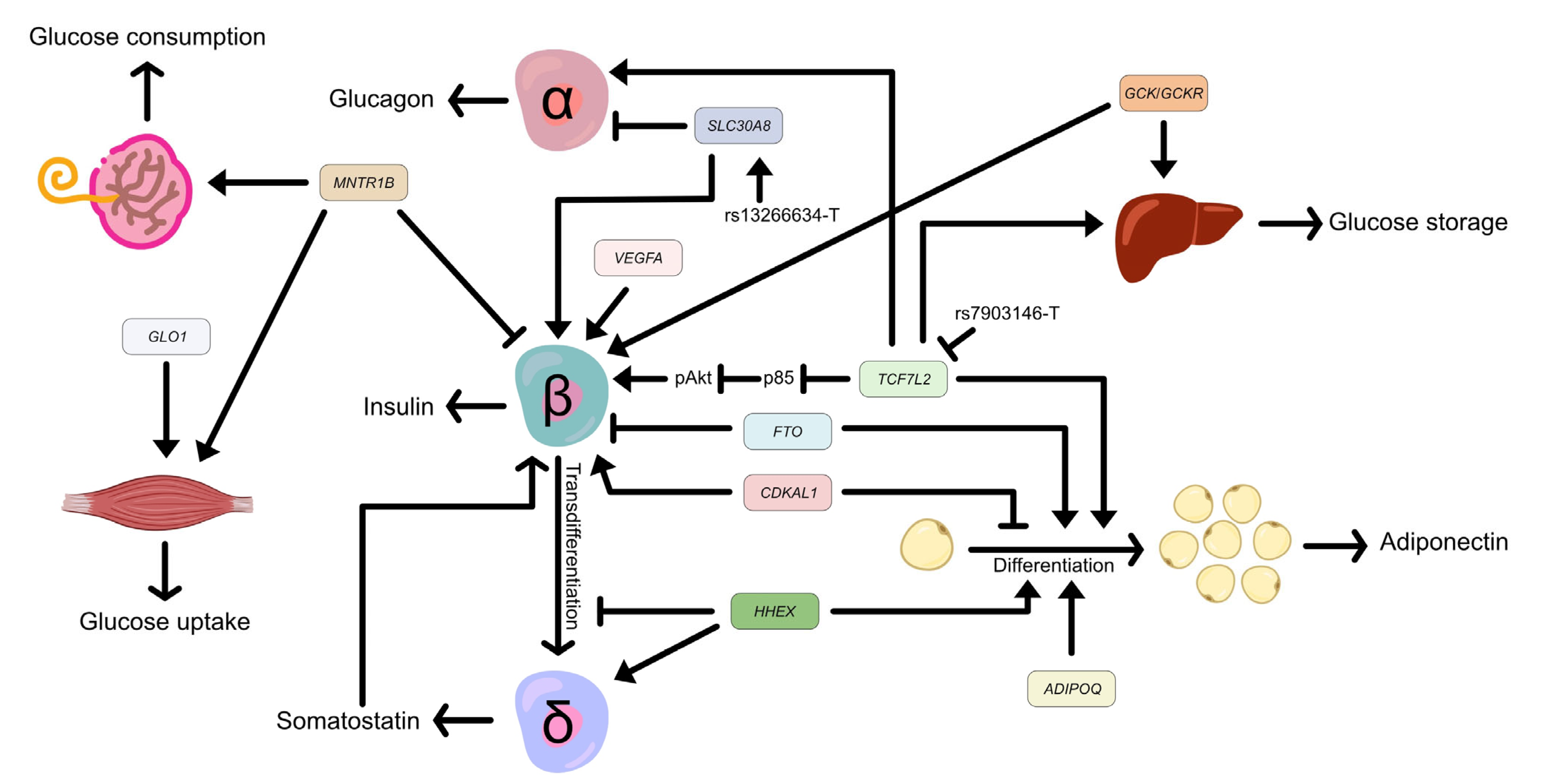
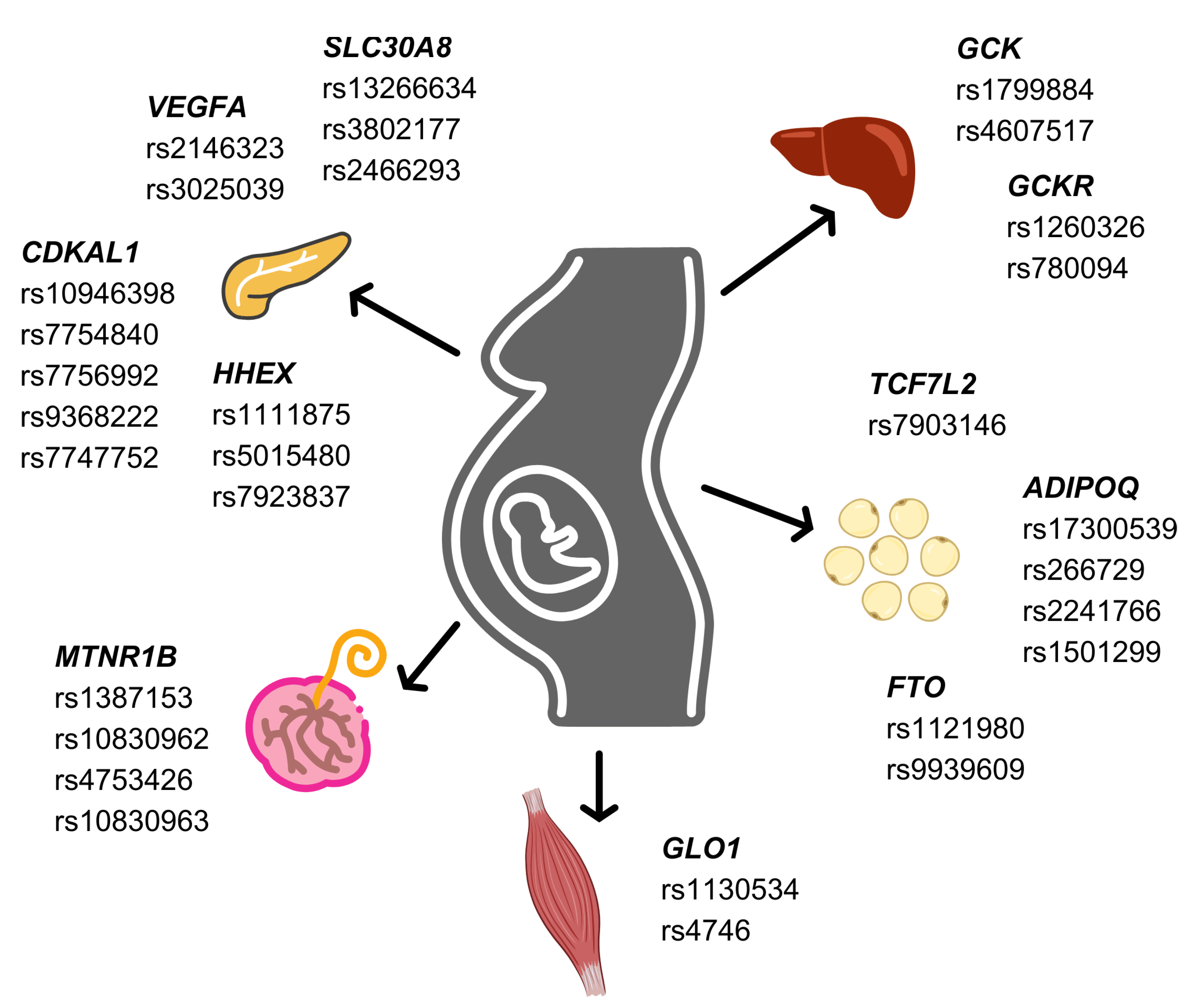
2. TCF7L2
3. HHEX
4. SLC30A8
5. ADIPOQ
6. FTO
7. VEGFA
8. CDKAL1
9. MTNR1B
10. GLO1
11. GCK and GCKR
12. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- ElSayed, N.A.; Aleppo, G.; Aroda, V.R.; Bannuru, R.R.; Brown, F.M.; Bruemmer, D.; Collins, B.S.; Hilliard, M.E.; Isaacs, D.; Johnson, E.L.; et al. 2. Classification and Diagnosis of Diabetes: Standards of Care in Diabetes-2023. Diabetes Care 2023, 46 (Suppl. S1), S19–S40. [Google Scholar] [CrossRef]
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Reynolds, C.M.; Vickers, M.H. The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3342. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Li, P. Novel single nucleotide polymorphisms in gestational diabetes mellitus. Clin. Chim. Acta 2023, 538, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Rauluseviciute, I.; Riudavets-Puig, R.; Blanc-Mathieu, R.; Castro-Mondragon, J.A.; Ferenc, K.; Kumar, V.; Lemma, R.B.; Lucas, J.; Cheneby, J.; Baranasic, D.; et al. JASPAR 2024: 20th anniversary of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2024, 52, D174–D182. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Del Bosque-Plata, L.; Martinez-Martinez, E.; Espinoza-Camacho, M.A.; Gragnoli, C. The Role of TCF7L2 in Type 2 Diabetes. Diabetes 2021, 70, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Chavarro, J.; Olsen, S.; Lin, Y.; Ley, S.H.; Bao, W.; Rawal, S.; Grunnet, L.G.; Thuesen, A.C.B.; Mills, J.L.; et al. Genetic variants of gestational diabetes mellitus: A study of 112 SNPs among 8722 women in two independent populations. Diabetologia 2018, 61, 1758–1768. [Google Scholar] [CrossRef]
- Chang, S.; Wang, Z.; Wu, L.; Lu, X.; Shangguan, S.; Xin, Y.; Li, L.; Wang, L. Association between TCF7L2 polymorphisms and gestational diabetes mellitus: A meta-analysis. J. Diabetes Investig. 2017, 8, 560–570. [Google Scholar] [CrossRef]
- Fang, C.; Wu, S.; Zhang, J.; Tian, Q.; Zhang, Z.; Wu, L. Impaired glucolipid metabolism in gestational diabetes mellitus with T variation of TCF7L2 rs7903146: A case–control study. Int. J. Diabetes Dev. Ctries. 2023, 1–8. [Google Scholar] [CrossRef]
- Zhang, P.; Deng, M.; Li, W.; Dai, Q.; He, H.; Zheng, W.; She, L.; Xiang, B.; Zeng, J.; Zhou, F.; et al. The correlation between transcription factor 7-like 2 gene polymorphisms and susceptibility of gestational diabetes mellitus in the population of central China: A case-control study. Front. Endocrinol. 2022, 13, 916590. [Google Scholar] [CrossRef]
- Franzago, M.; Fraticelli, F.; Nicolucci, A.; Celentano, C.; Liberati, M.; Stuppia, L.; Vitacolonna, E. Molecular Analysis of a Genetic Variants Panel Related to Nutrients and Metabolism: Association with Susceptibility to Gestational Diabetes and Cardiometabolic Risk in Affected Women. J. Diabetes Res. 2017, 2017, 4612623. [Google Scholar] [CrossRef]
- Verma, M.; Loh, N.Y.; Sabaratnam, R.; Vasan, S.K.; van Dam, A.D.; Todorcevic, M.; Neville, M.J.; Toledo, E.; Karpe, F.; Christodoulides, C. TCF7L2 plays a complex role in human adipose progenitor biology, which might contribute to genetic susceptibility to type 2 diabetes. Metabolism 2022, 133, 155240. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, R.N.; Warrington, N.M.; Cavadino, A.; Tyrrell, J.; Nodzenski, M.; Horikoshi, M.; Geller, F.; Myhre, R.; Richmond, R.C.; Paternoster, L.; et al. Genome-wide association study of offspring birth weight in 86 577 women identifies five novel loci and highlights maternal genetic effects that are independent of fetal genetics. Hum. Mol. Genet. 2018, 27, 742–756. [Google Scholar] [CrossRef]
- Tangjittipokin, W.; Chongjarean, N.; Plengvidhya, N.; Homsanit, M.; Yenchitsomanus, P.T. Transcription factor 7-like 2 (TCF7L2) variations associated with earlier age-onset of type 2 diabetes in Thai patients. J. Genet. 2012, 91, 251–255. [Google Scholar] [CrossRef] [PubMed]
- da Silva Xavier, G.; Mondragon, A.; Sun, G.; Chen, L.; McGinty, J.A.; French, P.M.; Rutter, G.A. Abnormal glucose tolerance and insulin secretion in pancreas-specific Tcf7l2-null mice. Diabetologia 2012, 55, 2667–2676. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.K.; Mondragon, A.; Chen, L.; McGinty, J.A.; French, P.M.; Ferrer, J.; Thorens, B.; Hodson, D.J.; Rutter, G.A.; Da Silva Xavier, G. Selective disruption of Tcf7l2 in the pancreatic beta cell impairs secretory function and lowers beta cell mass. Hum. Mol. Genet. 2015, 24, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.H.; Li, Y.L.; Liu, N.J.; Yang, Z.; Tao, X.M.; Du, Y.P.; Wang, X.C.; Lu, B.; Zhang, Z.Y.; Hu, R.M.; et al. TCF7L2 regulates pancreatic beta-cell function through PI3K/AKT signal pathway. Diabetol. Metab. Syndr. 2019, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- da Silva Xavier, G.; Mondragon, A.; Mourougavelou, V.; Cruciani-Guglielmacci, C.; Denom, J.; Herrera, P.L.; Magnan, C.; Rutter, G.A. Pancreatic alpha cell-selective deletion of Tcf7l2 impairs glucagon secretion and counter-regulatory responses to hypoglycaemia in mice. Diabetologia 2017, 60, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Facchinello, N.; Tarifeno-Saldivia, E.; Grisan, E.; Schiavone, M.; Peron, M.; Mongera, A.; Ek, O.; Schmitner, N.; Meyer, D.; Peers, B.; et al. Tcf7l2 plays pleiotropic roles in the control of glucose homeostasis, pancreas morphology, vascularization and regeneration. Sci. Rep. 2017, 7, 9605. [Google Scholar] [CrossRef]
- Chen, X.; Ayala, I.; Shannon, C.; Fourcaudot, M.; Acharya, N.K.; Jenkinson, C.P.; Heikkinen, S.; Norton, L. The Diabetes Gene and Wnt Pathway Effector TCF7L2 Regulates Adipocyte Development and Function. Diabetes 2018, 67, 554–568. [Google Scholar] [CrossRef]
- Geoghegan, G.; Simcox, J.; Seldin, M.M.; Parnell, T.J.; Stubben, C.; Just, S.; Begaye, L.; Lusis, A.J.; Villanueva, C.J. Targeted deletion of Tcf7l2 in adipocytes promotes adipocyte hypertrophy and impaired glucose metabolism. Mol. Metab. 2019, 24, 44–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; An, T.H.; Kim, H.; Jung, E.; Kim, G.; Oh, S.Y.; Kim, J.S.; Chun, H.J.; Jung, J.; Lee, E.W.; et al. Tcf7l2 in hepatocytes regulates de novo lipogenesis in diet-induced non-alcoholic fatty liver disease in mice. Diabetologia 2023, 66, 931–954. [Google Scholar] [CrossRef] [PubMed]
- Kuzmicki, M.; Telejko, B.; Wawrusiewicz-Kurylonek, N.; Kalejta, K.; Lemancewicz, A.; Zdrodowski, M.; Grabiec, M.; Pryszczepko-Wawreszuk, A.M.; Kretowski, A.; Gorska, M.; et al. The expression of transcription factor 7-like 2 (TCF7L2) in fat and placental tissue from women with gestational diabetes. Diabetes Res. Clin. Pract. 2011, 94, e43–e46. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Lu, F.; Dong, M.; Lin, Y.; Li, H.; Chen, J.; Shen, C.; Jin, G.; Hu, Z.; Shen, H. Genetic variants of IDE-KIF11-HHEX at 10q23.33 associated with type 2 diabetes risk: A fine-mapping study in Chinese population. PLoS ONE 2012, 7, e35060. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Zhang, L.; Wang, J.; Wang, Y. Association of HHEX and SLC30A8 Gene Polymorphisms with Gestational Diabetes Mellitus Susceptibility: A Meta-analysis. Biochem. Genet. 2023, 61, 2203–2221. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ding, Y.; Zhang, X.; Rao, J.; Yu, H.; Pan, H. The association between HHEX single-nucleotide polymorphism rs5015480 and gestational diabetes mellitus: A meta-analysis. Medicine 2020, 99, e19478. [Google Scholar] [CrossRef] [PubMed]
- Tarnowski, M.; Malinowski, D.; Safranow, K.; Dziedziejko, V.; Czerewaty, M.; Pawlik, A. Hematopoietically expressed homeobox (HHEX) gene polymorphism (rs5015480) is associated with increased risk of gestational diabetes mellitus. Clin. Genet. 2017, 91, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Cho, H.; Tayyebi, Z.; Shukla, A.; Luo, R.; Dixon, G.; Ursu, V.; Stransky, S.; Tremmel, D.M.; Sackett, S.D.; et al. CRISPR screening uncovers a central requirement for HHEX in pancreatic lineage commitment and plasticity restriction. Nat. Cell Biol. 2022, 24, 1064–1076. [Google Scholar] [CrossRef]
- Ito, R.; Kimura, A.; Hirose, Y.; Hatano, Y.; Mima, A.; Mae, S.I.; Keidai, Y.; Nakamura, T.; Fujikura, J.; Nishi, Y.; et al. Elucidation of HHEX in pancreatic endoderm differentiation using a human iPSC differentiation model. Sci. Rep. 2023, 13, 8659. [Google Scholar] [CrossRef]
- Zhang, J.; McKenna, L.B.; Bogue, C.W.; Kaestner, K.H. The diabetes gene Hhex maintains delta-cell differentiation and islet function. Genes Dev. 2014, 28, 829–834. [Google Scholar] [CrossRef]
- Liang, X.; Duan, H.; Mao, Y.; Koestner, U.; Wei, Y.; Deng, F.; Zhuang, J.; Li, H.; Wang, C.; Hernandez-Miranda, L.R.; et al. The SNAG Domain of Insm1 Regulates Pancreatic Endocrine Cell Differentiation and Represses beta- to delta-Cell Transdifferentiation. Diabetes 2021, 70, 1084–1097. [Google Scholar] [CrossRef]
- Evseeva, M.N.; Dyikanov, D.T.; Karagyaur, M.N.; Prikazchikova, T.A.; Sheptulina, A.F.; Balashova, M.S.; Zatsepin, T.S.; Rubtsov, Y.P.; Kulebyakin, K.Y. Hematopoietically-expressed homeobox protein HHEX regulates adipogenesis in preadipocytes. Biochimie 2021, 185, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Rajaraman, G.; Murthi, P.; Quinn, L.; Brennecke, S.P.; Kalionis, B. Homeodomain protein HLX is expressed primarily in cytotrophoblast cell types in the early pregnancy human placenta. Reprod. Fertil. Dev. 2008, 20, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Chimienti, F.; Devergnas, S.; Favier, A.; Seve, M. Identification and cloning of a beta-cell-specific zinc transporter, ZnT-8, localized into insulin secretory granules. Diabetes 2004, 53, 2330–2337. [Google Scholar] [CrossRef] [PubMed]
- Benny, P.; Ahn, H.J.; Burlingame, J.; Lee, M.J.; Miller, C.; Chen, J.; Urschitz, J. Genetic risk factors associated with gestational diabetes in a multi-ethnic population. PLoS ONE 2021, 16, e0261137. [Google Scholar] [CrossRef] [PubMed]
- Dereke, J.; Palmqvist, S.; Nilsson, C.; Landin-Olsson, M.; Hillman, M. The prevalence and predictive value of the SLC30A8 R325W polymorphism and zinc transporter 8 autoantibodies in the development of GDM and postpartum type 1 diabetes. Endocrine 2016, 53, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wang, Y.; Zhang, B.; Jin, Z. Association of type 2 diabetes susceptible genes GCKR, SLC30A8, and FTO polymorphisms with gestational diabetes mellitus risk: A meta-analysis. Endocrine 2018, 62, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Tan, B.; Han, F.; Huang, X.; Huang, J.; Wei, Y.; Guo, R. Association of solute carrier family 30 A8 zinc transporter gene variations with gestational diabetes mellitus risk in a Chinese population. Front. Endocrinol. 2023, 14, 1159714. [Google Scholar] [CrossRef] [PubMed]
- Rosta, K.; Al-Aissa, Z.; Hadarits, O.; Harreiter, J.; Nadasdi, A.; Kelemen, F.; Bancher-Todesca, D.; Komlosi, Z.; Nemeth, L.; Rigo, J.; et al. Association Study with 77 SNPs Confirms the Robust Role for the rs10830963/G of MTNR1B Variant and Identifies Two Novel Associations in Gestational Diabetes Mellitus Development. PLoS ONE 2017, 12, e0169781. [Google Scholar] [CrossRef]
- Zhang, Y.; Jian, W.; He, L.; Wu, J. ZnT8 in T2D: A novel therapeutic target for maintaining insulin secretion capacity. Acta Biochim. Biophys. Sin. 2020, 52, 1050–1051. [Google Scholar] [CrossRef]
- Ma, Q.; Xiao, Y.; Xu, W.; Wang, M.; Li, S.; Yang, Z.; Xu, M.; Zhang, T.; Zhang, Z.N.; Hu, R.; et al. ZnT8 loss-of-function accelerates functional maturation of hESC-derived beta cells and resists metabolic stress in diabetes. Nat. Commun. 2022, 13, 4142. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, O.P.; Lehtovirta, M.; Hastoy, B.; Chandra, V.; Krentz, N.A.J.; Kleiner, S.; Jain, D.; Richard, A.M.; Abaitua, F.; Beer, N.L.; et al. Loss of ZnT8 function protects against diabetes by enhanced insulin secretion. Nat. Genet. 2019, 51, 1596–1606. [Google Scholar] [CrossRef] [PubMed]
- Moyce Gruber, B.L.; Dolinsky, V.W. The Role of Adiponectin during Pregnancy and Gestational Diabetes. Life 2023, 13, 301. [Google Scholar] [CrossRef] [PubMed]
- Tangjittipokin, W.; Thanatummatis, B.; Wardati, F.; Narkdontri, T.; Teerawattanapong, N.; Boriboonhirunsarn, D. The genetic polymorphisms and levels of adipokines and adipocytokines that influence the risk of developing gestational diabetes mellitus in Thai pregnant women. Gene 2023, 860, 147228. [Google Scholar] [CrossRef] [PubMed]
- Takhshid, M.A.; Haem, Z.; Aboualizadeh, F. The association of circulating adiponectin and +45 T/G polymorphism of adiponectin gene with gestational diabetes mellitus in Iranian population. J. Diabetes Metab. Disord. 2015, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Tangjittipokin, W.; Narkdontri, T.; Teerawattanapong, N.; Thanatummatis, B.; Wardati, F.; Sunsaneevithayakul, P.; Boriboonhirunsarn, D. The Variants in ADIPOQ are Associated with Maternal Circulating Adipokine Profile in Gestational Diabetes Mellitus. J. Multidiscip. Healthc. 2023, 16, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, A.; Teler, J.; Maciejewska, A.; Sawczuk, M.; Safranow, K.; Dziedziejko, V. Adiponectin and leptin gene polymorphisms in women with gestational diabetes mellitus. J. Assist. Reprod. Genet. 2017, 34, 511–516. [Google Scholar] [CrossRef]
- Bai, Y.; Tang, L.; Li, L.; Li, L. The roles of ADIPOQ rs266729 and MTNR1B rs10830963 polymorphisms in patients with gestational diabetes mellitus: A meta-analysis. Gene 2020, 730, 144302. [Google Scholar] [CrossRef]
- Zhu, M.; Lv, Y.; Peng, Y.; Wu, Y.; Feng, Y.; Jia, T.; Xu, S.; Li, S.; Wang, W.; Tian, J.; et al. GCKR and ADIPOQ gene polymorphisms in women with gestational diabetes mellitus. Acta Diabetol. 2023, 60, 1709–1718. [Google Scholar] [CrossRef]
- Beltcheva, O.; Boyadzhieva, M.; Angelova, O.; Mitev, V.; Kaneva, R.; Atanasova, I. The rs266729 single-nucleotide polymorphism in the adiponectin gene shows association with gestational diabetes. Arch. Gynecol. Obstet. 2014, 289, 743–748. [Google Scholar] [CrossRef]
- Hivert, M.F.; Scholtens, D.M.; Allard, C.; Nodzenski, M.; Bouchard, L.; Brisson, D.; Lowe, L.P.; McDowell, I.; Reddy, T.; Dastani, Z.; et al. Genetic determinants of adiponectin regulation revealed by pregnancy. Obesity 2017, 25, 935–944. [Google Scholar] [CrossRef]
- Dias, S.; Adam, S.; Rheeder, P.; Pheiffer, C. No Association Between ADIPOQ or MTHFR Polymorphisms and Gestational Diabetes Mellitus in South African Women. Diabetes Metab. Syndr. Obes. 2021, 14, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.T.; Wu, S.L.; Liao, X.; Ma, S.J.; Tan, H.Z. Adiponectin gene polymorphisms and risk of gestational diabetes mellitus: A meta-analysis. World J. Clin. Cases 2019, 7, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Low, C.F.; Mohd Tohit, E.R.; Chong, P.P.; Idris, F. Adiponectin SNP45TG is associated with gestational diabetes mellitus. Arch. Gynecol. Obstet. 2011, 283, 1255–1260. [Google Scholar] [CrossRef]
- Feng, Y.; Jiang, C.D.; Chang, A.M.; Shi, Y.; Gao, J.; Zhu, L.; Zhang, Z. Interactions among insulin resistance, inflammation factors, obesity-related gene polymorphisms, environmental risk factors, and diet in the development of gestational diabetes mellitus. J. Matern. Fetal. Neonatal. Med. 2019, 32, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Howlader, M.; Sultana, M.I.; Akter, F.; Hossain, M.M. Adiponectin gene polymorphisms associated with diabetes mellitus: A descriptive review. Heliyon 2021, 7, e07851. [Google Scholar] [CrossRef] [PubMed]
- Cakina, S.; Ulu, S.; Beyazit, F.; Özen, E.; Postacı, E.S. Association between adiponectin and gene polymorphism in gestational diabetes mellitus patients. Rev. Romana De Med. De Lab. 2023, 31, 119–124. [Google Scholar]
- Lan, N.; Lu, Y.; Zhang, Y.; Pu, S.; Xi, H.; Nie, X.; Liu, J.; Yuan, W. FTO—A Common Genetic Basis for Obesity and Cancer. Front. Genet. 2020, 11, 559138. [Google Scholar] [CrossRef]
- Merkestein, M.; Laber, S.; McMurray, F.; Andrew, D.; Sachse, G.; Sanderson, J.; Li, M.; Usher, S.; Sellayah, D.; Ashcroft, F.M.; et al. FTO influences adipogenesis by regulating mitotic clonal expansion. Nat. Commun. 2015, 6, 6792. [Google Scholar] [CrossRef]
- Fan, H.Q.; He, W.; Xu, K.F.; Wang, Z.X.; Xu, X.Y.; Chen, H. FTO Inhibits Insulin Secretion and Promotes NF-kappaB Activation through Positively Regulating ROS Production in Pancreatic beta cells. PLoS ONE 2015, 10, e0127705. [Google Scholar]
- Cao, M.; Zhang, L.; Chen, T.; Shi, A.; Xie, K.; Li, Z.; Xu, J.; Chen, Z.; Ji, C.; Wen, J. Genetic Susceptibility to Gestational Diabetes Mellitus in a Chinese Population. Front. Endocrinol. 2020, 11, 247. [Google Scholar] [CrossRef]
- Santos, K.D.; Rosado, E.L.; da Fonseca, A.C.P.; Belfort, G.P.; da Silva, L.B.G.; Ribeiro-Alves, M.; Zembrzuski, V.M.; Martinez, J.A.; Saunders, C. FTO and ADRB2 Genetic Polymorphisms Are Risk Factors for Earlier Excessive Gestational Weight Gain in Pregnant Women with Pregestational Diabetes Mellitus: Results of a Randomized Nutrigenetic Trial. Nutrients 2022, 14, 1050. [Google Scholar] [CrossRef] [PubMed]
- Saucedo, R.; Valencia, J.; Gutierrez, C.; Basurto, L.; Hernandez, M.; Puello, E.; Rico, G.; Vega, G.; Zarate, A. Gene variants in the FTO gene are associated with adiponectin and TNF-alpha levels in gestational diabetes mellitus. Diabetol. Metab. Syndr. 2017, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Long, W.; Zhou, W.; Zhang, B.; Liu, J.; Yu, B. FTO, GCKR, CDKAL1 and CDKN2A/B gene polymorphisms and the risk of gestational diabetes mellitus: A meta-analysis. Arch. Gynecol. Obstet. 2018, 298, 705–715. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Cao, W.T.; Zeng, Y.H.; Huang, Z.Q.; Du, W.R.; Guan, N.D.; Zhao, Y.Z.; Wei, B.R.; Liu, Y.H.; Jing, C.X.; et al. Lack of associations between the FTO polymorphisms and gestational diabetes: A meta-analysis and trial sequential analysis. Gene 2018, 677, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Tarnowski, M.; Bujak, J.; Kopytko, P.; Majcher, S.; Ustianowski, P.; Dziedziejko, V.; Safranow, K.; Pawlik, A. Effect of FTO and IGF2BP2 gene polymorphisms on duration of pregnancy and Apgar scores in women with gestational diabetes. J. Obstet. Gynaecol. 2019, 39, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Amin, U.S.M.; Rahman, T.A.; Hasan, M.; Tofail, T.; Hasanat, M.A.; Seraj, Z.I.; Salimullah, M. Type 2 diabetes linked FTO gene variant rs8050136 is significantly associated with gravidity in gestational diabetes in a sample of Bangladeshi women: Meta-analysis and case-control study. PLoS ONE 2023, 18, e0288318. [Google Scholar] [CrossRef] [PubMed]
- Melincovici, C.S.; Bosca, A.B.; Susman, S.; Marginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Troncoso, F.; Acurio, J.; Herlitz, K.; Aguayo, C.; Bertoglia, P.; Guzman-Gutierrez, E.; Loyola, M.; Gonzalez, M.; Rezgaoui, M.; Desoye, G.; et al. Gestational diabetes mellitus is associated with increased pro-migratory activation of vascular endothelial growth factor receptor 2 and reduced expression of vascular endothelial growth factor receptor 1. PLoS ONE 2017, 12, e0182509. [Google Scholar] [CrossRef]
- Meng, Q.; Shao, L.; Luo, X.; Mu, Y.; Xu, W.; Gao, L.; Xu, H.; Cui, Y. Expressions of VEGF-A and VEGFR-2 in placentae from GDM pregnancies. Reprod. Biol. Endocrinol. 2016, 14, 61. [Google Scholar] [CrossRef]
- Dong, P.P. Association of vascular endothelial growth factor expression and polymorphisms with the risk of gestational diabetes mellitus. J. Clin. Lab. Anal. 2019, 33, e22686. [Google Scholar] [CrossRef]
- Sirico, A.; Rossi, E.D.; Degennaro, V.A.; Arena, V.; Rizzi, A.; Tartaglione, L.; Di Leo, M.; Pitocco, D.; Lanzone, A. Placental diabesity: Placental VEGF and CD31 expression according to pregestational BMI and gestational weight gain in women with gestational diabetes. Arch. Gynecol. Obstet. 2023, 307, 1823–1831. [Google Scholar] [CrossRef]
- Yang, W.; Jiang, Y.; Wang, Y.; Zhang, T.; Liu, Q.; Wang, C.; Swisher, G.; Wu, N.; Chao, C.; Prasadan, K.; et al. Placental growth factor in beta cells plays an essential role in gestational beta-cell growth. BMJ Open Diabetes Res. Care 2020, 8, e000921. [Google Scholar] [CrossRef] [PubMed]
- Staels, W.; Heremans, Y.; Leuckx, G.; Van Gassen, N.; Salinno, C.; De Groef, S.; Cools, M.; Keshet, E.; Dor, Y.; Heimberg, H.; et al. Conditional islet hypovascularisation does not preclude beta cell expansion during pregnancy in mice. Diabetologia 2017, 60, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Bolatai, A.; He, Y.; Wu, N. Vascular endothelial growth factor and its receptors regulation in gestational diabetes mellitus and eclampsia. J. Transl. Med. 2022, 20, 400. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, C.; Das, N.; Saha, S.; Kundu, T.; Sircar, D.; Roy, P. Involvement of Cdkal1 in the etiology of type 2 diabetes mellitus and microvascular diabetic complications: A review. J. Diabetes Metab. Disord. 2022, 21, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Okamura, T.; Yanobu-Takanashi, R.; Takeuchi, F.; Isono, M.; Akiyama, K.; Shimizu, Y.; Goto, M.; Liang, Y.Q.; Yamamoto, K.; Katsuya, T.; et al. Deletion of CDKAL1 affects high-fat diet-induced fat accumulation and glucose-stimulated insulin secretion in mice, indicating relevance to diabetes. PLoS ONE 2012, 7, e49055. [Google Scholar] [CrossRef] [PubMed]
- Take, K.; Waki, H.; Sun, W.; Wada, T.; Yu, J.; Nakamura, M.; Aoyama, T.; Yamauchi, T.; Kadowaki, T. CDK5 Regulatory Subunit-Associated Protein 1-like 1 Negatively Regulates Adipocyte Differentiation through Activation of Wnt Signaling Pathway. Sci. Rep. 2017, 7, 7326. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.J.; Bruckner, R.J.; Paulo, J.A.; Kazak, L.; Long, J.Z.; Mina, A.I.; Deng, Z.; LeClair, K.B.; Hall, J.A.; Hong, S.; et al. Cdkal1, a type 2 diabetes susceptibility gene, regulates mitochondrial function in adipose tissue. Mol. Metab. 2017, 6, 1212–1225. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Y.; Song, L.P.; Wei, S.D.; Wen, X.L.; Liu, D.B. CDK5 Regulatory Subunit-Associated Protein 1-Like 1 Gene Polymorphisms and Gestational Diabetes Mellitus Risk: A Trial Sequential Meta-Analysis of 13,306 Subjects. Front. Endocrinol. 2021, 12, 722674. [Google Scholar] [CrossRef]
- Mahdizade, A.H.; Bahreiny, S.S.; Bastani, M.-N.; Dabbagh, M.R.; Aghaei, M.; Ali Malayeri, F.; YousefiFard, A.; Taghizadeh, E. The influence of CDKAL1 (rs7754840) gene polymorphism on susceptibility to gestational diabetes mellitus in pregnant women: A systematic review and meta-analysis. Int. J. Diabetes Dev. Ctries. 2023, 1–10. [Google Scholar] [CrossRef]
- Noury, A.E.; Azmy, O.; Alsharnoubi, J.; Salama, S.; Okasha, A.; Gouda, W. Variants of CDKAL1 rs7754840 (G/C) and CDKN2A/2B rs10811661 (C/T) with gestational diabetes: Insignificant association. BMC Res. Notes 2018, 11, 181. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, M.; Li, W.; Ping, F.; Hu, Y.; Ma, L.; Gao, J.; Liu, J. Association of six single nucleotide polymorphisms with gestational diabetes mellitus in a Chinese population. PLoS ONE 2011, 6, e26953. [Google Scholar] [CrossRef] [PubMed]
- Amin, U.S.M.; Parvez, N.; Rahman, T.A.; Hasan, M.R.; Das, K.C.; Jahan, S.; Hasanat, M.A.; Seraj, Z.I.; Salimullah, M. CDKAL1 gene rs7756992 A/G and rs7754840 G/C polymorphisms are associated with gestational diabetes mellitus in a sample of Bangladeshi population: Implication for future T2DM prophylaxis. Diabetol. Metab. Syndr. 2022, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.F.; Xiao, X.H.; Zhang, Z.X.; Liu, Y.; Xu, T.; Zhu, X.L.; Zhang, Y.; Wu, X.P.; Li, W.H.; Zhang, H.B.; et al. Positive Association Between Type 2 Diabetes Risk Alleles Near CDKAL1 and Reduced Birthweight in Chinese Han Individuals. Chin. Med. J. 2015, 128, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, Q.; Feng, Y.; Yang, H.; Wu, W.; Zhang, P.; Wang, Y.; Ko, J.; Zhao, F.; Du, W.; et al. Single Nucleotide Polymorphisms in CDKAL1 Gene Are Associated with Risk of Gestational Diabetes Mellitus in Chinese Population. J. Diabetes Res. 2019, 2019, 3618103. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, W.; Liu, J.; Leng, J.; Li, W.; Yu, Z.; Li, J.; Ma, R.C.W.; Hu, G.; Fang, Z.; et al. Serum concentrations of SFAs and CDKAL1 single-nucleotide polymorphism rs7747752 are related to an increased risk of gestational diabetes mellitus. Am. J. Clin. Nutr. 2021, 114, 1698–1707. [Google Scholar] [CrossRef]
- Wang, H.; Li, J.; Leng, J.; Li, W.; Liu, J.; Yan, X.; Yu, Z.; Hu, G.; Ma, R.C.W.; Fang, Z.; et al. The CDKAL1 rs7747752-Bile Acids Interaction Increased Risk of Gestational Diabetes Mellitus: A Nested Case-Control Study. Front. Endocrinol. 2022, 13, 808956. [Google Scholar] [CrossRef]
- Ramos-Levi, A.; Barabash, A.; Valerio, J.; Garcia de la Torre, N.; Mendizabal, L.; Zulueta, M.; de Miguel, M.P.; Diaz, A.; Duran, A.; Familiar, C.; et al. Genetic variants for prediction of gestational diabetes mellitus and modulation of susceptibility by a nutritional intervention based on a Mediterranean diet. Front. Endocrinol. 2022, 13, 1036088. [Google Scholar] [CrossRef]
- Asghar, A.; Firasat, S.; Afshan, K.; Naz, S. Association of CDKAL1 gene polymorphism (rs10946398) with gestational diabetes mellitus in Pakistani population. Mol. Biol. Rep. 2023, 50, 57–64. [Google Scholar] [CrossRef]
- Tarnowski, M.; Malinowski, D.; Pawlak, K.; Dziedziejko, V.; Safranow, K.; Pawlik, A. GCK, GCKR, FADS1, DGKB/TMEM195 and CDKAL1 Gene Polymorphisms in Women with Gestational Diabetes. Can. J. Diabetes 2017, 41, 372–379. [Google Scholar] [CrossRef]
- Pervjakova, N.; Moen, G.H.; Borges, M.C.; Ferreira, T.; Cook, J.P.; Allard, C.; Beaumont, R.N.; Canouil, M.; Hatem, G.; Heiskala, A.; et al. Multi-ancestry genome-wide association study of gestational diabetes mellitus highlights genetic links with type 2 diabetes. Hum. Mol. Genet. 2022, 31, 3377–3391. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, Z.J.; Liu, H.Y.; Cai, J.; Lu, Q.K.; Ji, L.D.; Xu, J. The melatonin receptor 1B gene links circadian rhythms and type 2 diabetes mellitus: An evolutionary story. Ann. Med. 2023, 55, 1262–1286. [Google Scholar] [CrossRef]
- Wei, L.; Jiang, Y.; Gao, P.; Zhang, J.; Zhou, X.; Zhu, S.; Chen, Y.; Zhang, H.; Du, Y.; Fang, C.; et al. Role of melatonin receptor 1B gene polymorphism and its effect on the regulation of glucose transport in gestational diabetes mellitus. J. Zhejiang Univ. Sci. B 2023, 24, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, Y.; Qiao, B.; Xu, L.; Li, Y.; Li, C. Association Between a Melatonin Receptor 1B Genetic Polymorphism and Its Protein Expression in Gestational Diabetes Mellitus. Reprod. Sci. 2019, 26, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Shen, Y.; Shi, X.; Gu, X.; Zhang, P.; Liu, Y.; Zhu, A.; Jiang, L. MTNR1B gene on susceptibility to gestational diabetes mellitus: A two-stage hospital-based study in Southern China. Mol. Genet. Genom. 2020, 295, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Fei, X.; Li, M.; Zhang, Z.; Zhu, W.; Zhang, M.; Chen, X.; Xu, J.; Zhang, M.; Shen, Y.; et al. Associations of the MTNR1B rs10830963 and PPARG rs1801282 variants with gestational diabetes mellitus: A meta-analysis. Int. J. Diabetes Dev. Ctries. 2023, 43, 1029–1042. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, K.; Jin, L.; Zhou, Y.; Shang, X.; Wang, X.; Yu, H. MTNR1B gene variations and high pre-pregnancy BMI increase gestational diabetes mellitus risk in Chinese women. Gene 2023, 894, 148023. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, W.; Liu, B.; Dai, A.; Wang, Y.; She, L.; Zhang, P.; Zheng, W.; Dai, Q.; Yang, M. Melatonin Receptor 1B Genetic Variants on Susceptibility to Gestational Diabetes Mellitus: A Hospital-Based Case-Control Study in Wuhan, Central China. Diabetes Metab. Syndr. Obes. 2022, 15, 1207–1216. [Google Scholar] [CrossRef]
- Shan, D.; Wang, A.; Yi, K. MTNR1B rs1387153 Polymorphism and Risk of Gestational Diabetes Mellitus: Meta-Analysis and Trial Sequential Analysis. Public Health Genom. 2023, 26, 201–211. [Google Scholar] [CrossRef]
- Xie, K.; Chen, T.; Zhang, Y.; Wen, J.; Cui, X.; You, L.; Zhu, L.; Xu, B.; Ji, C.; Guo, X. Association of rs10830962 polymorphism with gestational diabetes mellitus risk in a Chinese population. Sci. Rep. 2019, 9, 5357. [Google Scholar] [CrossRef]
- Liang, Z.; Liu, H.; Wang, L.; Chen, Y.; Zhou, T.; Heianza, Y.; Li, W.; Leng, J.; Wang, J.; Gao, R.; et al. Maternal MTNR1B genotype, maternal gestational weight gain, and childhood obesity. Am. J. Clin. Nutr. 2020, 111, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Mey, J.T.; Haus, J.M. Dicarbonyl Stress and Glyoxalase-1 in Skeletal Muscle: Implications for Insulin Resistance and Type 2 Diabetes. Front. Cardiovasc. Med. 2018, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Yang, T.; Wei, W.; Zou, D.; Wei, Y.; Han, F.; He, J.; Huang, J.; Guo, R. Association between GLO1 variants and gestational diabetes mellitus susceptibility in a Chinese population: A preliminary study. Front. Endocrinol. 2023, 14, 1235581. [Google Scholar] [CrossRef] [PubMed]
- Gale, C.P.; Futers, T.S.; Summers, L.K. Common polymorphisms in the glyoxalase-1 gene and their association with pro-thrombotic factors. Diab. Vasc. Dis. Res. 2004, 1, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Matschinsky, F.M.; Magnuson, M.A.; Zelent, D.; Jetton, T.L.; Doliba, N.; Han, Y.; Taub, R.; Grimsby, J. The network of glucokinase-expressing cells in glucose homeostasis and the potential of glucokinase activators for diabetes therapy. Diabetes 2006, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Matschinsky, F.M.; Wilson, D.F. The Central Role of Glucokinase in Glucose Homeostasis: A Perspective 50 Years After Demonstrating the Presence of the Enzyme in Islets of Langerhans. Front. Physiol. 2019, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Chakera, A.J.; Steele, A.M.; Gloyn, A.L.; Shepherd, M.H.; Shields, B.; Ellard, S.; Hattersley, A.T. Recognition and Management of Individuals With Hyperglycemia Because of a Heterozygous Glucokinase Mutation. Diabetes Care 2015, 38, 1383–1392. [Google Scholar] [CrossRef]
- She, L.; Li, W.; Guo, Y.; Zhou, J.; Liu, J.; Zheng, W.; Dai, A.; Chen, X.; Wang, P.; He, H.; et al. Association of glucokinase gene and glucokinase regulatory protein gene polymorphisms with gestational diabetes mellitus: A case-control study. Gene 2022, 824, 146378. [Google Scholar] [CrossRef]
- Popova, P.V.; Klyushina, A.A.; Vasilyeva, L.B.; Tkachuk, A.S.; Bolotko, Y.A.; Gerasimov, A.S.; Pustozerov, E.A.; Kravchuk, E.N.; Predeus, A.; Kostareva, A.A.; et al. Effect of gene-lifestyle interaction on gestational diabetes risk. Oncotarget 2017, 8, 112024–112035. [Google Scholar] [CrossRef]
- Ao, D.; Zhao, Q.; Song, J.Y.; Liu, Z.; Wang, Y.; Wang, H.J.; Yang, H.X. The association of the glucokinase rs4607517 polymorphism with gestational diabetes mellitus and its interaction with sweets consumption in Chinese women. Public Health Nutr. 2021, 24, 2563–2569. [Google Scholar] [CrossRef]
- Anghebem-Oliveira, M.I.; Webber, S.; Alberton, D.; de Souza, E.M.; Klassen, G.; Picheth, G.; Rego, F.G. The GCKR Gene Polymorphism rs780094 is a Risk Factor for Gestational Diabetes in a Brazilian Population. J. Clin. Lab. Anal. 2017, 31, e22035. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Genes | SNPs | Location | Risk Allele | Predicted TF (JASPAR) | Expression in Placenta |
|---|---|---|---|---|---|
| TCF7L2 | rs34872471 | Intron 4 | C | ZNF354A, Six3, JUND | Y |
| rs7901695 | Intron 4 | C | n/a | ||
| rs4506565 | Intron 4 | T | PLAGL2 | ||
| rs7903146 | Intron 4 | T | ZNF211 | ||
| rs12243326 | Intron 4 | C | ZNF211 | ||
| rs12255372 | Intron 5 | T | n/a | ||
| rs290487 | Intron 8 | T | NFIC::TLX1 | ||
| HHEX | rs1111875 | Intergenic | C | ETV2::FOXI1, ERF::FOXI1, FOXO1::FLI1, SPIB, GABPA, Erg | |
| rs5015480 | Intergenic | C | n/a | Y | |
| rs7923837 | Intergenic | G | FOXO, FOXA, FOXP | ||
| SLC30A8 | rs13266634 | Missense | C | PAX1, PAX2, PAX9 | N |
| rs3802177 | 3′ UTR | G | PAX1, PAX9 | ||
| rs2466293 | 3′ UTR | G | NR1D2, RXRA::VDR | ||
| ADIPOQ | rs17300539 | Promoter | A | n/a | |
| rs266729 | Promoter | G | THAP1 | ||
| rs2241766 | Exon 2 | G | Tfcp2l1, RARB, RARG | ||
| rs1501299 | Intron 2 | T | ZNF354A, POU4F3 | ||
| FTO | rs1421085 | Intron 2 | No association | ONECUT1, ONECUT3, CUX1, CUX2 | Y |
| rs1121980 | Intron 2 | G | SIX2 | ||
| rs8050136 | Intron 2 | No association | ONECUT1, ONECUT2, CUX1 | ||
| rs9939609 | Intron 2 | A | POU2F1::SOX2 | ||
| VEGFA | rs2010963 | Exon 1 | No association | ZNF701, IRF2 | Y |
| rs833069 | Intron 2 | No association | ZNF257, ZKSCAN5, ZNF701, ZNF263, SP4, SP5 | ||
| rs2146323 | Intron 2 | A | Zfp335 | ||
| rs3025010 | Intron5 | No association | Ebf2, EBF3, ZNF449, ZNF682, KLF3, NFIC, ZNF324 | ||
| rs3025039 | Exon 7 | T | Bach1::Mafk, Mafg, MAFG::NFE2L, MAFK, MAF::NFE2 | ||
| CDKAL1 | rs10946398 | Intron 5 | C | CREB1, MAF | Y |
| rs7754840 | Intron 5 | C | n/a | ||
| rs7756992 | Intron 5 | G | Irf1, IRF7, MEF2A, MEF2C, MEF2D | ||
| rs9368222 | Intron 5 | A | SOX4, SOX10, SOX 12, Sox 11, Sox6 | ||
| rs7747752 | Intron 5 | C | MGA | ||
| MTNR1B | rs1387153 | Intergenic | T | NR5A1, ZBTB12, ZSCAN31, THAP1 | Y |
| rs10830962 | Intergenic | G | MEF2B, MEF2D, Rhox11, CDX2 | ||
| rs4753426 | Promoter | C | n/a | ||
| rs10830963 | Intron 1 | G | n/a | ||
| GLO1 | rs1130534 | Exon 4 | T | n/a | Y |
| rs4746 | Exon 4 | T | n/a | ||
| rs1781735 | Promoter | No association | n/a | ||
| GCK | rs1799884 | Promoter | T | n/a | Y |
| rs4607517 | Intergenic | A | PBX3, SREBF1, PKNOX1, Neurod2 | ||
| GCKR | rs1260326 | Missense | C | n/a | Y |
| rs780094 | Intron 16 | C | NRL, TBX19, TBXT, PKNOX1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suthon, S.; Tangjittipokin, W. Mechanisms and Physiological Roles of Polymorphisms in Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2024, 25, 2039. https://doi.org/10.3390/ijms25042039
Suthon S, Tangjittipokin W. Mechanisms and Physiological Roles of Polymorphisms in Gestational Diabetes Mellitus. International Journal of Molecular Sciences. 2024; 25(4):2039. https://doi.org/10.3390/ijms25042039
Chicago/Turabian StyleSuthon, Sarocha, and Watip Tangjittipokin. 2024. "Mechanisms and Physiological Roles of Polymorphisms in Gestational Diabetes Mellitus" International Journal of Molecular Sciences 25, no. 4: 2039. https://doi.org/10.3390/ijms25042039
APA StyleSuthon, S., & Tangjittipokin, W. (2024). Mechanisms and Physiological Roles of Polymorphisms in Gestational Diabetes Mellitus. International Journal of Molecular Sciences, 25(4), 2039. https://doi.org/10.3390/ijms25042039