
1,6-Hexanediol Is Inducing Homologous Recombination by Releasing BLM from Assemblysomes in Drosophila melanogaster

Abstract
1. Introduction
2. Results
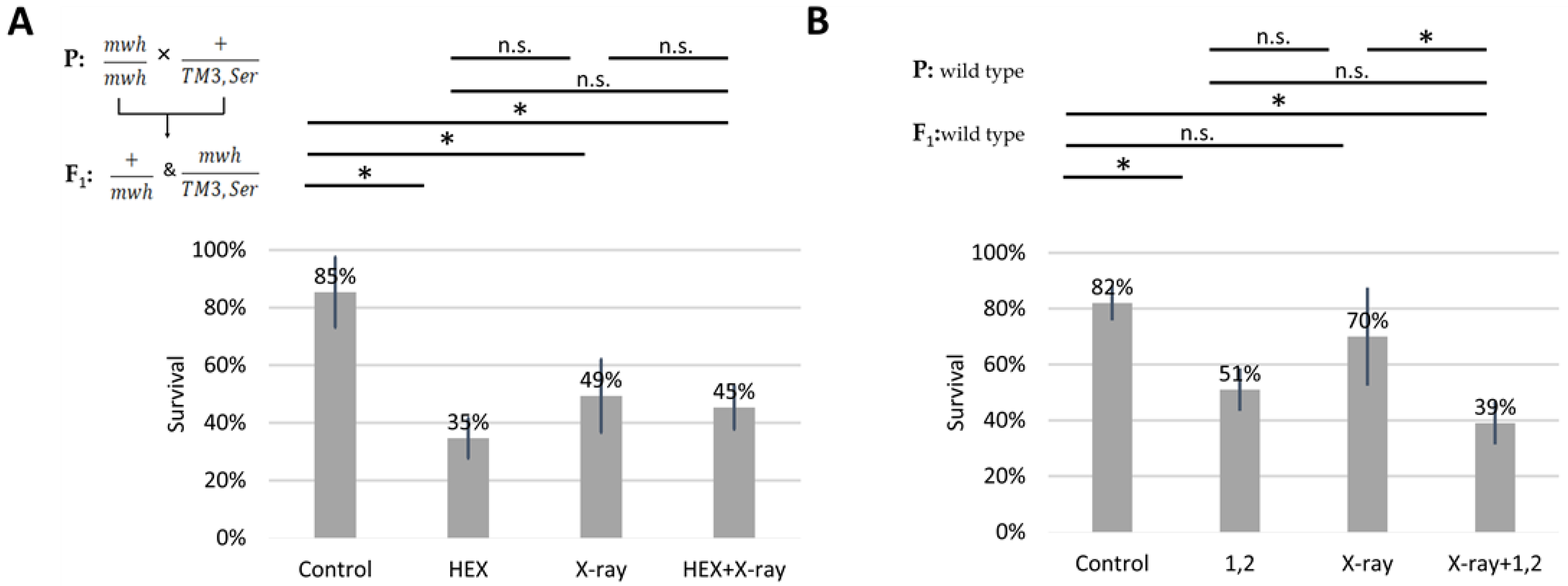
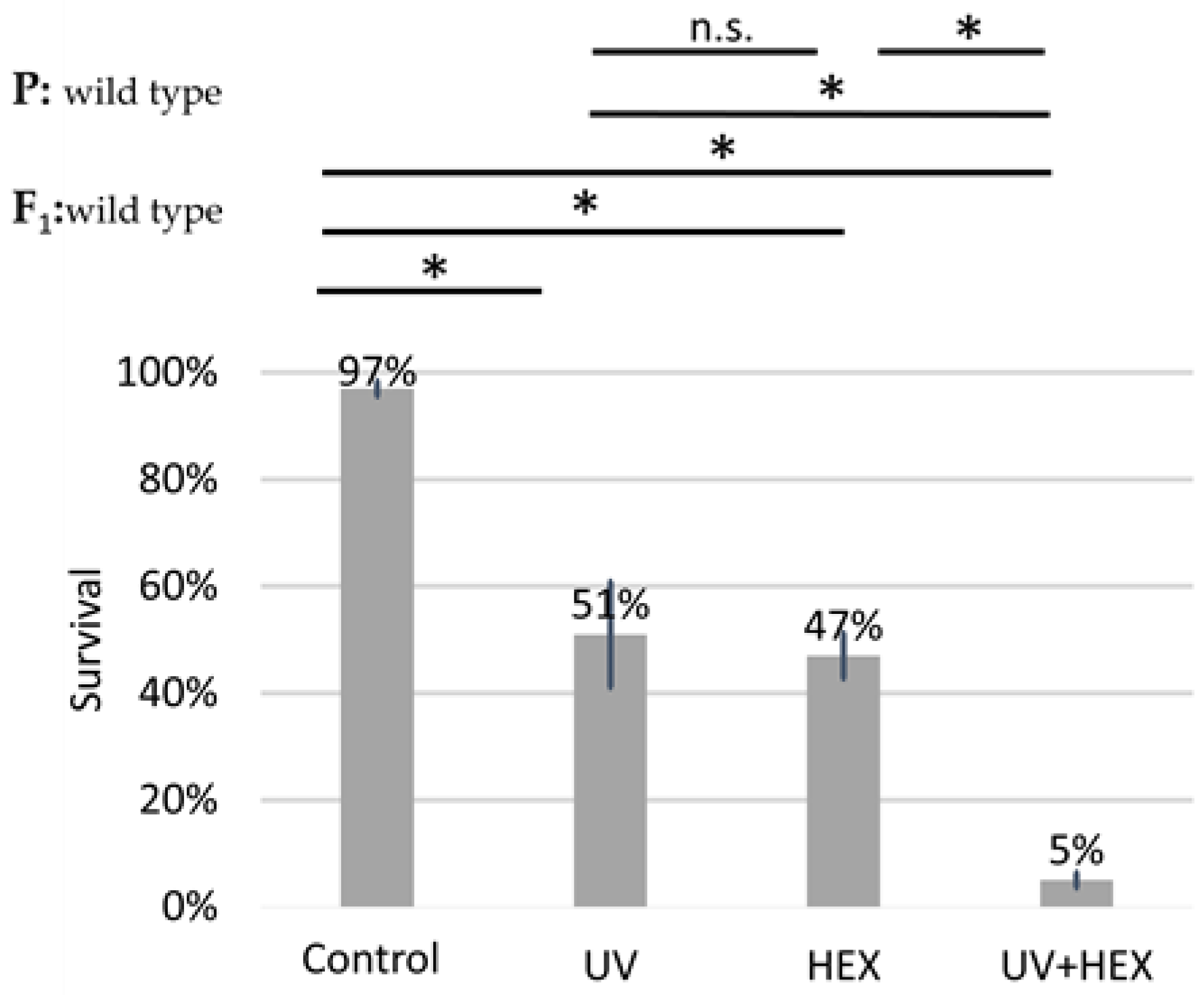
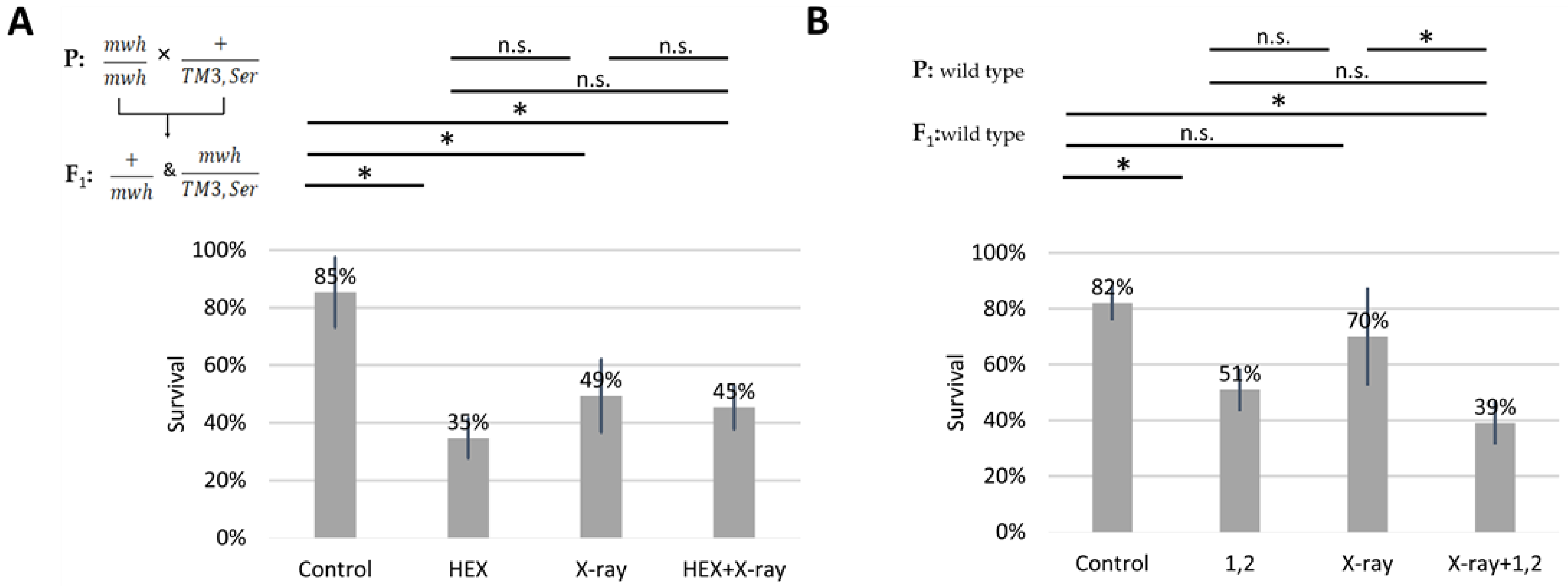
2.1. 1,6-Hexanediol Is Less Toxic If Larvae Are Treated with X-ray
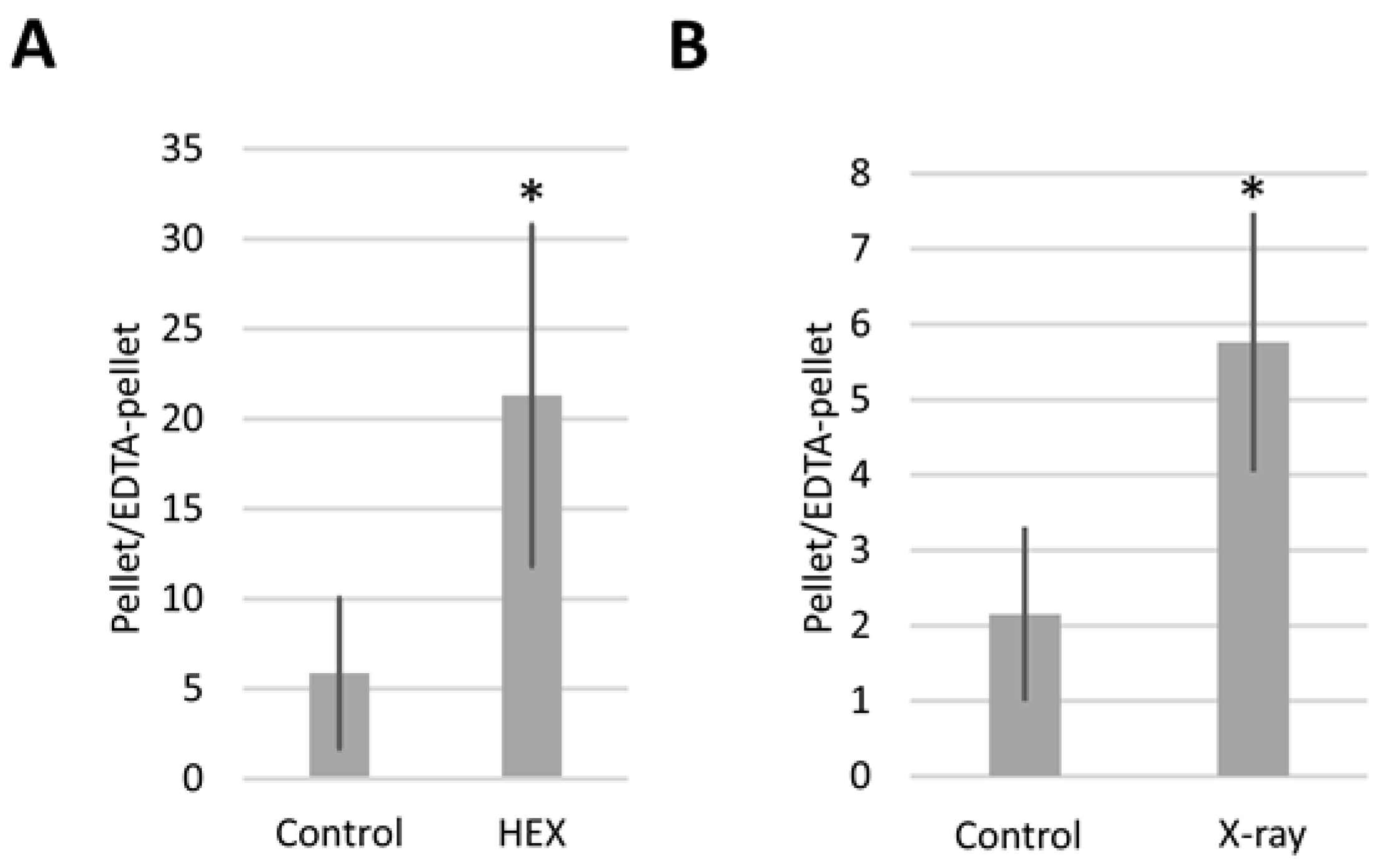
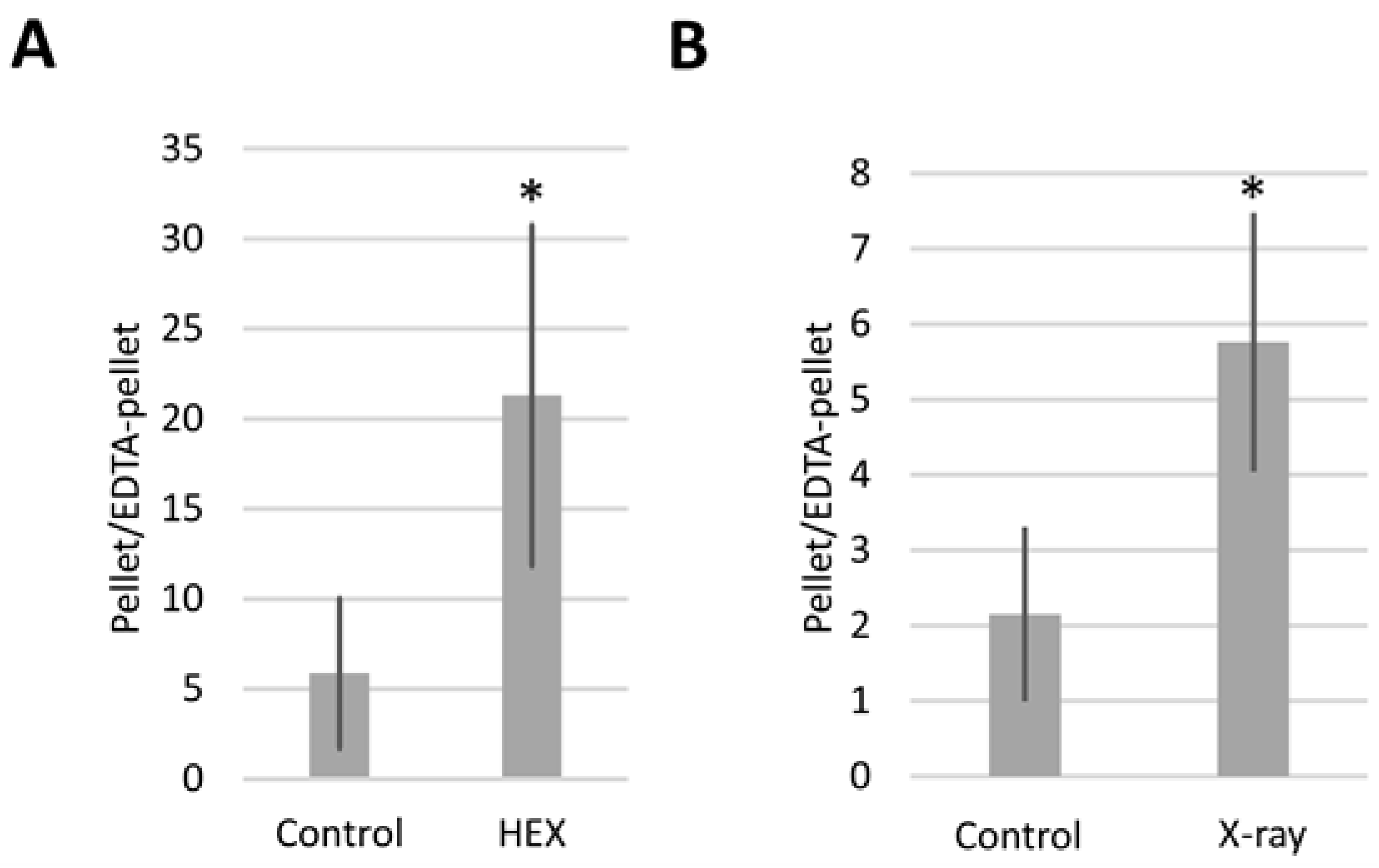
2.2. Bloom Syndrome Helicase RNA Containing Assemblysomes Switches to Translating Ribosomes in Response to X-ray or HEX Treatment
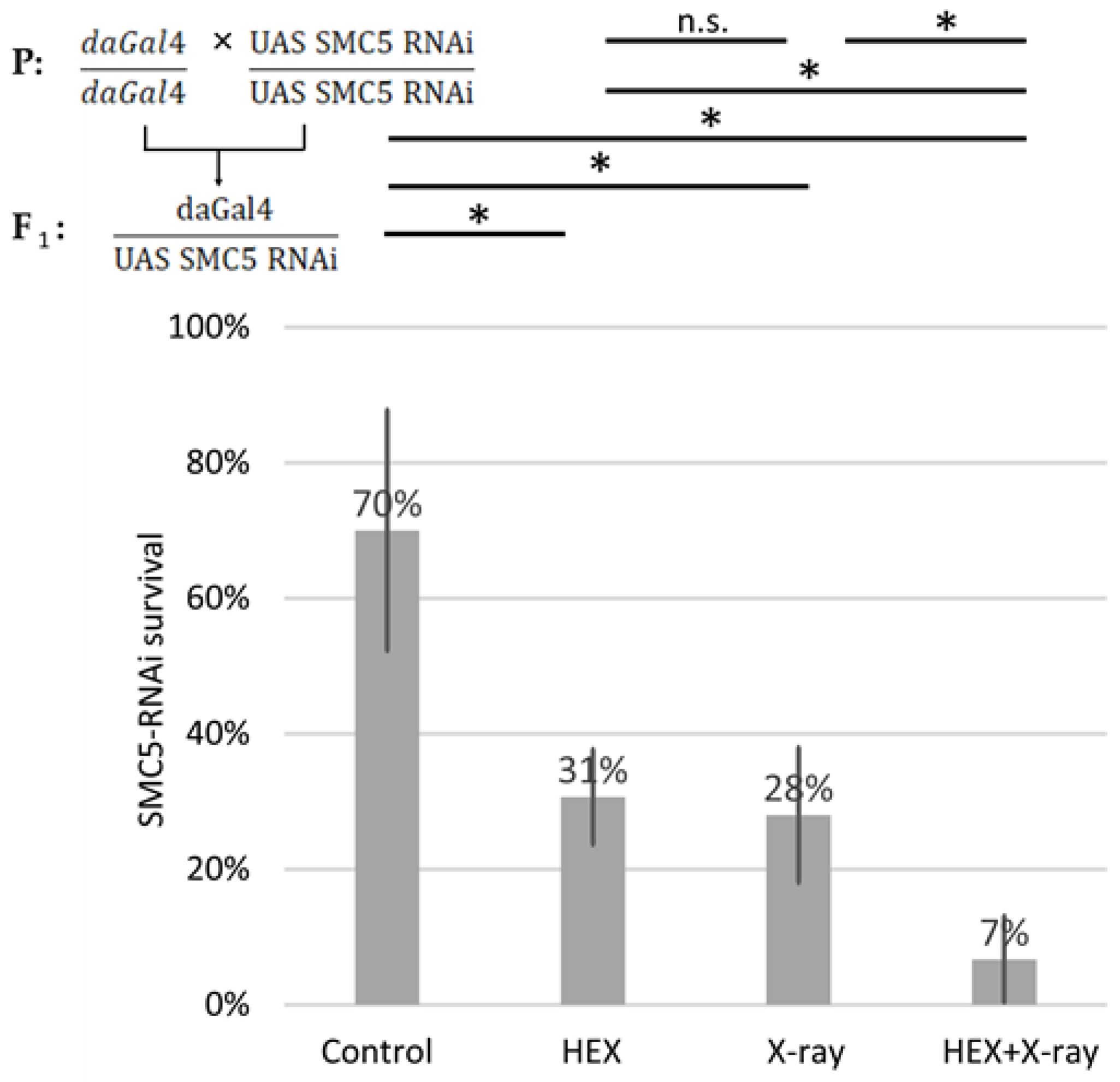
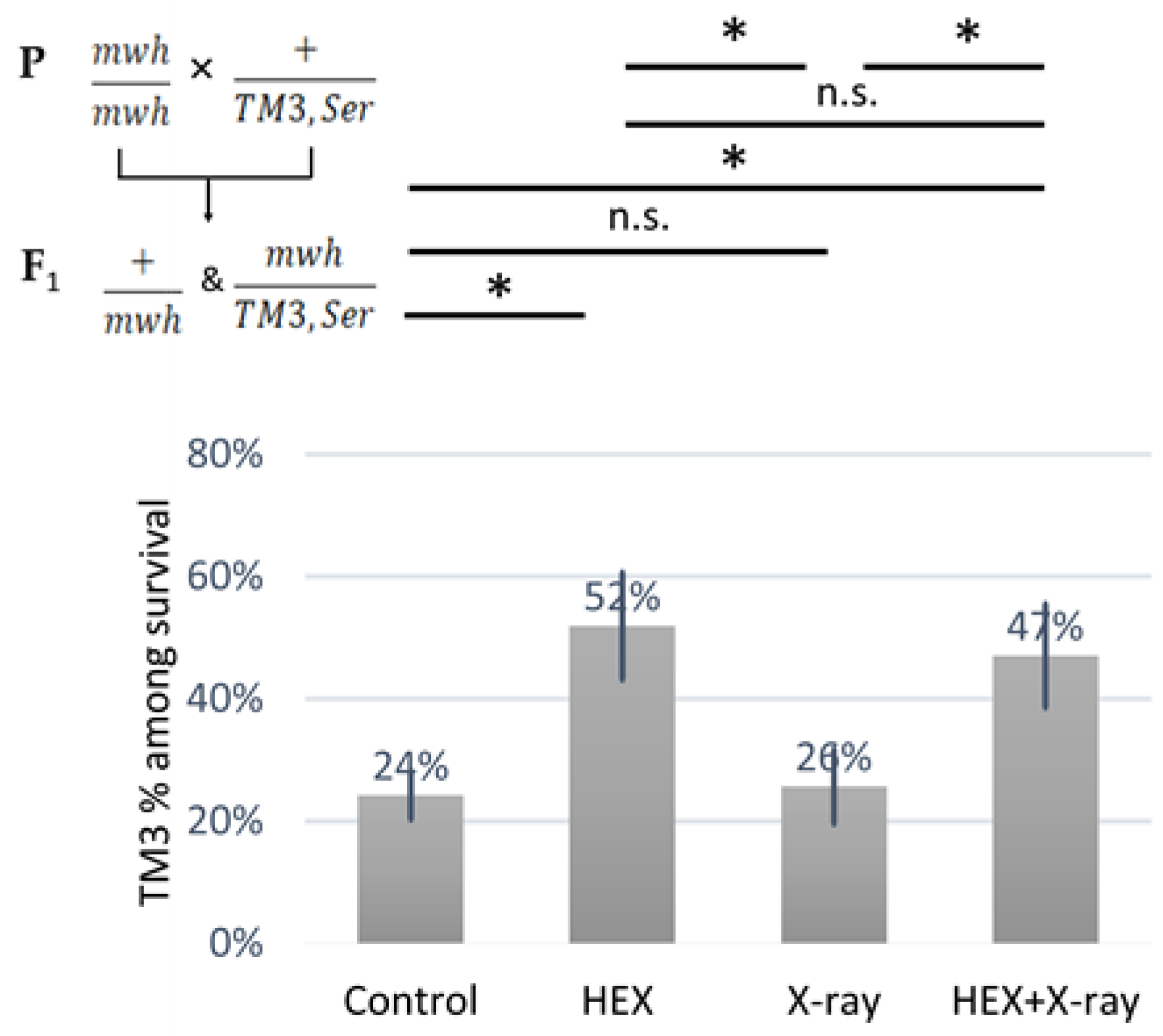
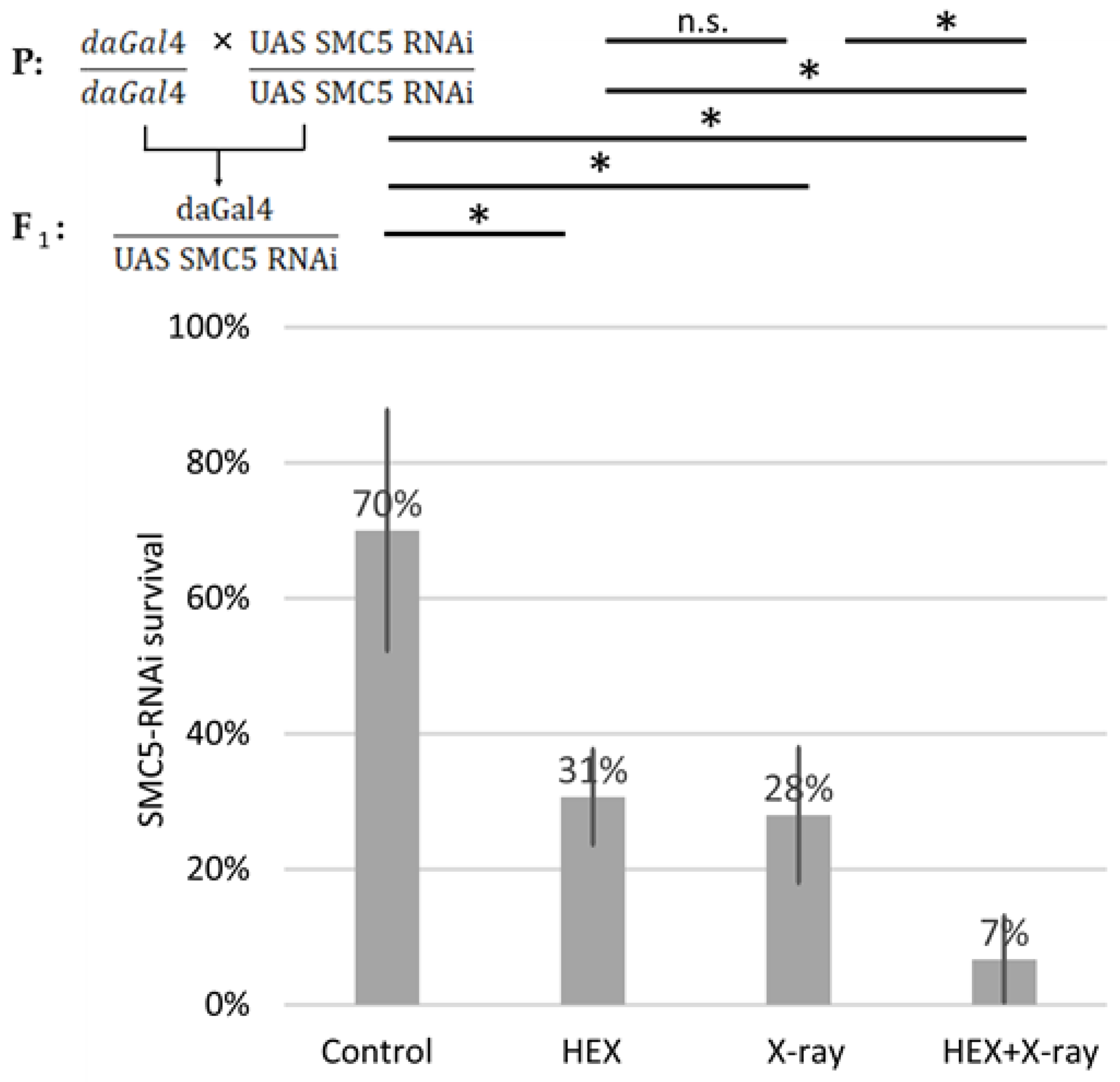
2.3. The SMC5/6 Complex Is Involved in the Recruitment of Assemblysome-Regulated DNA Repair Machinery to the Site of DNA Damage
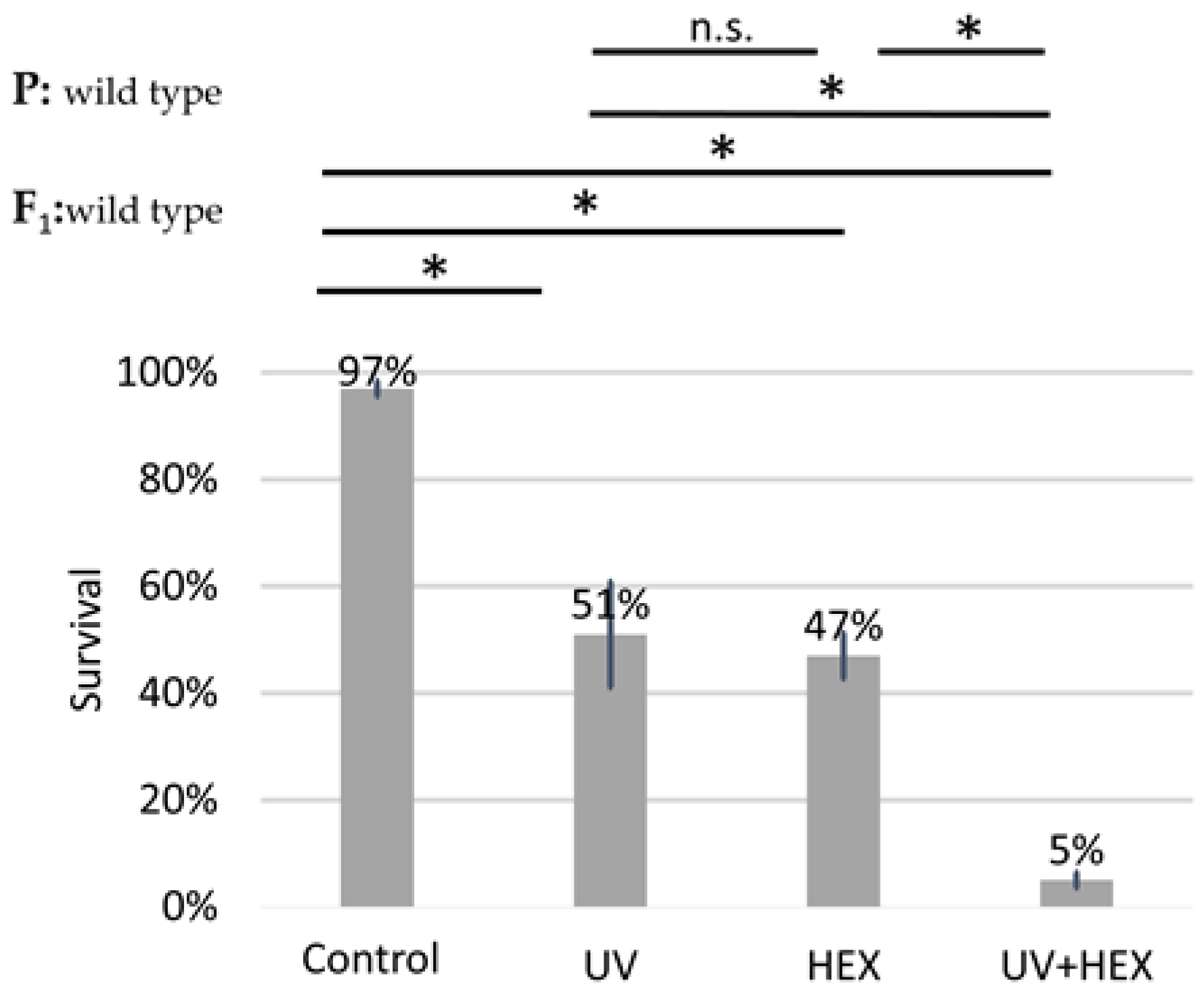
2.4. UV-Induced DNA Damage Is Additive to HEX’ Effect on Mortality
3. Discussion
4. Materials and Methods
4.1. Genotypes, Markers, Crosses, and Strains
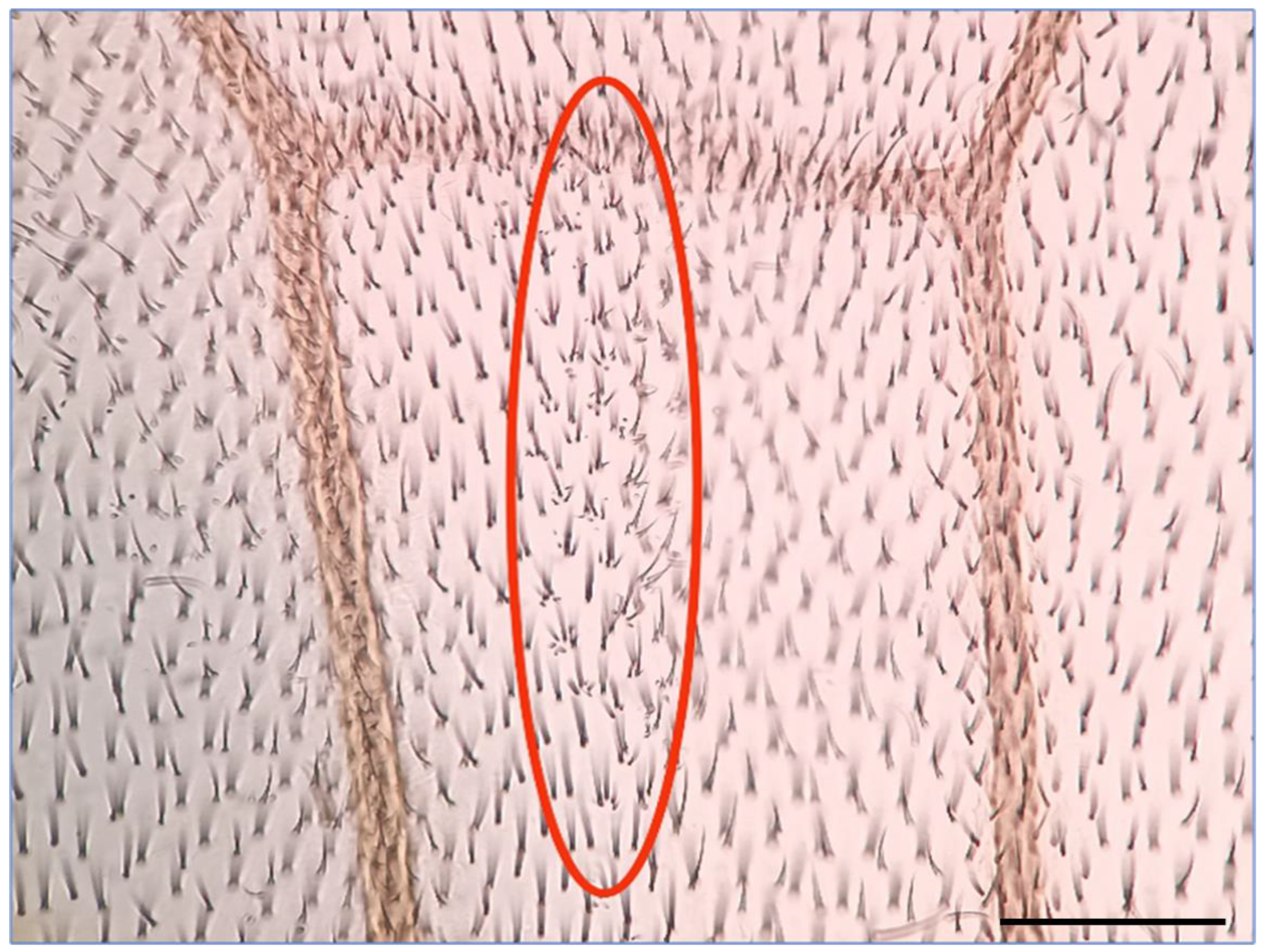
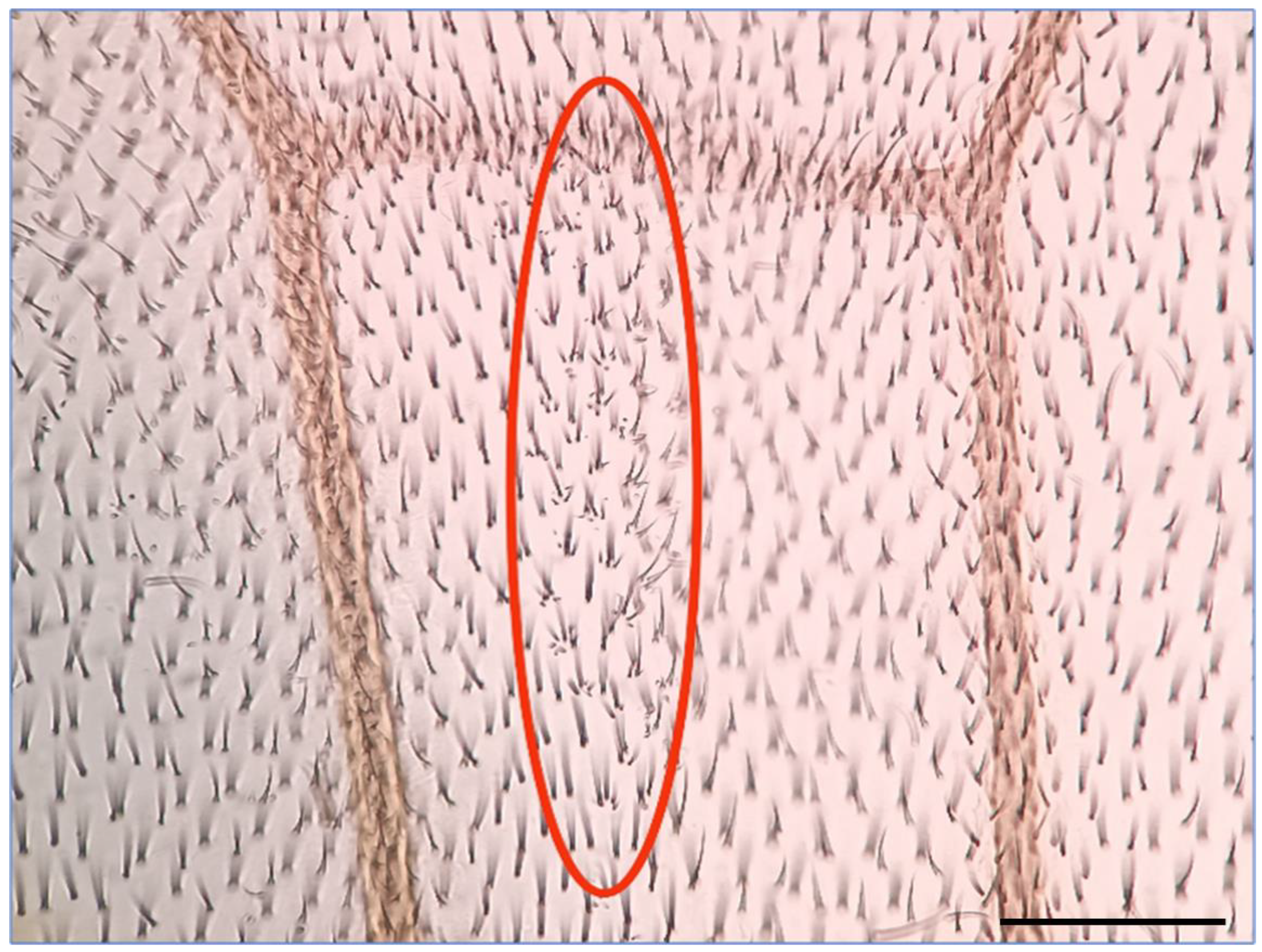
4.2. Somatic Mutation and Recombination Test (SMART)
4.3. Larval Treatments
4.4. Isolation of Assemblysomes
4.5. RNA Extraction—Quantitative Real-Time Polymerase Chain Reaction
4.6. Chemicals
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| Sgs1 | Slow growth suppressor 1 |
| BLM | Bloom syndrome helicase |
| SMC5 | Structural maintenance of chromosomes protein 5 |
| HR | Homologous recombination |
| DmBLM | Drosophila melanogaster bloom syndrome helicase |
| P-bodies | Processing bodies |
| SGs | Stress granules |
| mwh | multiple wing hairs |
| HEX | 1,6-hexanediol |
| RNCs | Ribosome nascent chain complexes |
| qPCR | quantitative polymerase chain reaction |
| EDTA | Ethylenediaminetetraacetic acid |
| UAS | Upstream activation sequence |
| RNAi | Ribonucleic acid interference |
| DSBs | Double-strand breaks |
| SMART | Somatic Mutation and Recombination Test |
| flr | flare |
| Ser | Serrate |
| TM3 | Third Multiple 3 |
| UV | Ultraviolet |
| RT | Reverse transcription |
| LLPS | Liquid–liquid phase separation |
References
- Nemeth-Szatmari, O.; Nagy-Miko, B.; Gyorkei, A.; Varga, D.; Kovacs, B.B.H.; Igaz, N.; Bognar, B.; Razga, Z.; Nagy, G.; Zsindely, N.; et al. Phase separated ribosome-nascent chain complexes in genotoxic stress response. RNA 2023, 29, 1557–1574. [Google Scholar] [CrossRef]
- Panasenko, O.O.; Somasekharan, S.P.; Villanyi, Z.; Zagatti, M.; Bezrukov, F.; Rashpa, R.; Cornut, J.; Iqbal, J.; Longis, M.; Carl, S.H.; et al. Co-translational assembly of proteasome subunits in NOT1-containing assemblysomes. Nat. Struct. Mol. Biol. 2019, 26, 110–120. [Google Scholar] [CrossRef]
- Kusano, K.; Berres, M.A.; Engels, W.R. Evolution of the RECQ Family of Helicases: A Drosophila Homolog, Dmblm, Is Similar to the Human Bloom Syndrome Gene. Genet. Soc. Am. 1999, 151, 1027–1039. [Google Scholar] [CrossRef]
- Birkbak, N.J.; Pathania, Y.L.; Green-Colozzi, A.; Dreze, M.; Bowman-Colin, C.; Sztupinszki, Z.; Krzystanek, M.; Diossy, M.; Tung, N.; Ryan, P.D.; et al. Overexpression of BLM promotes DNA damage and increased sensitivity to platinum salts in triple-negative breast and serous ovarian cancers. Annu. Oncol. 2018, 29, 903–909. [Google Scholar] [CrossRef]
- Ellis, N.A.; Groden, J.; Ye, T.-Z.; Straughen, J.; Lennon, D.J.; Ciocci, S.; Proytcheva, M.; German, J. The Bloom Gene Product Is Homologous to RecQ Helicases. Cell 1995, 83, 655–666. [Google Scholar] [CrossRef]
- Grabarz, A.; Guirouilh-Barbat, J.; Barascu, A.; Pennarun, G.; Genet, D.; Rass, E.; Germann, S.M.; Bertrand, P.; Hickson, I.D.; Lopez, B.S. A role for BLM in double-strand break repair pathway choice: Prevention of CtIP/Mre11-mediated alternative nonhomologous end-joining. Cell Rep. 2013, 5, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.B.; Hickson, I.D. RecQ Helicases: Conserved Guardians of Genomic Integrity. Adv. Exp. Med. Biol. 2013, 767, 161–184. [Google Scholar] [PubMed]
- Ertl, H.A.; Russo, D.P.; Srivastava, N.; Brooks, J.T.; Dao, T.N.; LaRocque, J.R. The Role of Blm Helicase in Homologous Recombination, Gene Conversion Tract Length, and Recombination Between Diverged Sequences in Drosophila melanogaster. Genetics 2017, 207, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Gravel, S.; Chapman, J.R.; Magill, C.; Jackson, S.P. DNA helicases Sgs1 and BLM promote DNA double-strand break resection. Genes Dev. 2008, 22, 2767–2772. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; McVey, M.; Sekelsky, J. Drosophila BLM in double-strand break repair by synthesis-dependent strand annealing. Science 2003, 299, 265–267. [Google Scholar] [CrossRef] [PubMed]
- Paques, F.; Haber, J.E. Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1999, 63, 349–404. [Google Scholar] [CrossRef]
- McVey, M.; Andersen, S.L.; Broze, Y.; Sekelsky, J. Multiple functions of Drosophila BLM helicase in maintenance of genome stability. Genetics 2007, 176, 1979–1992. [Google Scholar] [CrossRef]
- Kroschwald, S.; Maharana, S.; Alberti, S. Hexanediol: A chemical probe to investigate the material properties of membrane-less compartments. Matters 2017, 3, e201702000010. [Google Scholar] [CrossRef]
- Amankwaa, B.; Schoborg, T.; Labrador, M. Drosophila insulator proteins exhibit in vivo liquid-liquid phase separation properties. Life Sci. Alliance 2022, 5, 12. [Google Scholar] [CrossRef]
- Jones, C.E.; Forsburg, S.L. Impact of 1,6-hexanediol on Schizosaccharomyces pombe genome stability. Genes. Genomes Genet. 2023, 13, jkad123. [Google Scholar] [CrossRef]
- Inada, T. Quality control systems for aberrant mRNAs induced by aberrant translation elongation and termination. Biochim. Biophys. Acta-Gene Regul. Mech. 2013, 1829, 634–642. [Google Scholar] [CrossRef]
- Graf, U.; Würgler, F.E.; Katz, A.J.; Frei, H.; Juon, H.; Hall, C.B.; Kale, P.G. Somatic Mutation and Recombination Test in Drosophila melanogaster. Environ. Mutagen. 1984, 6, 153–188. [Google Scholar] [CrossRef]
- Würgler, F.E.; Graf, U.; Frei, H. Somatic mutation and recombination test in wings of Drosophila melanogaster. Prog. Mutat. Res. 1985, 5, 325–340. [Google Scholar]
- Szabad, J.; Soós, I.; Polgár, G.; Héjja, G. Testing the mutagenicity of malondialdehyde and formaldehyde by the Drosophila mosaic and the sex-linked recessive lethal tests. Mutat. Res./Environ. Mutagen. Relat. Subj. 1983, 113, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, D.L.; Zimm, G.G. The Genome of Drosophila Melanogaster; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Garcia-Bellido, A.; Merriam, J.R. Parameters of the Wing Imaginal Disc Development of Drosophila melanogaster. Dev. Biol. 1971, 24, 61–87. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bellido, A.; Dapena, J. Induction, detection and characterization of cell differentiation mutants in Drosophila. Mol. Gen. Genet. 1974, 128, 117–130. [Google Scholar]
- Hoffmann, G.R. Induction of genetic recombination: Consequences and model systems. Environ. Mol. Mutagen. 1994, 24, 59–66. [Google Scholar] [CrossRef]
- Bryant, P.J. Cell lineage relationships in the imaginal wing disc of Drosophila melanogaster. Dev. Biol. 1970, 22, 389–411. [Google Scholar] [CrossRef]
- Haynie, J.L.; Bryant, P.J. The effects of X-rays on the proliferation dynamics of the cells in the wing disc of Drosophila raelanogaster. Wilhelm Roux’ Arch. 1977, 183, 85–100. [Google Scholar] [CrossRef]
- Bradley, M.O.; Kohn, K.W. X-ray-induced DNA double strand break production and repair in mammalian cells as measured by neutral filter elution. Nucleic Acids Res. 1979, 7, 793–804. [Google Scholar] [CrossRef]
- Crown, N.K.; Miller, D.E.; Sekelsky, J.; Hawley, S.R. Local inversion heterozygosity alters recombination throughout the genome. Curr. Biol. 2018, 28, 2984–2990. [Google Scholar] [CrossRef]
- Bermúdez-López, M.; Villoria, M.T.; Esteras, M.; Jarmuz, A.; Torres-Rosell, J.; Clemente-Blanco, A.; Aragon, L. Sgs1′s roles in DNA end resection, HJ dissolution, and crossover suppression require a two-step SUMO regulation on Smc5/6. Genes Dev. 2016, 30, 1339–1356. [Google Scholar] [CrossRef] [PubMed]
- Eladad, S.; Ye, T.-Z.; Hu, P.; Leversha, M.; Beresten, S.; Matunis, M.J.; Ellis, N.A. Intra-nuclear trafficking of the BLM helicase to DNA damage-induced foci is regulated by SUMO modification. Hum. Mol. Genet. 2005, 10, 1351–1365. [Google Scholar] [CrossRef] [PubMed]
- Harvey, S.H.; Sheedy, D.M.; Cuddihy, A.R.; O’Connell, M.J. Coordination of DNA Damage Responses via the Smc5/Smc6 Complex. Mol. Cell. Biol. 2004, 24, 662–674. [Google Scholar] [CrossRef]
- Rossi, F.; Helbling-Leclerc, A.; Jegadesan, N.K.; Xu, X.; Devulder, P.; Abe, T.; Takata, M.; Xu, D.; Roselli, F.; Branzei, D. SMC5/6 acts jointly with Fanconi anemia factors to support DNA repair and genome stability. EMBO Rep. 2020, 21, e48222. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez-López, M.; Aragon, L. Smc5/6 complex regulates Sgs1 recombination functions. Curr. Genet. 2017, 63, 381–388. [Google Scholar] [CrossRef]
- Lehmann, A.R. The role of SMC proteins in the responses to DNA damage. DNA Repair. 2005, 4, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-F.; Yang, S.-A.; Gong, S.; Chang, C.-H.; Portilla, J.M.; Chatterjee, D.; Irianto, J.; Bao, H.; Huang, Y.-C.; Deng, W.-M. Polyploid mitosis and depolyploidization promote chromosomal instability and tumor progression in a Notch-induced tumor model. Dev. Cell 2021, 13, 1976–1988. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.-P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Kier, L.D.; Yamasaki, E.; Ames, B.N. Detection of Mutagenic Activity in Cigarette Smoke Condensates. Proc. Natl. Acad. Sci. USA 1974, 71, 4159–4163. [Google Scholar] [CrossRef]
- Nagy-Mikó, B.; Németh-Szatmári, O.; Faragó-Mészáros, R.; Csókási, A.; Bognár, B.; Ördög, N.; Borsos, B.N.; Majoros, H.; Ujfaludi, Z.; Oláh-Németh, O.; et al. Predictive Potential of RNA Polymerase B (II) Subunit 1 (RPB1) Cytoplasmic Aggregation for Neoadjuvant Chemotherapy Failure. Int. J. Mol. Sci. 2023, 24, 5869. [Google Scholar] [CrossRef]
- Szabad, J. Genetic Mosaicism; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2021. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | HEX | X-ray Treatment | HEX+X-ray Treatment | |
|---|---|---|---|---|
| Number of treated larvae | 150 | 150 | 150 | 150 |
| Number of mosaic spots (mwh) between mutants | 2 | 4 | 25 | 40 |
| Number of mwh/TM3,Ser survivals | 31 | 27 | 19 | 32 |
| Number of mwh/+ survivals | 97 | 25 | 55 | 36 |
| Number of examined wing pairs (all survivals) | 128 | 52 | 74 | 68 |
| Average size of mwh spots in trans-heterozygotes ± standard deviation | 3 ± 0 | 4.5 ± 3.4 | 27.6 ± 43.1 | 16.5 ± 30.9 |
| Frequency of mwh spots (mwh) | 0.0103 | 0.08 | 0.2273 | 0.5556 |
| Recombination frequency | 1.0309 × 10−6 | 1.0667 × 10−5 | 2.0939 × 10−4 | 3.0602 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gombás, B.G.; Villányi, Z. 1,6-Hexanediol Is Inducing Homologous Recombination by Releasing BLM from Assemblysomes in Drosophila melanogaster. Int. J. Mol. Sci. 2024, 25, 1611. https://doi.org/10.3390/ijms25031611
Gombás BG, Villányi Z. 1,6-Hexanediol Is Inducing Homologous Recombination by Releasing BLM from Assemblysomes in Drosophila melanogaster. International Journal of Molecular Sciences. 2024; 25(3):1611. https://doi.org/10.3390/ijms25031611
Chicago/Turabian StyleGombás, Bence György, and Zoltán Villányi. 2024. "1,6-Hexanediol Is Inducing Homologous Recombination by Releasing BLM from Assemblysomes in Drosophila melanogaster" International Journal of Molecular Sciences 25, no. 3: 1611. https://doi.org/10.3390/ijms25031611
APA StyleGombás, B. G., & Villányi, Z. (2024). 1,6-Hexanediol Is Inducing Homologous Recombination by Releasing BLM from Assemblysomes in Drosophila melanogaster. International Journal of Molecular Sciences, 25(3), 1611. https://doi.org/10.3390/ijms25031611