
Network Pharmacology and Transcriptomics to Explore the Pharmacological Mechanisms of 20(S)-Protopanaxatriol in the Treatment of Depression

 ,
,
Abstract
1. Introduction
2. Result
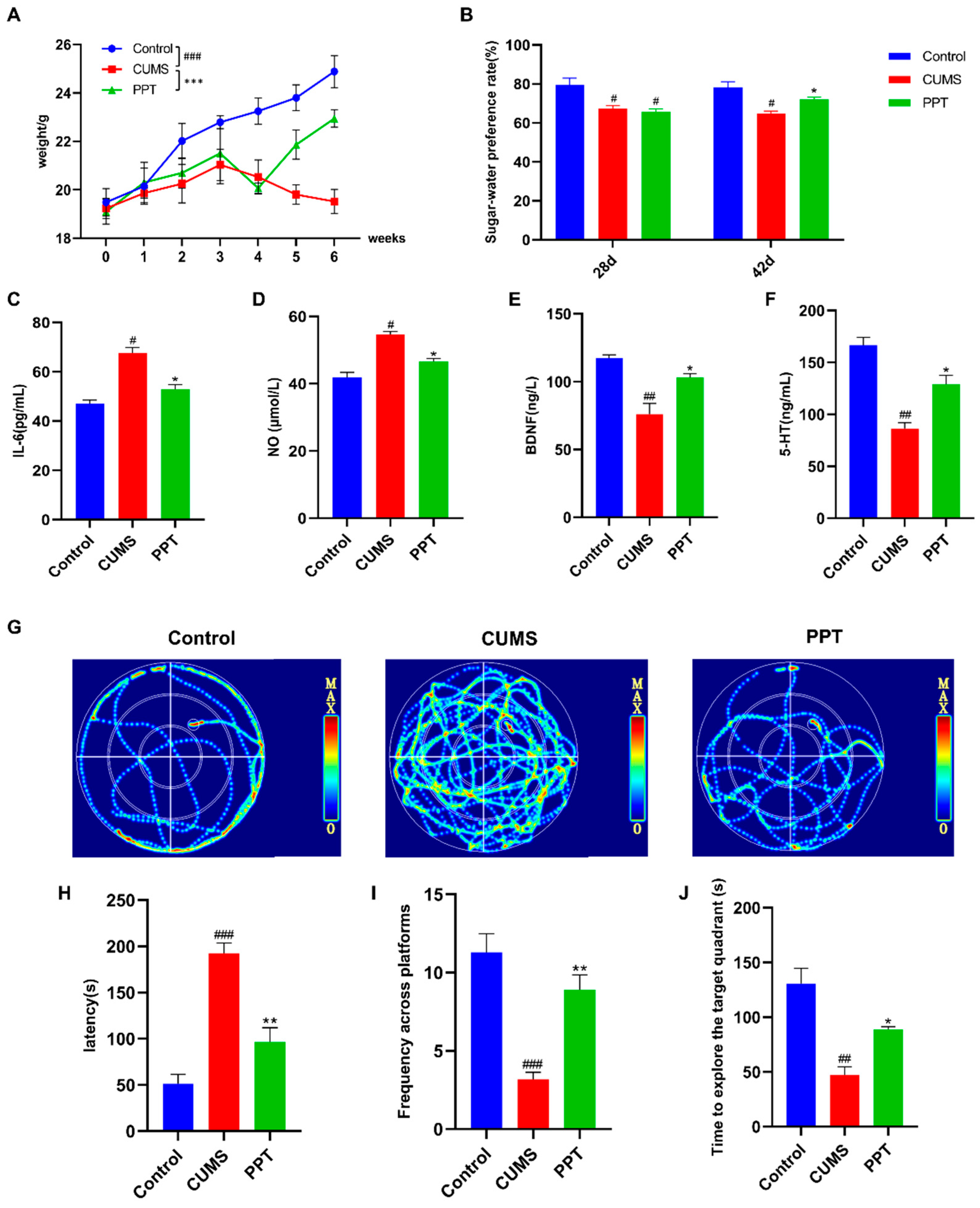
2.1. Weight Detection
2.2. Sucrose Preference Test (SPT)
2.3. Serum Biochemical Indicator Tests
2.4. Morris Water Maze (MWM)
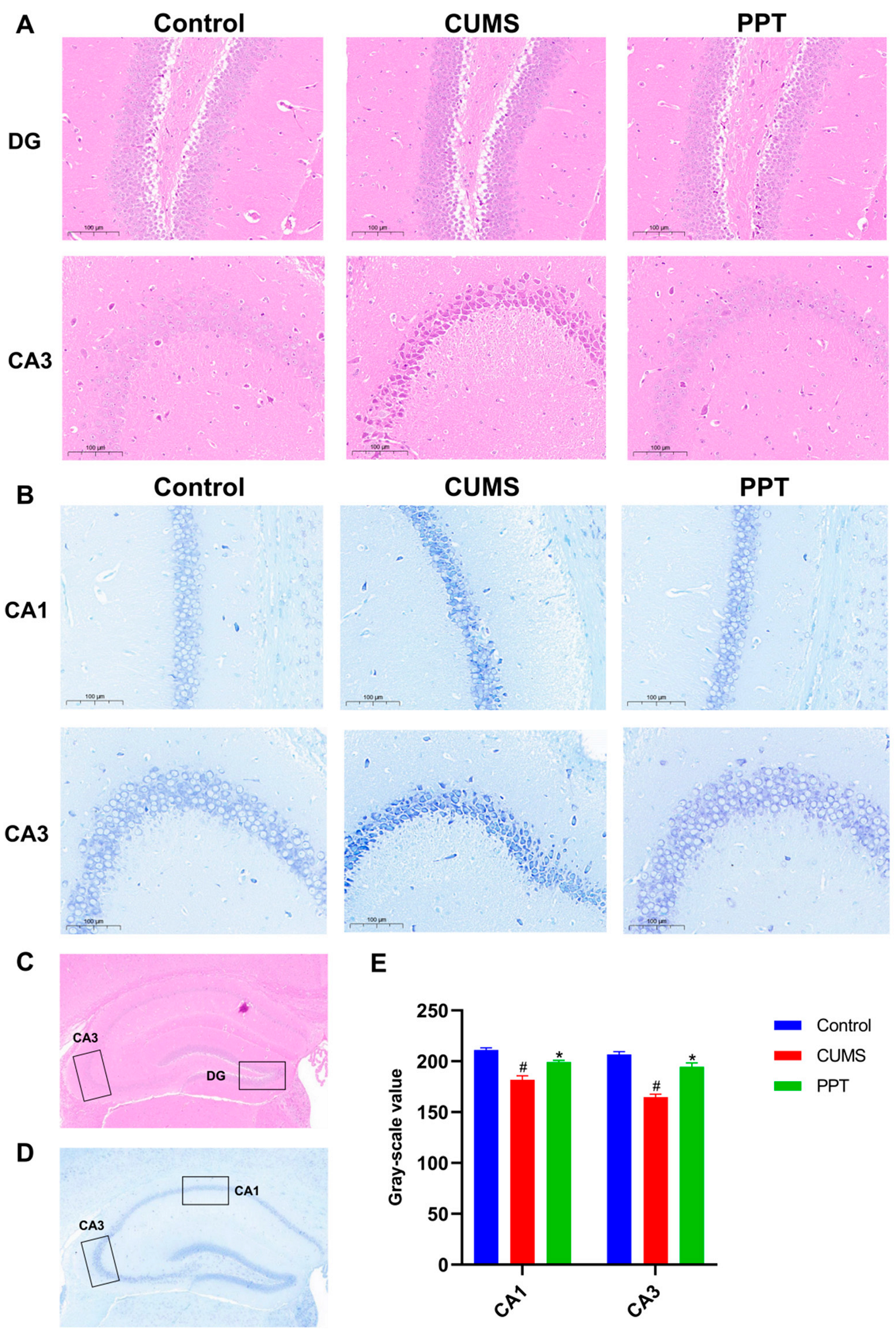
2.5. PPT Can Improve the Brain Histopathological Injury Induced by CUMS
2.6. Target Collection and PPI Network Analysis of PPT for Depression
2.7. GO and KEGG Pathway Analysis
2.8. Transcriptomics Analysis
2.9. Integrated Analysis of Transcriptome Sequencing and Network Pharmacology
2.10. Molecular Docking
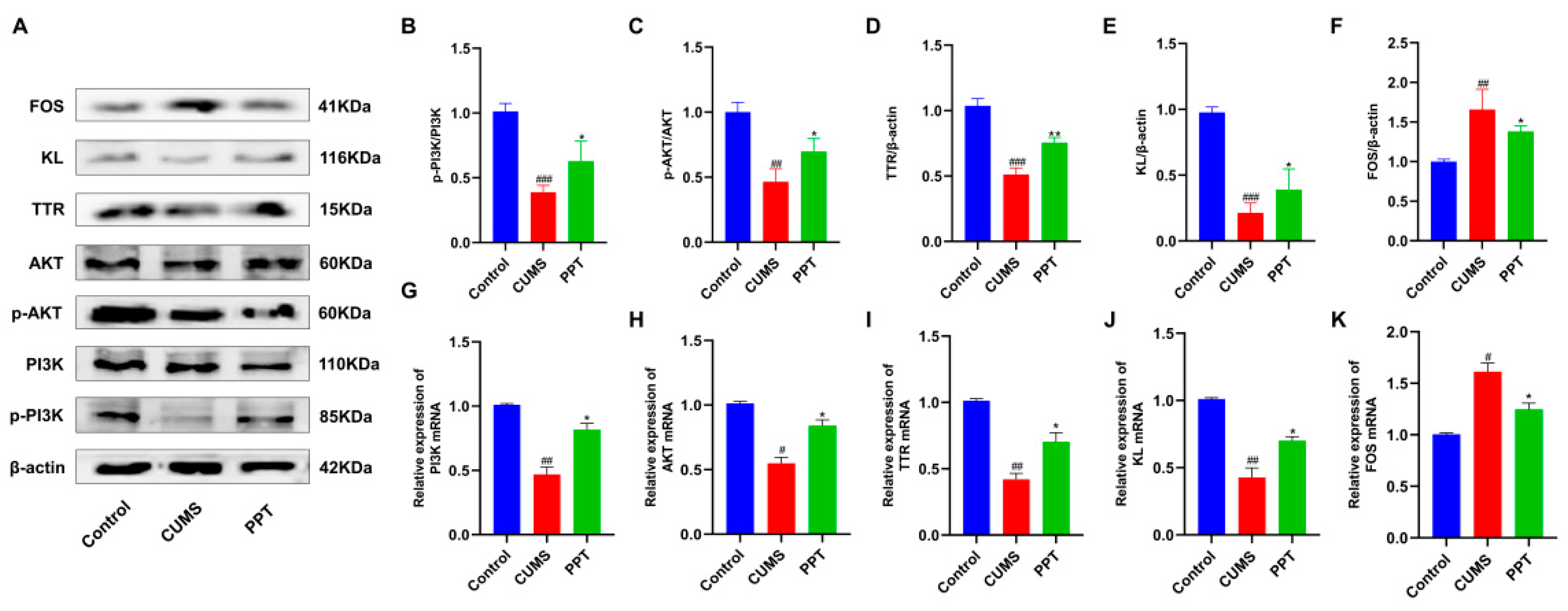
2.11. Effects of PPT on PI3K-AKT Signaling Pathway and TTR, KL, and FOS Protein Expression in Depressed Mice
2.12. qRT-PCR to Verify the Expression Level of Target Genes
3. Materials and Methods
3.1. Reagents and Materials
3.2. Animal Models and Therapy
3.3. Weight Detection
3.4. Sucrose Preference Test (SPT)
3.5. Serum Biochemical Indicator Tests
3.6. Morris Water Maze (MWM)
3.6.1. Localization Navigation Experiment
3.6.2. Spatial Search Experiment
3.7. Histopathological Examination
3.8. Target Collection and PPI Network Analysis of PPT for Depression
3.9. GO and KEGG Pathway Analysis
3.10. RNA Sequencing (RNA-Seq) and Data Analysis
3.11. Molecular Docking
3.12. Western Blotting
3.13. qRT-PCR to Verify Gene Expression Levels
3.14. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, M.; Fu, X.; Xie, W.; Guo, W.; Li, B.; Cui, R.; Yang, W. Effect of Early Life Stress on the Epigenetic Profiles in Depression. Front. Cell Dev. Biol. 2020, 8, 867. [Google Scholar] [CrossRef] [PubMed]
- W.H. Organization. Depressive Disorder (Depression). Available online: https://www.who.int/news-room/fact-sheets/detail/depression (accessed on 31 March 2023).
- Varghese, S.P.; Florentin, O.D.; Koola, M.M. Role of Spirituality in the Management of Major Depression and Stress-Related Disorders. Chronic Stress 2021, 5, 2470547020971232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Qin, X.-M.; Du, G.-H.; Gao, X.-X. Progress in study of antidepressants metabolism in vivo. Acta Pharm. Sin. 2017, 12, 1791–1800. [Google Scholar]
- Ding, Y.; Botchway, B.O.A.; Zhang, Y.; Liu, X. Ginsenosides can target brain-derived neurotrophic factor to improve Parkinson’s disease. Food Funct. 2023, 14, 5537–5550. [Google Scholar] [CrossRef] [PubMed]
- Hyun, S.H.; Bhilare, K.D.; In, G.; Park, C.-K.; Kim, J.-H. Effects of Panax ginseng and ginsenosides on oxidative stress and cardiovascular diseases: Pharmacological and therapeutic roles. J. Ginseng Res. 2022, 46, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Pan, Y.; Liu, Y.; Na, S.; Zhou, H.; Li, L.; Chen, F.; Song, H. A review of anti-tumour effects of ginsenoside in gastrointestinal cancer. J. Pharm. Pharmacol. 2021, 73, 1292–1301. [Google Scholar] [CrossRef]
- Fu, B.; Ma, R.; Liu, F.; Chen, X.; Teng, X.; Yang, P.; Liu, J.; Zhao, D.; Sun, L. Ginsenosides improve reproductive capability of aged female Drosophila through mechanism dependent on ecdysteroid receptor (ECR) and steroid signaling pathway. Front. Endocrinol. 2022, 13, 964069. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Qi, H.; Wang, Y.; Zhang, Z.; Liu, Q.; Guo, C.; Liu, J.; Chen, Z.; Li, J.; Chen, J.; et al. Recent advances in ginsenosides against respiratory diseases: Therapeutic targets and potential mechanisms. Biomed. Pharmacother. 2023, 158, 114096. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Cheng, Y.; Han, X.; Wang, T.; Zhang, H.; Yao, Q.; Chen, F.; Gu, L.; Yang, D.; Chen, L.; et al. 20(S)-Protopanaxadiol Exerts Antidepressive Effects in Chronic Corticosterone-Induced Rodent Animal Models as an Activator of Brain-Type Creatine Kinase. J. Agric. Food Chem. 2024, 72, 10376–10390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-J.; Hu, R.-Y.; Chen, K.-C.; Liu, Y.-B.; Hou, Y.-Y.; Zhang, Y.-Z.; Feng, Z.-M.; Chen, R.-X.; Zheng, Y.-N.; Liu, S.; et al. 20(S)-protopanaxatriol inhibited D-galactose-induced brain aging in mice via promoting mitochondrial autophagy flow. Phytother. Res. 2023, 37, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Nogales, C.; Mamdouh, Z.M.; List, M.; Kiel, C.; Casas, A.I.; Schmidt, H.H.H.W. Network pharmacology: Curing causal mechanisms instead of treating symptoms. Trends Pharmacol. Sci. 2022, 43, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Cui, H.; Liu, G.; Zhao, X.; Li, W.; Piao, G. How Can Synergism of Traditional Medicines Benefit from Network Pharmacology? Molecules 2017, 22, 1135. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Kui, L.; Tang, M.; Li, D.; Wei, K.; Chen, W.; Miao, J.; Dong, Y. High-Throughput Transcriptome Profiling in Drug and Biomarker Discovery. Front. Genet. 2020, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, S.; Shan, C.; Li, X.; Tan, B.; Chen, Q.; Yang, Y.; Yu, H.; Yang, A. Mechanism of protective effect of xuan-bai-cheng-qi decoction on LPS-induced acute lung injury based on an integrated network pharmacology and RNA-sequencing approach. Respir. Res. 2021, 22, 188. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Liang, J.; Guo, Z.; Bai, X.; Zhang, H.; Zhang, N.; Wang, H.; Chen, Q.; Li, W.; Dong, R.; et al. Network pharmacology and transcriptomics to determine Danggui Yifei Decoction mechanism of action for the treatment of chronic lung injury. J. Ethnopharmacol. 2023, 318, 116873. [Google Scholar] [CrossRef]
- Lu, C.; Lv, J.; Dong, L.; Jiang, N.; Wang, Y.; Wang, Q.; Li, Y.; Chen, S.; Fan, B.; Wang, F.; et al. Neuroprotective effects of 20(S)-protopanaxatriol (PPT) on scopolamine-induced cognitive deficits in mice. Phytother. Res. 2018, 32, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, X.; He, S.; Hu, L.; Guo, J.; Huang, X.; Hu, J.; Qi, Y.; Chen, B.; Shang, D.; et al. Regulation of the kynurenine metabolism pathway by Xiaoyao San and the underlying effect in the hippocampus of the depressed rat. J. Ethnopharmacol. 2018, 214, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, N.; Lei, J.; Jing, B.; Li, M.; Tian, H.; Xue, B.; Li, X. Fluoxetine shows neuroprotective effects against LPS-induced neuroinflammation via the Notch signaling pathway. Int. Immunopharmacol. 2022, 113, 109417. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. Swiss Target Prediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-Y.; Zhang, Y.-Q.; Liu, Z.-M.; Chen, T.; Lv, C.-Y.; Tang, S.-H.; Zhang, X.-B.; Zhang, W.; Li, Z.-Y.; Zhou, R.-R.; et al. ETCM: An encyclopaedia of traditional Chinese medicine. Nucleic Acids Res. 2018, 47, D976–D982. [Google Scholar] [CrossRef] [PubMed]
- Fishilevich, S.; Zimmerman, S.; Kohn, A.; Stein, T.I.; Olender, T.; Kolker, E.; Safran, M.; Lancet, D. Genic insights from integrated human proteomics in GeneCards. Database 2016, 2016, baw030. [Google Scholar] [CrossRef] [PubMed]
- Hamosh, A.; Scott, A.F.; Amberger, J.; Bocchini, C.; Valle, D.; McKusick, V.A. Online Mendelian Inheritance in Man (OMIM), a knowledgebase of human genes and genetic disorders. Nucleic Acids Res. 2002, 30, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Saüch, J.; Sanz, F.; Furlong, L.I. The DisGeNET cytoscape app: Exploring and visualizing disease genomics data. Comput. Struct. Biotechnol. J. 2021, 19, 2960–2967. [Google Scholar] [CrossRef] [PubMed]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed]
- Snel, B.; Lehmann, G.; Bork, P.; Huynen, M.A. STRING: A web-server to retrieve and display the repeatedly occurring neighbourhood of a gene. Nucleic Acids Res. 2000, 28, 3442–3444. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, J.; Wang, C.; Zheng, T.; Di, S.; Wang, Y.; Fei, W.; Liang, W.; Wang, L. Ameliorative Effect of Ethanolic Echinacea purpurea against Hyperthyroidism-Induced Oxidative Stress via AMRK and PPAR Signal Pathway Using Transcriptomics and Network Pharmacology Analysis. Int. J. Mol. Sci. 2023, 24, 187. [Google Scholar] [CrossRef]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A free online platform for data visualization and graphing. PLoS ONE 2023, 18, e0294236. [Google Scholar] [CrossRef]
- Berman, H.; Henrick, K.; Nakamura, H. Announcing the worldwide Protein Data Bank. Nat. Struct. Mol. Biol. 2003, 10, 980. [Google Scholar] [CrossRef]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; McVeigh, R.; O‘Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database J. Biol. Databases Curation 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Chen, T.; Cheng, L.; Ma, J.; Yuan, J.; Pi, C.; Xiong, L.; Chen, J.; Liu, H.; Tang, J.; Zhong, Y.; et al. Molecular mechanisms of rapid-acting antidepressants: New perspectives for developing antidepressants. Pharmacol. Res. 2023, 194, 106837. [Google Scholar] [CrossRef]
- Tang, M.; Ai, Y.; Zhu, S.; Song, N.; Xu, X.; Liang, L.; Rong, B.; Zheng, X.; Zhang, L.; He, T. Antidepressant-Like Effect of Essential Oils From Citrus reticulata in Reserpine-Induced Depressive Mouse. Nat. Prod. Commun. 2022, 17, 1934578X221093916. [Google Scholar]
- Oh, H.A.; Kim, D.-E.; Choi, H.J.; Kim, N.J.; Kim, D.-H. Anti-stress Effects of 20(S)-Protopanaxadiol and 20(S)-Protopanaxatriol in Immobilized Mice. Biol. Pharm. Bull. 2015, 38, 331–335. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef]
- Yan, Z.-Y.; Jiao, H.-Y.; Chen, J.-B.; Zhang, K.-W.; Wang, X.-H.; Jiang, Y.-M.; Liu, Y.-Y.; Xue, Z.; Ma, Q.-Y.; Li, X.-J.; et al. Antidepressant Mechanism of Traditional Chinese Medicine Formula Xiaoyaosan in CUMS-Induced Depressed Mouse Model via RIPK1-RIPK3-MLKL Mediated Necroptosis Based on Network Pharmacology Analysis. Front. Pharmacol. 2021, 12, 773562. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, N.; Yang, C.; Li, X.M.; Zhou, Z.Q.; Yang, J.J. Ketamine-induced antidepressant effects are associated with AMPA receptors-mediated upregulation of mTOR and BDNF in rat hippocampus and prefrontal cortex. Eur. Psychiatry J. Assoc. Eur. Psychiatr. 2014, 29, 419–423. [Google Scholar] [CrossRef]
- Majd, M.; Saunders, E.F.H.; Engeland, C.G. Inflammation and the dimensions of depression: A review. Front. Neuroendocrinol. 2020, 56, 100800. [Google Scholar] [CrossRef]
- Roohi, E.; Jaafari, N.; Hashemian, F. On inflammatory hypothesis of depression: What is the role of IL-6 in the middle of the chaos? J. Neuroinflammation 2021, 18, 45. [Google Scholar] [CrossRef]
- Hainmueller, T.; Bartos, M. Dentate gyrus circuits for encoding, retrieval and discrimination of episodic memories. Nat. Rev. Neurosci. 2020, 21, 153–168. [Google Scholar] [CrossRef]
- Jeong, N.; Singer, A.C. Learning from inhibition: Functional roles of hippocampal CA1 inhibition in spatial learning and memory. Curr. Opin. Neurobiol. 2022, 76, 102604. [Google Scholar] [CrossRef]
- Song, D.; Wang, D.; Yang, Q.; Yan, T.; Wang, Z.; Yan, Y.; Zhao, J.; Xie, Z.; Liu, Y.; Ke, Z.; et al. The lateralization of left hippocampal CA3 during the retrieval of spatial working memory. Nat. Commun. 2020, 11, 2901. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, C.; Qin, C.; Zhang, X.; Feng, H.; Wang, Z.; Li, J. Evaluation of the effect of myelotomy on nerve function in rats with spinal cord injury by diffusion tensor imaging. Acta Radiol. 2021, 62, 1666–1673. [Google Scholar] [CrossRef]
- Law, N.C.; Hunzicker-Dunn, M.E. Insulin Receptor Substrate 1, the Hub Linking Follicle-stimulating Hormone to Phosphatidylinositol 3-Kinase Activation. J. Biol. Chem. 2016, 291, 4547–4560. [Google Scholar] [CrossRef]
- Kang, K.; Wang, D.P.; Lv, Q.L.; Chen, F. VEGF-A ameliorates ischemia hippocampal neural injury via regulating autophagy and Akt/CREB signaling in a rat model of chronic cerebral hypoperfusion. J. Stroke Cerebrovasc. Dis. Off. J. Natl. Stroke Assoc. 2023, 32, 107367. [Google Scholar] [CrossRef]
- Tao, W.; Dong, Y.; Su, Q.; Wang, H.; Chen, Y.; Xue, W.; Chen, C.; Xia, B.; Duan, J.; Chen, G. Liquiritigenin reverses depression-like behavior in unpredictable chronic mild stress-induced mice by regulating PI3K/Akt/mTOR mediated BDNF/TrkB pathway. Behav. Brain Res. 2016, 308, 177–186. [Google Scholar] [CrossRef]
- Matsuda, S.; Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y.J.D. Roles of PI3K/AKT/GSK3 Pathway Involved in Psychiatric Illnesses. Diseases 2019, 7, 22. [Google Scholar] [CrossRef]
- Ignácio, Z.M.; Réus, G.Z.; Arent, C.O.; Abelaira, H.M.; Pitcher, M.R.; Quevedo, J. New perspectives on the involvement of mTOR in depression as well as in the action of antidepressant drugs. Br. J. Clin. Pharmacol. 2016, 82, 1280–1290. [Google Scholar] [CrossRef]
- He, W.; Wang, Y.; Yang, R.; Ma, H.; Qin, X.; Yan, M.; Rong, Y.; Xie, Y.; Li, L.; Si, J.; et al. Molecular Mechanism of Naringenin Against High-Glucose-Induced Vascular Smooth Muscle Cells Proliferation and Migration Based on Network Pharmacology and Transcriptomic Analyses. Front. Pharmacol. 2022, 13, 862709. [Google Scholar] [CrossRef]
- Frye, M.A.; Nassan, M.; Jenkins, G.D.; Kung, S.; Veldic, M.; Palmer, B.A.; Feeder, S.E.; Tye, S.J.; Choi, D.S.; Biernacka, J.M. Feasibility of investigating differential proteomic expression in depression: Implications for biomarker development in mood disorders. Transl. Psychiatry 2015, 5, e689. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qi, S.M.; Wang, Y.; Jiang, H.W.; Li, Y.H.; Zhao, B.C.; Zhang, T.; Sun, Y.; Gao, X.Z.; Quan, S.X.; et al. Antidepressant effect of electroacupuncture on modulating the expression of c-Fos/AP-1 through the JNK signaling pathway. Anat. Rec. 2021, 304, 2480–2493. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.G.; Chang, Y.; Ryu, D.R.; Song, T.J. Plasma Klotho concentration is associated with the presence, burden and progression of cerebral small vessel disease in patients with acute ischaemic stroke. PLoS ONE 2019, 14, e0220796. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | PDB ID | Molecular Docking Score (kcal/mol) |
|---|---|---|
| PI3K | 1E8X | −7.9 |
| AKT | 7WM1 | −7.6 |
| FOS | 1S9K | −8.8 |
| TTR | 3D7P | −7.6 |
| KL | 5VAK | −6.8 |
| Gene Name | Primer Sequences (5′ → 3′) |
|---|---|
| GAPDH | F:GCCTCCTCCAATTCAACCCT |
| R:CTCGTGGTTCACACCCATCA | |
| PI3K | F:ACCTTAAATGGTGAGCACGGA |
| R:GGCCCGCACTGTAACCTATT | |
| AKT | F:CCGCCTGATCAAGTTCTCCT |
| R:AGAGGGAGAGGGCCAGTTAG | |
| TTR | F:TTCCGTCTGCTCCTCCTTT |
| R:AACACCTTCACACCCACATTC | |
| FOS | F:CTGAGTGTCTCAAGTGCCTC |
| R:CTCGGGTTGTAGGATTGAG | |
| KL | F:CCTCCTTTACCTGAGAACCA |
| R:GCACATCCCACAGATAGACA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Su, L.; Shi, M.; Sun, L.; Chen, W.; Geng, J.; Li, J.; Zong, Y.; He, Z.; Du, R. Network Pharmacology and Transcriptomics to Explore the Pharmacological Mechanisms of 20(S)-Protopanaxatriol in the Treatment of Depression. Int. J. Mol. Sci. 2024, 25, 7574. https://doi.org/10.3390/ijms25147574
Guo X, Su L, Shi M, Sun L, Chen W, Geng J, Li J, Zong Y, He Z, Du R. Network Pharmacology and Transcriptomics to Explore the Pharmacological Mechanisms of 20(S)-Protopanaxatriol in the Treatment of Depression. International Journal of Molecular Sciences. 2024; 25(14):7574. https://doi.org/10.3390/ijms25147574
Chicago/Turabian StyleGuo, Xiangjuan, Lili Su, Meiling Shi, Li Sun, Weijia Chen, Jianan Geng, Jianming Li, Ying Zong, Zhongmei He, and Rui Du. 2024. "Network Pharmacology and Transcriptomics to Explore the Pharmacological Mechanisms of 20(S)-Protopanaxatriol in the Treatment of Depression" International Journal of Molecular Sciences 25, no. 14: 7574. https://doi.org/10.3390/ijms25147574
APA StyleGuo, X., Su, L., Shi, M., Sun, L., Chen, W., Geng, J., Li, J., Zong, Y., He, Z., & Du, R. (2024). Network Pharmacology and Transcriptomics to Explore the Pharmacological Mechanisms of 20(S)-Protopanaxatriol in the Treatment of Depression. International Journal of Molecular Sciences, 25(14), 7574. https://doi.org/10.3390/ijms25147574