
Pharmacological Mechanisms of Bile Acids Targeting the Farnesoid X Receptor

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
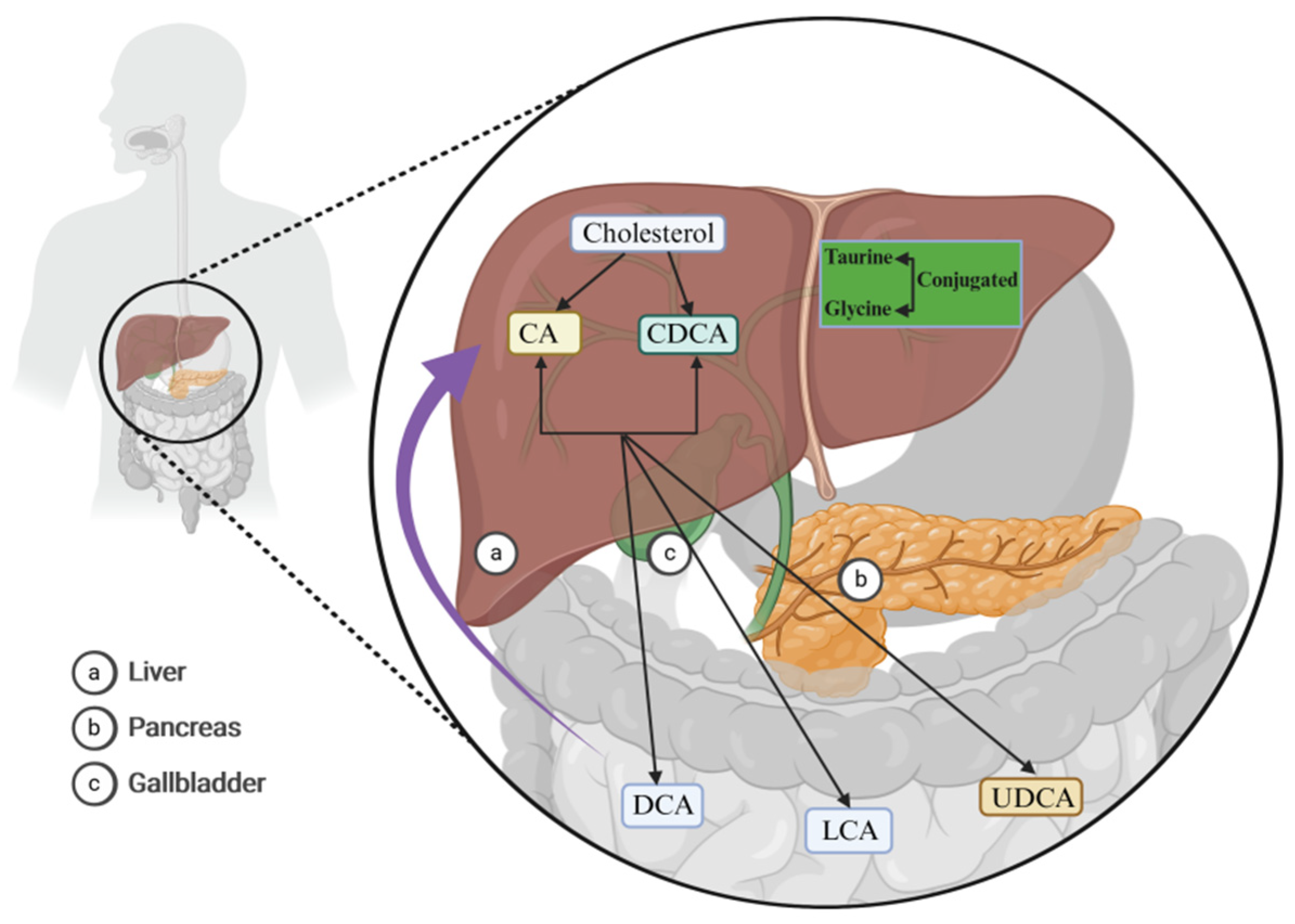
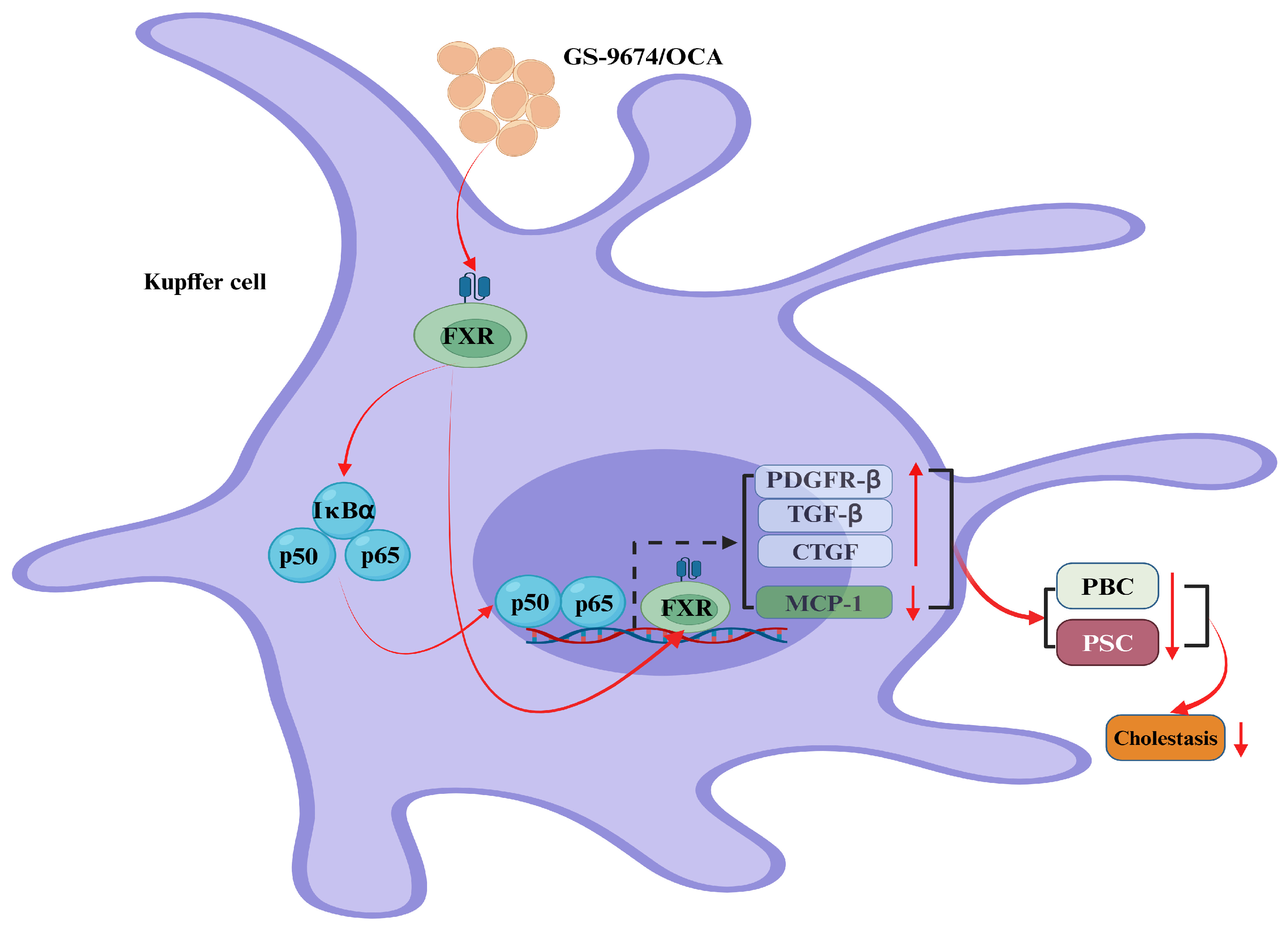
2. BAs Regulate Cholestasis via the FXR
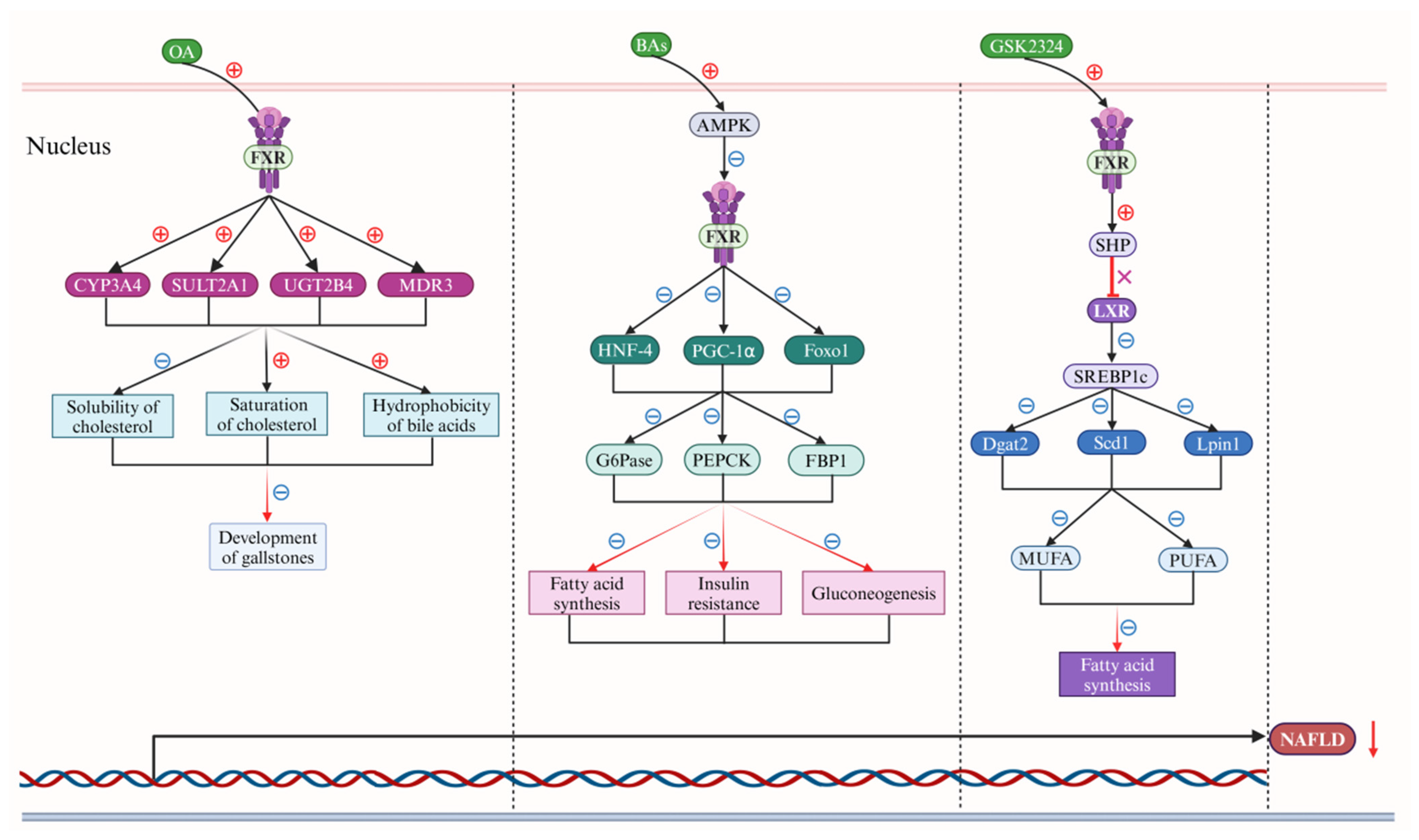
3. BAs Regulate NAFLD via the FXR
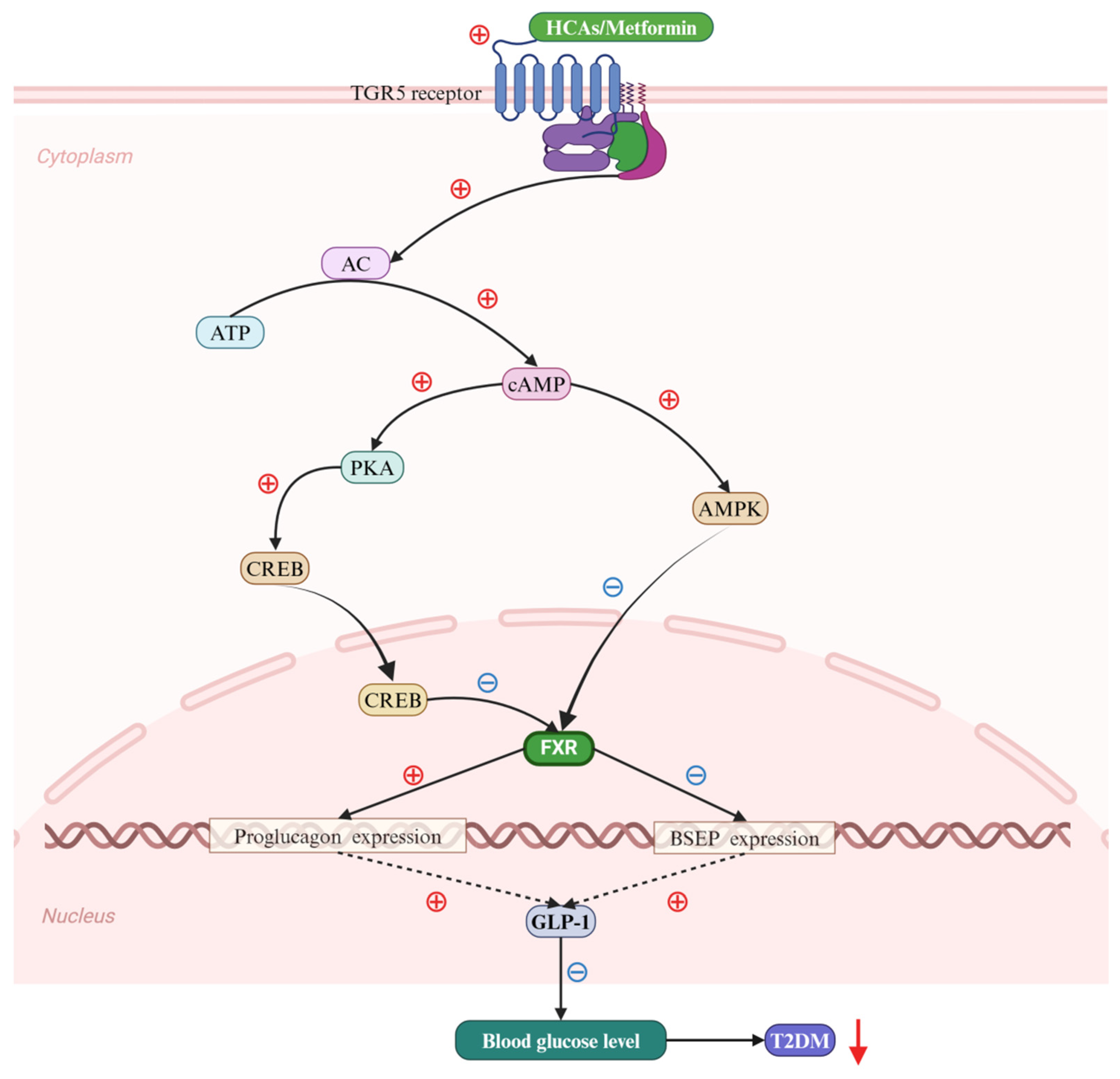
4. BAs Regulate T2DM via the FXR
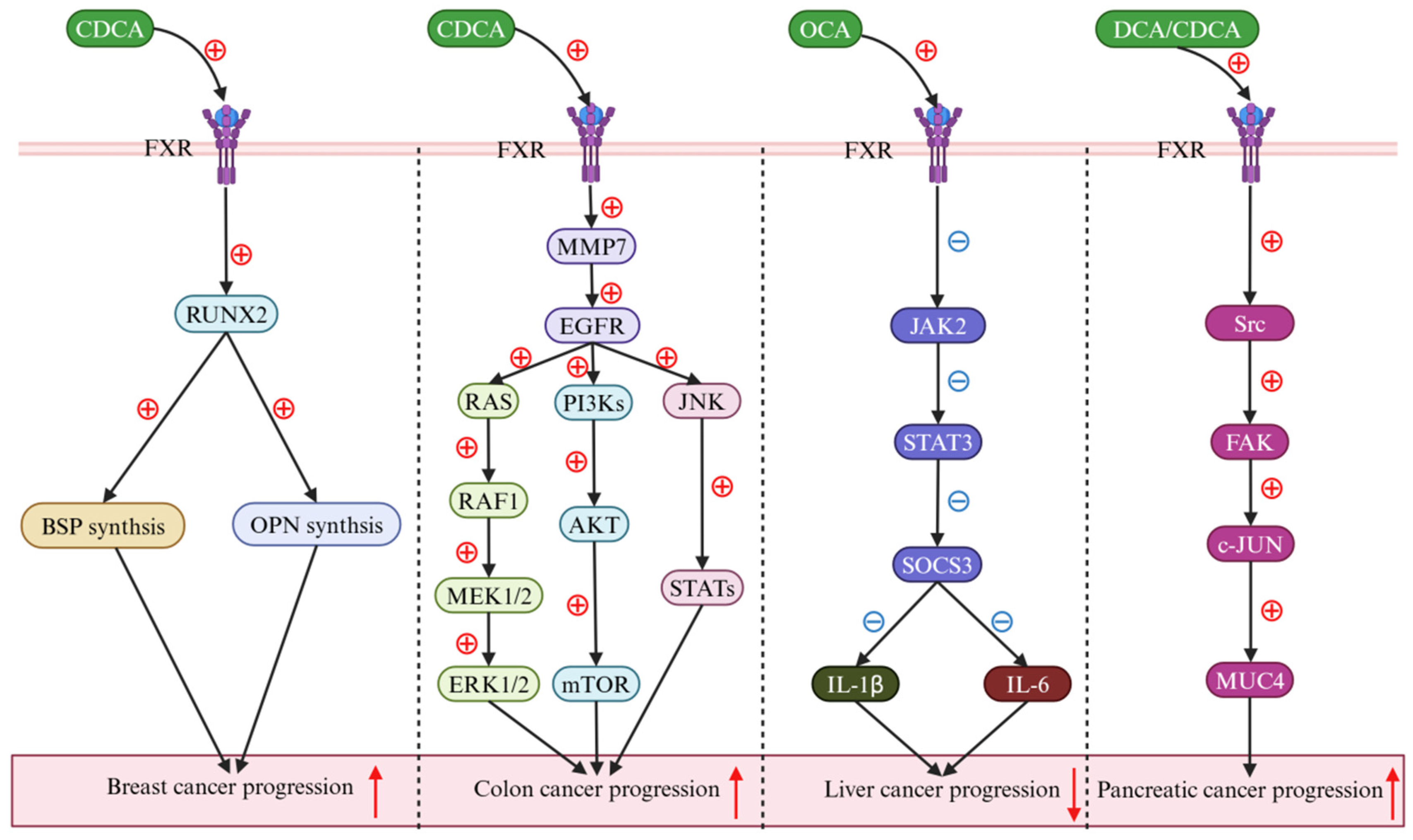
5. BAs Regulate Cancer via the FXR
6. Conclusions and Outlook
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chiang, J.Y.L. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [PubMed]
- Hu, H.; Shao, W.T.; Liu, Q.; Liu, N.; Wang, Q.; Xu, J.; Zhang, X.; Weng, Z.; Lu, Q.; Jiao, L.; et al. Gut microbiota promotes cholesterol gallstone formation by modulating bile acid composition and biliary cholesterol secretion. Nat. Commun. 2022, 13, 252. [Google Scholar] [CrossRef] [PubMed]
- di Gregorio, M.C.; Cautela, J.; Galantini, L. Physiology and physical chemistry of bile acids. Int. J. Mol. Sci. 2021, 22, 1780. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, K.; Wang, X.; Pang, Y.; Jiang, C. The role of the gut microbiome and its metabolites in metabolic diseases. Protein Cell 2021, 12, 360–373. [Google Scholar] [CrossRef]
- Thomas, C.; Pellicciari, R.; Pruzanski, M.; Auwerx, J.; Schoonjans, K. Targeting bile-acid signalling for metabolic diseases. Nat. Rev. Drug Discov. 2008, 7, 678–693. [Google Scholar] [CrossRef]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary bile acids and short chain fatty acids in the colon: A focus on colonic microbiome, cell proliferation, inflammation, and cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef] [PubMed]
- Perino, A.; Schoonjans, K. Metabolic messengers: Bile acids. Nat. Metab. 2022, 4, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, C.D.; Trauner, M. Role of bile acids and their receptors in gastrointestinal and hepatic pathophysiology. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 432–450. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Traxinger, B.R.; Richert-Spuhler, L.E.; Lund, J.M. Mucosal tissue regulatory T cells are integral in balancing immunity and tolerance at portals of antigen entry. Mucosal Immunol. 2022, 15, 398–407. [Google Scholar] [CrossRef]
- Pinho, S.S.; Alves, I.; Gaifem, J.; Rabinovich, G.A. Immune regulatory networks coordinated by glycans and glycan-binding proteins in autoimmunity and infection. Cell Mol. Immunol. 2023, 20, 1101–1113. [Google Scholar] [CrossRef]
- Peng, Y.L.; Wang, S.H.; Zhang, Y.L.; Chen, M.Y.; He, K.; Li, Q.; Huang, W.H.; Zhang, W. Effects of bile acids on the growth, composition and metabolism of gut bacteria. npj Biofilms Microbiomes. 2024, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Kumar, V.; Yang, G.; Kajbaf, K.; Rubio, M.B.; Overturf, K.; Brezas, A.; Hardy, R. Bile acid metabolism in fish: Disturbances caused by fishmeal alternatives and some mitigating effects from dietary bile inclusions. Rev. Aquacult. 2022, 12, 1792–1817. [Google Scholar] [CrossRef]
- Hagey, L.R.; Moller, P.R.; Hofmann, A.F.; Krasowski, M.D. Diversity of bile salts in fish and amphibians: Evolution of a complex biochemical pathway. Physiol Biochem Zool. 2010, 83, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, I.; Allaband, C.; Mannochio-Russo, H.; El Abiead, Y.; Hagey, L.R.; Knight, R.; Dorrestein, P.C. The changing metabolic landscape of bile acids-keys to metabolism and immune regulation. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 493–516. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, T.; Zhao, X.; Gao, Y. New insights into the bile acid-based regulatory mechanisms and therapeutic perspectives in alcohol-related liver disease. Cell Mol Life Sci. 2022, 79, 486. [Google Scholar] [CrossRef] [PubMed]
- Guzior, D.V.; Quinn, R.A. Review: Microbial transformations of human bile acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- Fleishman, J.S.; Kumar, S. Bile acid metabolism and signaling in health and disease: Molecular mechanisms and therapeutic targets. Sig. Transduct. Target. Ther. 2024, 9, 97. [Google Scholar] [CrossRef] [PubMed]
- Jiao, T.; Ma, Y.; Guo, X.; Ye, Y.F.; Xie, C. Bile acid and receptors: Biology and drug discovery for nonalcoholic fatty liver disease. Acta Pharmacol. Sin. 2022, 43, 1103–1119. [Google Scholar] [CrossRef]
- Keitel, V.; Dröge, C.; Häussinge, D. Targeting FXR in cholestasis. Handb. Exp. Pharmacol. 2019, 256, 299–324. [Google Scholar] [PubMed]
- Chen, W.D.; Wang, Y.D.; Meng, Z.; Zhang, L.; Huang, W. Nuclear bile acid receptor FXR in the hepatic regeneration. Biochim. Biophys. Acta 2011, 1812, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Song, H.K.; Cha, J.J.; Han, J.Y.; Kang, Y.S.; Cha, D.R. Farnesoid X receptor (FXR) agonist ameliorates systemic insulin resistance, dysregulation of lipid metabolism, and alterations of various organs in a type 2 diabetic kidney animal model. Acta Diabetol. 2021, 58, 495–503. [Google Scholar] [CrossRef]
- Armstrong, L.E.; Guo, G.L. Role of FXR in liver inflammation during nonalcoholic steatohepatitis. Curr. Pharmacol. Rep. 2017, 3, 92–100. [Google Scholar] [CrossRef]
- Li, K.; Zou, J.A.; Li, S.; Guo, J.; Shi, W.; Wang, B.; Han, X.; Zhang, H.; Zhang, P.; Miao, Z.; et al. Farnesoid X receptor contributes to body weight-independent improvements in glycemic control after Roux-en-Y gastric bypass surgery in diet-induced obese mice. Mol. Metab. 2020, 37, 100980. [Google Scholar] [CrossRef]
- Tian, S.; Chen, S.; Pan, C.; Li, Y. FXR: Structures, biology, and drug development for NASH and fibrosis diseases. Acta Pharmacol. Sin. 2022, 43, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, H.; Xiao, D.; Wei, H.; Chen, Y. Farnesoid X receptor (FXR): Structures and ligands. Comput. Struct. Biotechnol. J. 2021, 19, 2148–2159. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Huang, W.K.; Young, R.L.; Jones, K.L.; Horowitz, M.; Rayner, C.K.; Wu, T. Role of bile acids in the regulation of food intake, and their dysregulation in metabolic disease. Nutrients 2021, 13, 1104. [Google Scholar] [CrossRef] [PubMed]
- Marchioni Beery, R.M.; Vaziri, H.; Forouhar, F. Primary biliary cirrhosis and primary sclerosing cholangitis: A review featuring a women’s health perspective. J. Clin. Transl. Hepatol. 2014, 2, 266–284. [Google Scholar] [PubMed]
- Hou, C.; Ren, C.; Luan, L.; Li, S. A case report of primary biliary cholangitis combined with ankylosing spondylitis. Medicine 2023, 102, e35655. [Google Scholar] [CrossRef] [PubMed]
- Song, J.M.; Li, Y.; Bowlus, C.L.; Yang, G.; Leung, P.S.; Gershwin, M.E. Cholangiocarcinoma in patients with primary sclerosing cholangitis (PSC): A comprehensive review. Clin. Rev. Allerg. Immu. 2020, 58, 134–149. [Google Scholar] [CrossRef]
- Kwo, P.Y.; Cohen, S.M.; Lim, J.K. ACG clinical guideline: Evaluation of abnormal liver chemistries. Am. J. Gastroenterol. 2017, 12, 18–35. [Google Scholar] [CrossRef]
- Trauner, M.; Gulamhusein, A.; Hameed, B.; Caldwell, S.; Shiffman, M.L.; Landis, C.; Eksteen, B.; Agarwal, K.; Muir, A.; Rushbrook, S.; et al. The nonsteroidal farnesoid X receptor agonist cilofexor (GS-9674) improves markers of cholestasis and liver injury in patients with primary sclerosing cholangitis. Hepatology 2019, 70, 788–801. [Google Scholar] [CrossRef]
- Zeng, T.T.; Zhang, C.L.; Xiao, M.; Yang, R.; Xie, K.Q. Critical roles of kupffer cells in the pathogenesis of alcoholic liver disease: From basic science to clinical trials. Front. Immunol. 2016, 7, 538. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Wu, S.B.; Kau, H.C.; Wei, Y.H. Essential role of connective tissue growth factor (CTGF) in transforming growth factor-β1 (TGF-β1)-induced myofibroblast transdifferentiation from Graves’ orbital fibroblasts. Sci. Rep. 2018, 8, 7276. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, H.; Meyer, C.; Li, J.; Nadalin, S.; Königsrainer, A.; Weng, H.; Dooley, S.; Ten Dijke, P. Transforming growth factor-β (TGF-β)-mediated connective tissue growth factor (CTGF) expression in hepatic stellate cells requires Stat3 signaling activation. J. Biol. Chem. 2023, 288, 30708–30719. [Google Scholar] [CrossRef] [PubMed]
- Gillard, J.; Clerbaux, L.A.; Nachit, M.; Sempoux, C.; Staels, B.; Bindels, L.B.; Tailleux, A.; Leclercq, I.A. Bile acids contribute to the development of non-alcoholic steatohepatitis in mice. Jhep Rep. 2021, 4, 100387. [Google Scholar] [CrossRef] [PubMed]
- Xun, Z.; Yao, X.; Ou, Q. Emerging roles of bile acids in chronic hepatitis, liver cirrhosis, and hepatocellular carcinoma. Cell Mol. Immunol. 2023, 20, 1087–1089. [Google Scholar] [CrossRef]
- Sommersberger, S.; Gunawan, S.; Elger, T.; Fererberger, T.; Loibl, J.; Huss, M.; Kandulski, A.; Krautbauer, S.; Müller, M.; Liebisch, G.; et al. Altered fecal bile acid composition in active ulcerative colitis. Lipids Health Dis. 2023, 22, 199. [Google Scholar] [CrossRef]
- Panzitt, K.; Wagner, M. FXR in liver physiology: Multiple faces to regulate liver metabolism. BBA-Mol. Basis Dis. 2021, 1867, 166133. [Google Scholar] [CrossRef]
- Zhang, Y.; Jackson, J.P., 3rd; St Claire, R.L.; Freeman, K.; Brouwer, K.R.; Edwards, J.E. Obeticholic acid, a selective farnesoid X receptor agonist, regulates bile acid homeostasis in sandwich-cultured human hepatocytes. Pharmacol. Res. Perspect. 2017, 5, e00329. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Jun, D.W.; Yoon, E.L.; Oh, J.H.; Roh, Y.J.; Lee, E.J.; Shin, J.H.; Nam, Y.D.; Kim, H.S. Discovery biomarker to optimize obeticholic acid treatment for non-alcoholic fatty liver disease. Biol. Direct. 2023, 18, 50. [Google Scholar] [CrossRef] [PubMed]
- Suriano, F.; Vieira-Silva, S.; Falony, G.; Roumain, M.; Paquot, A.; Pelicaen, R.; Régnier, M.; Delzenne, N.M.; Raes, J.; Muccioli, G.G.; et al. Novel insights into the genetically obese (ob/ob) and diabetic (db/db) mice: Two sides of the same coin. Microbiome 2021, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Nguyen Dinh Cat, A.; Callera, G.E.; Friederich-Persson, M.; Sanchez, A.; Dulak-Lis, M.G.; Tsiropoulou, S.; Montezano, A.C.; He, Y.; Briones, A.M.; Jaisser, F.; et al. Vascular dysfunction in obese diabetic db/db mice involves the interplay between aldosterone/mineralocorticoid receptor and Rho kinase signaling. Sci. Rep. 2018, 8, 2952. [Google Scholar] [CrossRef]
- Li, Y.Y.; Yu, L.F.; Zhang, L.N.; Qiu, B.Y.; Su, M.B.; Wu, F.; Chen, D.K.; Pang, T.; Gu, M.; Zhang, W.; et al. Novel small-molecule AMPK activator orally exerts beneficial effects on diabetic db/db mice. Toxicol. Appl. Pharm. 2013, 36, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.S.; Shin, S.; Li, H.Y.; Park, E.Y.; Lee, S.M.; Choi, C.S.; Lim, Y.; Jung, H.S.; Jun, H.S. Betacellulin ameliorates hyperglycemia in obese diabetic db/db mice. J. Mol. Med. 2015, 93, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Tschuck, J.; Theilacker, L.; Rothenaigner, I.; Weiß, S.A.; Akdogan, B.; Lam, V.T.; Müller, C.; Graf, R.; Brandner, S.; Pütz, C.; et al. Farnesoid X receptor activation by bile acids suppresses lipid peroxidation and ferroptosis. Nat. Commun. 2023, 14, 6908. [Google Scholar] [CrossRef]
- Clifford, B.L.; Sedgeman, L.R.; Williams, K.J.; Morand, P.; Cheng, A.; Jarrett, K.E.; Chan, A.P.; Brearley-Sholto, M.C.; Wahlström, A.; Ashby, J.W.; et al. FXR activation protects against NAFLD via bile-acid-dependent reductions in lipid absorption. Cell Metab. 2021, 33, 1671–1684. [Google Scholar] [CrossRef]
- Hendrix, S.; Kingma, J.; Ottenhoff, R.; Valiloo, M.; Svecla, M.; Zijlstra, L.F.; Sachdev, V.; Kovac, K.; Levels, J.H.; Jongejan, A.; et al. Hepatic SREBP signaling requires SPRING to govern systemic lipid metabolism in mice and humans. Nat. Commun. 2023, 14, 5181. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Fitzinger, J.; Rodriguez-Blanco, G.; Herrmann, M.; Borenich, A.; Stauber, R.; Aigner, E.; Mangge, H. Gender-specific bile acid profiles in non-alcoholic fatty liver disease. Nutrients 2024, 16, 250. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Jena, P.K.; Liu, H.X.; Kalanetra, K.M.; Gonzalez, F.J.; French, S.W.; Krishnan, V.V.; Mills, D.A.; Wan, Y.J. Gender differences in bile acids and microbiota in relationship with gender dissimilarity in steatosis induced by diet and FXR inactivation. Sci. Rep. 2017, 7, 1748. [Google Scholar] [CrossRef] [PubMed]
- Phelps, T.; Snyder, E.; Rodriguez, E.; Child, H.; Harvey, P. The influence of biological sex and sex hormones on bile acid synthesis and cholesterol homeostasis. Biol. Sex Differ. 2019, 10, 52. [Google Scholar] [CrossRef]
- Osei-Ntansah, A.; Oliver, T.; Lofton, T.; Falzarano, C.; Carr, K.; Huang, R.; Wilson, A.; Damaser, E.; Harvey, G.; Rahman, M.A.; et al. Liver androgen receptor knockout improved high-fat diet induced glucose dysregulation in female mice but not male mice. J. Endocr. Soc. 2024, 8, bvae021. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Wu, Z.; Wang, S.; Yu, Z.; Ullah, R.; Liang, X.; Wu, W.; Huang, K.; Ni, Y.; Wang, J.; et al. Gender differences in non-alcoholic fatty liver disease in obese children and adolescents: A large cross-sectional study. Hepatol. Int. 2024, 18, 179–187. [Google Scholar] [CrossRef]
- Bao, R.X.; Chen, B.B.; Wang, A.; Wang, D.; Pan, J.; Chen, Q.; Wu, Y.; Zhu, Z.; Yu, H.; Zhang, Y.; et al. Intestinal FXR deficiency induces dysregulation of xanthine oxidase and accounts for sex difference in hyperuricemia. Free Radic. Biol. Med. 2024, 226, 374–388. [Google Scholar] [CrossRef]
- Lonardo, A.; Nascimbeni, F.; Ballestri, S.; Fairweather, D.; Win, S.; Than, T.A.; Abdelmalek, M.F.; Suzuki, A. Sex differences in nonalcoholic fatty liver disease: State of the art and identification of research gaps. Hepatology 2019, 70, 1457–1469. [Google Scholar] [CrossRef]
- Ke, C.; Narayan, K.M.V.; Chan, J.C.N.; Jha, P.; Shah, B.R. Pathophysiology, phenotypes and management of type 2 diabetes mellitus in Indian and Chinese populations. Nat. Rev. Endocrinol. 2022, 18, 413–432. [Google Scholar] [CrossRef] [PubMed]
- Antar, S.A.; Ashour, N.A.; Sharaky, M.; Khattab, M.; Ashour, N.A.; Zaid, R.T.; Roh, E.J.; Elkamhawy, A.; Al-Karmalawy, A.A. Diabetes mellitus: Classification, mediators, and complications; A gate to identify potential targets for the development of new effective treatments. Biomed. Pharmacother. 2023, 168, 115734. [Google Scholar] [CrossRef] [PubMed]
- Cloete, L. Diabetes mellitus: An overview of the types, symptoms, complications and management. Nurs. Stand. 2022, 37, 61–66. [Google Scholar] [CrossRef]
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of diabetes: An overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36, S67–S74. [Google Scholar] [CrossRef]
- Vulichi, S.R.; Runthala, A.; Begari, N.; Rupak, K.; Chunduri, V.R.; Kapur, S.; Chippada, A.R.; Sistla, D.S. Type-2 diabetes mellitus-associated cancer risk: In pursuit of understanding the possible link. Diabetes Metab. Syndr. 2022, 16, 102591. [Google Scholar] [CrossRef]
- Pearson-Stuttard, J.; Papadimitriou, N.; Markozannes, G.; Cividini, S.; Kakourou, A.; Gill, D.; Rizos, E.C.; Monori, G.; Ward, H.A.; Kyrgiou, M.; et al. Type 2 Diabetes and Cancer: An Umbrella Review of Observational and Mendelian Randomization Studies. Cancer Epidemiol. Biomarkers Prev. 2021, 30, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A review of colorectal cancer in terms of epidemiology, risk factors, development, symptoms and diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef]
- Douaiher, J.; Ravipati, A.; Grams, B.; Chowdhury, S.; Alatise, O.; Are, C. Colorectal cancer-global burden, trends, and geographical variations. J. Surg. Oncol. 2017, 115, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wu, H.; Cao, M.; Li, H.; He, S.; Yang, F.; Yan, X.; Zhang, S.; Teng, Y.; Xia, C.; et al. Colorectal cancer burden, trends and risk factors in China: A review and comparison with the United States. Chin. J. Cancer Res. 2022, 34, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, X.; Yu, T.; Liu, Y.; Jing, Y. Education reform and change driven by digital technology: A bibliometric study from a global perspective. Humanit. Soc. Sci. Commun. 2024, 11, 256. [Google Scholar] [CrossRef]
- Cladis, A.E. A shifting paradigm: An evaluation of the pervasive effects of digital technologies on language expression, creativity, critical thinking, political discourse, and interactive processes of human communications. E-Learn. Digit. Media 2020, 17, 341–364. [Google Scholar] [CrossRef]
- Hou, Y.; Zhai, X.; Wang, X.; Wu, Y.; Wang, H.; Qin, Y.; Han, J.; Meng, Y. Research progress on the relationship between bile acid metabolism and type 2 diabetes mellitus. Diabetol. Metab. Syndr. 2023, 15, 235. [Google Scholar] [CrossRef] [PubMed]
- Sim, A.Y.; Choi, D.H.; Kim, J.Y.; Kim, E.R.; Goh, A.R.; Lee, Y.H.; Lee, J.E. SGLT2 and DPP4 inhibitors improve Alzheimer’s disease-like pathology and cognitive function through distinct mechanisms in a T2D-AD mouse model. Biomed. Pharmacother. 2023, 168, 115755. [Google Scholar] [CrossRef]
- Tahrani, A.; Barnett, A.; Bailey, C. Pharmacology and therapeutic implications of current drugs for type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2016, 12, 566–592. [Google Scholar] [CrossRef]
- Lissi, C.B.; Zanetti, M.O.B. Pharmacotherapeutic options for the management of type 2 diabetes mellitus: Literature review. Res. Soc. Dev. 2023, 12, e15112340605. [Google Scholar] [CrossRef]
- Wong, K.H.; Li, G.Q.; Li, K.M.; Razmovski-Naumovski, V.; Chan, K. Kudzu root: Traditional uses and potential medicinal benefits in diabetes and cardiovascular diseases. J. Ethnopharmacol. 2011, 134, 584–607. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Shao, Q.Q.; Xia, Q.S.; Hu, M.; Zhao, Y.; Wang, D.; Fang, K.; Xu, L.; Zou, X.; Chen, Z.; et al. A bioinformatics and transcriptomics based investigation reveals an inhibitory role of Huanglian-Renshen-Decoction on hepatic glucose production of T2DM mice via PI3K/Akt/FoxO1 signaling pathway. Phytomedicine 2021, 83, 153487. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, L.; Yue, R.; Ding, N.; Yang, H. Large dosage Huangqin (Scutellaria) and Huanglian (Rhizoma Coptidis) for T2DM: A protocol of systematic review and meta-analysis of randomized clinical trials. Medicine 2020, 299, e22032. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.L.; Ferrell, J.M. Discovery of farnesoid X receptor and its role in bile acid metabolism. Mol. Cell Endocrinol. 2022, 548, 111618. [Google Scholar] [CrossRef]
- Zhang, Y. Farnesoid X receptor-Acting through bile acids to treat metabolic disorders. Drugs Future 2010, 35, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Zhang, H.; Zheng, X.; Zhao, A.; Jia, W. Novel microbial modifications of bile acids and their functional implications. Imeta. 2024, 3, e243. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Dong, N. Effects ofbile acid metabolism on intestinal health of livestock andpoultry. J. Anim. Physiol. Anim. Nutr. 2024, 108, 1258–1269. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.J.; Chen, T.L.; Jiang, R.Q.; Zhao, A.; Wu, Q.; Kuang, J.; Sun, D.; Ren, Z.; Li, M.; Zhao, M.; et al. Hyocholic acid species improve glucose homeostasis through a distinct TGR5 and FXR signaling mechanism. Cell Metab. 2021, 33, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; He, X.; Gao, X.; Liu, Q.; Zhao, Y.; Hong, Y.; Zhu, W.; Yan, J.; Li, Y.; Li, Y.; et al. Hyodeoxycholic acid ameliorates nonalcoholic fatty liver disease by inhibiting RAN-mediated PPARα nucleus-cytoplasm shuttling. Nat. Commun. 2023, 14, 5451. [Google Scholar] [CrossRef] [PubMed]
- Minoretti, P.; Emanuele, E. Bile Acids in Cardiovascular Diseases: Don’t Forget Hyocholic Acid. Aging Dis. 2024, 15, 5451. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Xi, B.; Yang, L.; Sun, J.; Zhao, M.; Bovet, P. Trends in the prevalence of overweight, obesity, and abdominal obesity among Chinese adults between 1993 and 2015. Int. J. Obes. 2021, 45, 427–437. [Google Scholar] [CrossRef]
- Taylor, R.E.; Bhattacharya, A.; Guo, G.L. Environmental Chemical Contribution to the Modulation of Bile Acid Homeostasis and Farnesoid X Receptor Signaling. Drug Metab Dispos. 2022, 50, 456–467. [Google Scholar] [CrossRef]
- Simbrunner, B.; Hofer, B.S.; Schwabl, P.; Zinober, K.; Petrenko, O.; Fuchs, C.; Semmler, G.; Marculescu, R.; Mandorfer, M.; Datz, C.; et al. FXR-FGF19 signaling in the gut–liver axis is dysregulated in patients with cirrhosis and correlates with impaired intestinal defence. Hepatol. Int. 2024, 18, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Yang, Q.; Zhang, F.L.; Chen, L.; Su, H.; Yang, X.; He, H.; Liu, F.; Zheng, J.; Ling, M.; et al. Hyodeoxycholic acid (HDCA) suppresses intestinal epithelial cell proliferation through FXR-PI3K/AKT pathway, accompanied by alteration of bile acids metabolism profiles induced by gut bacteria. FASEB J. 2020, 34, 7103–7117. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Bray, F.; Parkin, D.M. Cancer in sub-Saharan Africa in 2020: A review of current estimates of the national burden, data gaps, and future needs. Lancet Oncol. 2022, 23, 719–728. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ott, J.J.; Ullrich, A.; Mascarenhas, M. Global cancer incidence and mortality caused by behavior and infection. J. Public Health 2011, 33, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Girisa, S.; Henamayee, S.; Parama, D.; Rana, V.; Dutta, U.; Kunnumakkara, A.B. Targeting Farnesoid X receptor (FXR) for developing novel therapeutics against cancer. Mol. Biomed. 2021, 2, 21. [Google Scholar] [CrossRef]
- Absil, L.; Journé, F.; Larsimont, D.; Body, J.J.; Tafforeau, L.; Nonclercq, D. Farnesoid X receptor as marker of osteotropism of breast cancers through its role in the osteomimetism of tumor cells. BMC Cancer 2020, 20, 640. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, J.; Drachenberg, C.B.; Raufman, J.P.; Xie, G. Farnesoid X receptor represses matrix metalloproteinase 7 expression, revealing this regulatory axis as a promising therapeutic target in colon cancer. J. Biol. Chem. 2019, 294, 8529–8542. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.M.; Tawfiq, R.A.; Ali, A.A.; Elmazar, M.M. The FXR agonist, obeticholic acid, suppresses HCC proliferation & metastasis: Role of IL-6/STAT3 signalling wathway. Sci. Rep. 2017, 7, 12502. [Google Scholar]
- Joshi, S.; Cruz, E.; Rachagani, S.; Guha, S.; Brand, R.E.; Ponnusamy, M.P.; Kumar, S.; Batra, S.K. Bile acids-mediated overexpression of MUC4 via FAK-dependent c-Jun activation in pancreatic cancer. Mol. Oncol. 2016, 10, 1063–1077. [Google Scholar] [CrossRef]
- Luo, Z.; Zhou, W.; Xie, T.; Xu, W.; Shi, C.; Xiao, Z.; Si, Y.; Ma, Y.; Ren, Q.; Di, L.; et al. The role of botanical triterpenoids and steroids in bile acid metabolism, transport, and signaling: Pharmacological and toxicological implications. Acta Pharm. Sin. B 2024, 14, 3385–3415. [Google Scholar] [CrossRef]
- Kay, H.Y.; Kim, W.D.; Hwang, S.J.; Choi, H.S.; Gilroy, R.K.; Wan, Y.J.; Kim, S.G. Nrf2 inhibits LXRα-dependent hepatic lipogenesis by competing with FXR for acetylase binding. Antioxid. Redox Signal. 2011, 15, 2135–2146. [Google Scholar] [CrossRef]
- Maliha, S.; Guo, G.L. Farnesoid X receptor and fibroblast growth factor 15/19 as pharmacological targets. Liver Res. 2021, 5, 142–150. [Google Scholar] [CrossRef]
- Yu, J.; Li, S.; Guo, J.; Xu, Z.; Zheng, J.; Sun, X. Farnesoid X receptor antagonizes Wnt/β-catenin signaling in colorectal tumorigenesis. Cell Death Dis. 2020, 11, 640. [Google Scholar] [CrossRef]
- Lin, Q.; Zhang, B.; Dai, M.; Cheng, Y.; Li, F. Aspirin caused intestinal damage through FXR and ET-1 signaling pathways. Int. J. Mol. Sci. 2024, 25, 3424. [Google Scholar] [CrossRef]
- Henry, Z.; Meadows, V.; Guo, G.L. FXR and NASH: An avenue for tissue-specific regulation. Hepatol. Commun. 2023, 7, e0127. [Google Scholar] [CrossRef] [PubMed]
- Habib, S. Team players in the pathogenesis of metabolic dysfunctions-associated steatotic liver disease: The basis of development of pharmacotherapy. World J. Gastrointest Pathophysiol. 2024, 15, 93606. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Shang, B.; Wang, J.; Sun, J.; Li, J.; Liang, B.; Wang, X.; Su, L.; You, W.; Jiang, S. Farnesoid X receptor promotes non-small cell lung cancer metastasis by activating Jak2/STAT3 signaling via transactivation of IL-6ST and IL-6 genes. Cell Death Dis. 2024, 15, 148. [Google Scholar] [CrossRef]
- Kuhls, S.; Osswald, A.; Ocvirk, S. Bile acids, bile pigments and colorectal cancer risk. Curr. Opin. Gastroen. 2022, 38, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.X.; Gao, Y.; Xu, X.F.; Jin, X.; Zhang, Y.; Xu, Q.; Ding, H.X.; Li, B.J.; Du, F.K.; Li, L.C.; et al. Bile acids inhibit ferroptosis sensitivity through activating farnesoid X receptor in gastric cancer cells. World J. Gastroen. 2024, 30, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, Q.; Sun, Q.; Luo, Q.; Wang, Y.; Wang, C.; Zhu, A.; Zhao, L.; Yin, L.; Lou, J.; et al. Investigating the mechanisms of Jieduquyuziyin prescription improves lupus nephritis and fibrosis via FXR in MRL/lpr Mice. Oxid. Med. Cell Longev. 2022, 9, 4301033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Khakisahneh, S.; Han, S.Y.; Song, E.J.; Nam, Y.D.; Kim, H. Ginseng extracts improve circadian clock gene expression and reduce inflammation directly and indirectly through gut microbiota and PI3K signaling pathway. npj Biofilms Microbi. 2024, 10, 24. [Google Scholar] [CrossRef]
- Premkumar, V.; Dey, M.; Dorn, R.; Raskin, I. MyD88-dependent and independent pathways of Toll-Like Receptors are engaged in biological activity of Triptolide in ligand-stimulated macrophages. BMC Chem. Biol. 2010, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Issinger, O.G.; Guerra, B. Phytochemicals in cancer and their effect on the PI3K/AKT-mediated cellular signalling. Biomed. Pharmacother. 2021, 139, 111650. [Google Scholar] [CrossRef]
- Tang, B.B.; Yuan, Y.F.; Wang, S. The Chinese herbal formula QiangGuYin regulates the Wnt/β-catenin pathway in osteoblasts by intervening in the expression of miRNAs derived from osteoclast exosomes. Pharmacol. Res. 2022, 3, 100087. [Google Scholar] [CrossRef]
- Zhu, M.; Sun, Y.; Bai, H.; Wang, Y.; Yang, B.; Wang, Q.; Kuang, H. Effects of saponins from Chinese herbal medicines on signal transduction pathways in cancer: A review. Front. Pharmacol. 2023, 14, 1159985. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.C.; Duan, G.Z.; Fan, G.H. Effect of Lycium barbarum polysaccharides on cell signal transduction pathways. Biomed. Pharmacother. 2022, 147, 112620. [Google Scholar] [CrossRef]
- Tang, X.W.; Liao, Q.; Li, Q.Q.; Jiang, L.; Li, W.; Xu, J.; Xiong, A.; Wang, R.; Zhao, J.; Wang, Z.; et al. Lusianthridin ameliorates high fat diet-induced metabolic dysfunction-associated fatty liver disease via activation of FXR signaling pathway. Eur. J. Pharmacol. 2024, 965, 176196. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Lu, X.; Liu, D.; Zhang, Y.; Zhai, X.; Zhou, L.; Gao, J. Fucogalactan sulfate (FS) from Laminaria japonica regulates lipid metabolism in diet-induced humanized dyslipidemia mice via an intestinal FXR-FGF19-CYP7A1/CYP8B1 pathway. J. Agric. Food Chem. 2023, 71, 14027–14037. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, T.; Moschetta, A. Farnesoid X receptor (FXR) regulation of bile acid homeostasis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2023, 1868 Pt 10, 110856. [Google Scholar]
- Panzitt, K.; Zollner, G.; Marschall, H.U.; Wagner, M. Recent advances on FXR-targeting therapeutics. Mol. Cell Endocrinol. 2022, 552, 111678. [Google Scholar] [CrossRef] [PubMed]
- Kosters, A.; Karpen, S.J. Bile acid signaling in liver disease and therapy. J. Hepatol. 2021, 74, 789–802. [Google Scholar]
- Pathak, P.; Liu, H.; Boehme, S.; Xie, C.; Krausz, K.W.; Gonzalez, F.; Chiang, J.Y.L. Farnesoid X receptor induces Takeda G-protein receptor 5 cross-talk to regulate bile acid synthesis and hepatic metabolism. J. Biol. Chem. 2017, 292, 11055–11069. [Google Scholar] [CrossRef]
- Wang, X.X.; Xie, C.; Libby, A.E.; Ranjit, S.; Levi, J.; Myakala, K.; Bhasin, K.; Jones, B.A.; Orlicky, D.J.; Takahashi, S. The role of FXR and TGR5 in reversing and preventing progression of Western diet–induced hepatic steatosis, inflammation, and fibrosis in mice. J. Biol. Chem. 2022, 298, 102530. [Google Scholar] [CrossRef] [PubMed]
- Vavassori, P.; Mencarelli, A.; Fiorucci, S. The bile acid-FXR axis: A promising target for the treatment of liver and metabolic diseases. Drug Discov. Today. 2022, 27, 1074–1084. [Google Scholar]
- Fiorucci, S.; Rizzo, G.; Donini, A.; Distrutti, E.; Santucci, L. Targeting farnesoid X receptor for liver and metabolic disorders. Trends Mol. Med. 2007, 13, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Kosters, A.; Karpen, S.J. Farnesoid X receptor in liver diseases: Molecular mechanisms and clinical applications. J. Gastroenterol. 2020, 55, 585–598. [Google Scholar]
- Fiorucci, S.; Baldoni, M.; Ricci, P.; Zampella, A.; Distrutti, E.; Biagioli, M. Bile acid-activated receptors and the regulation of macrophages function in metabolic disorders. Curr. Opin. Pharmacol. 2020, 53, 45–54. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Ma, Y.; Duan, G. Pharmacological Mechanisms of Bile Acids Targeting the Farnesoid X Receptor. Int. J. Mol. Sci. 2024, 25, 13656. https://doi.org/10.3390/ijms252413656
Qi Y, Ma Y, Duan G. Pharmacological Mechanisms of Bile Acids Targeting the Farnesoid X Receptor. International Journal of Molecular Sciences. 2024; 25(24):13656. https://doi.org/10.3390/ijms252413656
Chicago/Turabian StyleQi, Youchao, Yonggui Ma, and Guozhen Duan. 2024. "Pharmacological Mechanisms of Bile Acids Targeting the Farnesoid X Receptor" International Journal of Molecular Sciences 25, no. 24: 13656. https://doi.org/10.3390/ijms252413656
APA StyleQi, Y., Ma, Y., & Duan, G. (2024). Pharmacological Mechanisms of Bile Acids Targeting the Farnesoid X Receptor. International Journal of Molecular Sciences, 25(24), 13656. https://doi.org/10.3390/ijms252413656