
Evaluation of the LDN-0060609 PERK Inhibitor as a Selective Treatment for Primary Open-Angle Glaucoma: An In Vitro Study on Human Retinal Astrocytes

 , , , , , , and
, , , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
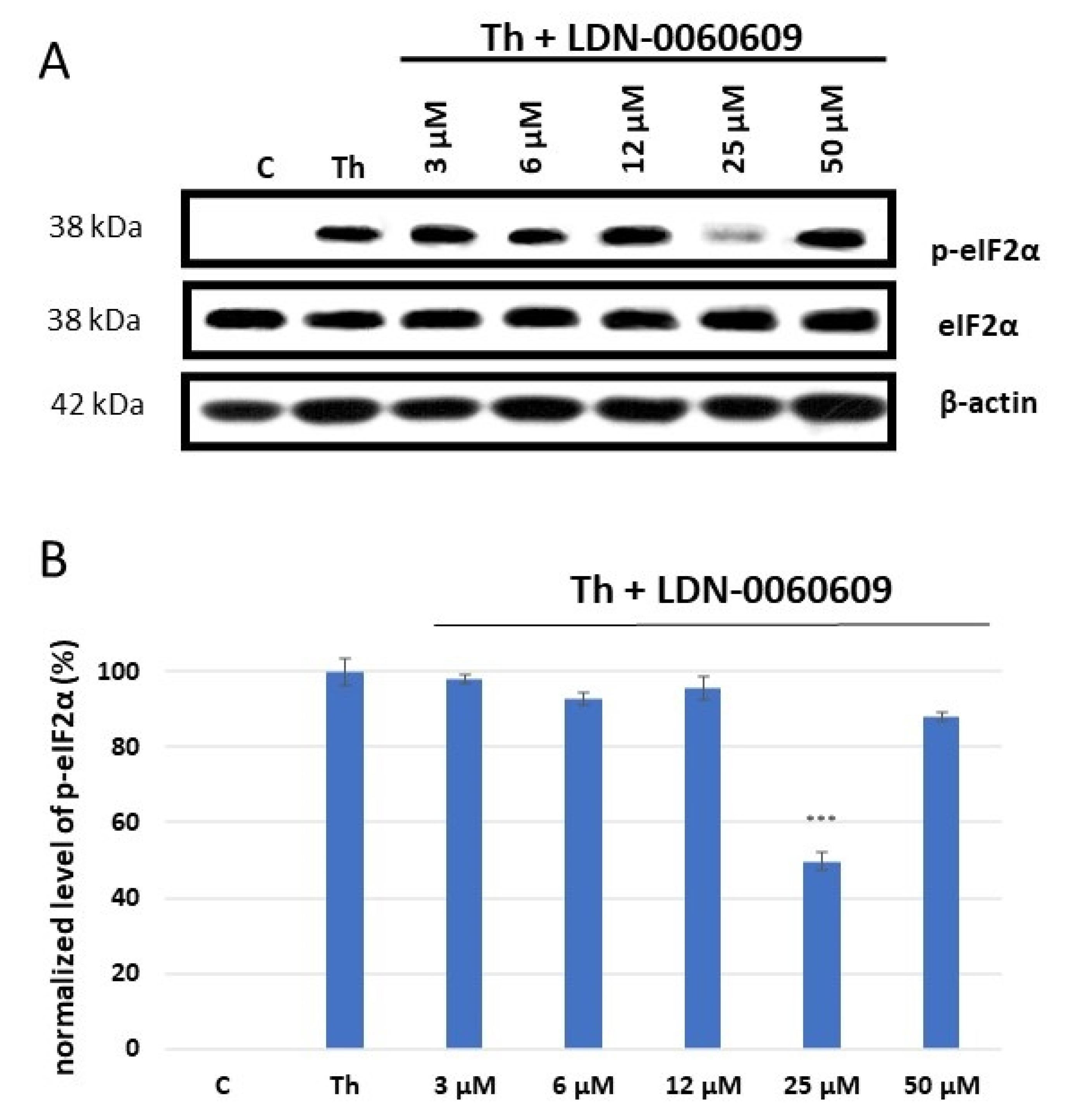
2.1. The Effect of the LDN-0060609 PERK Inhibitor on HRA Cells with Activated ER Stress Conditions
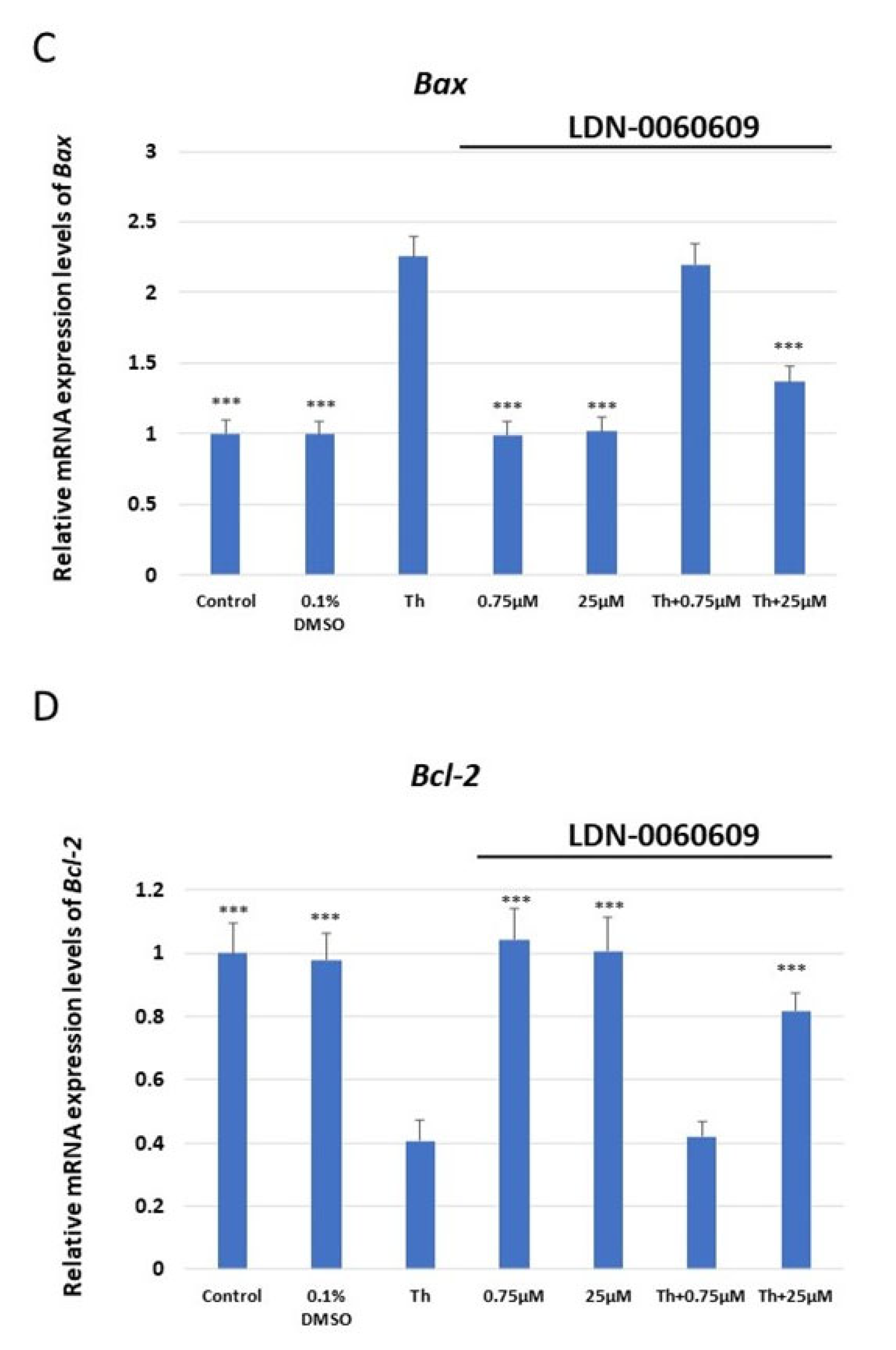
2.2. Evaluation of mRNA Expression via ER Stress-Related Genes in HRA Cells Treated with LDN-0060609
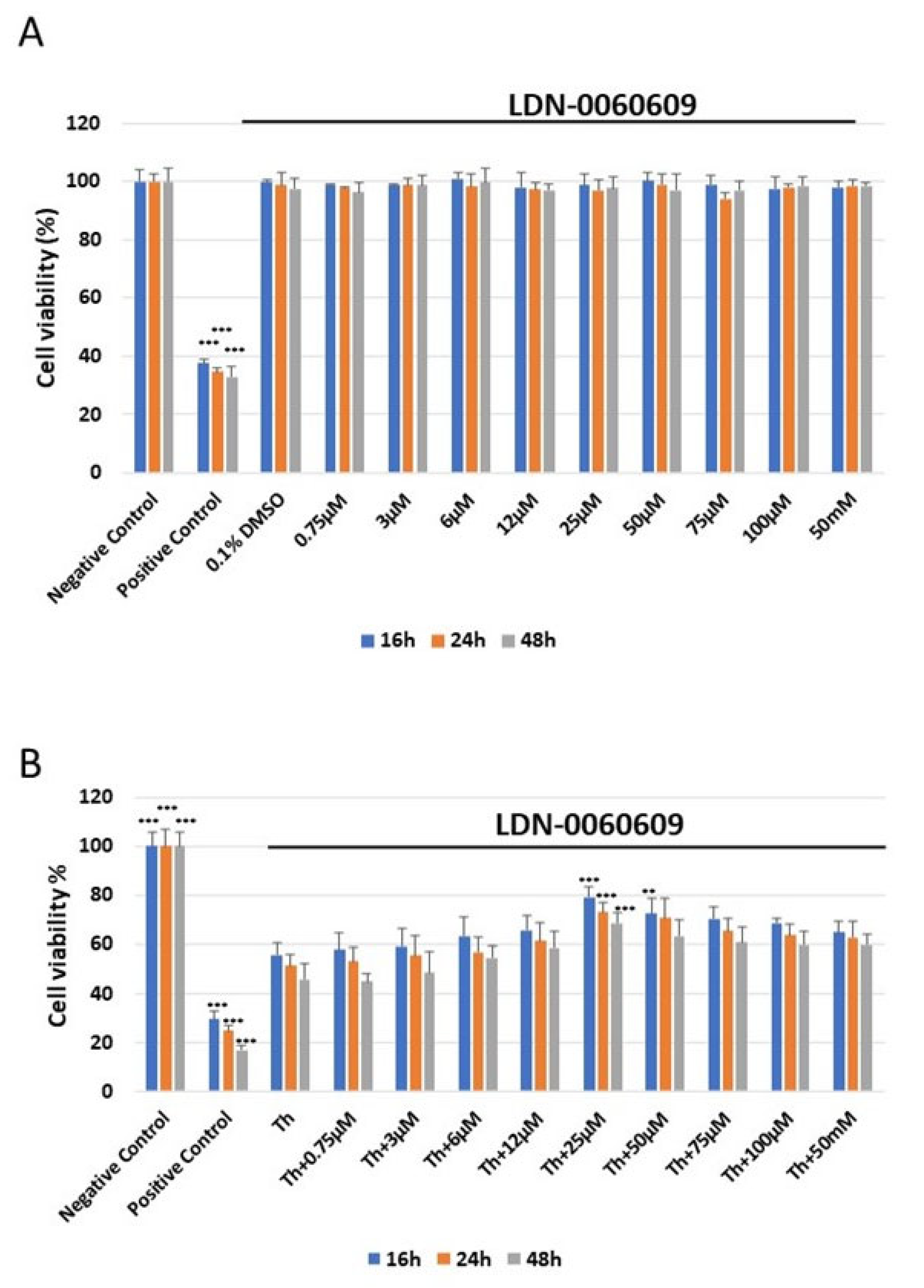
2.3. Assessment of the Potential Cytotoxic Acivity of the LDN-0060609 Compound
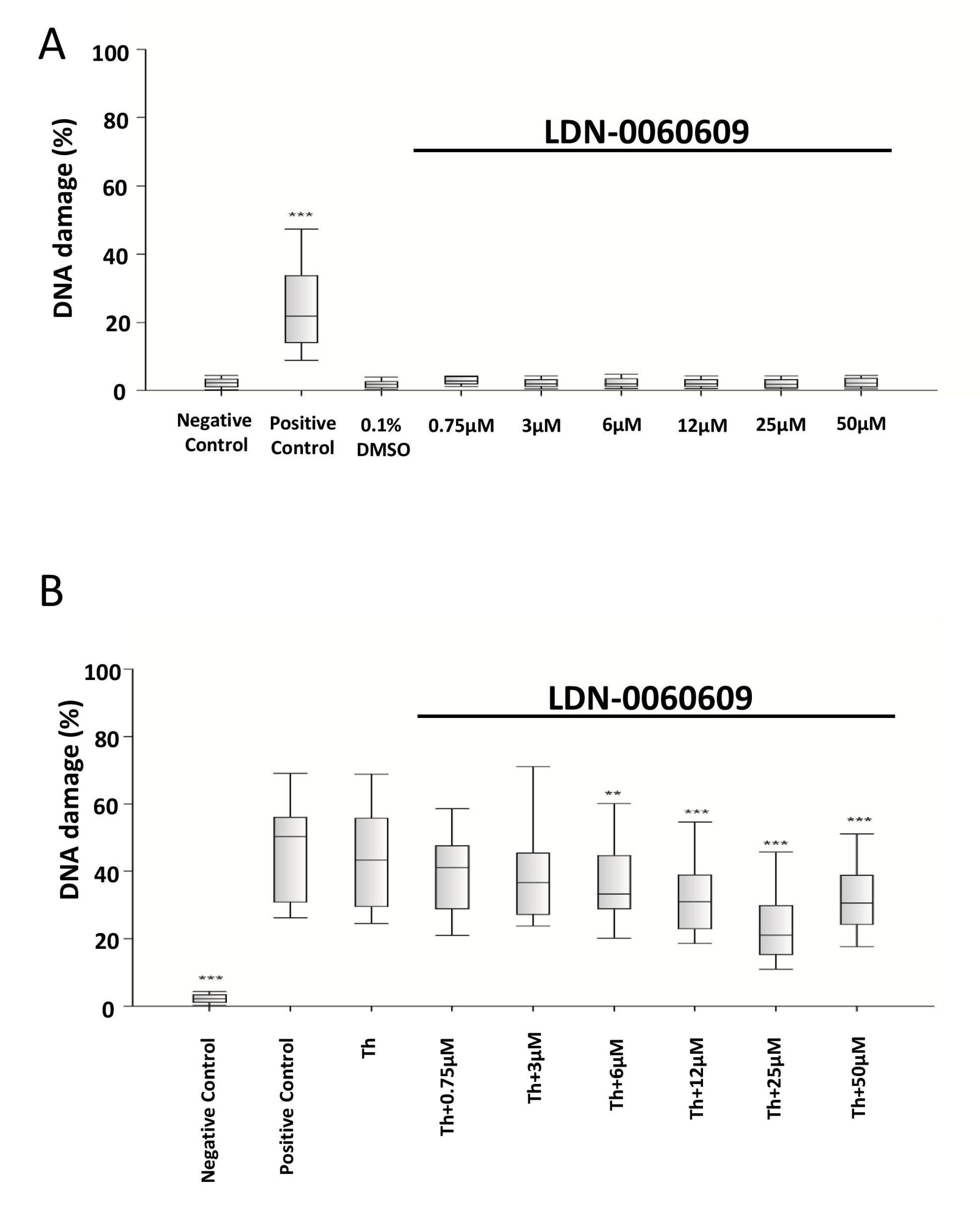
2.4. Evaluation of the Genotoxic Poential of the LDN-0060609 Compound
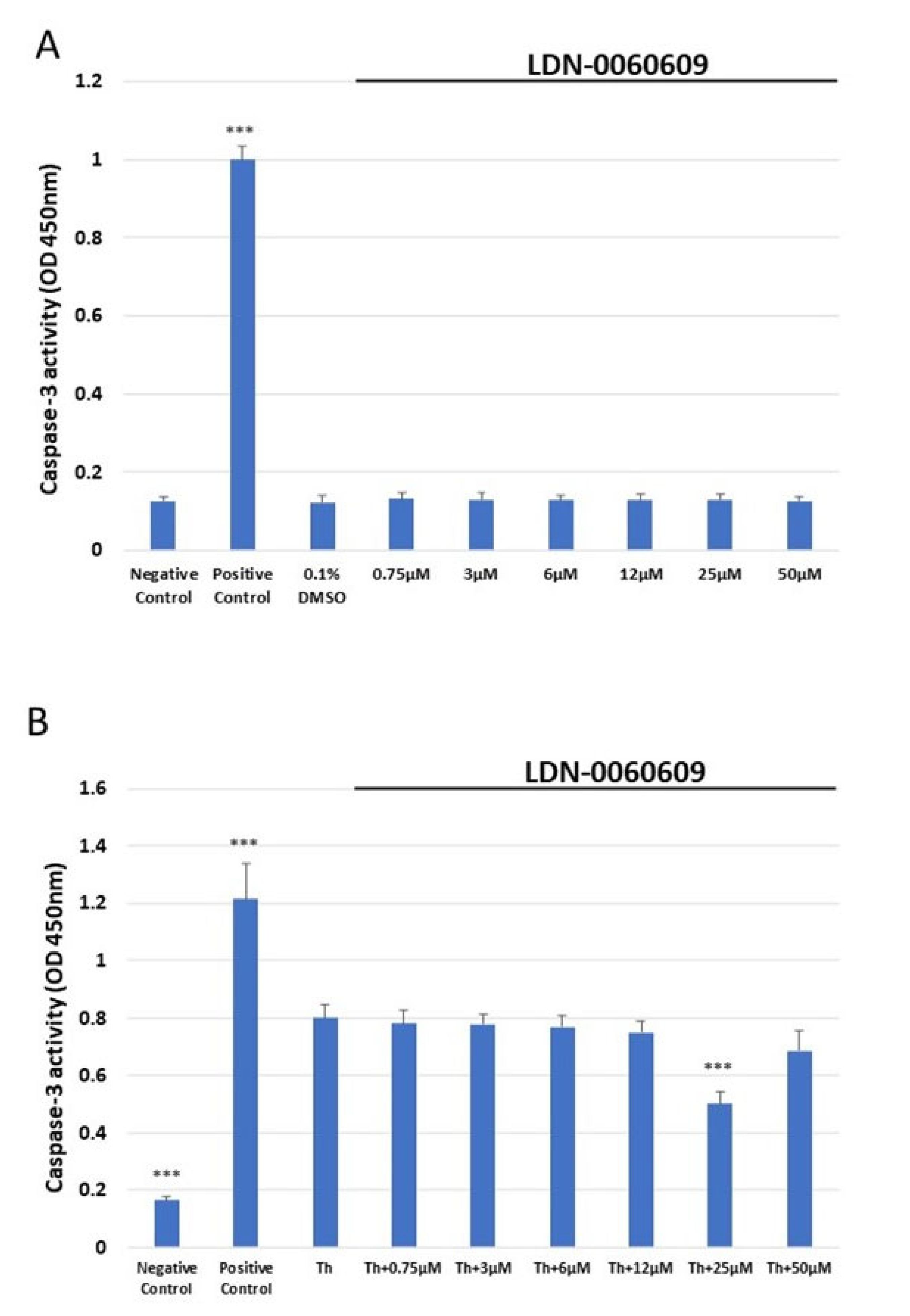
2.5. Analysis of the Level of Apoptosis in HRA Cells Using a Caspase-3 Assay
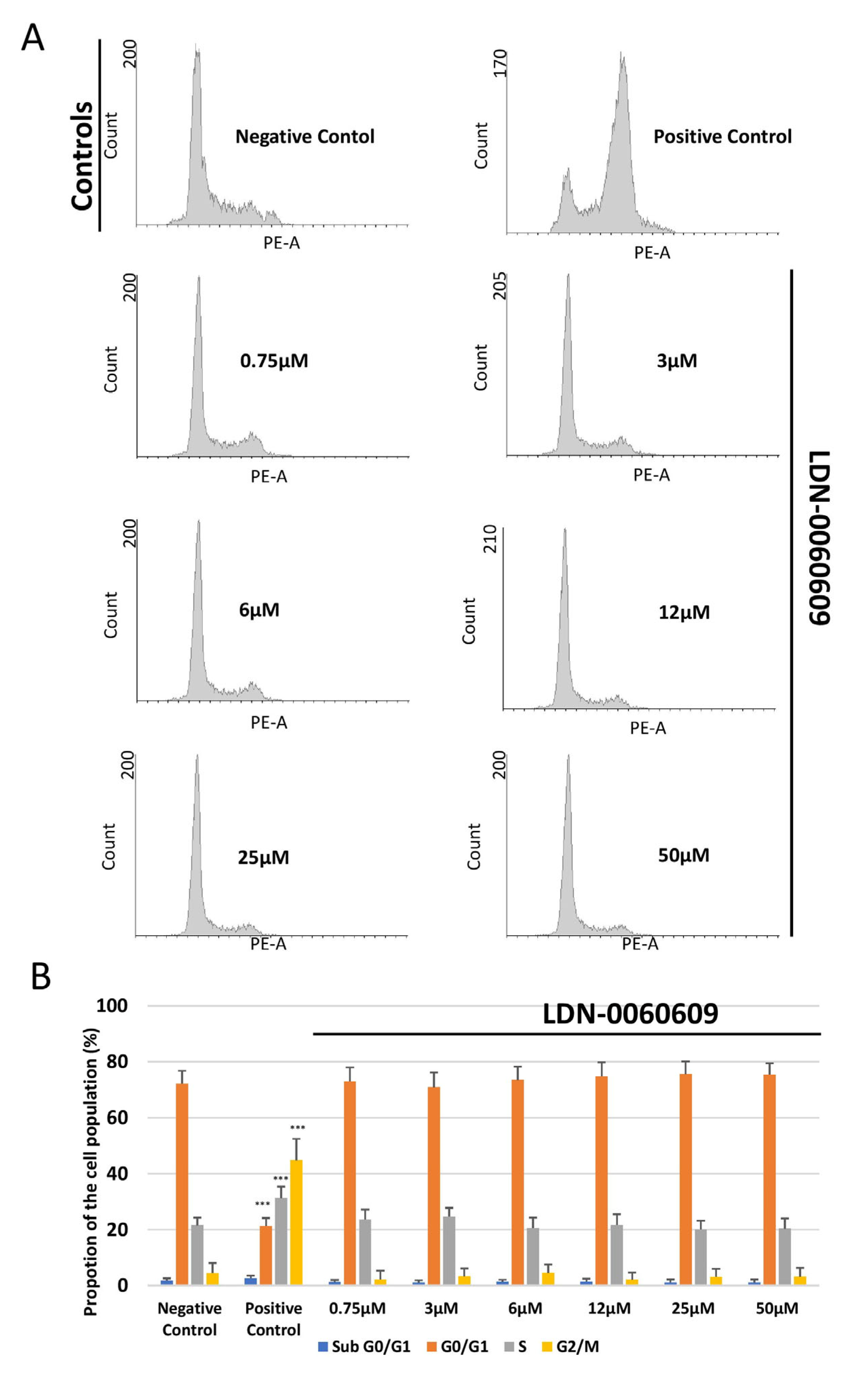
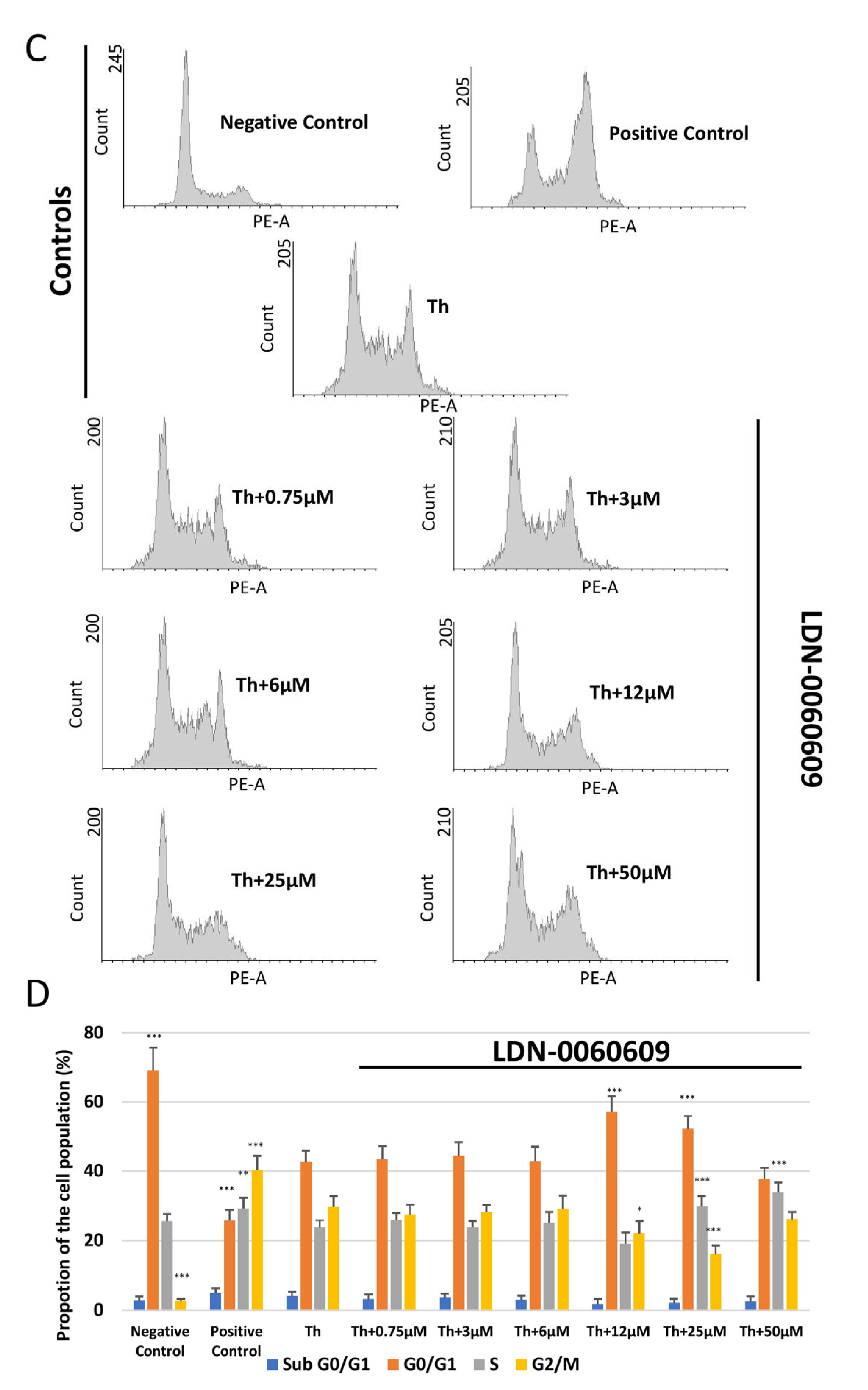
2.6. Assessment of the Effect of LDN-0060609 on Cell Cycle Distribution and Progression in HRA Cells
3. Discussion
4. Materials and Methods
4.1. The Identification of the LDN-0060609 PERK Inhibitor
4.2. Cell Culture
4.3. Western Blotting
4.4. Assesment of the Expression of the ER Stress-Related Genes
4.5. Cytotoxicity Analysis
4.6. Genotoxicity Analysis
4.7. Apoptosis Analysis
4.8. Cell Cycle Analysis
4.9. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jayaram, H.; Kolko, M.; Friedman, D.S.; Gazzard, G. Glaucoma: Now and Beyond. Lancet 2023, 402, 1788–1801. [Google Scholar] [CrossRef] [PubMed]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global Prevalence of Glaucoma and Projections of Glaucoma Burden through 2040: A Systematic Review and Meta-Analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Burton, M.J.; Ramke, J.; Marques, A.P.; Bourne, R.R.A.; Congdon, N.; Jones, I.; Ah Tong, B.A.M.; Arunga, S.; Bachani, D.; Bascaran, C.; et al. The Lancet Global Health Commission on Global Eye Health: Vision beyond 2020. Lancet Glob. Health 2021, 9, e489–e551. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, T.; Medina, A.; Perkins, J.; Yera, M.; Wang, J.J.; Zhang, S.X. Cellular Stress Signaling and the Unfolded Protein Response in Retinal Degeneration: Mechanisms and Therapeutic Implications. Mol. Neurodegener. 2022, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shi, C.; He, M.; Xiong, S.; Xia, X. Endoplasmic Reticulum Stress: Molecular Mechanism and Therapeutic Targets. Signal Transduct. Target. Ther. 2023, 8, 352. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Qian, T.; He, R.; Wan, C.; Liu, Y.; Yu, H. Endoplasmic Reticulum–Plasma Membrane Contact Sites: Regulators, Mechanisms, and Physiological Functions. Front. Cell Dev. Biol. 2021, 9, 627700. [Google Scholar] [CrossRef] [PubMed]
- Rozpędek-Kamińska, W.; Wojtczak, R.; Szaflik, J.P.; Szaflik, J.; Majsterek, I. The Genetic and Endoplasmic Reticulum-Mediated Molecular Mechanisms of Primary Open-Angle Glaucoma. Int. J. Mol. Sci. 2020, 21, 4171. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, R.L.; Ma, M.T. Molecular Insights into Myocilin and Its Glaucoma-Causing Misfolded Olfactomedin Domain Variants. Acc. Chem. Res. 2021, 54, 2205–2215. [Google Scholar] [CrossRef]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, Regulation and Functions of the Unfolded Protein Response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef]
- Cullen, P.F.; Sun, D. Astrocytes of the Eye and Optic Nerve: Heterogeneous Populations with Unique Functions Mediate Axonal Resilience and Vulnerability to Glaucoma. Front. Ophthalmol. 2023, 3, 1217137. [Google Scholar] [CrossRef]
- Yoo, H.-S.; Shanmugalingam, U.; Smith, P.D. Harnessing Astrocytes and Müller Glial Cells in the Retina for Survival and Regeneration of Retinal Ganglion Cells. Cells 2021, 10, 1339. [Google Scholar] [CrossRef]
- Müller, A.; Hauk, T.G.; Leibinger, M.; Marienfeld, R.; Fischer, D. Exogenous CNTF Stimulates Axon Regeneration of Retinal Ganglion Cells Partially via Endogenous CNTF. Mol. Cell. Neurosci. 2009, 41, 233–246. [Google Scholar] [CrossRef]
- Boia, R.; Ruzafa, N.; Aires, I.D.; Pereiro, X.; Ambrósio, A.F.; Vecino, E.; Santiago, A.R. Neuroprotective Strategies for Retinal Ganglion Cell Degeneration: Current Status and Challenges Ahead. Int. J. Mol. Sci. 2020, 21, 2262. [Google Scholar] [CrossRef] [PubMed]
- Calkins, D.J. Adaptive Responses to Neurodegenerative Stress in Glaucoma. Prog. Retin. Eye Res. 2021, 84, 100953. [Google Scholar] [CrossRef] [PubMed]
- Alqawlaq, S.; Flanagan, J.G.; Sivak, J.M. All Roads Lead to Glaucoma: Induced Retinal Injury Cascades Contribute to a Common Neurodegenerative Outcome. Exp. Eye Res. 2019, 183, 88–97. [Google Scholar] [CrossRef] [PubMed]
- García-Bermúdez, M.Y.; Freude, K.K.; Mouhammad, Z.A.; van Wijngaarden, P.; Martin, K.K.; Kolko, M. Glial Cells in Glaucoma: Friends, Foes, and Potential Therapeutic Targets. Front. Neurol. 2021, 12, 624983. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Mesentier-Louro, L.A.; Oh, A.J.; Heng, K.; Shariati, M.A.; Huang, H.; Hu, Y.; Liao, Y.J. Increased ER Stress after Experimental Ischemic Optic Neuropathy and Improved RGC and Oligodendrocyte Survival after Treatment with Chemical Chaperon. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1953–1966. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, R.; Pappas, A.C.; Seifert, P.; Savol, A.; Sadreyev, R.I.; Sun, D.; Jakobs, T.C. Astrocytes in the Optic Nerve Are Heterogeneous in Their Reactivity to Glaucomatous Injury. Cells 2023, 12, 2131. [Google Scholar] [CrossRef]
- Hu, Y.; Park, K.K.; Yang, L.; Wei, X.; Yang, Q.; Cho, K.S.; Thielen, P.; Lee, A.H.; Cartoni, R.; Glimcher, L.H.; et al. Differential Effects of Unfolded Protein Response Pathways on Axon Injury-Induced Death of Retinal Ganglion Cells. Neuron 2012, 73, 445–452. [Google Scholar] [CrossRef]
- Yang, L.; Li, S.; Miao, L.; Huang, H.; Liang, F.; Teng, X.; Xu, L.; Wang, Q.; Xiao, W.; Ridder, W.H.; et al. Rescue of Glaucomatous Neurodegeneration by Differentially Modulating Neuronal Endoplasmic Reticulum Stress Molecules. J. Neurosci. 2016, 36, 5891–5903. [Google Scholar] [CrossRef]
- Peters, J.C.; Bhattacharya, S.; Clark, A.F.; Zode, G.S. Increased Endoplasmic Reticulum Stress in Human Glaucomatous Trabecular Meshwork Cells and Tissues. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3860–3868. [Google Scholar] [CrossRef] [PubMed]
- Kasetti, R.B.; Patel, P.D.; Maddineni, P.; Patil, S.; Kiehlbauch, C.; Millar, J.C.; Searby, C.C.; Raghunathan, V.K.; Sheffield, V.C.; Zode, G.S. ATF4 Leads to Glaucoma by Promoting Protein Synthesis and ER Client Protein Load. Nat. Commun. 2020, 11, 5594. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, S.J.; Garcia-Bonilla, L.; Hu, J.; Harding, H.P.; Ron, D. Activation-Dependent Substrate Recruitment by the Eukaryotic Translation Initiation Factor 2 Kinase PERK. J. Cell Biol. 2006, 172, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Tehrani, S.; Davis, L.; Cepurna, W.O.; Choe, T.E.; Lozano, D.C.; Monfared, A.; Cooper, L.; Cheng, J.; Johnson, E.C.; Morrison, J.C. Astrocyte Structural and Molecular Response to Elevated Intraocular Pressure Occurs Rapidly and Precedes Axonal Tubulin Rearrangement within the Optic Nerve Head in a Rat Model. PLoS ONE 2016, 11, e0167364. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.L.; Crish, S.D.; Inman, D.M.; Horner, P.J.; Calkins, D.J. Early Astrocyte Redistribution in the Optic Nerve Precedes Axonopathy in the DBA/2J Mouse Model of Glaucoma. Exp. Eye Res. 2016, 150, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Guttenplan, K.A.; Stafford, B.K.; El-Danaf, R.N.; Adler, D.I.; Münch, A.E.; Weigel, M.K.; Huberman, A.D.; Liddelow, S.A. Neurotoxic Reactive Astrocytes Drive Neuronal Death after Retinal Injury. Cell Rep. 2020, 31, 107776. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhao, G.-L.; Cheng, S.; Wang, Z.; Yang, X.-L. Activation of Retinal Glial Cells Contributes to the Degeneration of Ganglion Cells in Experimental Glaucoma. Prog. Retin. Eye Res. 2023, 93, 101169. [Google Scholar] [CrossRef]
- Cooper, M.L.; Collyer, J.W.; Calkins, D.J. Astrocyte Remodeling without Gliosis Precedes Optic Nerve Axonopathy. Acta Neuropathol. Commun. 2018, 6, 38. [Google Scholar] [CrossRef]
- Sun, D.; Moore, S.; Jakobs, T.C. Optic Nerve Astrocyte Reactivity Protects Function in Experimental Glaucoma and Other Nerve Injuries. J. Exp. Med. 2017, 214, 1411–1430. [Google Scholar] [CrossRef]
- Cooper, M.L.; Pasini, S.; Lambert, W.S.; D’Alessandro, K.B.; Yao, V.; Risner, M.L.; Calkins, D.J. Redistribution of Metabolic Resources through Astrocyte Networks Mitigates Neurodegenerative Stress. Proc. Natl. Acad. Sci. USA 2020, 117, 18810–18821. [Google Scholar] [CrossRef]
- Gomes, C.; VanderWall, K.B.; Pan, Y.; Lu, X.; Lavekar, S.S.; Huang, K.C.; Fligor, C.M.; Harkin, J.; Zhang, C.; Cummins, T.R.; et al. Astrocytes Modulate Neurodegenerative Phenotypes Associated with Glaucoma in OPTN(E50K) Human Stem Cell-Derived Retinal Ganglion Cells. Stem Cell Rep. 2022, 17, 1636–1649. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L.; Freeman, O.J.; Butcher, A.J.; Holmqvist, S.; Humoud, I.; Schätzl, T.; Hughes, D.T.; Verity, N.C.; Swinden, D.P.; Hayes, J.; et al. Astrocyte Unfolded Protein Response Induces a Specific Reactivity State That Causes Non-Cell-Autonomous Neuronal Degeneration. Neuron 2020, 105, 855–866.e5. [Google Scholar] [CrossRef] [PubMed]
- Ojino, K.; Shimazawa, M.; Izawa, H.; Nakano, Y.; Tsuruma, K.; Hara, H. Involvement of Endoplasmic Reticulum Stress in Optic Nerve Degeneration after Chronic High Intraocular Pressure in DBA/2J Mice. J. Neurosci. Res. 2015, 93, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- Mesentier-Louro, L.A.; Shariati, M.A.; Dalal, R.; Camargo, A.; Kumar, V.; Shamskhou, E.A.; de Jesus Perez, V.; Liao, Y.J. Systemic Hypoxia Led to Little Retinal Neuronal Loss and Dramatic Optic Nerve Glial Response. Exp. Eye Res. 2020, 193, 107957. [Google Scholar] [CrossRef] [PubMed]
- Benavides, A.; Pastor, D.; Santos, P.; Tranque, P.; Calvo, S. Plays a Pivotal Role in the Astrocyte Death Induced by Oxygen and Glucose Deprivation. Glia 2005, 52, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Han, J.C.; Park, D.Y.; Kee, C. A Neuroglia-Based Interpretation of Glaucomatous Neuroretinal Rim Thinning in the Optic Nerve Head. Prog. Retin. Eye Res. 2020, 77, 100840. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, L.; Pembroke, W.G.; Rexach, J.E.; Godoy, M.I.; Condro, M.C.; Alvarado, A.G.; Harteni, M.; Chen, Y.W.; Stiles, L.; et al. Conservation and Divergence of Vulnerability and Responses to Stressors between Human and Mouse Astrocytes. Nat. Commun. 2021, 12, 3958. [Google Scholar] [CrossRef]
- Mazumder, A.G.; Julé, A.M.; Sun, D. Astrocytes of the Optic Nerve Exhibit a Region-Specific and Temporally Distinct Response to Elevated Intraocular Pressure. Mol. Neurodegener. 2023, 18, 68. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, Y.; Chen, D. The Heterogeneity of Astrocytes in Glaucoma. Front. Neuroanat. 2022, 16, 995369. [Google Scholar] [CrossRef]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive Astrocyte Nomenclature, Definitions, and Future Directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Rozpędek-Kamińska, W.; Galita, G.; Siwecka, N.; Carroll, S.L.; Diehl, J.A.; Kucharska, E.; Pytel, D.; Majsterek, I. The Potential Role of Small-Molecule Perk Inhibitor Ldn-0060609 in Primary Open-Angle Glaucoma Treatment. Int. J. Mol. Sci. 2021, 22, 4494. [Google Scholar] [CrossRef]
- Pytel, D.; Seyb, K.; Liu, M.; Ray, S.S.; Concannon, J.; Huang, M.; Cuny, G.D.; Diehl, J.A.; Glicksman, M.A. Enzymatic Characterization of ER Stress-Dependent Kinase, PERK, and Development of a High-Throughput Assay for Identification of PERK Inhibitors. J. Biomol. Screen. 2014, 19, 1024–1034. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozpędek-Kamińska, W.; Galita, G.; Saramowicz, K.; Granek, Z.; Barczuk, J.; Siwecka, N.; Pytel, D.; Majsterek, I. Evaluation of the LDN-0060609 PERK Inhibitor as a Selective Treatment for Primary Open-Angle Glaucoma: An In Vitro Study on Human Retinal Astrocytes. Int. J. Mol. Sci. 2024, 25, 728. https://doi.org/10.3390/ijms25020728
Rozpędek-Kamińska W, Galita G, Saramowicz K, Granek Z, Barczuk J, Siwecka N, Pytel D, Majsterek I. Evaluation of the LDN-0060609 PERK Inhibitor as a Selective Treatment for Primary Open-Angle Glaucoma: An In Vitro Study on Human Retinal Astrocytes. International Journal of Molecular Sciences. 2024; 25(2):728. https://doi.org/10.3390/ijms25020728
Chicago/Turabian StyleRozpędek-Kamińska, Wioletta, Grzegorz Galita, Kamil Saramowicz, Zuzanna Granek, Julia Barczuk, Natalia Siwecka, Dariusz Pytel, and Ireneusz Majsterek. 2024. "Evaluation of the LDN-0060609 PERK Inhibitor as a Selective Treatment for Primary Open-Angle Glaucoma: An In Vitro Study on Human Retinal Astrocytes" International Journal of Molecular Sciences 25, no. 2: 728. https://doi.org/10.3390/ijms25020728
APA StyleRozpędek-Kamińska, W., Galita, G., Saramowicz, K., Granek, Z., Barczuk, J., Siwecka, N., Pytel, D., & Majsterek, I. (2024). Evaluation of the LDN-0060609 PERK Inhibitor as a Selective Treatment for Primary Open-Angle Glaucoma: An In Vitro Study on Human Retinal Astrocytes. International Journal of Molecular Sciences, 25(2), 728. https://doi.org/10.3390/ijms25020728