


Application of Flow Cytometry Using Advanced Chromatin Analyses for Assessing Changes in Sperm Structure and DNA Integrity in a Porcine Model

 , ,
, ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Results for the Clustering of Motility Data
2.2. Extensive Modifications of Sperm Chromatin Are Evident after Long Refrigerated Storage, with a Relevant Boar Effect
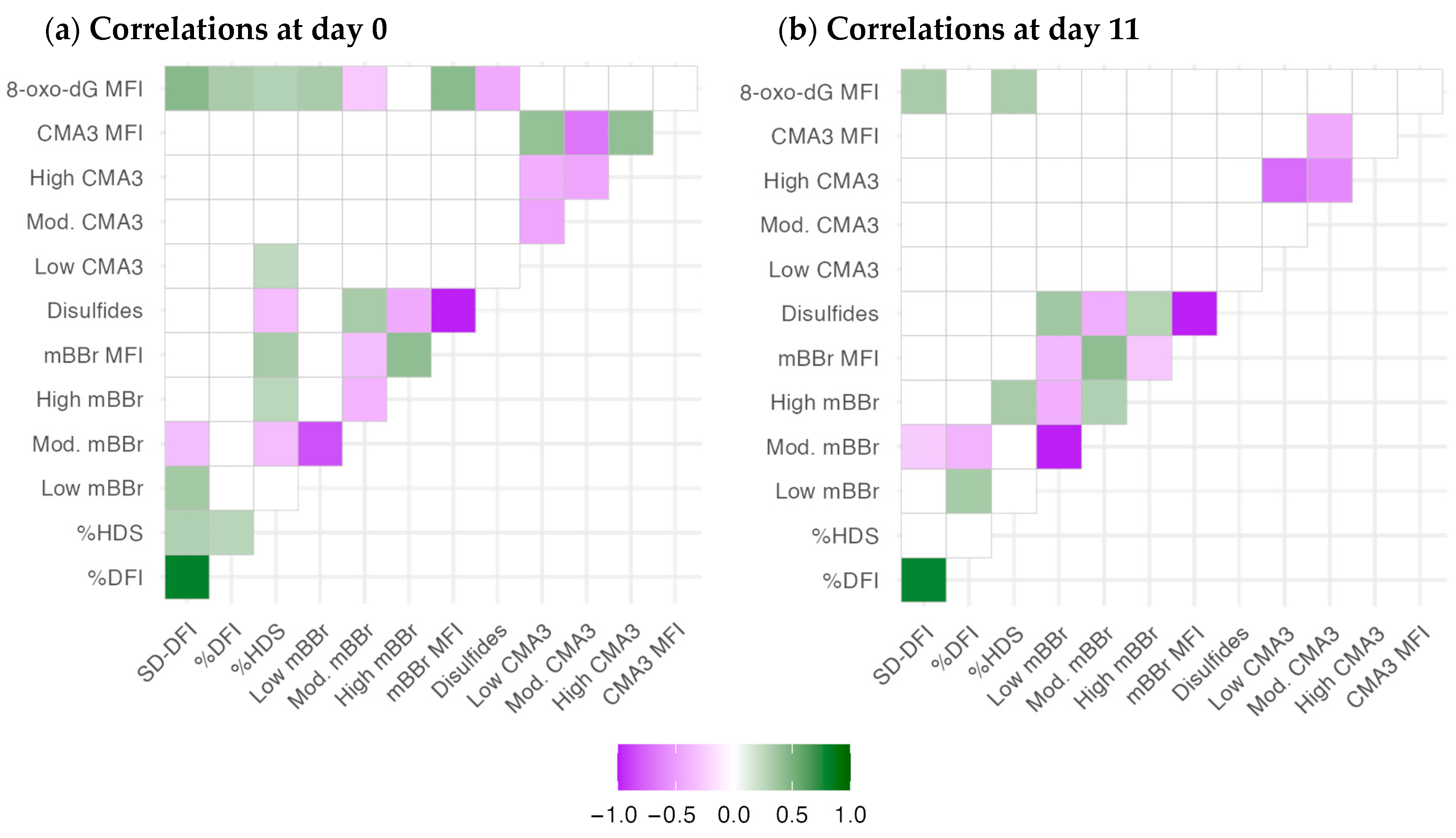
2.3. Chromatin Structure Variables Are Related to Different Nuclear Features
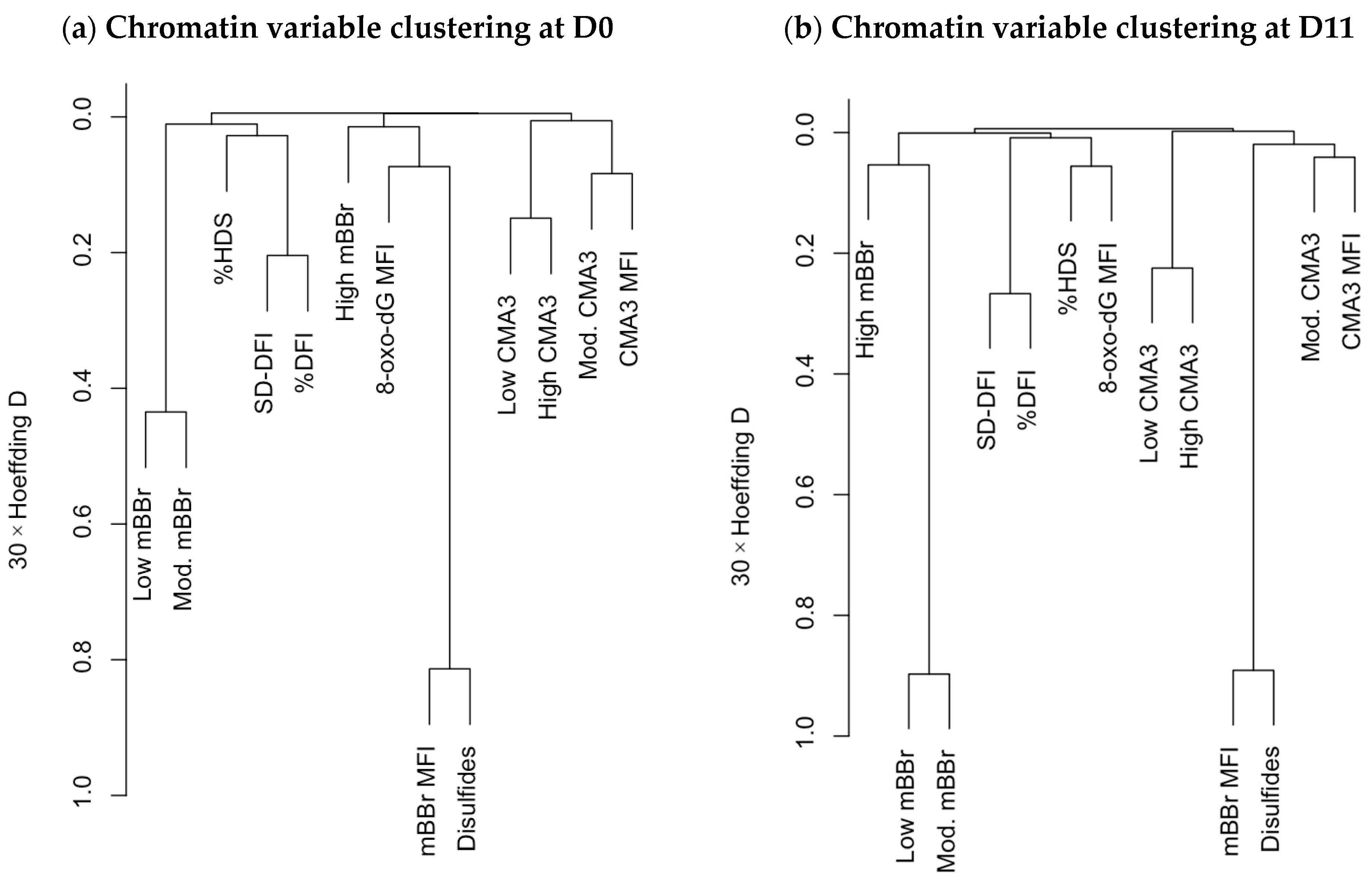
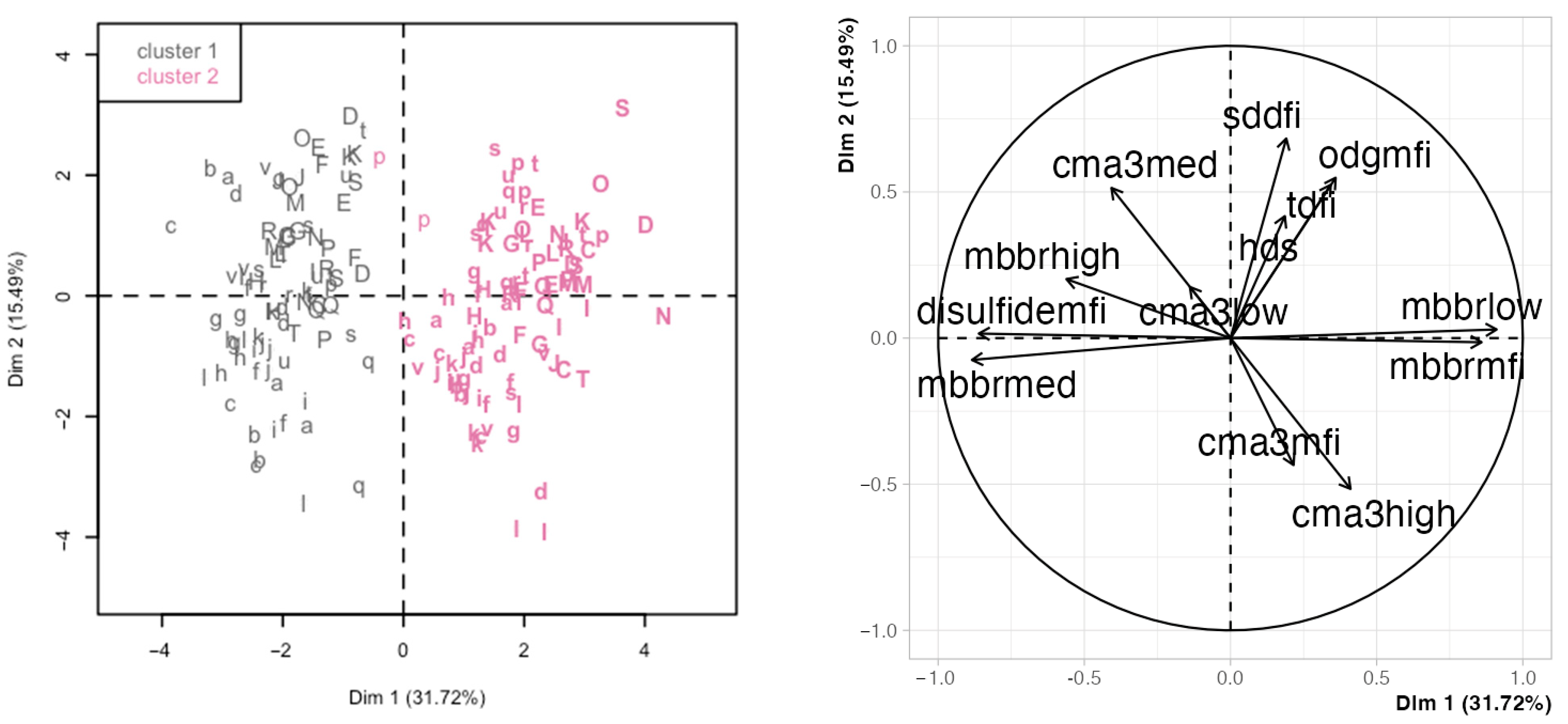
2.4. Hierarchical Clustering and Principal Components Analysis Unveils Relationships among Chromatin Variables and with Sperm Motility and Physiology Variables
3. Discussion
3.1. Analysis of the Chromatin Structure in the Pig Model
3.2. Detection of 8-oxo-dG as a Marker of DNA Oxidative Insult and a Proxy for SDF
3.3. CMA3 for the Evaluation of Chromatin Compaction in Pig Spermatozoa
3.4. Multivariate Analyses Enhance Discovering Variable Associations and Potential New Research Avenues
3.5. Implications for Animal Breeding and the Human Clinic
4. Materials and Methods
4.1. Solutions and Reagents
4.2. Animals and Samples Collection
4.3. Experimental Design
4.4. Sperm Functionality Assessment
4.5. Sperm Motility Analysis
4.6. Oxidative DNA Damage Analysis
4.7. Sperm Chromatin Structure Assay (SCSA)
4.8. Chromatin Disulfide Bridge Assessment
4.9. Chromatin Condensation Assay
4.10. Flow Cytometry Analyses
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamb, D.J.; Marinaro, J.A. Can Semen Parameters Predict Pregnancy Outcomes? Fertil. Steril. 2023, 120, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Maside, C.; Recuero, S.; Salas-Huetos, A.; Ribas-Maynou, J.; Yeste, M. Animal Board Invited Review: An Update on the Methods for Semen Quality Evaluation in Swine—From Farm to the Lab. Animal 2023, 17, 100720. [Google Scholar] [CrossRef] [PubMed]
- Conflitti, A.C.; Cicolani, G.; Buonacquisto, A.; Pallotti, F.; Faja, F.; Bianchini, S.; Blaconà, G.; Bruno, S.M.; Linari, A.; Lucarelli, M.; et al. Sperm DNA Fragmentation and Sperm-Borne miRNAs: Molecular Biomarkers of Embryo Development? Int. J. Mol. Sci. 2023, 24, 1007. [Google Scholar] [CrossRef] [PubMed]
- Llavanera, M.; Delgado-Bermúdez, A.; Ribas-Maynou, J.; Salas-Huetos, A.; Yeste, M. A Systematic Review Identifying Fertility Biomarkers in Semen: A Clinical Approach through Omics to Diagnose Male Infertility. Fertil. Steril. 2022, 118, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Maynou, J.; Novo, S.; Torres, M.; Salas-Huetos, A.; Rovira, S.; Antich, M.; Yeste, M. Sperm DNA Integrity Does Play a Crucial Role for Embryo Development after ICSI, Notably When Good-Quality Oocytes from Young Donors Are Used. Biol. Res. 2022, 55, 41. [Google Scholar] [CrossRef] [PubMed]
- Kumaresan, A.; Das Gupta, M.; Datta, T.K.; Morrell, J.M. Sperm DNA Integrity and Male Fertility in Farm Animals: A Review. Front. Vet. Sci. 2020, 7, 321. [Google Scholar] [CrossRef] [PubMed]
- Santi, D.; Spaggiari, G.; Simoni, M. Sperm DNA Fragmentation Index as a Promising Predictive Tool for Male Infertility Diagnosis and Treatment Management—Meta-Analyses. Reprod. BioMedicine Online 2018, 37, 315–326. [Google Scholar] [CrossRef]
- Batista, C.; van Lier, E.; Petrocelli, H. Dynamics of Sperm DNA Fragmentation in Raw Boar Semen and Fertility. Reprod. Domest. Anim. 2016, 51, 774–780. [Google Scholar] [CrossRef]
- Vallet-Buisan, M.; Mecca, R.; Jones, C.; Coward, K.; Yeste, M. Contribution of Semen to Early Embryo Development: Fertilization and Beyond. Hum. Reprod. Update 2023, 29, 395–433. [Google Scholar] [CrossRef]
- Moritz, L.; Hammoud, S.S. The Art of Packaging the Sperm Genome: Molecular and Structural Basis of the Histone-To-Protamine Exchange. Front. Endocrinol. 2022, 13, 895502. [Google Scholar] [CrossRef]
- Ribas-Maynou, J.; Nguyen, H.; Wu, H.; Ward, W.S. Functional Aspects of Sperm Chromatin Organization. Results Probl. Cell Differ. 2022, 70, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R. Protamines and Male Infertility. Hum. Reprod. Update 2006, 12, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Brinkworth, M.; Iles, D. Paternal DNA Packaging in Spermatozoa: More than the Sum of Its Parts? DNA, Histones, Protamines and Epigenetics. Reproduction 2010, 139, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Gosálvez, J.; López-Fernández, C.; Fernández, J.L.; Gouraud, A.; Holt, W.V. Relationships between the Dynamics of Iatrogenic DNA Damage and Genomic Design in Mammalian Spermatozoa from Eleven Species. Mol. Reprod. Dev. 2011, 78, 951–961. [Google Scholar] [CrossRef]
- Didion, B.A.; Kasperson, K.M.; Wixon, R.L.; Evenson, D.P. Boar Fertility and Sperm Chromatin Structure Status: A Retrospective Report. J. Androl. 2009, 30, 655–660. [Google Scholar] [CrossRef]
- Boe-Hansen, G.B.; Christensen, P.; Vibjerg, D.; Nielsen, M.B.F.; Hedeboe, A.M. Sperm Chromatin Structure Integrity in Liquid Stored Boar Semen and Its Relationships with Field Fertility. Theriogenology 2008, 69, 728–736. [Google Scholar] [CrossRef]
- Broekhuijse, M.L.W.J.; Šoštarić, E.; Feitsma, H.; Gadella, B.M. Relationship of Flow Cytometric Sperm Integrity Assessments with Boar Fertility Performance under Optimized Field Conditions. J. Anim. Sci. 2012, 90, 4327–4336. [Google Scholar] [CrossRef] [PubMed]
- Myromslien, F.D.; Tremoen, N.H.; Andersen-Ranberg, I.; Fransplass, R.; Stenseth, E.B.; Zeremichael, T.T.; van Son, M.; Grindflek, E.; Gaustad, A.H. Sperm DNA Integrity in Landrace and Duroc Boar Semen and Its Relationship to Litter Size. Reprod. Domest. Anim. 2019, 54, 160–166. [Google Scholar] [CrossRef]
- Mateo-Otero, Y.; Llavanera, M.; Recuero, S.; Delgado-Bermúdez, A.; Barranco, I.; Ribas-Maynou, J.; Yeste, M. Sperm DNA Damage Compromises Embryo Development, but Not Oocyte Fertilisation in Pigs. Biol. Res. 2022, 55, 15. [Google Scholar] [CrossRef]
- Evenson, D.P.; Thompson, L.; Jost, L. Flow Cytometric Evaluation of Boar Semen by the Sperm Chromatin Structure Assay as Related to Cryopreservation and Fertility. Theriogenology 1994, 41, 637–651. [Google Scholar] [CrossRef]
- Virro, M.R.; Larson-Cook, K.L.; Evenson, D.P. Sperm Chromatin Structure Assay (SCSA®) Parameters Are Related to Fertilization, Blastocyst Development, and Ongoing Pregnancy in in Vitro Fertilization and Intracytoplasmic Sperm Injection Cycles. Fertil. Steril. 2004, 81, 1289–1295. [Google Scholar] [CrossRef]
- Manicardi, G.C.; Bianchi, P.G.; Pantano, S.; Azzoni, P.; Bizzaro, D.; Bianchi, U.; Sakkas, D. Presence of Endogenous Nicks in DNA of Ejaculated Human Spermatozoa and Its Relationship to Chromomycin A3 Accessibility. Biol. Reprod. 1995, 52, 864–867. [Google Scholar] [CrossRef]
- Lolis, D.; Georgiou, I.; Syrrou, M.; Zikopoulos, K.; Konstantelli, M.; Messinis, I. Chromomycin A3-Staining as an Indicator of Protamine Deficiency and Fertilization. Int. J. Androl. 1996, 19, 23–27. [Google Scholar] [CrossRef]
- Iranpour, F.G.; Nasr-Esfahani, M.H.; Valojerdi, M.R.; Taki Al-Taraihi, T.M. Chromomycin A3 Staining as a Useful Tool for Evaluation of Male Fertility. J. Assist. Reprod. Genet. 2000, 17, 60–66. [Google Scholar] [CrossRef]
- Ménézo, Y. Evaluation of the Human Sperm Nucleus: Ambiguity and Risk of Confusion with Chromomycin Staining. Zygote 2021, 29, 257–259. [Google Scholar] [CrossRef]
- Noblanc, A.; Kocer, A.; Chabory, E.; Vernet, P.; Saez, F.; Cadet, R.; Conrad, M.; Drevet, J.R. Glutathione Peroxidases at Work on Epididymal Spermatozoa: An Example of the Dual Effect of Reactive Oxygen Species on Mammalian Male Fertilizing Ability. J. Androl. 2011, 32, 641–650. [Google Scholar] [CrossRef]
- Betarelli, R.P.; Rocco, M.; Yeste, M.; Fernández-Novell, J.M.; Placci, A.; Azevedo Pereira, B.; Castillo-Martín, M.; Estrada, E.; Peña, A.; Zangeronimo, M.G.; et al. The Achievement of Boar Sperm in Vitro Capacitation Is Related to an Increase of Disrupted Disulphide Bonds and Intracellular Reactive Oxygen Species Levels. Andrology 2018, 6, 781–797. [Google Scholar] [CrossRef] [PubMed]
- Reyes, R.; Rosado, A.; Hernández, O.; Delgado, N.M. Heparin and Glutathione: Physiological Decondensing Agents of Human Sperm Nuclei. Gamete Res. 1989, 23, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Lacalle, E.; Fernández-Alegre, E.; Soriano-Úbeda, C.; Martínez-Martínez, S.; Domínguez, J.C.; González-Montaña, J.R.; Morrell, J.M.; Martínez-Pastor, F. Single Layer Centrifugation (SLC) for Bacterial Removal with Porcicoll Positively Modifies Chromatin Structure in Boar Spermatozoa. Theriogenology 2023, 201, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Lacalle, E.; Núñez, A.; Fernández-Alegre, E.; Crespo-Félez, I.; Domínguez, J.C.; Alonso, M.E.; González-Urdiales, R.; Martínez-Pastor, F. Cold-Shock Test Is a Practical Method for Selecting Boar Ejaculates Yielding Appropriate Seminal Plasma for Post-Thawing Supplementation. Animals 2021, 11, 871. [Google Scholar] [CrossRef] [PubMed]
- Zubkova, E.V.; Wade, M.; Robaire, B. Changes in Spermatozoal Chromatin Packaging and Susceptibility to Oxidative Challenge during Aging. Fertil. Steril. 2005, 84, 1191–1198. [Google Scholar] [CrossRef]
- Aitken, R.J.; Smith, T.B.; Jobling, M.S.; Baker, M.A.; De Iuliis, G.N. Oxidative Stress and Male Reproductive Health. Asian J. Androl. 2014, 16, 31–38. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, Y.; Duan, R.; Wei, H.; Jiang, S.; Peng, J. Lower Dietary N-6:N-3 Ratio and High-Dose Vitamin E Supplementation Improve Sperm Morphology and Oxidative Stress in Boars. Reprod. Fertil. Dev. 2017, 29, 940–949. [Google Scholar] [CrossRef]
- De Iuliis, G.N.; Thomson, L.K.; Mitchell, L.A.; Finnie, J.M.; Koppers, A.J.; Hedges, A.; Nixon, B.; Aitken, R.J. DNA Damage in Human Spermatozoa Is Highly Correlated with the Efficiency of Chromatin Remodeling and the Formation of 8-Hydroxy-2′-Deoxyguanosine, a Marker of Oxidative Stress. Biol. Reprod. 2009, 81, 517–524. [Google Scholar] [CrossRef]
- Ramón, M.; Pérez-Guzmán, M.D.; Jiménez-Rabadán, P.; Esteso, M.C.; García-Álvarez, O.; Maroto-Morales, A.; Anel-López, L.; Soler, A.J.; Fernández-Santos, M.R.; Garde, J.J. Sperm Cell Population Dynamics in Ram Semen during the Cryopreservation Process. PLoS ONE 2013, 8, e59189. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gago, R.; Domínguez, J.C.; Martínez-Pastor, F. Seminal Plasma Applied Post-Thawing Affects Boar Sperm Physiology: A Flow Cytometry Study. Theriogenology 2013, 80, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Corzett, M.; Mazrimas, J.; Balhorn, R. Protamine 1: Protamine 2 Stoichiometry in the Sperm of Eutherian Mammals. Mol. Reprod. Dev. 2002, 61, 519–527. [Google Scholar] [CrossRef]
- Ausejo, R.; Martínez, J.M.; Mendoza, N.; Bolarin, A.; Tejedor, M.T.; Falceto, M.V. Nuclear DNA Fragmentation in Boar Spermatozoa: Measurement Methods and Reproductive Performance Implications. Front. Vet. Sci. 2022, 9, 929858. [Google Scholar] [CrossRef] [PubMed]
- Czubaszek, M.; Andraszek, K.; Banaszewska, D. Influence of the Age of the Individual on the Stability of Boar Sperm Genetic Material. Theriogenology 2020, 147, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Maynou, J.; Garcia-Bonavila, E.; Bonet, S.; Catalán, J.; Salas-Huetos, A.; Yeste, M. The TUNEL Assay Underestimates the Incidence of DNA Damage in Pig Sperm Due to Chromatin Condensation: The TUNEL Assay Underestimates Sperm DNA Fragmentation. Theriogenology 2021, 174, 94–101. [Google Scholar] [CrossRef]
- Soria-meneses, P.J.; Jurado-campos, A.; Gómez-Rubio, V.; Sánchez-Ajofrín, I.; Soler, A.J.; Garde, J.J.; Fernández-Santos, M.d.R. Determination of Ram (Ovis aries) Sperm DNA Damage Due to Oxidative Stress: 8-OHdG Immunodetection Assay vs. SCSA®. Animals 2022, 12, 3286. [Google Scholar] [CrossRef]
- Lacalle, E.; Martínez-Martínez, S.; Fernández-Alegre, E.; Soriano-Úbeda, C.; Morrell, J.M.; Martínez-Pastor, F. Low-Density Colloid Centrifugation Removes Bacteria from Boar Semen Doses after Spiking with Selected Species. Res. Vet. Sci. 2023, 158, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Viñolas-Vergés, E.; Ribas-Maynou, J.; Barranco, I.; Peres Rubio, C.; Bonet, S.; Roca, J.; Yeste, M. Chromatin Condensation but Not DNA Integrity of Pig Sperm Is Greater in the Sperm-Rich Fraction. J. Anim. Sci. Biotechnol. 2023, 14, 139. [Google Scholar] [CrossRef] [PubMed]
- Peña, S.T.; Gummow, B.; Parker, A.J.; Paris, D.B.B.P. Revisiting Summer Infertility in the Pig: Could Heat Stress-Induced Sperm DNA Damage Negatively Affect Early Embryo Development? Anim. Prod. Sci. 2017, 57, 1975–1983. [Google Scholar] [CrossRef]
- Luño, V.; Gil, L.; Olaciregui, M.; Jerez, R.A.; de Blas, I.; Hozbor, F. Antioxidant Effect of Lemon Balm (Melissa Officinalis) and Mate Tea (Ilex Paraguensys) on Quality, Lipid Peroxidation and DNA Oxidation of Cryopreserved Boar Epididymal Spermatozoa. Andrologia 2015, 47, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gago, R.; Álvarez-Rodríguez, M.; Alonso, M.E.; Ramiro González, J.; Alegre, B.; Domínguez, J.C.; Martínez-Pastor, F. Thawing Boar Semen in the Presence of Seminal Plasma Improves Motility, Modifies Subpopulation Patterns and Reduces Chromatin Alterations. Reprod. Fertil. Dev. 2017, 29, 1576–1584. [Google Scholar] [CrossRef]
- Shalgi, R.; Seligman, J.; Kosower, N.S. Dynamics of the Thiol Status of Rat Spermatozoa during Maturation: Analysis with the Fluorescent Labeling Agent Monobromobimane. Biol. Reprod. 1989, 40, 1037–1045. [Google Scholar] [CrossRef]
- Hamilton, L.E.E.; Oko, R.; Miranda-Vizuete, A.; Sutovsky, P. Sperm Redox System Equilibrium: Implications for Fertilization and Male Fertility. Adv. Exp. Med. Biol. 2022, 1358, 345–367. [Google Scholar] [CrossRef]
- Peña, S.T.; Stone, F.; Gummow, B.; Parker, A.J.; Paris, D.B.B.P. Tropical Summer Induces DNA Fragmentation in Boar Spermatozoa: Implications for Evaluating Seasonal Infertility. Reprod. Fertil. Dev. 2019, 31, 590–601. [Google Scholar] [CrossRef]
- Abah, K.O.; Fontbonne, A.; Partyka, A.; Nizanski, W. Effect of Male Age on Semen Quality in Domestic Animals: Potential for Advanced Functional and Translational Research? Vet. Res. Commun. 2023, 47, 1125–1137. [Google Scholar] [CrossRef]
- Martínez-Pastor, F.; Lacalle, E.; Martínez-Martínez, S.; Fernández-Alegre, E.; Álvarez-Fernández, L.; Martinez-Alborcia, M.J.; Bolarin, A.; Morrell, J.M. Low Density Porcicoll Separates Spermatozoa from Bacteria and Retains Sperm Quality. Theriogenology 2021, 165, 28–36. [Google Scholar] [CrossRef]
- Gill, K.; Rosiak, A.; Gaczarzewicz, D.; Jakubik, J.; Kurzawa, R.; Kazienko, A.; Rymaszewska, A.; Laszczynska, M.; Grochans, E.; Piasecka, M. The Effect of Human Sperm Chromatin Maturity on ICSI Outcomes. Hum. Cell 2018, 31, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Spiess, A.N.; Schuppe, H.C.; Steger, K. The Impact of Sperm Protamine Deficiency and Sperm DNA Damage on Human Male Fertility: A Systematic Review and Meta-Analysis. Andrology 2016, 4, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, T.R.D.S.; Siqueira, A.F.P.; De Castro, L.S.; Mendes, C.M.; Delgado, J.D.C.; De Assis, P.M.; Mesquita, L.P.; Maiorka, P.C.; Nichi, M.; Goissis, M.D.; et al. Effect of Heat Stress on Sperm DNA: Protamine Assessment in Ram Spermatozoa and Testicle. Oxidative Med. Cell. Longev. 2018, 2018, 5413056. [Google Scholar] [CrossRef]
- Ausejo, R.; Martínez, J.M.; Soler-llorens, P.; Bolarín, A.; Tejedor, T.; Falceto, M.V. Seasonal Changes of Nuclear DNA Fragmentation in Boar Spermatozoa in Spain. Animals 2021, 11, 465. [Google Scholar] [CrossRef]
- Peña, S.T.; Stone, F.; Gummow, B.; Parker, A.J.; Paris, D.B.B.P. Susceptibility of Boar Spermatozoa to Heat Stress Using in Vivo and in Vitro Experimental Models. Trop. Anim. Health Prod. 2021, 53, 97. [Google Scholar] [CrossRef]
- Narud, B.; Klinkenberg, G.; Khezri, A.; Zeremichael, T.T.; Stenseth, E.-B.; Nordborg, A.; Haukaas, T.H.; Morrell, J.M.; Heringstad, B.; Myromslien, F.D.; et al. Differences in Sperm Functionality and Intracellular Metabolites in Norwegian Red Bulls of Contrasting Fertility. Theriogenology 2020, 157, 24–32. [Google Scholar] [CrossRef]
- Evenson, D.P.; Djira, G.; Kasperson, K.; Christianson, J. Relationships between the Age of 25,445 Men Attending Infertility Clinics and Sperm Chromatin Structure Assay (SCSA®) Defined Sperm DNA and Chromatin Integrity. Fertil. Steril. 2020, 114, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A.J. Usefulness of Bovine and Porcine IVM/IVF Models for Reproductive Toxicology. Reprod. Biol. Endocrinol. 2014, 12, 117. [Google Scholar] [CrossRef]
- Zigo, M.; Maňásková-Postlerová, P.; Zuidema, D.; Kerns, K.; Jonáková, V.; Tůmová, L.; Bubeníčková, F.; Sutovsky, P. Porcine Model for the Study of Sperm Capacitation, Fertilization and Male Fertility. Cell Tissue Res. 2020, 380, 237–262. [Google Scholar] [CrossRef]
- Ledesma, A.; Zalazar, L.; Fernández-Alegre, E.; Hozbor, F.; Cesari, A.; Martínez-Pastor, F. Seminal Plasma Proteins Modify the Distribution of Sperm Subpopulations in Cryopreserved Semen of Rams with Lesser Fertility. Anim. Reprod. Sci. 2017, 184, 44–50. [Google Scholar] [CrossRef]
- Evenson, D.P. Sperm Chromatin Structure Assay (SCSA®) for Fertility Assessment. Curr. Protoc. 2022, 2, e508. [Google Scholar] [CrossRef] [PubMed]
- R Core Team: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2023. Available online: http://www.r-project.org (accessed on 5 February 2024).
- Hahne, F.; LeMeur, N.; Brinkman, R.R.; Ellis, B.; Haaland, P.; Sarkar, D.; Spidlen, J.; Strain, E.; Gentleman, R. flowCore: A Bioconductor Package for High Throughput Flow Cytometry. BMC Bioinform. 2009, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Finak, G.; Frelinger, J.; Jiang, W.; Newell, E.W.; Ramey, J.; Davis, M.M.; Kalams, S.A.; De Rosa, S.C.; Gottardo, R. OpenCyto: An Open Source Infrastructure for Scalable, Robust, Reproducible, and Automated, End-to-End Flow Cytometry Data Analysis. PLoS Comput. Biol. 2014, 10, e1003806. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Cluster | VCL (µm/s) | VSL (µm/s) | VAP (µm/s) | LIN (%) | STR (%) | WOB (%) | ALH (µm) | BCF (Hz) | DNC (µm2/s) | DNCm (µm) | % |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Slow | 50.1 ± 22.6 | 14.4 ± 8.5 | 27.4 ± 13.2 | 31.1 ± 17.2 | 58.9 ± 23.3 | 57.4 ± 14.8 | 1.4 ± 0.4 | 9 ± 5.9 | 72 ± 51.3 | 4.7 ± 3.3 | 29.9 |
| Fast | 117.3 ± 51.8 | 34.5 ± 14.4 | 67.8 ± 26.2 | 32.7 ± 20.4 | 58.9 ± 27.7 | 58.2 ± 12.4 | 2.6 ± 1.1 | 20 ± 7.4 | 304.8 ± 238.1 | 8.2 ± 7.1 | 51.8 |
| Hyper | 113.5 ± 51.6 | 9.1 ± 5.8 | 51.7 ± 28 | 8.2 ± 5.8 | 18.5 ± 13.7 | 45.5 ± 9.4 | 2.9 ± 1.1 | 13 ± 7.4 | 331.8 ± 255.9 | 33.6 ± 27 | 18.3 |
| Variable | Day 0 | Day 11 | Day | Boar | Day × Boar |
|---|---|---|---|---|---|
| SD-DFI | 15.08 ± 0.20 | 14.91 ± 0.20 | 0.997 | <0.001 | 0.490 |
| %DFI (%) | 1.12 ± 0.07 | 1.41 ± 0.07 | 0.003 | <0.001 | 0.468 |
| %HDS (%) | 4.31 ± 0.14 | 4.75 ± 0.14 | 0.132 | <0.001 | 0.869 |
| Low mBBr (%) | 6.88 ± 0.59 | 34.18 ± 0.60 | <0.001 | <0.001 | <0.001 |
| Moderate mBBr (%) | 88.27 ± 0.66 | 64.55 ± 0.67 | <0.001 | <0.001 | <0.001 |
| High mBBr (%) | 4.84 ± 0.20 | 1.27 ± 0.20 | <0.001 | <0.001 | 0.005 |
| Median mBBr (MFI) | 0.95 ± 0.01 | 1.18 ± 0.01 | <0.001 | <0.001 | <0.001 |
| Disulfide levels | 33.28 ± 0.31 | 21.31 ± 0.31 | <0.001 | <0.001 | <0.001 |
| Low CMA3 (%) | 6.09 ± 0.20 | 5.66 ± 0.20 | 0.431 | 0.643 | 0.385 |
| Moderate CMA3 (%) | 87.46 ± 0.33 | 86.82 ± 0.33 | 0.040 | 0.674 | 0.355 |
| High CMA3 (%) | 6.45 ± 0.24 | 7.51 ± 0.24 | <0.001 | 0.301 | 0.412 |
| Median CMA3 (MFI) | 1.083 ± 0.004 | 1.099 ± 0.004 | 0.002 | 0.495 | 0.893 |
| 8-oxo-dG MFI (%) | 2.54 ± 0.05 | 2.80 ± 0.05 | <0.001 | <0.001 | <0.001 |
| Variable | Loadings | Contributions | ||
|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | |
| SD-DFI | 0.189 | 0.686 | 0.87 | 23.38 |
| %DFI | 0.341 | 0.527 | 2.82 | 13.82 |
| %HDS | 0.185 | 0.417 | 0.83 | 8.64 |
| Low mBBr fluor. | 0.910 | 0.029 | 20.10 | 0.04 |
| Moderate mBBr fluor. | −0.884 | −0.075 | 18.97 | 0.28 |
| High mBBr fluor. | −0.562 | 0.202 | 7.65 | 2.03 |
| mBBr MFI | −0.859 | −0.015 | 17.90 | 0.01 |
| Disulfide levels | −0.861 | 0.016 | 17.99 | 0.01 |
| Low CMA3 fluor. | −0.139 | 0.169 | 0.47 | 1.42 |
| Moderate CMA3 fluor. | −0.407 | 0.513 | 4.03 | 13.07 |
| High CMA3 fluor. | 0.411 | −0.511 | 4.10 | 12.96 |
| CMA3 MFI | 0.216 | −0.434 | 1.13 | 9.37 |
| 8-oxo-dG MFI | 0.359 | 0.549 | 3.13 | 14.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacalle, E.; Fernández-Alegre, E.; Gómez-Giménez, B.; Álvarez-Rodríguez, M.; Martín-Fernández, B.; Soriano-Úbeda, C.; Martínez-Pastor, F. Application of Flow Cytometry Using Advanced Chromatin Analyses for Assessing Changes in Sperm Structure and DNA Integrity in a Porcine Model. Int. J. Mol. Sci. 2024, 25, 1953. https://doi.org/10.3390/ijms25041953
Lacalle E, Fernández-Alegre E, Gómez-Giménez B, Álvarez-Rodríguez M, Martín-Fernández B, Soriano-Úbeda C, Martínez-Pastor F. Application of Flow Cytometry Using Advanced Chromatin Analyses for Assessing Changes in Sperm Structure and DNA Integrity in a Porcine Model. International Journal of Molecular Sciences. 2024; 25(4):1953. https://doi.org/10.3390/ijms25041953
Chicago/Turabian StyleLacalle, Estíbaliz, Estela Fernández-Alegre, Belén Gómez-Giménez, Manuel Álvarez-Rodríguez, Beatriz Martín-Fernández, Cristina Soriano-Úbeda, and Felipe Martínez-Pastor. 2024. "Application of Flow Cytometry Using Advanced Chromatin Analyses for Assessing Changes in Sperm Structure and DNA Integrity in a Porcine Model" International Journal of Molecular Sciences 25, no. 4: 1953. https://doi.org/10.3390/ijms25041953
APA StyleLacalle, E., Fernández-Alegre, E., Gómez-Giménez, B., Álvarez-Rodríguez, M., Martín-Fernández, B., Soriano-Úbeda, C., & Martínez-Pastor, F. (2024). Application of Flow Cytometry Using Advanced Chromatin Analyses for Assessing Changes in Sperm Structure and DNA Integrity in a Porcine Model. International Journal of Molecular Sciences, 25(4), 1953. https://doi.org/10.3390/ijms25041953