
Effect of the Flavonoid Rutin on the Modulation of the Myenteric Plexuses in an Experimental Model of Parkinson’s Disease

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Rutin Treatment Reduced the Number of Contralateral Rotations Induced by 6-OHDA
2.2. Rutin Treatment Improves the Motor Capacity of Animals
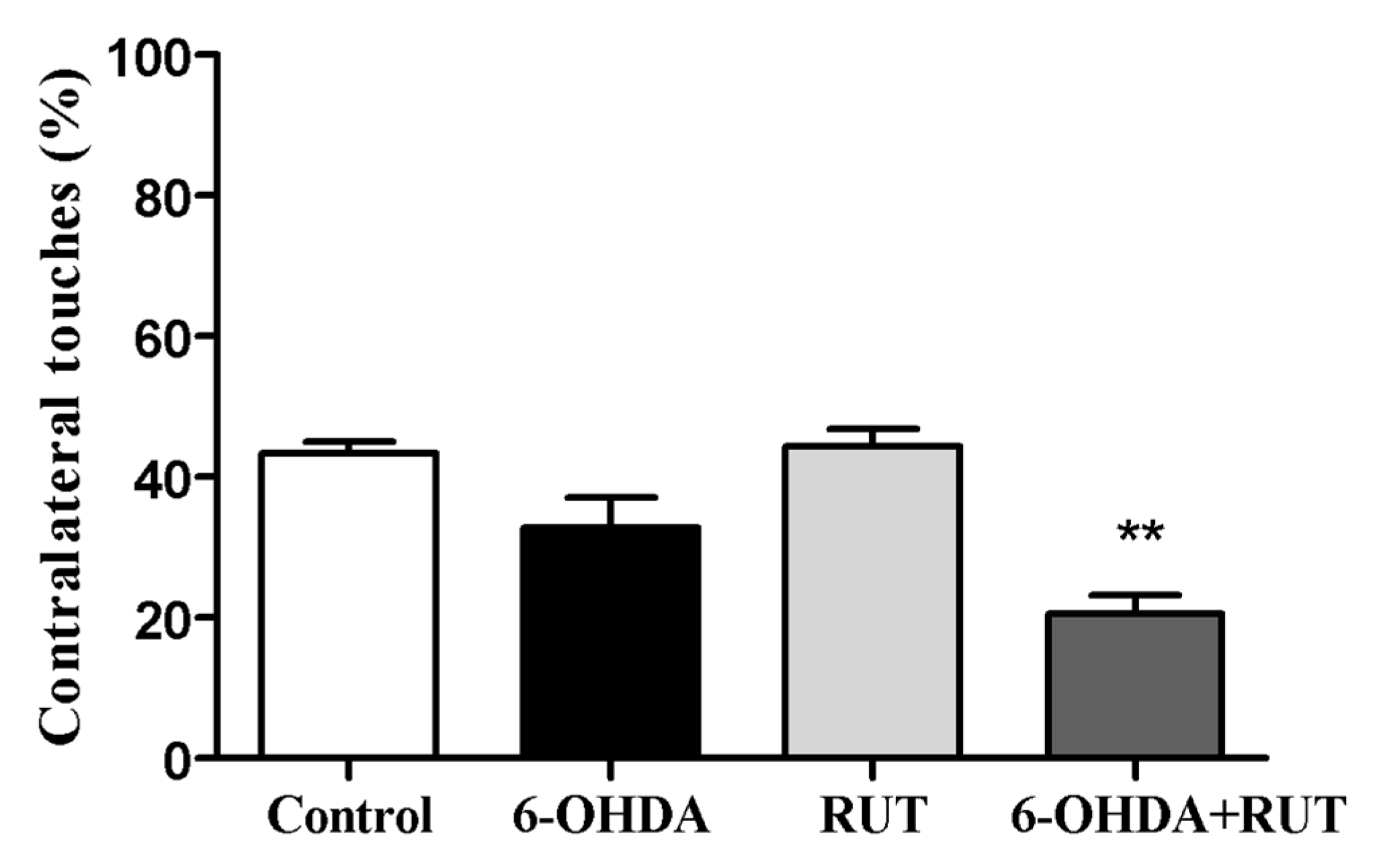
2.3. Rutin Reduces 6-OHDA-Induced Contralateral Deviation
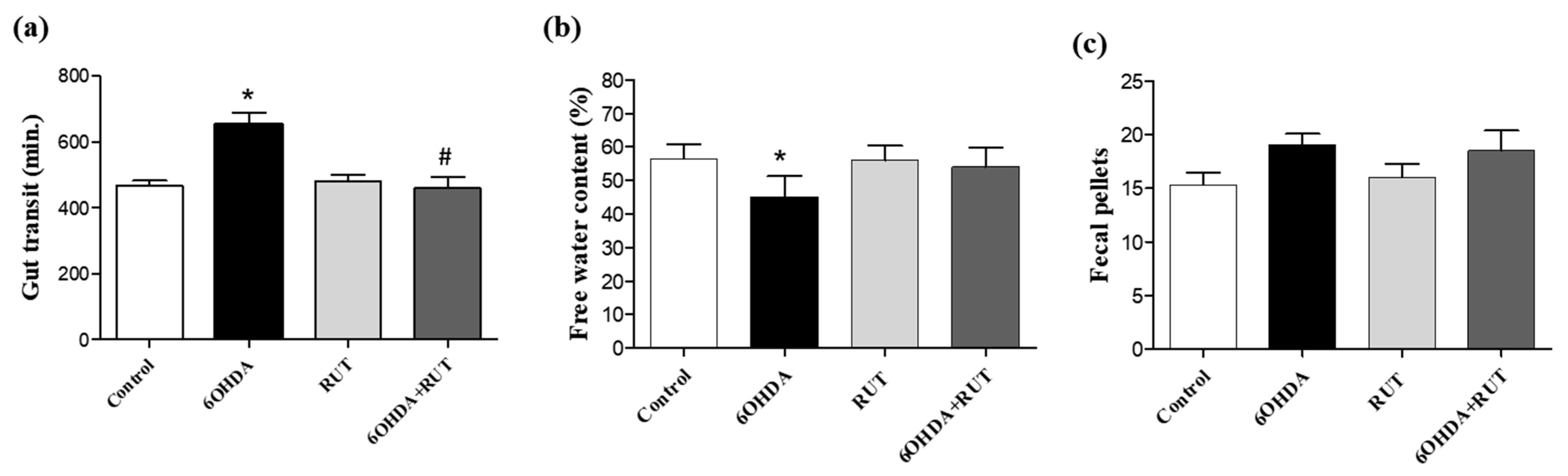
2.4. Rutin Improves Gastrointestinal Transit and Increases Fecal Production in Parkinsonian Animals
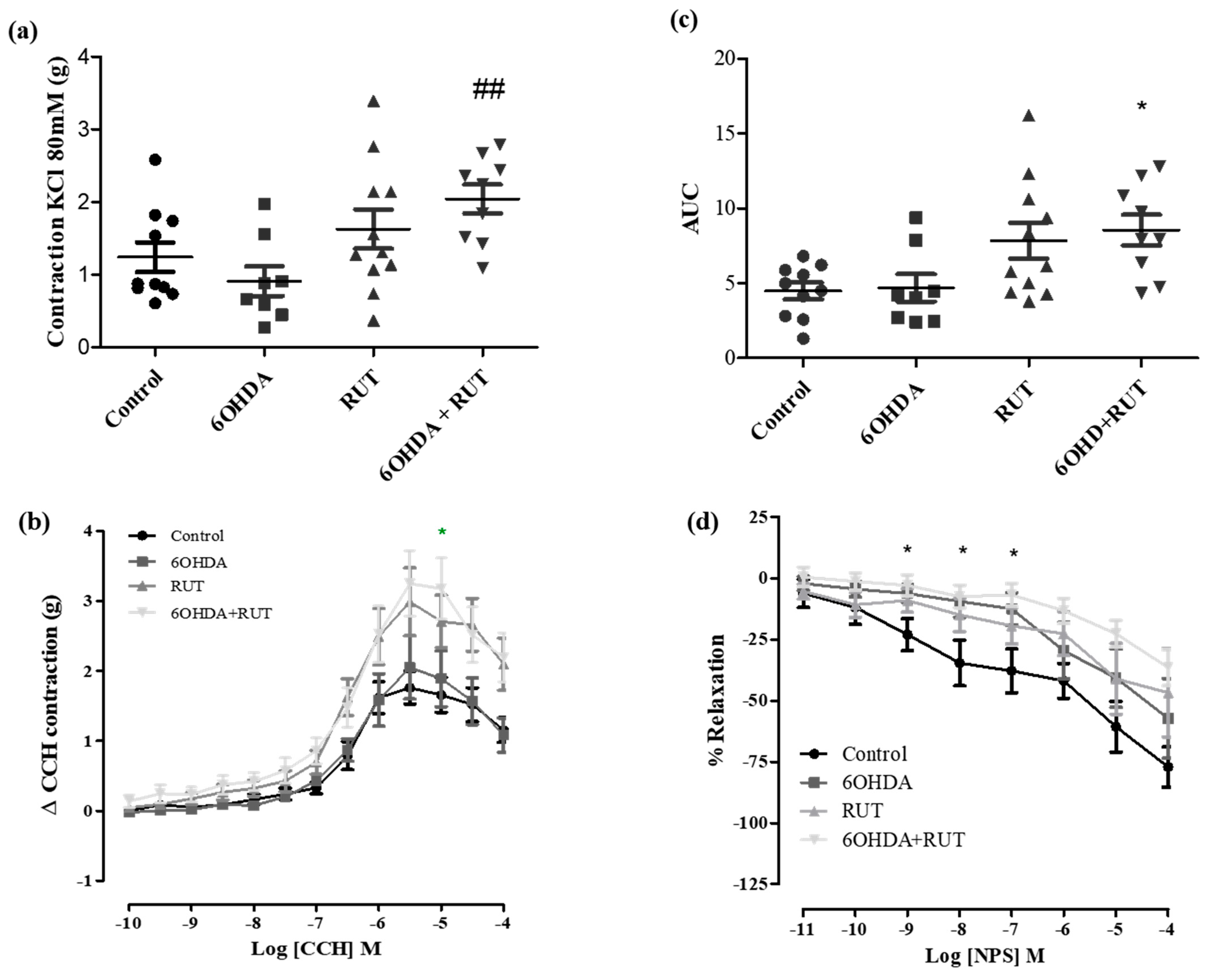
2.5. Rutin Improves Reactivity to Muscarinic Receptors and Hinders Nitric Oxide Signaling in the Ileal Segments from Parkinsonian Animals
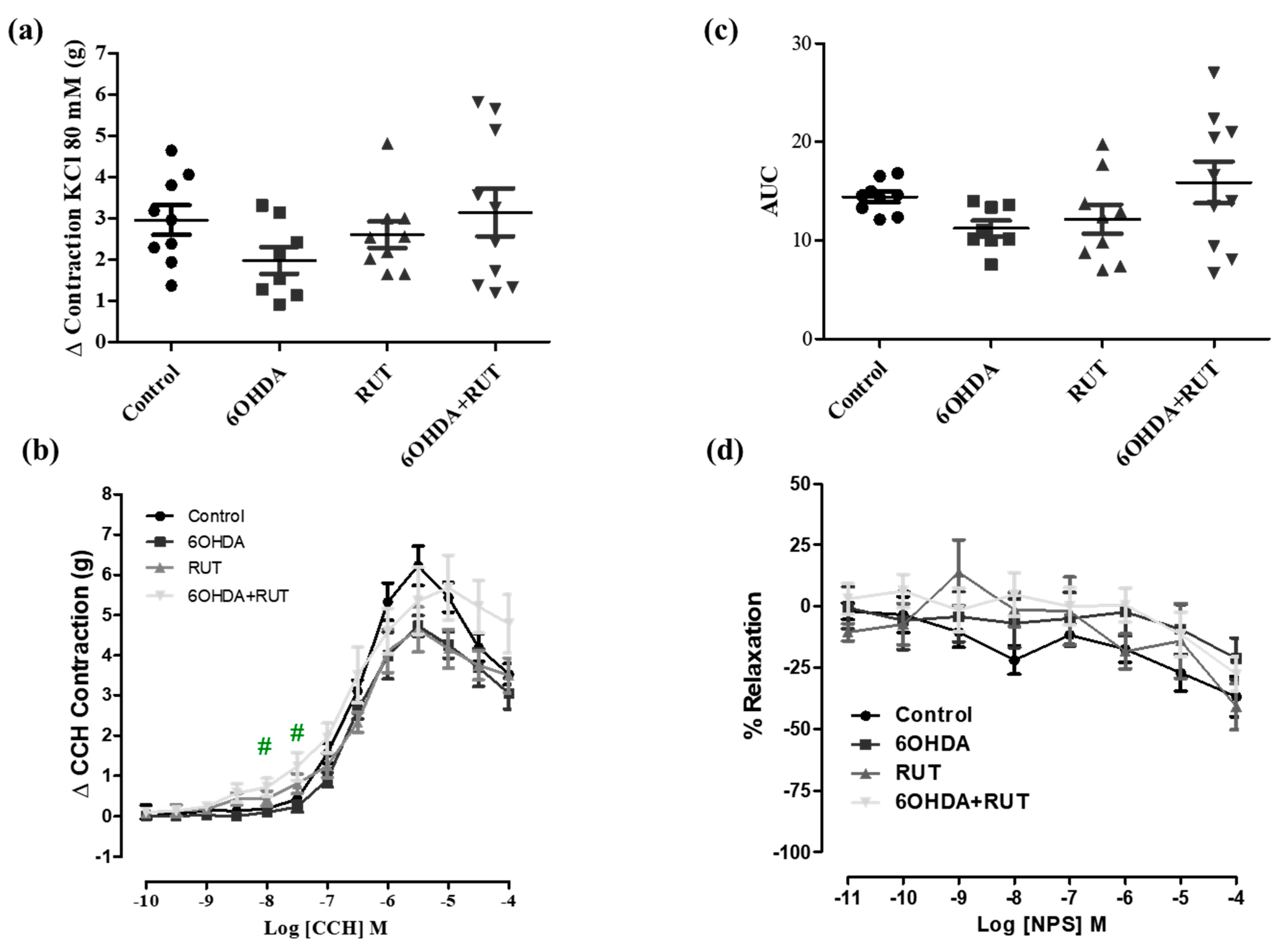
2.6. Treatment with Rutin Increases the Contractility of the Colonic Longitudinal Muscle from Parkinsonian Animals
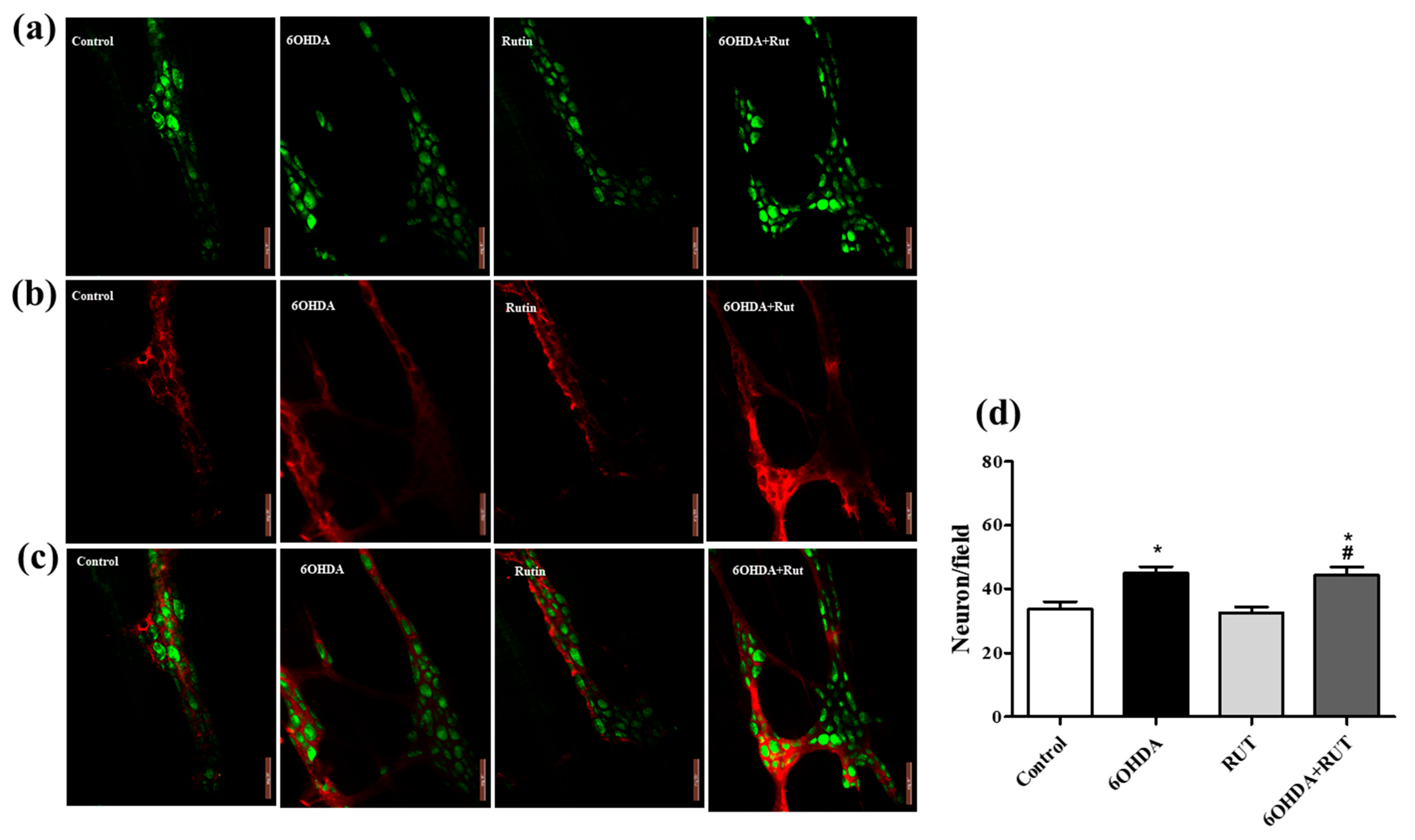
2.7. Rutin Does Not Interfere in the Population of Glial Cells and Enteric Neurons of 6-OHDA-Injured Animals
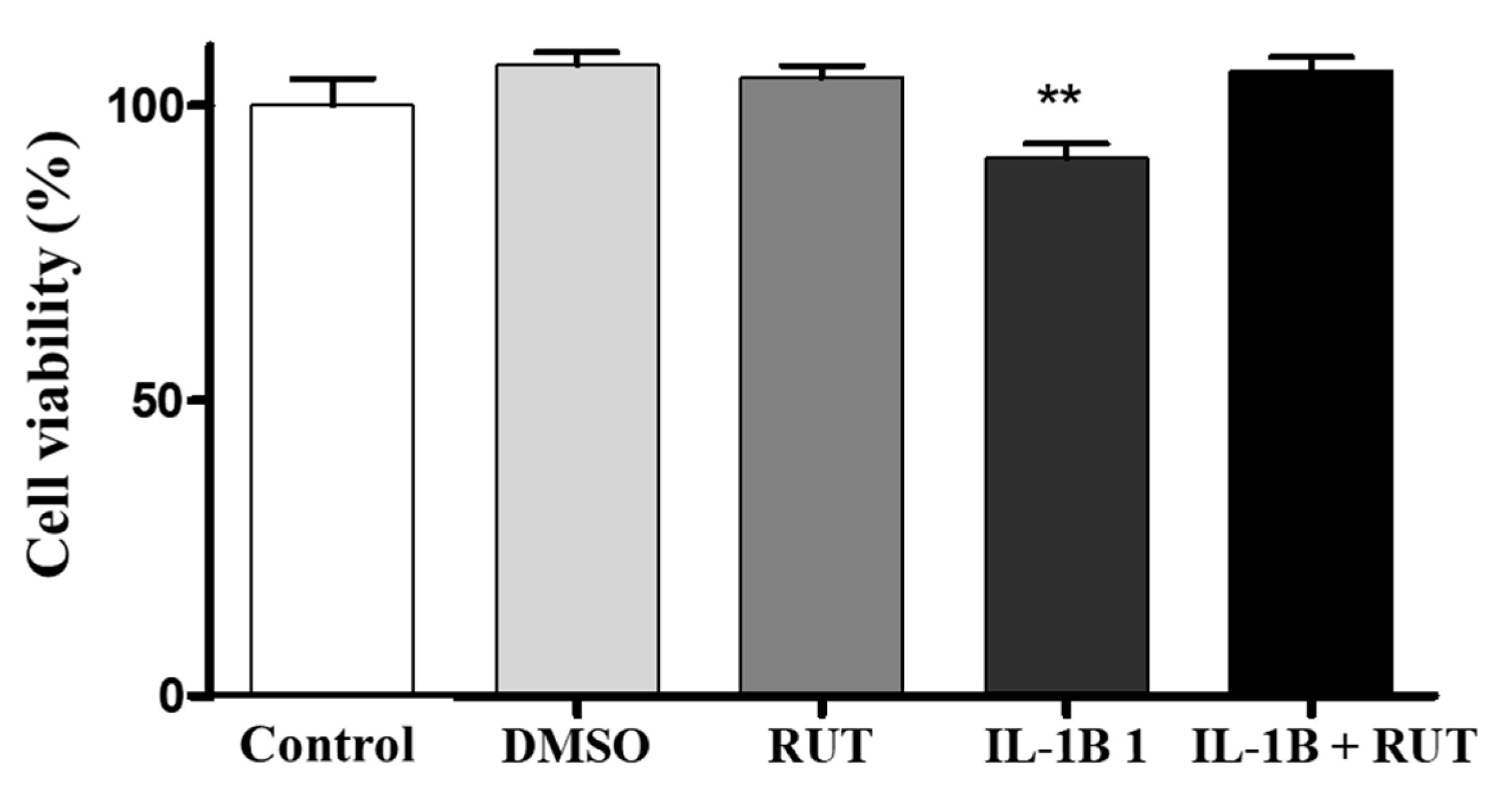
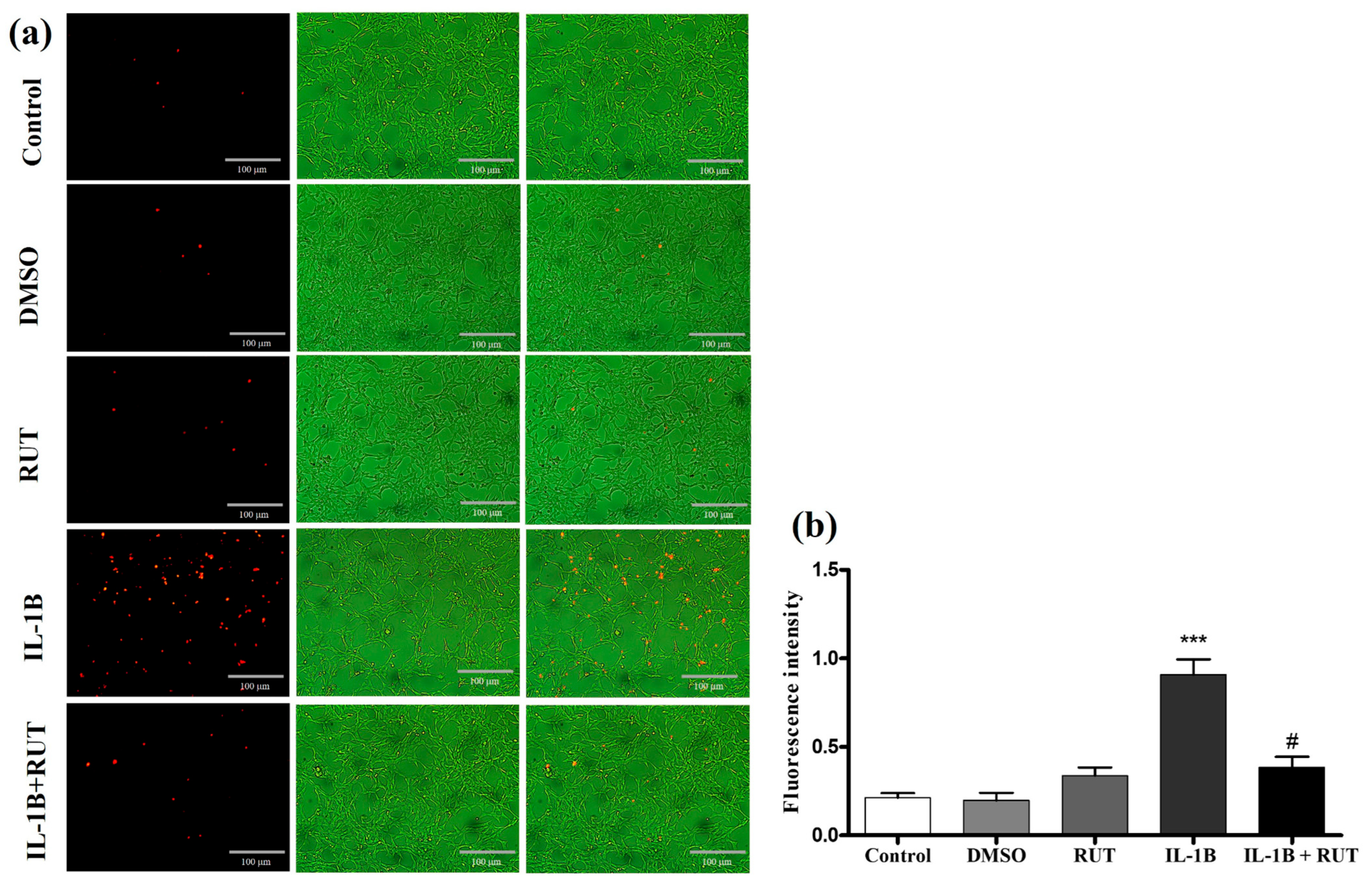
2.8. Treatment with Rutin Protects from Damage Caused by IL-1β in Enteroglial Cells (EGCs)
3. Discussion
4. Materials and Methods
4.1. Animals and Parkinson’s Disease Model
4.2. Open-Field Test
4.3. Cylinder Test
4.4. Apomorphine-Induced Rotation Behavior
4.5. Whole Gut Transit Test and Fecal Output
4.6. Study of Intestinal Contractility
4.7. Immunohistochemical Analysis
4.8. Enteroglial Cell Culture (EGCs)
4.9. Evaluation of Cell Viability in EGC Cultures
4.10. Propidium Iodide (PI) Incorporation Test in EGC Cultures
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.-J. The Enteric Nervous System and Gastrointestinal Innervation: Integrated local and central control. In Experimental Medicine and Biology; Springer: New York, NY, USA, 2014; pp. 39–71. [Google Scholar] [CrossRef]
- Rao, M.; Rastelli, D.; Dong, L.; Chiu, S.; Setlik, W.; Gershon, M.D.; Corfas, G. Enteric Glia Regulate Gastrointestinal Motility but Are Not Required for Maintenance of the Epithelium in Mice. Gastroenterology 2017, 153, 1068–1081.e7. [Google Scholar] [CrossRef]
- Gulbransen, B.D.; Sharkey, K.A. Novel functional roles for enteric glia in the gastrointestinal tract. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Spencer, N.J.; Hu, H. Enteric nervous system: Sensory transduction, neural circuits and gastrointestinal motility. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 338–351. [Google Scholar] [CrossRef]
- Gershon, M.D.; Rothman, T.P. Enteric glia. Glia 1991, 4, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.; Berghe, P.V. The enteric nervous system in PD: Gateway, bystander victim, or source of solutions. Cell Tissue Res. 2018, 373, 313–326. [Google Scholar] [CrossRef]
- Westfall, S.; Lomis, N.; Kahouli, I.; Dia, S.Y.; Singh, S.P.; Prakash, S. Microbiome, probiotics and neurodegenerative diseases: Deciphering the gut brain axis. Cell. Mol. Life Sci. 2017, 74, 3769–3787. [Google Scholar] [CrossRef]
- Herrero, M.T.; Morelli, M. Multiple mechanisms of neurodegeneration and progression. Prog. Neurobiol. 2017, 155, 1. [Google Scholar] [CrossRef]
- Schapira, A.H.V.; Chaudhuri, K.R.; Jenner, P. Non-motor features of Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 435–450. [Google Scholar] [CrossRef]
- Schrag, A.; Horsfall, L.; Walters, K.; Noyce, A.; Petersen, I. Prediagnostic presentations of Parkinson’s disease in primary care: A case-control study. Lancet Neurol. 2015, 14, 57–64. [Google Scholar] [CrossRef]
- Filograna, R.; Beltramini, M.; Bubacco, L.; Bisaglia, M. Anti-Oxidants in Parkinson’s Disease Therapy: A critical point of view. Curr. Neuropharmacol. 2016, 14, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative Stress in the Aging Substantia Nigra and the Etiology of Parkinson’s Disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef]
- De Araújo, F.M.; Cuenca-Bermejo, L.; Fernández-Villalba, E.; Costa, S.L.; Silva, V.D.A.; Herrero, M.T. Role of Microgliosis and NLRP3 Inflammasome in Parkinson’s Disease Pathogenesis and Therapy. Cell. Mol. Neurobiol. 2021, 42, 1283–1300. [Google Scholar] [CrossRef] [PubMed]
- Kujawska, M.; Jodynis-Liebert, J. Polyphenols in Parkinson’s Disease: A systematic review of in vivo studies. Nutrients 2018, 10, 642. [Google Scholar] [CrossRef]
- De Araújo, F.M.; Ferreira, R.S.; Souza, C.S.; Dos Santos, C.C.; Rodrigues, T.L.R.S.; Silva, J.H.C.E.; Gasparotto, J.; Gelain, D.P.; El-Bachá, R.S.; Costa, M.D.F.D.; et al. Aminochrome decreases NGF, GDNF and induces neuroinflammation in organotypic midbrain slice cultures. Neurotoxicology 2018, 66, 98–106. [Google Scholar] [CrossRef]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115. [Google Scholar] [CrossRef]
- Silva, A.R.; Pinheiro, A.M.; Souza, C.S.; Freitas, S.R.V.-B.; Vasconcellos, V.; Freire, S.M.; Velozo, E.S.; Tardy, M.; El-Bachá, R.S.; Costa, M.F.D.; et al. The flavonoid rutin induces astrocyte and microglia activation and regulates TNF-alpha and NO release in primary glial cell cultures. Cell Biol. Toxicol. 2007, 24, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Raza, S.S.; Javed, H.; Ahmad, A.; Khan, A.; Islam, F.; Safhi, M.M.; Islam, F. Rutin Protects Dopaminergic Neurons from Oxidative Stress in an Animal Model of Parkinson’s Disease. Neurotox. Res. 2011, 22, 1–15. [Google Scholar] [CrossRef]
- Ren, R.; Shi, C.; Cao, J.; Sun, Y.; Zhao, X.; Guo, Y.; Wang, C.; Lei, H.; Jiang, H.; Ablat, N.; et al. Neuroprotective Effects of A Standardized Flavonoid Extract of Safflower Against Neurotoxin-Induced Cellular and Animal Models of Parkinson’s Disease. Sci. Rep. 2016, 6, 22135. [Google Scholar] [CrossRef]
- Berridge, K.C. Comparative Fine Structure of Action: Rules of Form and Sequence in the Grooming Patterns of Six Rodent Species. Behaviour 1990, 113, 21–56. [Google Scholar] [CrossRef]
- Richmond, G.; Sachs, B.D. Grooming in Norway Rats: The development and adult expression of a complex motor pattern. Behaviour 1980, 75, 82–95. [Google Scholar] [CrossRef]
- Young, R.K.; Thiessen, D.D. Washing, drying, and anointing in adult humans (Homo sapiens): Commonalities with grooming sequences in rodents. J. Comp. Psychol. 1991, 105, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C. Substantia nigra 6-OHDA lesions mimic striatopallidal disruption of syntactic grooming chains: A neural systems analysis of sequence control. Psychobiology 1989, 17, 377–385. [Google Scholar] [CrossRef]
- Tozzi, A.; Tantucci, M.; Marchi, S.; Mazzocchetti, P.; Morari, M.; Pinton, P.; Mancini, A.; Calabresi, P. Dopamine D2 receptor-mediated neuroprotection in a G2019S Lrrk2 genetic model of Parkinson’s disease. Cell Death Dis. 2018, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Naidu, P.S.; Singh, A.; Kulkarni, S.K. D2-dopamine receptor and alpha2-adrenoreceptor-mediated analgesic response of quercetin. Indian J. Exp. Biol. 2003, 41, 1400–1404. [Google Scholar] [PubMed]
- Lebouvier, T.; Chaumette, T.; Paillusson, S.; Duyckaerts, C.; Varannes, S.B.D.; Neunlist, M.; Derkinderen, P. The second brain and Parkinson’s disease. Eur. J. Neurosci. 2009, 30, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.S.; Teles-Souza, J.; Souza, C.D.S.; Pereira, É.P.L.; De Araðjo, F.M.; Da Silva, A.B.; Silva, J.H.C.E.; Nonose, Y.; Nðñez-Figueredo, Y.; de Assis, A.M.; et al. Rutin improves glutamate uptake and inhibits glutamate excitotoxicity in rat brain slices. Mol. Biol. Rep. 2021, 48, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Chinchilla, T.; Quiroga-Varela, A.; Molinet-Dronda, F.; Belloso-Iguerategui, A.; Merino-Galan, L.; Jimenez-Urbieta, H.; Gago, B.; Rodriguez-Oroz, M.C. [18F]-DPA-714 PET as a specific in vivo marker of early microglial activation in a rat model of progressive dopaminergic degeneration. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 2602–2612. [Google Scholar] [CrossRef]
- Gil-Cardoso, K.; Ginés, I.; Pinent, M.; Ardévol, A.; Blay, M.; Terra, X. Effects of flavonoids on intestinal inflammation, barrier integrity and changes in gut microbiota during diet-induced obesity. Nutr. Res. Rev. 2016, 29, 234–248. [Google Scholar] [CrossRef]
- Dawson, T.M.; Dawson, V.L. Molecular Pathways of Neurodegeneration in Parkinson’s Disease. Science 2003, 302, 819–822. [Google Scholar] [CrossRef]
- Das, N.R.; Sharma, S.S. Cognitive Impairment Associated with Parkinson’s Disease: Role of mitochondria. Curr. Neuropharmacol. 2016, 14, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Devos, D.; Lebouvier, T.; Lardeux, B.; Biraud, M.; Rouaud, T.; Pouclet, H.; Coron, E.; Des Varannes, S.B.; Naveilhan, P.; Nguyen, J.-M.; et al. Colonic inflammation in Parkinson’s disease. Neurobiol. Dis. 2013, 50, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Pallares, J.; Parga, J.A.; Muñoz, A.; Rey, P.; Guerra, M.J.; Labandeira-Garcia, J.L. Mechanism of 6-hydroxydopamine neurotoxicity: The role of nadph oxidase and microglial activation in 6-hydroxydopamine-induced degeneration of dopaminergic neurons. J. Neurochem. 2007, 103, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Murillo, M.D.P.; Johansson, E.; Bryntesson, V.; Aronsson, P.; Tobin, G.; Winder, M.; Carlsson, T. 6-OHDA-Induced Changes in Colonic Segment Contractility in the Rat Model of Parkinson’s Disease. Gastroenterol. Res. Pract. 2023, 2023, 9090524. [Google Scholar] [CrossRef] [PubMed]
- Benvenuti, L.; D’antongiovanni, V.; Pellegrini, C.; Antonioli, L.; Bernardini, N.; Blandizzi, C.; Fornai, M. Enteric Glia at the Crossroads between Intestinal Immune System and Epithelial Barrier: Implications for parkinson disease. Int. J. Mol. Sci. 2020, 21, 9199. [Google Scholar] [CrossRef] [PubMed]
- Rivera, L.R.; Poole, D.P.; Thacker, M.; Furness, J.B. The involvement of nitric oxide synthase neurons in enteric neuropathies. Neurogastroenterol. Motil. 2011, 23, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Groneberg, D.; Voussen, B.; Friebe, A. Integrative Control of Gastrointestinal Motility by Nitric Oxide. Curr. Med. Chem. 2016, 23, 2715–2735. [Google Scholar] [CrossRef] [PubMed]
- Wiley, J.W. The many faces of nitric oxide: Cytotoxic, cytoprotective or both. Neurogastroenterol. Motil. 2007, 19, 541–544. [Google Scholar] [CrossRef]
- Martins-Perles, J.V.C.; Bossolani, G.D.P.; Zignani, I.; de Souza, S.R.G.; Frez, F.C.V.; de Souza Melo, C.G.; Barili, E.; Neto, F.P.d.S.; Guarnier, F.A.; Armani, A.L.C.; et al. Quercetin increases bioavailability of nitric oxide in the jejunum of euglycemic and diabetic rats and induces neuronal plasticity in the myenteric plexus. Auton. Neurosci. 2020, 227, 102675. [Google Scholar] [CrossRef]
- Clairembault, T.; Kamphuis, W.; Leclair-Visonneau, L.; Rolli-Derkinderen, M.; Coron, E.; Neunlist, M.; Hol, E.M.; Derkinderen, P. Enteric GFAP expression and phosphorylation in Parkinson’s disease. J. Neurochem. 2014, 130, 805–815. [Google Scholar] [CrossRef]
- Christmann, A.; Gries, M.; Scholz, P.; Stahr, P.L.; Law, J.K.Y.; Schulte, S.; Martin, M.; Lilischkis, R.; Ingebrandt, S.; Keck, C.M.; et al. The antioxidant Rutin counteracts the pathological impact of α-synuclein on the enteric nervous system in vitro. Biol. Chem. 2021, 403, 103–122. [Google Scholar] [CrossRef] [PubMed]
- De Araújo, F.M.; Frota, A.F.; de Jesus, L.B.; Cuenca-Bermejo, L.; Ferreira, K.M.S.; Santos, C.C.; Soares, E.N.; Souza, J.T.; Sanches, F.S.; Costa, A.C.S.; et al. Protective Effects of Flavonoid Rutin Against Aminochrome Neurotoxicity. Neurotox. Res. 2023, 41, 224–241. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Elsevier: San Diego, CA, USA; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Broadhurst, P.L. Application of Biometrical Genetics to Behaviour in Rats. Nature 1959, 184, 1517–1518. [Google Scholar] [CrossRef]
- Archer, J. Tests for emotionality in rats and mice: A review. Anim. Behav. 1973, 21, 205–235. [Google Scholar] [CrossRef]
- Schallert, T.; Fleming, S.M.; Leasure, J.L.; Tillerson, J.L.; Bland, S.T. CNS plasticity and assessment of forelimb sensorimotor outcome in unilateral rat models of stroke, cortical ablation, parkinsonism and spinal cord injury. Neuropharmacology 2000, 39, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Joo, W.S.; Jin, B.K.; Cho, Y.H.; Baik, H.H.; Park, C.W. Melatonin protects 6-OHDA-induced neuronal death of nigrostriatal dopaminergic system. Neuroreport 1998, 9, 2387–2390. [Google Scholar] [CrossRef]
- Devries, M.P.; Vessalo, M.; Galligan, J.J. A deleção das subunidades dos receptores P2X2 e P2X3 não altera a motilidade do cólon de camundongos. Frente Neurosci. 2010, 19, 22. [Google Scholar] [CrossRef]
- Farmer, S.G.; Laniyonu, A.A. Effects of p-chlorophenylalanine on the sensitivity of rat intestine to agonists and on intestinal 5-hydroxytryptamine levels during Nippostrongylus brasiliensis infection. Br. J. Pharmacol. 1984, 82, 883–889. [Google Scholar] [CrossRef]
- Frias, B.; Phillips, A.A.; Squair, J.W.; Lee, A.H.X.; Laher, I.; Krassioukov, A.V. Reduced colonic smooth muscle cholinergic responsiveness is associated with impaired bowel motility after chronic experimental high-level spinal cord injury. Auton. Neurosci. 2019, 216, 33–38. [Google Scholar] [CrossRef]
- Lin, Z.; Gao, N.; Hu, H.-Z.; Liu, S.; Gao, C.; Kim, G.; Ren, J.; Xia, Y.; Peck, O.C.; Wood, J.D. Immunoreactivity of Hu proteins facilitates identification of myenteric neurones in guinea-pig small intestine. Neurogastroenterol. Motil. 2002, 14, 197–204. [Google Scholar] [CrossRef]
- Schneider, L.C.L.; Nascimento, J.C.P.D.; Trevizan, A.R.; Góis, M.B.; Borges, S.C.; Beraldi, E.J.; Garcia, J.L.; Sant’Ana, D.M.G.; Buttow, N.C. Toxoplasma gondii promotes changes in VIPergic submucosal neurons, mucosal intraepithelial lymphocytes, and goblet cells during acute infection in the ileum of rats. Neurogastroenterol. Motil. 2018, 30, e13264. [Google Scholar] [CrossRef] [PubMed]
- Trevizan, A.R.; Schneider, L.C.L.; Araújo, E.J.d.A.; Garcia, J.L.; Buttow, N.C.; Nogueira-Melo, G.d.A.; Sant’Ana, D.d.M.G. Acute Toxoplasma gondii infection alters the number of neurons and the proportion of enteric glial cells in the duodenum in Wistar rats. Neurogastroenterol. Motil. 2019, 31, e13523. [Google Scholar] [CrossRef] [PubMed]
- Hermes-Uliana, C.; Panizzon, C.P.D.N.B.; Trevizan, A.R.; Sehaber, C.C.; Ramalho, F.V.; Martins, H.A.; Zanoni, J.N. Is l-Glutathione More Effective Than l-Glutamine in Preventing Enteric Diabetic Neuropathy? Dig. Dis. Sci. 2013, 59, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Ferri, G.-L.; Probert, L.; Cocchia, D.; Michetti, F.; Marangos, P.J.; Polak, J.M. Evidence for the presence of S-100 protein in the glial component of the human enteric nervous system. Nature 1982, 297, 409–410. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Jesus, L.B.; Frota, A.F.; de Araújo, F.M.; Jesus, R.L.C.d.; Costa, M.d.F.D.; de Vasconcelos, D.F.S.A.; Gois, M.B.; Baccan, G.C.; da Silva, V.D.A.; Costa, S.L. Effect of the Flavonoid Rutin on the Modulation of the Myenteric Plexuses in an Experimental Model of Parkinson’s Disease. Int. J. Mol. Sci. 2024, 25, 1037. https://doi.org/10.3390/ijms25021037
de Jesus LB, Frota AF, de Araújo FM, Jesus RLCd, Costa MdFD, de Vasconcelos DFSA, Gois MB, Baccan GC, da Silva VDA, Costa SL. Effect of the Flavonoid Rutin on the Modulation of the Myenteric Plexuses in an Experimental Model of Parkinson’s Disease. International Journal of Molecular Sciences. 2024; 25(2):1037. https://doi.org/10.3390/ijms25021037
Chicago/Turabian Stylede Jesus, Livia Bacelar, Annyta Fernandes Frota, Fillipe Mendes de Araújo, Rafael Leonne Cruz de Jesus, Maria de Fátima Dias Costa, Darizy Flavia Silva Amorim de Vasconcelos, Marcelo Biondaro Gois, Gyselle Chrystina Baccan, Victor Diogenes Amaral da Silva, and Silvia Lima Costa. 2024. "Effect of the Flavonoid Rutin on the Modulation of the Myenteric Plexuses in an Experimental Model of Parkinson’s Disease" International Journal of Molecular Sciences 25, no. 2: 1037. https://doi.org/10.3390/ijms25021037
APA Stylede Jesus, L. B., Frota, A. F., de Araújo, F. M., Jesus, R. L. C. d., Costa, M. d. F. D., de Vasconcelos, D. F. S. A., Gois, M. B., Baccan, G. C., da Silva, V. D. A., & Costa, S. L. (2024). Effect of the Flavonoid Rutin on the Modulation of the Myenteric Plexuses in an Experimental Model of Parkinson’s Disease. International Journal of Molecular Sciences, 25(2), 1037. https://doi.org/10.3390/ijms25021037