
Association of Inflammasome Gene Expression Levels with Pathogenesis of Familial Mediterranean Fever in Armenians

 and
and
Abstract
1. Introduction
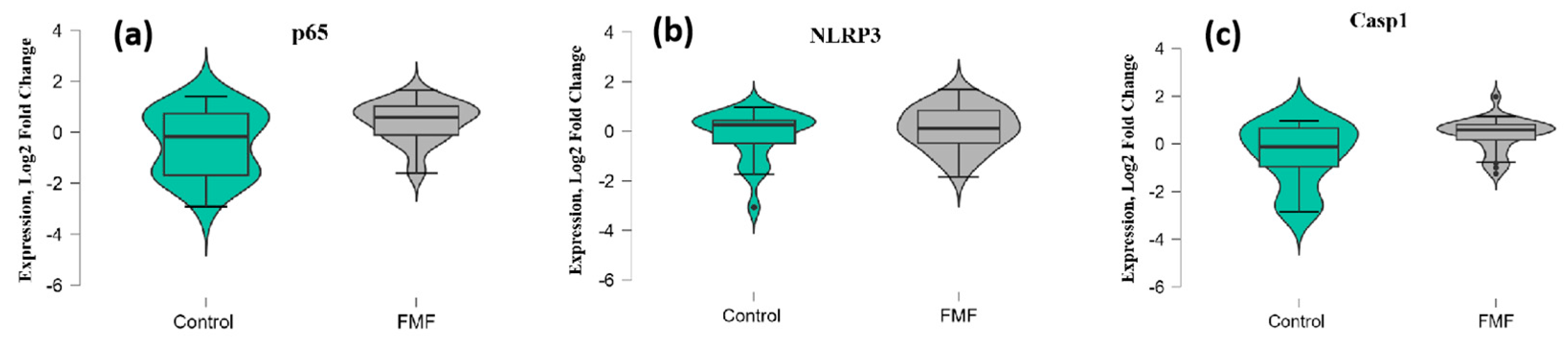
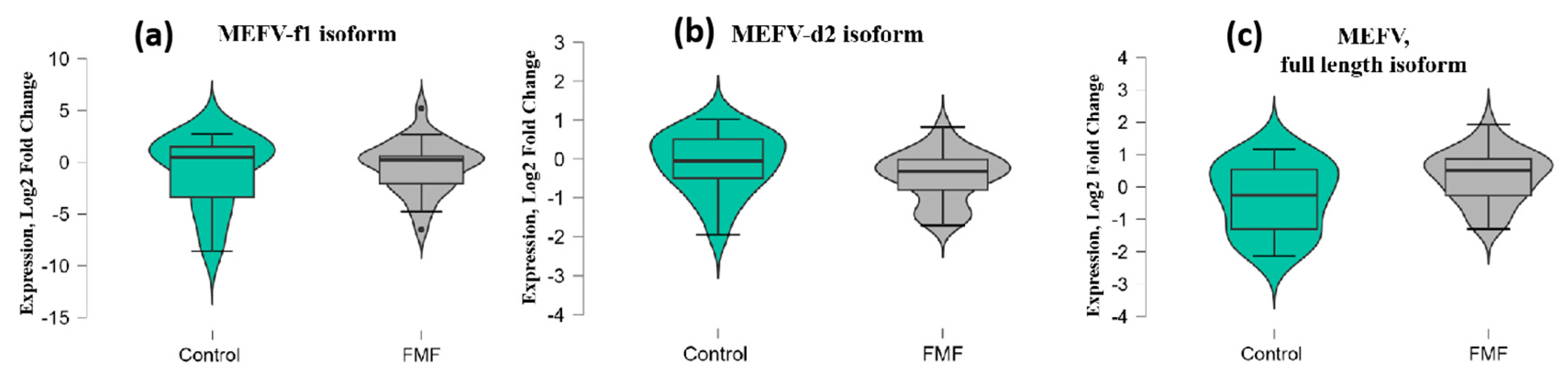
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population and Selection Criteria
4.2. RNA Extraction and cDNA Synthesis
4.3. Evaluation of Inflammasome Gene Expression
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Touitou, I. The spectrum of Familial Mediterranean Fever (FMF) mutations. Eur. J. Hum. Genet. 2001, 9, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Schnappauf, O.; Chae, J.J.; Kastner, D.L.; Aksentijevich, I. The pyrin inflammasome in health and disease. Front. Immunol. 2019, 10, 1745. [Google Scholar] [CrossRef] [PubMed]
- Milhavet, F.; Cuisset, L.; Hoffman, H.M.; Slim, R.; El-Shanti, H.; Aksentijevich, I.; Lesage, S.; Waterham, H.; Wise, C.; Menthiere, C.S.; et al. The infevers autoinflammatory mutation online registry: Update with new genes and functions. Hum. Mutat. 2008, 29, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Grandemange, S.; Soler, S.; Touitou, I. Expression of the familial Mediterranean fever gene is regulated by nonsense-mediated decay. Hum. Mol. Genet. 2009, 18, 4746–4755. [Google Scholar] [CrossRef] [PubMed]
- Papin, S.; Duquesnoy, P.; Cazeneuve, C.; Pantel, J.; Coppey-Moisan, M.; Dargemont, C.; Amselem, S. Alternative splicing at the MEFV locus involved in familial Mediterranean fever regulates translocation of the marenostrin/pyrin protein to the nucleus. Hum. Mol. Genet. 2000, 9, 3001–3009. [Google Scholar] [CrossRef] [PubMed]
- Erdem, G.C.; Erdemir, S.; Abaci, I.; Aydin, A.K.K.; Everest, E.; Turanli, E.T. Alternatively spliced MEFV transcript lacking exon 2 and its protein isoform pyrin-2d implies an epigenetic regulation of the gene in inflammatory cell culture models. Genet. Mol. Biol. 2017, 40, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Matzner, Y.; Abedat, S.; Shapiro, E.; Eisenberg, S.; Bar-Gil-Shitrit, A.; Stepensky, P.; Calco, S.; Azar, Y.; Urieli-Shoval, S. Expression of the familial Mediterranean fever gene and activity of the C5a inhibitor in human primary fibroblast cultures. Blood 2000, 96, 727–731. [Google Scholar] [CrossRef] [PubMed]
- de Torre-Minguela, C.; Del Castillo, P.M.; Pelegrín, P. The NLRP3 and pyrin inflammasomes: Implications in the pathophysiology of autoinflammatory diseases. Front. Immunol. 2017, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.; Ablasser, A.; Bartok, E.; Kim, S.; Schmid-Burgk, J.; Cavlar, T.; Hornung, V. Inflammasomes: Current understanding and open questions. Cell Mol. Life Sci. 2011, 68, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Manukyan, G.; Aminov, R. Update on pyrin functions and mechanisms of Familial Mediterranean fever. Front. Microbiol. 2016, 7, 456. [Google Scholar] [CrossRef] [PubMed]
- Heilig, R.; Broz, P. Function and mechanism of the pyrin inflammasome. Eur. J. Immunol. 2018, 48, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Ozkurede, V.U.; Franchi, L. Immunology in clinic review series; focus on autoinflammatory diseases: Role of inflammasomes in autoinflammatory syndromes. Clin. Exp. Immunol. 2012, 167, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Eigenbrod, T.; Munoz-Planillo, R.; Nunez, G. The inflammasome: A caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nat. Immunol. 2009, 10, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Repa, A.; Bertsias, G.K.; Petraki, E.; Choulaki, C.; Vassou, D.; Kambas, K.; Sidiropoulos, P. Dysregulated production of interleukin-1β upon activation of the NLRP3 inflammasome in patients with familial Mediterranean fever. Hum. Immunol. 2015, 76, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.J.; Wood, G.; Masters, S.L.; Richard, K.; Park, G.; Smith, B.J.; Kastner, D.L. The B30. 2 domain of pyrin, the familial Mediterranean fever protein, interacts directly with caspase-1 to modulate IL-1β production. Proc. Natl. Acad. Sci. USA 2006, 103, 9982–9987. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.J.; Wood, G.; Richard, K.; Jaffe, H.; Colburn, N.T.; Masters, S.L.; Kastner, D.L. The familial Mediterranean fever protein, pyrin, is cleaved by caspase-1 and activates NF-κB through its N-terminal fragment. Blood 2008, 112, 1794–1803. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, H.; Livneh, A.; Lidar, M.; Feld, O.; Yahia, S.H.; Grossman, C.; Ben-Zvi, I. Toll-like receptor 2 is overexpressed in Familial Mediterranean fever patients and is inhibited by colchicine treatment. Best Pract. Res. Clin. Rheumatol. 2018, 32, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Jamilloux, Y.; Lefeuvre, L.; Magnotti, F.; Martin, A.; Benezech, S.; Allatif, O.; Henry, T. Familial Mediterranean fever mutations are hypermorphic mutations that specifically decrease the activation threshold of the pyrin inflammasome. Rheumatology 2018, 57, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| 1 | GAPDH forward | AGCCTCAAGATCATCAGCAAT |
| GAPDH reverse | GTCATGAGTCCTTCCACGATAC | |
| 2 | P65 forward | CTGTCCTTTCTCATCCCATCTT |
| P65 reverse | ACACCTCAATGTCCTCTTTCTG | |
| 4 | Casp1 forward | TTTCCGCAAGGTTCGATTT |
| Casp1 reverse | CCTGGGAAGAGGTAGAAACA | |
| 5 | MEFV i1 forward | ATTCGGTCACCGGAAGG |
| MEFV i1 reverse | GGAATCACGCACACAGGTA | |
| 6 | MEFV i2 forward | ACTCTGCTGGTCACCTACTAT |
| MEFV i2 reverse | GGTGGCCTTCCCTGAATG | |
| 7 | MEFV i2 forward | GGCAGCCATTCAGGGAAG |
| MEFV i2 reverse | GGAATCACGCACACAGGTA | |
| 8 | MEFV i1.2 (full-length) forward | AGATCAGGAAGGCATATGACAC |
| MEFV i1.2 (full-length) reverse | CTCCAATGTCCTGCAGAAGT | |
| 9 | NLRP3 forward | GAAGAGGAGTGGATGGGTTTAC |
| NLRP3 reverse | CTTTCTGTACTTCTTACGGTAATCTTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayrapetyan, V.; Karapetyan, L.; Ghukasyan, L.; Atshemyan, S.; Ghazaryan, H.; Vardanyan, V.; Mukuchyan, V.; Arakelyan, A.; Zakharyan, R. Association of Inflammasome Gene Expression Levels with Pathogenesis of Familial Mediterranean Fever in Armenians. Int. J. Mol. Sci. 2024, 25, 12958. https://doi.org/10.3390/ijms252312958
Hayrapetyan V, Karapetyan L, Ghukasyan L, Atshemyan S, Ghazaryan H, Vardanyan V, Mukuchyan V, Arakelyan A, Zakharyan R. Association of Inflammasome Gene Expression Levels with Pathogenesis of Familial Mediterranean Fever in Armenians. International Journal of Molecular Sciences. 2024; 25(23):12958. https://doi.org/10.3390/ijms252312958
Chicago/Turabian StyleHayrapetyan, Varduhi, Lana Karapetyan, Lilit Ghukasyan, Sofi Atshemyan, Hovsep Ghazaryan, Valentina Vardanyan, Vahan Mukuchyan, Arsen Arakelyan, and Roksana Zakharyan. 2024. "Association of Inflammasome Gene Expression Levels with Pathogenesis of Familial Mediterranean Fever in Armenians" International Journal of Molecular Sciences 25, no. 23: 12958. https://doi.org/10.3390/ijms252312958
APA StyleHayrapetyan, V., Karapetyan, L., Ghukasyan, L., Atshemyan, S., Ghazaryan, H., Vardanyan, V., Mukuchyan, V., Arakelyan, A., & Zakharyan, R. (2024). Association of Inflammasome Gene Expression Levels with Pathogenesis of Familial Mediterranean Fever in Armenians. International Journal of Molecular Sciences, 25(23), 12958. https://doi.org/10.3390/ijms252312958