
Exploring the Replication and Pathogenic Characteristics of Alpha, Delta, and Omicron Variants of SARS-CoV-2

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
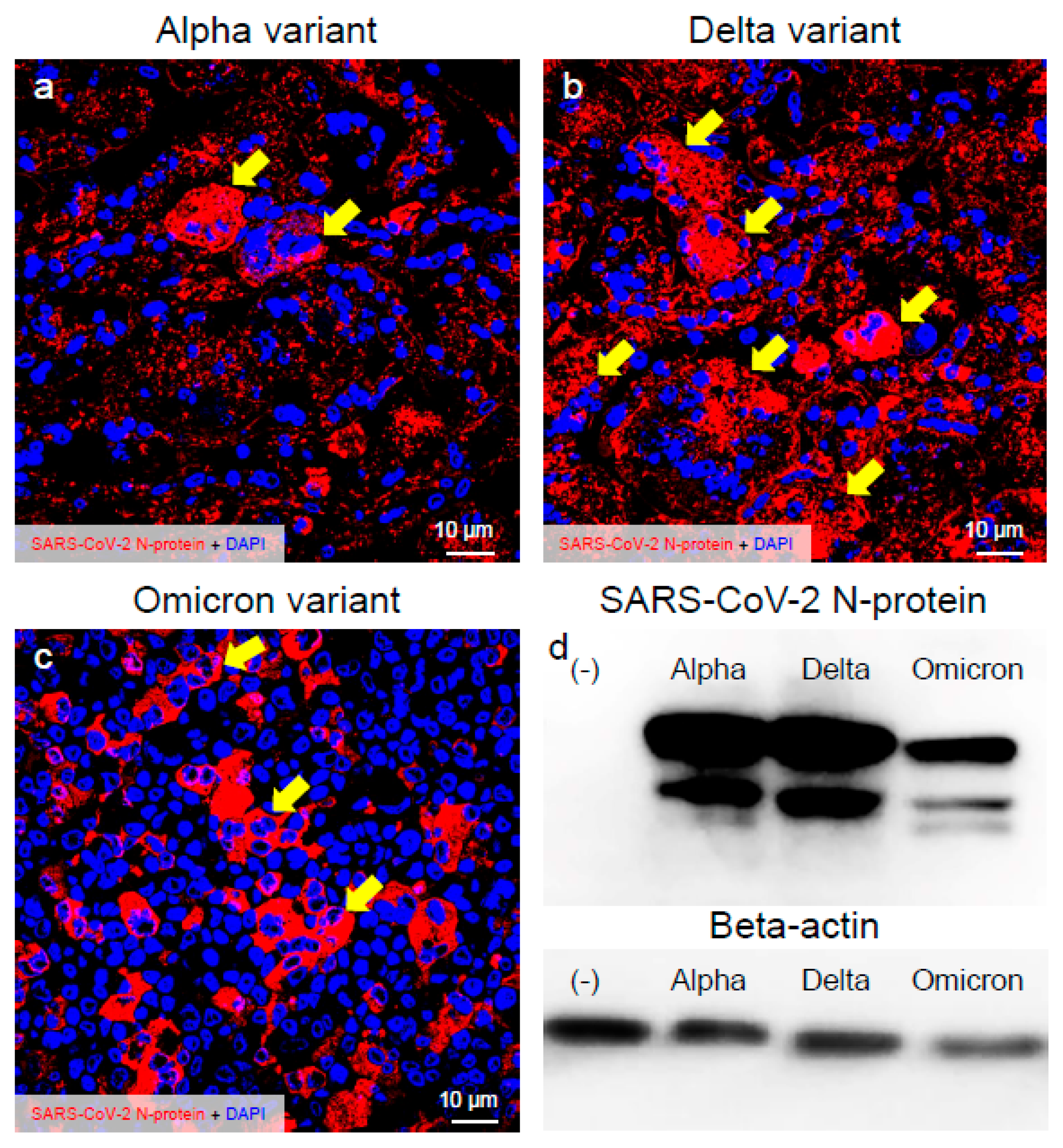
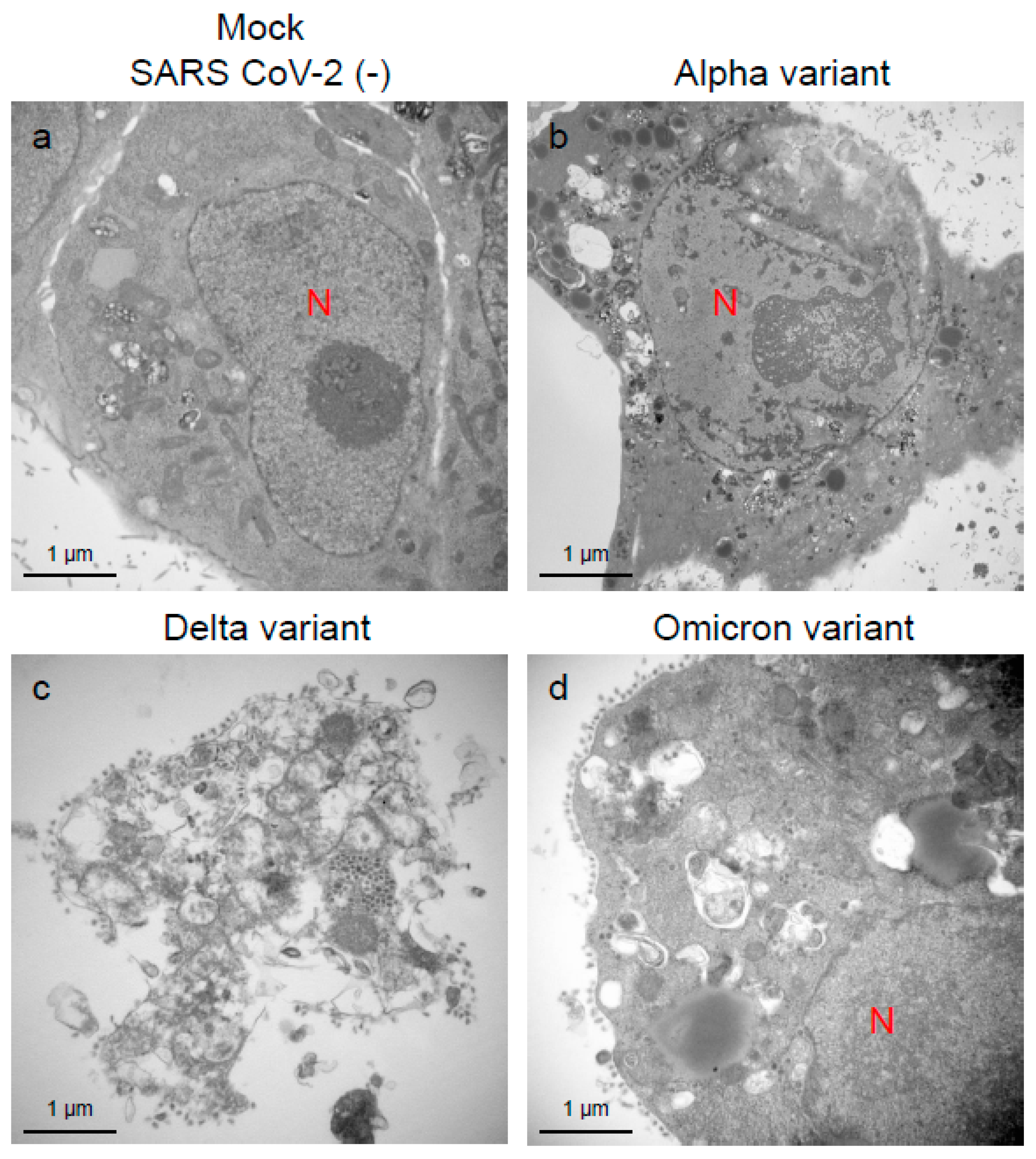
2.1. Delta Variant of SARS-CoV-2 Showed More Infectivity than the Alpha or Omicron Variant in VeroE6/TMPRSS2 Cells
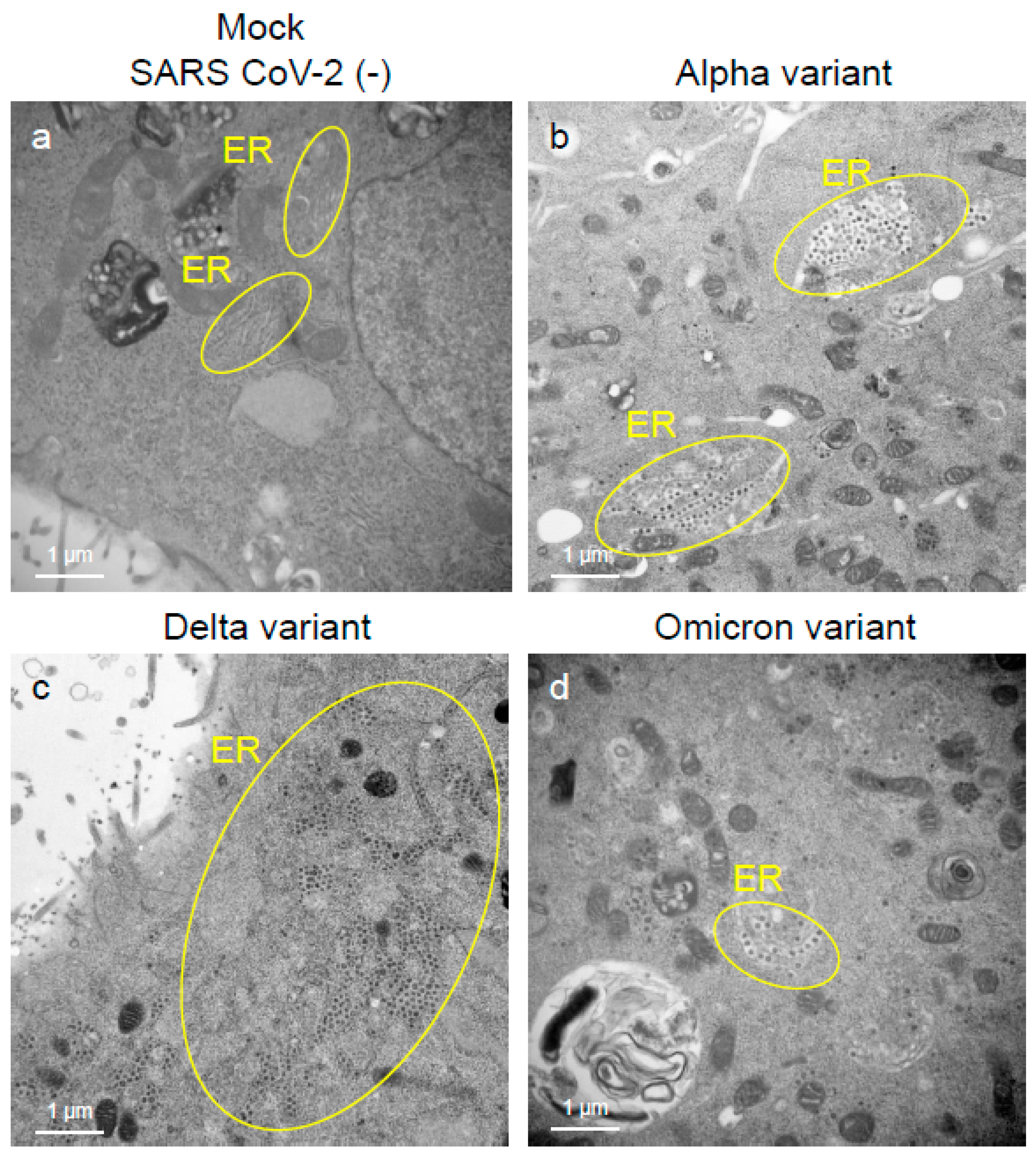
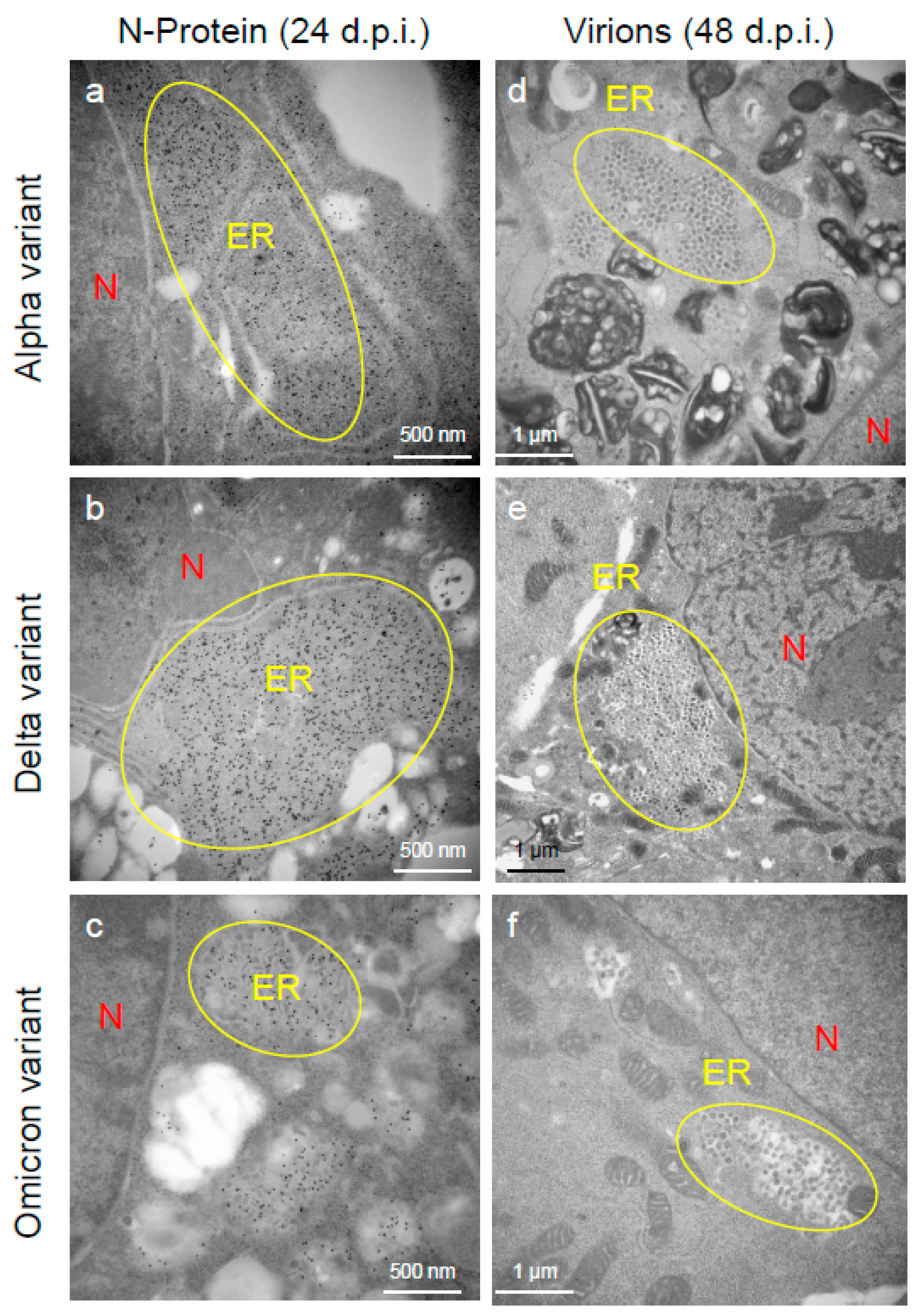
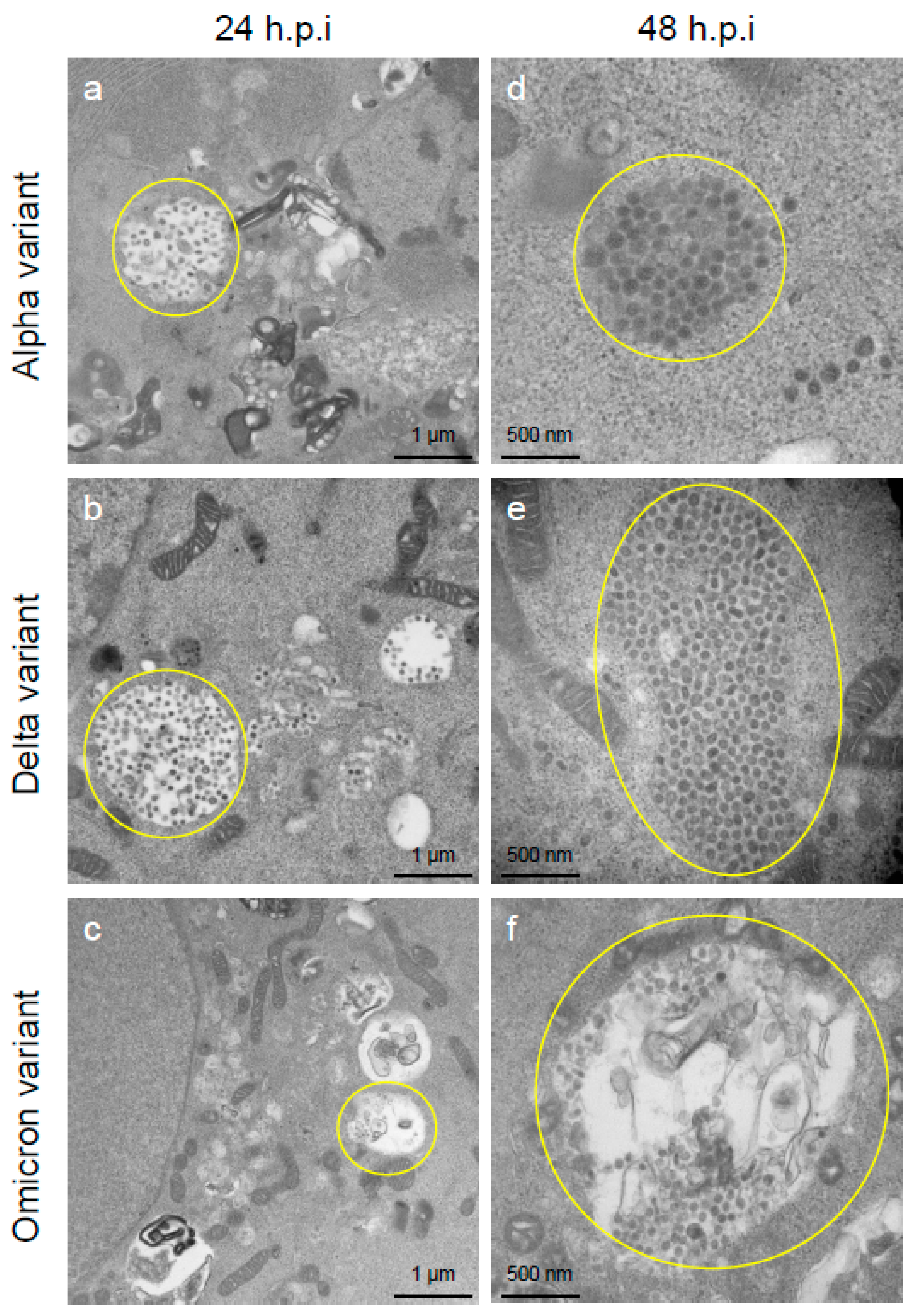
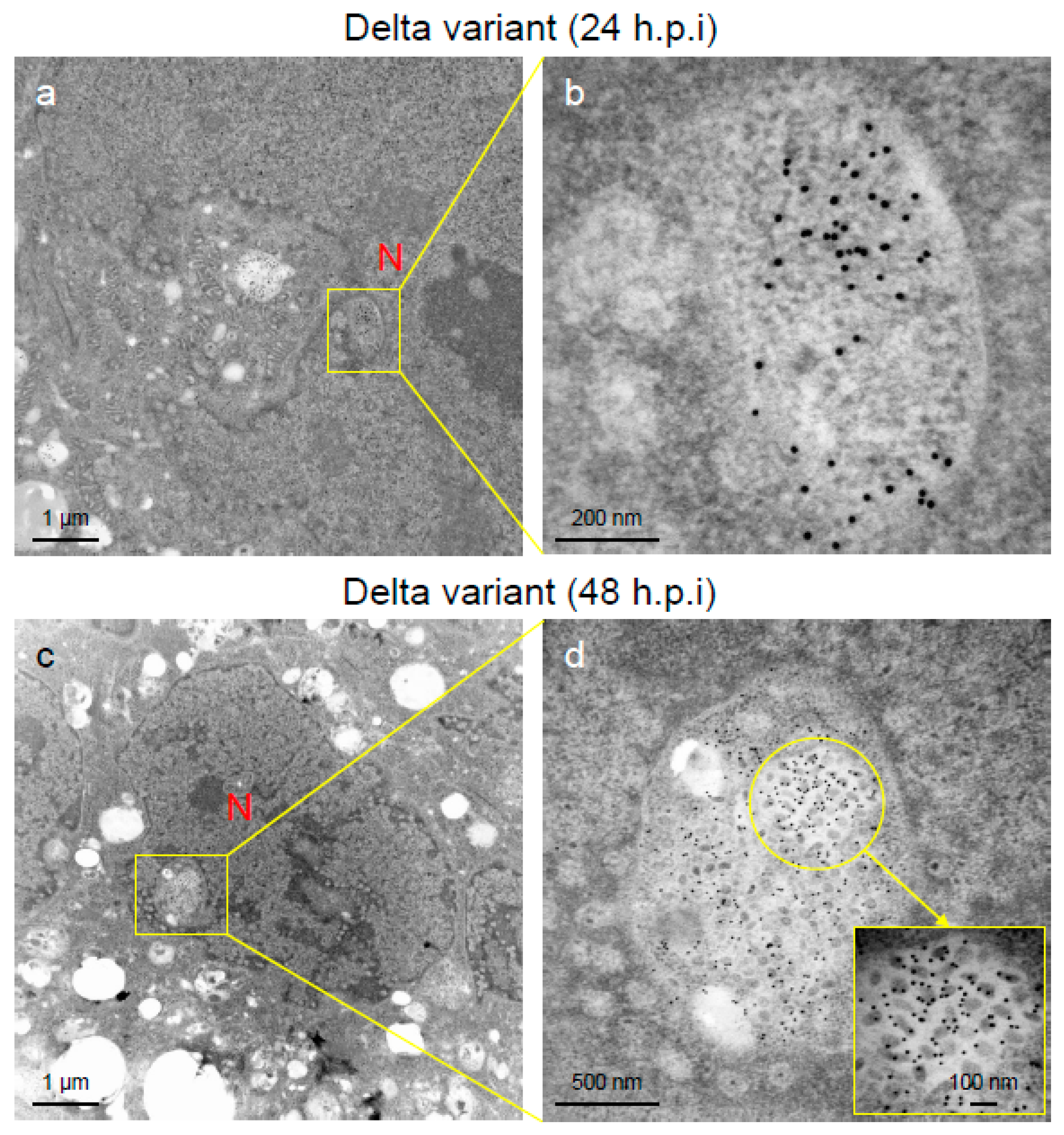
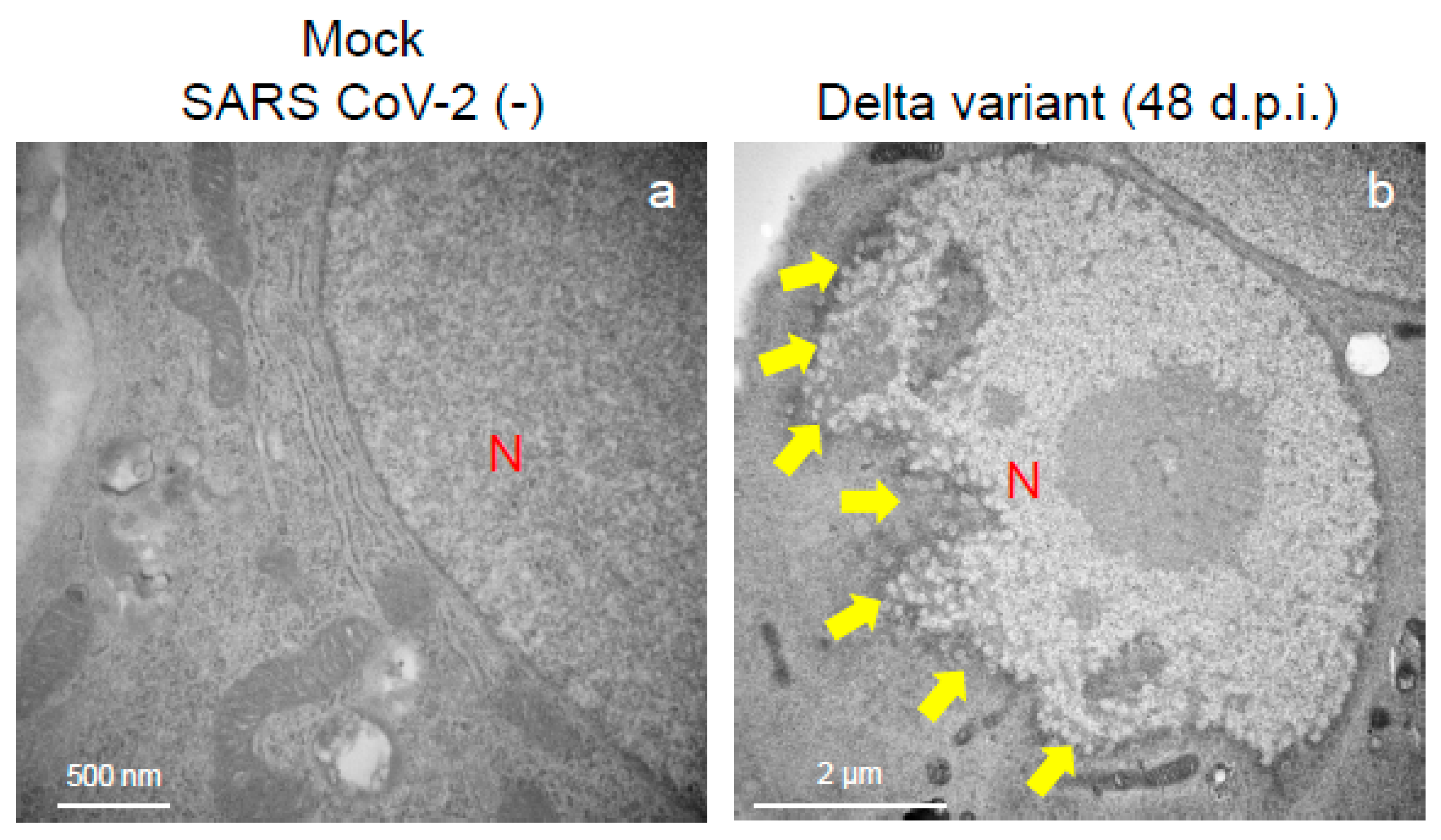
2.2. Delta Variant of SARS-CoV-2 Showed More Replicative and Pathogenic Characteristics than the Alpha or Omicron Variant in VeroE6/TMPRSS2 Cells
2.3. Unique Mutations in N- and Spike-Protein May Link to Increased Infectivity and Pathogenicity of the Delta Variant of SARS-CoV-2
3. Discussion
4. Materials and Methods
4.1. Cells and SARS-CoV-2 Infection
4.2. Analysis of Infection Patterns of VOCs of SARS-Cov-2 by Immunofluorescence Staining and Western Blotting
4.3. Analysis of Viral Replication and Pathogenic Features of VOCs of SARS-CoV-2 by Electron Microscopy
4.4. Analysis of Pathological Features of VOCs of SARS-CoV-2 by Hematoxylin Staining
4.5. Analysis of Genomic Variations in VOCs of SARS-CoV-2
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO COVID-19 Dashboard. Available online: https://data.who.int/dashboards/covid19/cases (accessed on 1 June 2024).
- Gopalan, A.; Tyagi, H. How reliable are test numbers for revealing the COVID-19 ground truth and applying Interventions? J. Indian Inst. Sci. 2020, 100, 863–884. [Google Scholar] [CrossRef] [PubMed]
- Jewell, N.P.; Lewnard, J.A. On the use of the reproduction number for SARS-CoV-2: Estimation, misinterpretations and relationships with other ecological measures. J. R. Stat. Soc. Ser. A Stat. Soc. 2022, 185, S16–S27. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Cases in Tokyo Likely 4 Times the Official Count. Available online: https://www.asahi.com/ajw/articles/14481175 (accessed on 8 November 2023).
- Carabelli, A.M.; Peacock, T.P.; Thorne, L.G.; Harvey, W.T.; Hughes, J.; COVID-19 Genomics UK Consortium; Peacock, S.J.; Barclay, W.S.; de Silva, T.I.; Towers, G.J.; et al. SARS-CoV-2 variant biology: Immune escape, transmission and fitness. Nat. Rev. Microbiol. 2023, 21, 162–177. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 Genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Denaro, M.; Ferro, E.; Barrano, G.; Meli, S.; Busacca, M.; Corallo, D.; Capici, A.; Zisa, A.; Cucuzza, L.; Gradante, S.; et al. Monitoring of SARS-CoV-2 Infection in Ragusa Area: Next Generation Sequencing and Serological Analysis. Int. J. Mol. Sci. 2023, 24, 4742. [Google Scholar] [CrossRef]
- Saberiyan, M.; Karimi, E.; Khademi, Z.; Movahhed, P.; Safi, A.; Mehri-Ghahfarrokhi, A. SARS-CoV-2: Phenotype, genotype, and characterization of different variants. Cell Mol. Biol. Lett. 2022, 27, 50. [Google Scholar] [CrossRef]
- Sigal, A. Milder disease with Omicron: Is it the virus or the pre-existing immunity? Nat. Rev. Immunol. 2022, 22, 69–71. [Google Scholar] [CrossRef]
- Xia, Q.; Yang, Y.; Wang, F.; Huang, Z.; Qiu, W.; Mao, A. Case fatality rates of COVID-19 during epidemic periods of variants of concern: A meta-analysis by continents. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2024, 141, 106950. [Google Scholar] [CrossRef]
- Auvigne, V.; Vaux, S.; Strat, Y.L.; Schaeffer, J.; Fournier, L.; Tamandjou, C.; Montagnat, C.; Coignard, B.; Levy-Bruhl, D.; Parent du Châtelet, I. Severe hospital events following symptomatic infection with Sars-CoV-2 Omicron and Delta variants in France, December 2021-January 2022: A retrospective, population-based, matched cohort study. EClinicalMedicine 2022, 48, 101455. [Google Scholar] [CrossRef]
- Jung, C.; Kmiec, D.; Koepke, L.; Zech, F.; Jacob, T.; Sparrer, K.M.J.; Kirchhoff, F. Omicron: What Makes the Latest SARS-CoV-2 Variant of Concern So Concerning? J. Virol. 2022, 96, e0207721. [Google Scholar] [CrossRef]
- Fan, Y.; Li, X.; Zhang, L.; Wan, S.; Zhang, L.; Zhou, F. SARS-CoV-2 Omicron variant: Recent progress and future perspectives. Signal Transduct. Target. Ther. 2022, 7, 141. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Cho, K.; Song, J.; Rahmati, M.; Koyanagi, A.; Lee, S.W.; Yon, D.K.; Il Shin, J.; Smith, L. The case fatality rate of COVID-19 during the Delta and the Omicron epidemic phase: A meta-analysis. J. Med. Virol. 2023, 95, e28522. [Google Scholar] [CrossRef] [PubMed]
- Qinfen, Z.; Jinming, C.; Xiaojun, H.; Huanying, Z.; Jicheng, H.; Ling, F.; Kunpeng, L.; Jingqiang, Z. The life cycle of SARS coronavirus in Vero E6 cells. J. Med. Virol. 2004, 73, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Irie, T.; Suzuki, R.; Maemura, T.; Nasser, H.; Uriu, K.; Kosugi, Y.; Shirakawa, K.; Sadamasu, K.; Kimura, I.; et al. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation. Nature 2022, 602, 300–306. [Google Scholar] [CrossRef]
- Zhao, H.; Nguyen, A.; Wu, D.; Li, Y.; Hassan, S.A.; Chen, J.; Shroff, H.; Piszczek, G.; Schuck, P. Plasticity in structure and assembly of SARS-CoV-2 nucleocapsid protein. PNAS Nexus 2022, 1, pgac049. [Google Scholar] [CrossRef]
- Hui, K.P.Y.; Ho, J.C.W.; Cheung, M.C.; Ng, K.C.; Ching, R.H.H.; Lai, K.L.; Kam, T.T.; Gu, H.; Sit, K.Y.; Hsin, M.K.Y.; et al. SARS-CoV-2 Omicron variant replication in human bronchus and lung ex vivo. Nature 2022, 603, 715–720. [Google Scholar] [CrossRef]
- Wei, L.; Song, L.; Dunker, A.K.; Foster, J.A.; Uversky, V.N.; Goh, G.K. A Comparative Experimental and Computational Study on the Nature of the Pangolin-CoV and COVID-19 Omicron. Int. J. Mol. Sci. 2024, 25, 7537. [Google Scholar] [CrossRef]
- Goh, G.K.; Dunker, A.K.; Foster, J.A.; Uversky, V.N. A Study on the Nature of SARS-CoV-2 Using the Shell Disorder Models: Reproducibility, Evolution, Spread, and Attenuation. Biomolecules 2022, 12, 1353. [Google Scholar] [CrossRef]
- Shuai, H.; Chan, J.F.; Hu, B.; Chai, Y.; Yuen, T.T.; Yin, F.; Huang, X.; Yoon, C.; Hu, J.C.; Liu, H.; et al. Attenuated replication and pathogenicity of SARS-CoV-2 B.1.1.529 Omicron. Nature 2022, 603, 693–699. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Gómez, S.A.; Rojas-Valencia, N.; Gómez, S.; Egidi, F.; Cappelli, C.; Restrepo, A. Binding of SARS-CoV-2 to Cell Receptors: A Tale of Molecular Evolution. ChemBioChem A Eur. J. Chem. Biol. 2021, 22, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhou, H.; Huang, W.; Zhou, J.; Qiu, M.; Deng, Z.; Chen, L.; Weng, Y.; Cai, L.; Gu, Y.; et al. Cell morphological analysis of SARS-CoV-2 infection by transmission electron microscopy. J. Thorac. Dis. 2020, 12, 4368–4373. [Google Scholar] [CrossRef]
- Siddell, S.; Wege, H.; Ter Meulen, V. The biology of coronaviruses. J. Gen. Virol. 1983, 64, 761–776. [Google Scholar] [CrossRef]
- Chen, H.; Wurm, T.; Britton, P.; Brooks, G.; Hiscox, J.A. Interaction of the coronavirus nucleoprotein with nucleolar antigens and the host cell. J. Virol. 2002, 76, 5233–5250. [Google Scholar] [CrossRef]
- Nguyen, V.P.; Hogue, B.G. Protein interactions during coronavirus assembly. J. Virol. 1997, 71, 9278–9284. [Google Scholar] [CrossRef]
- Evans, M.R.; Simpson, R.W. The coronavirus avian infectious bronchitis virus requires the cell nucleus and host transcriptional factors. Virology 1980, 105, 582–591. [Google Scholar] [CrossRef]
- Wilhelmsen, K.C.; Leibowitz, J.L.; Bond, C.W.; Robb, J.A. The replication of murine coronaviruses in enucleated cells. Virology 1981, 110, 225–230. [Google Scholar] [CrossRef]
- Chen, M.; Ma, Y.; Chang, W. SARS-CoV-2 and the Nucleus. Int. J. Biol. Sci. 2022, 18, 4731–4743. [Google Scholar] [CrossRef]
- McBride, R.; van Zyl, M.; Fielding, B.C. The coronavirus nucleocapsid is a multifunctional protein. Viruses 2014, 6, 2991–3018. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Cheng, Y.; Zhou, H.; Sun, C.; Zhang, S. The SARS-CoV-2 nucleocapsid protein: Its role in the viral life cycle, structure and functions, and use as a potential target in the development of vaccines and diagnostics. Virol. J. 2023, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Zhang, H.; Zhang, Y.; Lin, K.; Zhang, Y.; Wu, J.; Wan, Y.; Huang, Y.; Song, J.; Fu, Z.; et al. Omicron variant showed lower neutralizing sensitivity than other SARS-CoV-2 variants to immune sera elicited by vaccines after boost. Emerg. Microbes Infect. 2022, 11, 337–343. [Google Scholar] [CrossRef]
- Duś-Ilnicka, I.; Mazur, M.; Rybińska, A.; Radwan-Oczko, M.; Jurczyszyn, K.; Paradowska-Stolarz, A. SARS CoV-2 IgG seropositivity post-vaccination among dental professionals: A prospective study. BMC Infect. Dis. 2023, 23, 539. [Google Scholar] [CrossRef]
- Chen, C.; Haupert, S.R.; Zimmermann, L.; Shi, X.; Fritsche, L.G.; Mukherjee, B. Global Prevalence of Post-Coronavirus Disease 2019 (COVID-19) Condition or Long COVID: A Meta-Analysis and Systematic Review. J. Infect. Dis. 2022, 226, 1593–1607. [Google Scholar] [CrossRef]
- Nyasulu, P.S.; Tamuzi, J.L.; Erasmus, R.T. Burden, causation, and particularities of Long COVID in African populations: A rapid systematic review. IJID Reg. 2023, 8, 137–144. [Google Scholar] [CrossRef]
- Kumar, S.; Thambiraja, T.S.; Karuppanan, K.; Subramaniam, G. Omicron and Delta variant of SARS-CoV-2: A comparative computational study of spike protein. J. Med. Virol. 2022, 94, 1641–1649. [Google Scholar] [CrossRef]
- Jack, A.; Ferro, L.S.; Trnka, M.J.; Wehri, E.; Nadgir, A.; Nguyenla, X.; Fox, D.; Costa, K.; Stanley, S.; Schaletzky, J.; et al. SARS-CoV-2 nucleocapsid protein forms condensates with viral genomic RNA. PLoS Biol. 2021, 19, e3001425. [Google Scholar] [CrossRef]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata., N.; Sekizuka, T.; Katoh, H.; Kato, F.; et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.; Yahiro, T.; Kimitsuki, K.; Hashimoto, T.; Matsuura, K.; Yano, S.; Noguchi, K.; Sonezaki, A.; Yoshizawa, K.; Kumasako, Y.; et al. Exploring the Replication and Pathogenic Characteristics of Alpha, Delta, and Omicron Variants of SARS-CoV-2. Int. J. Mol. Sci. 2024, 25, 12641. https://doi.org/10.3390/ijms252312641
Khan S, Yahiro T, Kimitsuki K, Hashimoto T, Matsuura K, Yano S, Noguchi K, Sonezaki A, Yoshizawa K, Kumasako Y, et al. Exploring the Replication and Pathogenic Characteristics of Alpha, Delta, and Omicron Variants of SARS-CoV-2. International Journal of Molecular Sciences. 2024; 25(23):12641. https://doi.org/10.3390/ijms252312641
Chicago/Turabian StyleKhan, Sakirul, Takaaki Yahiro, Kazunori Kimitsuki, Takehiro Hashimoto, Keiko Matsuura, Shinji Yano, Kazuko Noguchi, Akane Sonezaki, Kaori Yoshizawa, Yoko Kumasako, and et al. 2024. "Exploring the Replication and Pathogenic Characteristics of Alpha, Delta, and Omicron Variants of SARS-CoV-2" International Journal of Molecular Sciences 25, no. 23: 12641. https://doi.org/10.3390/ijms252312641
APA StyleKhan, S., Yahiro, T., Kimitsuki, K., Hashimoto, T., Matsuura, K., Yano, S., Noguchi, K., Sonezaki, A., Yoshizawa, K., Kumasako, Y., Akbar, S. M. F., & Nishizono, A. (2024). Exploring the Replication and Pathogenic Characteristics of Alpha, Delta, and Omicron Variants of SARS-CoV-2. International Journal of Molecular Sciences, 25(23), 12641. https://doi.org/10.3390/ijms252312641