

Cannabinoid and Orexigenic Systems Interplay as a New Focus of Research in Alzheimer’s Disease
 , and
, and {kind=link}
{kind=link}
Abstract
1. Introduction
2. GPCRs and Alzheimer’s Disease
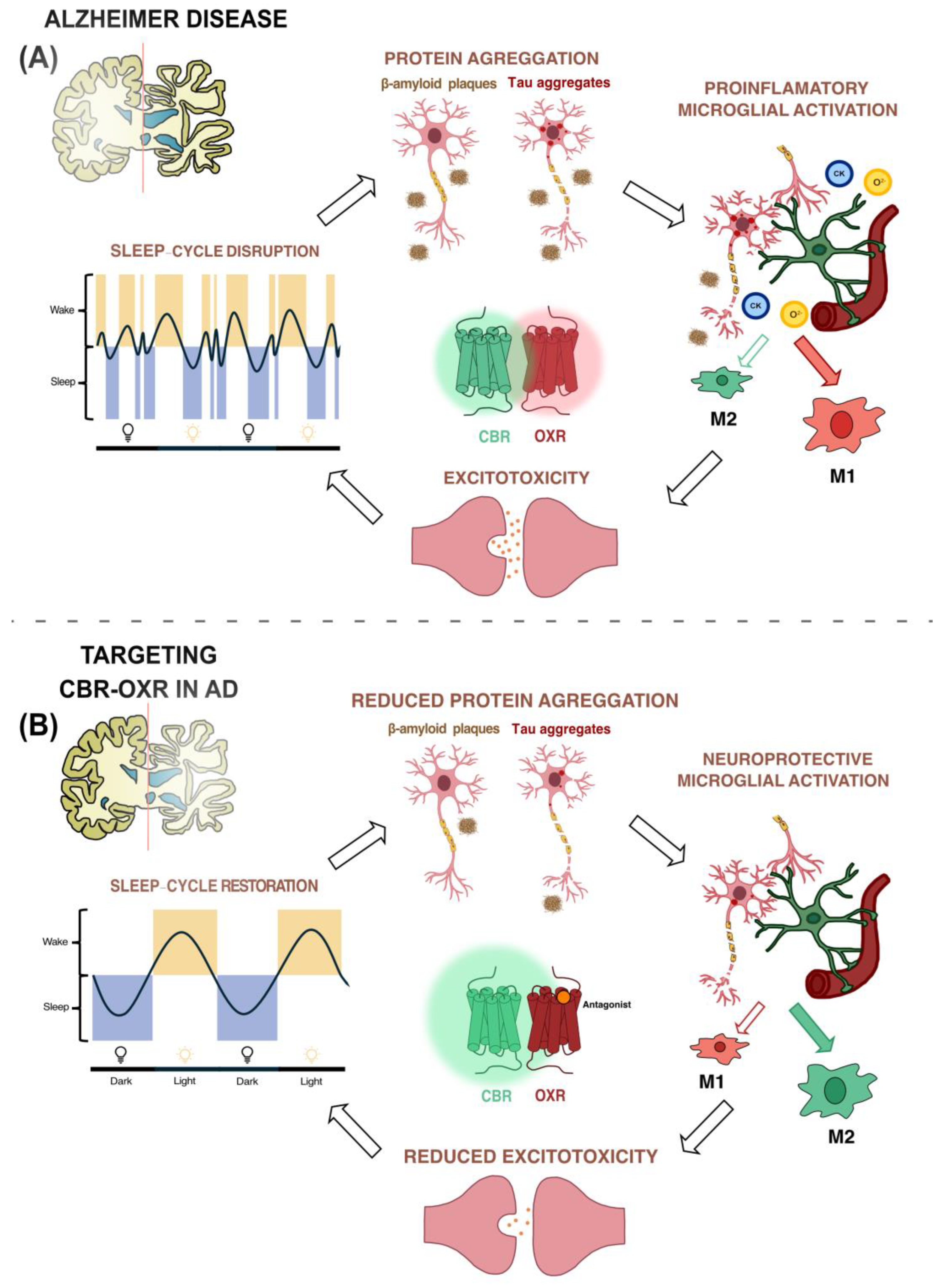
3. Involvement of Orexin System in Alzheimer’s Disease
4. Role of Endocannabinoid System in Alzheimer’s Disease
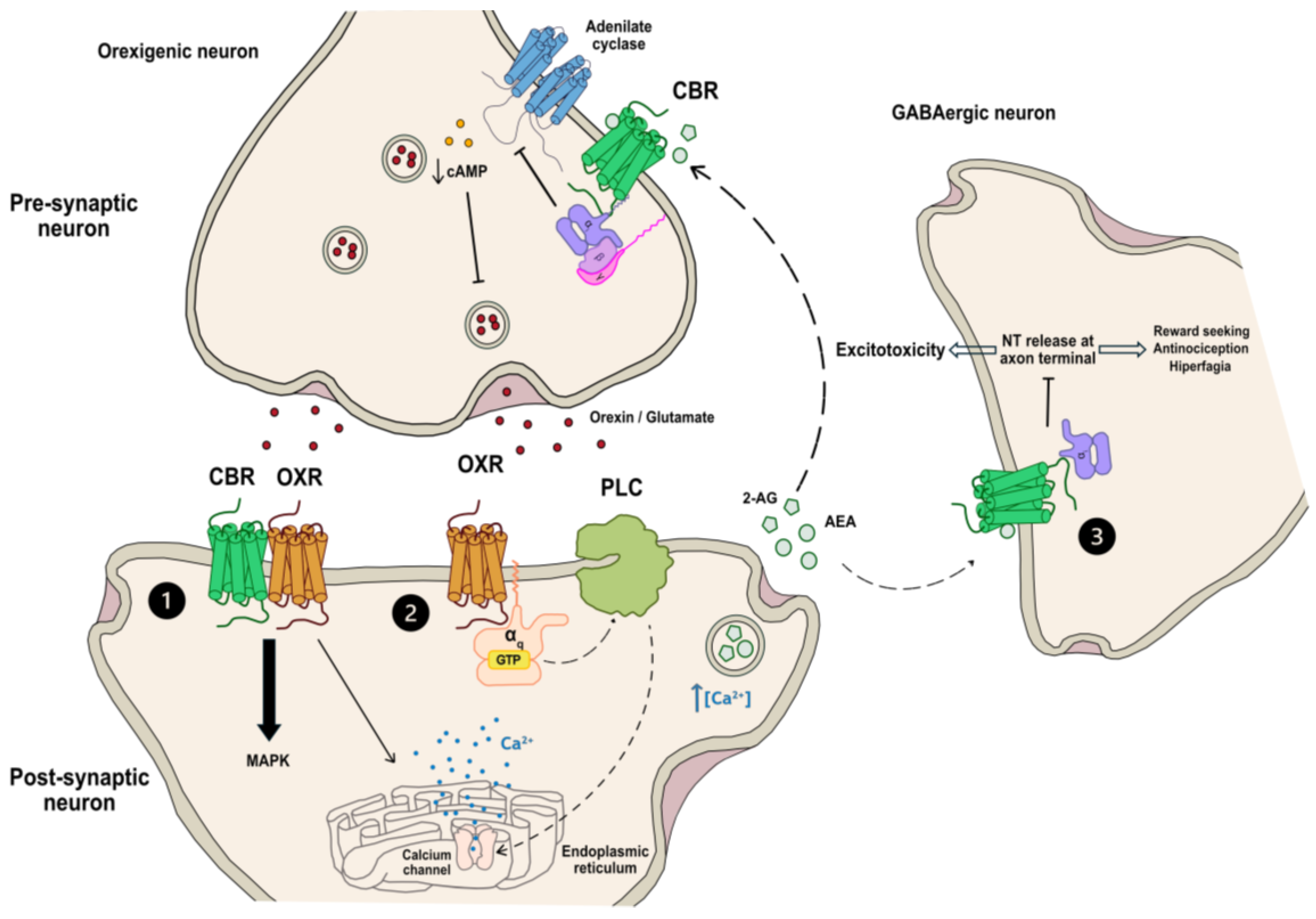
5. Interactions between Orexigenic and Endocannabinoid Systems
6. Orexin–Cannabinoid Heteromer Formation as a Potential Target to Combat Alzheimer’s Disease
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fonseca, C.; Ettcheto, M.; Bicker, J.; Fernandes, M.J.; Falcão, A.; Camins, A.; Fortuna, A. Under the Umbrella of Depression and Alzheimer’s Disease Physiopathology: Can Cannabinoids Be a Dual-Pleiotropic Therapy? Ageing Res. Rev. 2023, 90, 101998. [Google Scholar] [CrossRef] [PubMed]
- Tanzi, R.E. The Genetics of Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006296. [Google Scholar] [CrossRef] [PubMed]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The Diagnosis of Dementia Due to Alzheimer’s Disease: Recommendations from the National Institute on Aging-Alzheimer’s Association Workgroups on Diagnostic Guidelines for Alzheimer’s Disease. Alzheimer’s Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of Acetylcholinesterase Inhibitors in Alzheimer’s Disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef]
- Kabir, M.T.; Sufian, M.A.; Uddin, M.S.; Begum, M.M.; Akhter, S.; Islam, A.; Mathew, B.; Islam, M.S.; Amran, M.S.; Md Ashraf, G. NMDA Receptor Antagonists: Repositioning of Memantine as a Multitargeting Agent for Alzheimer’s Therapy. Curr. Pharm. Des. 2019, 25, 3506–3518. [Google Scholar] [CrossRef]
- Bonifazi, A.; Del Bello, F.; Giorgioni, G.; Piergentili, A.; Saab, E.; Botticelli, L.; Cifani, C.; Micioni Di Bonaventura, E.; Micioni Di Bonaventura, M.V.; Quaglia, W. Targeting Orexin Receptors: Recent Advances in the Development of Subtype Selective or Dual Ligands for the Treatment of Neuropsychiatric Disorders. Med. Res. Rev. 2023, 43, 1607–1667. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Ravichandran, M. Orexin Receptors: Targets and Applications. Fundam. Clin. Pharmacol. 2022, 36, 72–80. [Google Scholar] [CrossRef]
- Ye, L.; Cao, Z.; Wang, W.; Zhou, N. New Insights in Cannabinoid Receptor Structure and Signaling. Curr. Mol. Pharmacol. 2019, 12, 239–248. [Google Scholar] [CrossRef]
- Zhang, Y.; Devries, M.E.; Skolnick, J. Structure Modeling of All Identified G Protein–Coupled Receptors in the Human Genome. PLoS Comput. Biol. 2006, 2, 88–99. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, Q.; Wu, B. Structural Studies of G Protein-Coupled Receptors. Mol. Cells 2015, 38, 836. [Google Scholar] [CrossRef]
- Changeux, J.P.; Edelstein, S.J. Allosteric Mechanisms of Signal Transduction. Science 2005, 308, 1424–1428. [Google Scholar] [CrossRef] [PubMed]
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and Dynamics of GPCR Signaling Complexes. Nat. Struct. Mol. Biol. 2018, 25, 4. [Google Scholar] [CrossRef] [PubMed]
- George, S.R.; O’Dowd, B.F.; Lee, S.P. G-Protein-Coupled Receptor Oligomerization and Its Potential for Drug Discovery. Nat. Rev. Drug Discov. 2002, 1, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Jordan, B.A.; Devi, L.A. G-Protein-Coupled Receptor Heterodimerization Modulates Receptor Function. Nature 1999, 399, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological Alterations in Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, B.C.; Bakkour, A.; Salat, D.H.; Feczko, E.; Pacheco, J.; Greve, D.N.; Grodstein, F.; Wright, C.I.; Blacker, D.; Rosas, H.D.; et al. The Cortical Signature of Alzheimer’s Disease: Regionally Specific Cortical Thinning Relates to Symptom Severity in Very Mild to Mild AD Dementia and Is Detectable in Asymptomatic Amyloid-Positive Individuals. Cereb. Cortex 2009, 19, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Todd, N.; Thathiah, A. The Role of GPCRs in Neurodegenerative Diseases: Avenues for Therapeutic Intervention. Curr. Opin. Pharmacol. 2017, 32, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Liguori, C. Orexin and Alzheimer’s Disease. Curr. Top. Behav. Neurosci. 2016, 33, 305–322. [Google Scholar] [CrossRef] [PubMed]
- Bedse, G.; Romano, A.; Lavecchia, A.M.; Cassano, T.; Gaetani, S. The Role of Endocannabinoid Signaling in the Molecular Mechanisms of Neurodegeneration in Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 43, 1115–1136. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Q.; Ji, B.; Pan, Y.; Xu, C.; Cheng, B.; Bai, B.; Chen, J. The Orexin/Receptor System: Molecular Mechanism and Therapeutic Potential for Neurological Diseases. Front. Mol. Neurosci. 2018, 11, 384999. [Google Scholar] [CrossRef]
- Thathiah, A.; Spittaels, K.; Hoffmann, M.; Staes, M.; Cohen, A.; Horré, K.; Vanbrabant, M.; Coun, F.; Baekelandt, V.; Delacourte, A.; et al. The Orphan G Protein-Coupled Receptor 3 Modulates Amyloid-Beta Peptide Generation in Neurons. Science 2009, 323, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Thathiah, A.; De Strooper, B. The Role of G Protein-Coupled Receptors in the Pathology of Alzheimer’s Disease. Nat. Rev. Neurosci. 2011, 12, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, X.; Zeng, X.; Bossers, K.; Swaab, D.F.; Zhao, J.; Pei, G. β-Arrestin1 Regulates γ-Secretase Complex Assembly and Modulates Amyloid-β Pathology. Cell Res. 2012, 23, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Maia, M.A.; Sousa, E. BACE-1 and γ-Secretase as Therapeutic Targets for Alzheimer’s Disease. Pharmaceuticals 2019, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhou, Q.; Labroska, V.; Qin, S.; Darbalaei, S.; Wu, Y.; Yuliantie, E.; Xie, L.; Tao, H.; Cheng, J.; et al. G Protein-Coupled Receptors: Structure- and Function-Based Drug Discovery. Signal Transduct. Target. Ther. 2021, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Schöne, C.; Burdakov, D. Glutamate and GABA as Rapid Effectors of Hypothalamic “Peptidergic” Neurons. Front. Behav. Neurosci. 2012, 6, 35573. [Google Scholar] [CrossRef]
- Stadel, J.M.; Wilson, S.; Bergsma, D.J. Orphan G Protein-Coupled Receptors: A Neglected Opportunity for Pioneer Drug Discovery. Trends Pharmacol. Sci. 1997, 18, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Chiou, L.-C.; Lee, H.-J.; Ho, Y.-C.; Chen, S.-P.; Liao, Y.-Y.; Ma, C.-H.; Fan, P.-C.; Fuh, J.-L.; Wang, S.-J. Orexins/Hypocretins: Pain Regulation and Cellular Actions. Curr. Pharm. Des. 2010, 16, 3089–3100. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and Orexin Receptors: A Family of Hypothalamic Neuropeptides and G Protein-Coupled Receptors That Regulate Feeding Behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef]
- Couvineau, A.; Voisin, T.; Nicole, P.; Gratio, V.; Abad, C.; Tan, Y.V. Orexins as Novel Therapeutic Targets in Inflammatory and Neurodegenerative Diseases. Front. Endocrinol. 2019, 10, 488305. [Google Scholar] [CrossRef]
- Gencik, M.; Dahmen, N.; Wieczorek, S.; Kasten, M.; Bierbrauer, J.; Anghelescu, I.; Szegedi, A.; Menezes Saecker, A.M.; Epplen, J.T. A Prepro-Orexin Gene Polymorphism Is Associated with Narcolepsy. Neurology 2001, 56, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Tsujino, N.; Sakurai, T. Orexin/Hypocretin: A Neuropeptide at the Interface of Sleep, Energy Homeostasis, and Reward System. Pharmacol. Rev. 2009, 61, 162–176. [Google Scholar] [CrossRef] [PubMed]
- De Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.B.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.F.; Gautvik, V.T.; Bartlett, F.S.; et al. The Hypocretins: Hypothalamus-Specific Peptides with Neuroexcitatory Activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Ammoun, S.; Holmqvist, T.; Shariatmadari, R.; Oonk, H.B.; Detheux, M.; Parmentier, M.; Åkerman, K.E.O.; Kukkonen, J.P. Distinct Recognition of OX1 and OX2 Receptors by Orexin Peptides. J. Pharmacol. Exp. Ther. 2003, 305, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Turunen, P.M.; Jäntti, M.H.; Kukkonen, J.P. OX1 Orexin/Hypocretin Receptor Signaling through Arachidonic Acid and Endocannabinoid Release. Mol. Pharmacol. 2012, 82, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Marcus, J.N.; Aschkenasi, C.J.; Lee, C.E.; Chemelli, R.M.; Saper, C.B.; Yanagisawa, M.; Elmquist, J.K. Differential Expression of Orexin Receptors 1 and 2 in the Rat Brain. J. Comp. Neurol. 2001, 435, 6–25. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Liu, T.; Tuo, M.; Chi, S. The Role of Orexin in Alzheimer Disease: From Sleep-Wake Disturbance to Therapeutic Target. Neurosci. Lett. 2021, 765, 136247. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Winer, J.R.; Jagust, W.J.; Walker, M.P. Sleep: A Novel Mechanistic Pathway, Biomarker, and Treatment Target in the Pathology of Alzheimer’s Disease? Trends Neurosci. 2016, 39, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Liguori, C.; Maestri, M.; Spanetta, M.; Placidi, F.; Bonanni, E.; Mercuri, N.B.; Guarnieri, B. Sleep-Disordered Breathing and the Risk of Alzheimer’s Disease. Sleep Med. Rev. 2021, 55, 101375. [Google Scholar] [CrossRef]
- Wilckens, K.A.; Tudorascu, D.L.; Snitz, B.E.; Price, J.C.; Aizenstein, H.J.; Lopez, O.L.; Erickson, K.I.; Lopresti, B.J.; Laymon, C.M.; Minhas, D.; et al. Sleep Moderates the Relationship between Amyloid Beta and Memory Recall. Neurobiol. Aging 2018, 71, 142–148. [Google Scholar] [CrossRef]
- Um, Y.H.; Lim, H.K. Orexin and Alzheimer’s Disease: A New Perspective. Psychiatry Investig. 2020, 17, 616. [Google Scholar] [CrossRef] [PubMed]
- Mavanji, V.; Perez-Leighton, C.E.; Kotz, C.M.; Billington, C.J.; Parthasarathy, S.; Sinton, C.M.; Teske, J.A. Promotion of Wakefulness and Energy Expenditure by Orexin-A in the Ventrolateral Preoptic Area. Sleep 2015, 38, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Van Egroo, M.; Narbutas, J.; Chylinski, D.; Villar González, P.; Maquet, P.; Salmon, E.; Bastin, C.; Collette, F.; Vandewalle, G. Sleep–Wake Regulation and the Hallmarks of the Pathogenesis of Alzheimer’s Disease. Sleep 2019, 42, zsz017. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Jiang, W.; Zhang, E.E. Orexin Signaling Regulates Both the Hippocampal Clock and the Circadian Oscillation of Alzheimer’s Disease-Risk Genes. Sci. Rep. 2016, 6, 36035. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.H.; Jiang, H.; Finn, M.B.; Stewart, F.R.; Mahan, T.E.; Cirrito, J.R.; Heda, A.; Joy Snider, B.; Li, M.; Yanagisawa, M.; et al. Potential Role of Orexin and Sleep Modulation in the Pathogenesis of Alzheimer’s Disease. J. Exp. Med. 2014, 211, 2487–2496. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Cho, M.H.; Kim, D.H.; Chung, S.; Yoon, S.Y. Orexin Impairs the Phagocytosis and Degradation of Amyloid-β Fibrils by Microglial Cells. J. Alzheimer’s Dis. 2017, 58, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-Mediated Control of Synaptic Transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Cannabinoid Pharmacology: The First 66 Years. Br. J. Pharmacol. 2006, 147, S163. [Google Scholar] [CrossRef]
- Di Marzo, V.; Piscitelli, F. The Endocannabinoid System and Its Modulation by Phytocannabinoids. Neurotherapeutics 2015, 12, 692. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.J.; Williams, C.M.; Whalley, B.J.; Stephens, G.J. Phytocannabinoids as Novel Therapeutic Agents in CNS Disorders. Pharmacol. Ther. 2012, 133, 79–97. [Google Scholar] [CrossRef]
- Abellanas, M.A.; Aymerich, M.S. Cannabinoid Receptor Type 2 as a Therapeutic Target for Parkinson’s Disease. In Diagnosis and Management in Parkinson’s Disease; Academic Press: Cambridge, MA, USA, 2020; Volume 1, pp. 557–573. [Google Scholar] [CrossRef]
- Ibeas Bih, C.; Chen, T.; Nunn, A.V.W.; Bazelot, M.; Dallas, M.; Whalley, B.J. Molecular Targets of Cannabidiol in Neurological Disorders. Neurotherapeutics 2015, 12, 699–730. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Dysarz, F.A.; Johnson, M.R.; Melvin, L.S.; Howlett, A.C. Determination and Characterization of a Cannabinoid Receptor in Rat Brain. Mol. Pharmacol. 1988, 34, 605–613. [Google Scholar] [PubMed]
- Devane, W.A.; Hanuš, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and Structure of a Brain Constituent That Binds to the Cannabinoid Receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R.; et al. Identification of an Endogenous 2-Monoglyceride, Present in Canine Gut, That Binds to Cannabinoid Receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Brusco, A.; Tagliaferro, P.A.; Saez, T.; Onaivi, E.S. Ultrastructural Localization of Neuronal Brain CB2 Cannabinoid Receptors. Ann. N. Y. Acad. Sci. 2008, 1139, 450–457. [Google Scholar] [CrossRef]
- Gong, J.P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 Receptors: Immunohistochemical Localization in Rat Brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef]
- Bietar, B.; Tanner, S.; Lehmann, C. Neuroprotection and Beyond: The Central Role of CB1 and CB2 Receptors in Stroke Recovery. Int. J. Mol. Sci. 2023, 24, 16728. [Google Scholar] [CrossRef]
- Duncan, R.S.; Riordan, S.M.; Gernon, M.C.; Koulen, P. Cannabinoids and Endocannabinoids as Therapeutics for Nervous System Disorders: Preclinical Models and Clinical Studies. Neural Regen. Res. 2024, 19, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Motta, C.; Musella, A.; Centonze, D. The Interplay between Inflammatory Cytokines and the Endocannabinoid System in the Regulation of Synaptic Transmission. Neuropharmacology 2015, 96, 105–112. [Google Scholar] [CrossRef]
- Magham, S.V.; Thaggikuppe Krishnamurthy, P.; Shaji, N.; Mani, L.; Balasubramanian, S. Cannabinoid Receptor 2 Selective Agonists and Alzheimer’s Disease: An Insight into the Therapeutic Potentials. J. Neurosci. Res. 2021, 99, 2888–2905. [Google Scholar] [CrossRef]
- Tolón, R.M.; Núñez, E.; Pazos, M.R.; Benito, C.; Castillo, A.I.; Martínez-Orgado, J.A.; Romero, J. The Activation of Cannabinoid CB2 Receptors Stimulates in Situ and in Vitro Beta-Amyloid Removal by Human Macrophages. Brain Res. 2009, 1283, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Zarruk, J.G.; Fernández-López, D.; García-Yébenes, I.; García-Gutiérrez, M.S.; Vivancos, J.; Nombela, F.; Torres, M.; Burguete, M.C.; Manzanares, J.; Lizasoain, I.; et al. Cannabinoid Type 2 Receptor Activation Downregulates Stroke-Induced Classic and Alternative Brain Macrophage/Microglial Activation Concomitant to Neuroprotection. Stroke 2012, 43, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Bie, B.; Wu, J.; Foss, J.F.; Naguib, M. An Overview of the Cannabinoid Type 2 Receptor System and Its Therapeutic Potential. Curr. Opin. Anaesthesiol. 2018, 31, 407–414. [Google Scholar] [CrossRef]
- Ellis, J.; Pediani, J.D.; Canals, M.; Milasta, S.; Milligan, G. Orexin-1 Receptor-Cannabinoid CB1 Receptor Heterodimerization Results in Both Ligand-Dependent and -Independent Coordinated Alterations of Receptor Localization and Function. J. Biol. Chem. 2006, 281, 38812–38824. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.J.; Zagzoog, A.; Smolyakova, A.M.; Ezeaka, U.C.; Benko, M.J.; Holt, T.; Laprairie, R.B. In Vivo Evidence for Brain Region-Specific Molecular Interactions Between Cannabinoid and Orexin Receptors. Front. Neurosci. 2021, 15, 790546. [Google Scholar] [CrossRef] [PubMed]
- Jäntti, M.H.; Putula, J.; Turunen, P.M.; Näsman, J.; Reijonen, S.; Lindqvist, C.; Kukkonen, J.P. Autocrine Endocannabinoid Signaling through CB1 Receptors Potentiates OX1 Orexin Receptor Signaling. Mol. Pharmacol. 2013, 83, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Rezende, B.; Alencar, A.K.N.; de Bem, G.F.; Fontes-Dantas, F.L.; Montes, G.C. Endocannabinoid System: Chemical Characteristics and Biological Activity. Pharmaceuticals 2023, 16, 148. [Google Scholar] [CrossRef]
- Lu, H.C.; MacKie, K. An Introduction to the Endogenous Cannabinoid System. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Covelo, A.; Araque, A. Lateral Regulation of Synaptic Transmission by Astrocytes. Neuroscience 2016, 323, 62–66. [Google Scholar] [CrossRef]
- Huang, H.; Acuna-Goycolea, C.; Li, Y.; Cheng, H.M.; Obrietan, K.; Van Den Pol, A.N. Cannabinoids Excite Hypothalamic Melanin-Concentrating Hormone But Inhibit Hypocretin/Orexin Neurons: Implications for Cannabinoid Actions on Food Intake and Cognitive Arousal. J. Neurosci. 2007, 27, 4870–4881. [Google Scholar] [CrossRef]
- Crespo, I.; Gómez de Heras, R.; Rodríguez de Fonseca, F.; Navarro, M. Pretreatment with Subeffective Doses of Rimonabant Attenuates Orexigenic Actions of Orexin A-Hypocretin 1. Neuropharmacology 2008, 54, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Merroun, I.; El Mlili, N.; Martinez, R.; Porres, J.M.; Llopis, J.; Ahabrach, H.; Aranda, P.; Sanchez Gonzalez, C.; Errami, M.; Lopez-Jurado, M. Interaction between Orexin A and Cannabinoid System in the Lateral Hypothalamus of Rats and Effects of Subchronic Intraperitoneal Administration of Cannabinoid Receptor Inverse Agonist on Food Intake and the Nutritive Utilization of Protein. J. Physiol. Pharmacol. 2015, 66, 181–190. [Google Scholar] [PubMed]
- Flores, Á.; Maldonado, R.; Berrendero, F. The Hypocretin/Orexin Receptor-1 as a Novel Target to Modulate Cannabinoid Reward. Biol. Psychiatry 2014, 75, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Fride, E. Physiology. A Hunger for Cannabinoids. Nature 2001, 410, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, F.; Jahangirvand, M.; Pirasteh, A.H.; Moradi, M.; Haghparast, A. Functional Interaction between OX2 and CB1 Receptors in the Ventral Tegmental Area and the Nucleus Accumbens in Response to Place Preference Induced by Chemical Stimulation of the Lateral Hypothalamus. Pharmacol. Biochem. Behav. 2015, 139, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Herring, W.J.; Roth, T.; Krystal, A.D.; Michelson, D. Orexin Receptor Antagonists for the Treatment of Insomnia and Potential Treatment of Other Neuropsychiatric Indications. J. Sleep Res. 2019, 28, e12782. [Google Scholar] [CrossRef] [PubMed]
- Jäntti, M.H.; Mandrika, I.; Kukkonen, J.P. Human Orexin/Hypocretin Receptors Form Constitutive Homo- and Heteromeric Complexes with Each Other and with Human CB1 Cannabinoid Receptors. Biochem. Biophys. Res. Commun. 2014, 445, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Raïch, I.; Rebassa, J.B.; Lillo, J.; Cordomi, A.; Rivas-Santisteban, R.; Lillo, A.; Reyes-Resina, I.; Franco, R.; Navarro, G. Antagonization of OX1 Receptor Potentiates CB2 Receptor Function in Microglia from APPSw/Ind Mice Model. Int. J. Mol. Sci. 2022, 23, 12801. [Google Scholar] [CrossRef] [PubMed]
- Maggio, R.; Millan, M.J. Dopamine D2-D3 Receptor Heteromers: Pharmacological Properties and Therapeutic Significance. Curr. Opin. Pharmacol. 2010, 10, 100–107. [Google Scholar] [CrossRef]
- Navarro, G.; Cordomí, A.; Brugarolas, M.; Moreno, E.; Aguinaga, D.; Pérez-Benito, L.; Ferre, S.; Cortés, A.; Casadó, V.; Mallol, J.; et al. Cross-Communication between Gi and Gs in a G-Protein-Coupled Receptor Heterotetramer Guided by a Receptor C-Terminal Domain. BMC Biol. 2018, 16, 24. [Google Scholar] [CrossRef]
- Chandrasekera, P.C.; Wan, T.C.; Gizewski, E.T.; Auchampach, J.A.; Lasley, R.D. Adenosine A1 Receptors Heterodimerize with Β1- and Β2-Adrenergic Receptors Creating Novel Receptor Complexes with Altered G Protein Coupling and Signaling. Cell. Signal. 2013, 25, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Hilairet, S.; Bouaboula, M.; Carrière, D.; Le Fur, G.; Casellas, P. Hypersensitization of the Orexin 1 Receptor by the CB1 Receptor: Evidence for Cross-Talk Blocked by the Specific CB1 Antagonist, SR141716. J. Biol. Chem. 2003, 278, 23731–23737. [Google Scholar] [CrossRef] [PubMed]
- Sim-Selley, L.J.; Brunk, L.K.; Selley, D.E. Inhibitory Effects of SR141716A on G-Protein Activation in Rat Brain. Eur. J. Pharmacol. 2001, 414, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.J.; Pediani, J.D.; Milligan, G. Heteromultimerization of Cannabinoid CB 1 Receptor and Orexin OX 1 Receptor Generates a Unique Complex in Which Both Protomers Are Regulated by Orexin A. J. Biol. Chem. 2011, 286, 37414–37428. [Google Scholar] [CrossRef]
- Imperatore, R.; Palomba, L.; Morello, G.; Spiezio, A.D.; Piscitelli, F.; Di Marzo, V.; Cristino, L. Formation of OX-1R/CB1R Heteromeric Complexes in Embryonic Mouse Hypothalamic Cells: Effect on Intracellular Calcium, 2-Arachidonoyl-Glycerol Biosynthesis and ERK Phosphorylation. Pharmacol. Res. 2016, 111, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Angers, S.; Salahpour, A.; Bouvier, M. Dimerization: An Emerging Concept for G Protein-Coupled Receptor Ontogeny and Function. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 409–435. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G.; Ramsay, D.; Pascal, G.; Carrillo, J.J. GPCR Dimerisation. Life Sci. 2003, 74, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Szidonya, L.; Cserzo, M.; Hunyady, L. Dimerization and Oligomerization of G-Protein-Coupled Receptors: Debated Structures with Established and Emerging Functions. J. Endocrinol. 2008, 196, 435–453. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G. G Protein-Coupled Receptor Dimerization: Function and Ligand Pharmacology. Mol. Pharmacol. 2004, 66, 1–7. [Google Scholar] [CrossRef]
- Bulenger, S.; Marullo, S.; Bouvier, M. Emerging Role of Homo- and Heterodimerization in G-Protein-Coupled Receptor Biosynthesis and Maturation. Trends Pharmacol. Sci. 2005, 26, 131–137. [Google Scholar] [CrossRef]
- Navarro, G.; Borroto-Escuela, D.; Angelats, E.; Etayo, Í.; Reyes-Resina, I.; Pulido-Salgado, M.; Rodríguez-Pérez, A.I.; Canela, E.I.; Saura, J.; Lanciego, J.L.; et al. Receptor-Heteromer Mediated Regulation of Endocannabinoid Signaling in Activated Microglia. Role of CB1 and CB2 Receptors and Relevance for Alzheimer’s Disease and Levodopa-Induced Dyskinesia. Brain. Behav. Immun. 2018, 67, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santisteban, R.; Lillo, A.; Lillo, J.; Rebassa, J.B.; Contestí, J.S.; Saura, C.A.; Franco, R.; Navarro, G. N-Methyl-D-Aspartate (NMDA) and Cannabinoid CB2 Receptors Form Functional Complexes in Cells of the Central Nervous System: Insights into the Therapeutic Potential of Neuronal and Microglial NMDA Receptors. Alzheimer’s Res. Ther. 2021, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Lillo, A.; Raïch, I.; Lillo, J.; Pérez-Olives, C.; Navarro, G.; Franco, R. Expression of the Adenosine A2A-A3 Receptor Heteromer in Different Brain Regions and Marked Upregulation in the Microglia of the Transgenic APPSw,Ind Alzheimer’s Disease Model. Biomedicines 2022, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Reyes-Resina, I.; Aguinaga, D.; Lillo, A.; Jiménez, J.; Raïch, I.; Borroto-Escuela, D.O.; Ferreiro-Vera, C.; Canela, E.I.; Sánchez de Medina, V.; et al. Potentiation of Cannabinoid Signaling in Microglia by Adenosine A2A Receptor Antagonists. Glia 2019, 67, 2410–2423. [Google Scholar] [CrossRef] [PubMed]
- Rapaka, D.; Bitra, V.R.; Challa, S.R.; Adiukwu, P.C. Potentiation of Microglial Endocannabinoid Signaling Alleviates Neuroinflammation in Alzheimer’s Disease. Neuropeptides 2021, 90, 102196. [Google Scholar] [CrossRef] [PubMed]
- Aso, E.; Ferrer, I. CB2 Cannabinoid Receptor As Potential Target against Alzheimer’s Disease. Front. Neurosci. 2016, 10, 202288. [Google Scholar] [CrossRef]
- Wu, J.; Hocevar, M.; Foss, J.F.; Bie, B.; Naguib, M. Activation of CB2 Receptor System Restores Cognitive Capacity and Hippocampal Sox2 Expression in a Transgenic Mouse Model of Alzheimer’s Disease. Eur. J. Pharmacol. 2017, 811, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Olives, C.; Rivas-Santisteban, R.; Lillo, J.; Navarro, G.; Franco, R. Recent Advances in the Potential of Cannabinoids for Neuroprotection in Alzheimer’s, Parkinson’s, and Huntington’s Diseases. Adv. Exp. Med. Biol. 2021, 1264, 81–92. [Google Scholar] [CrossRef]
- Navarro, G.; Morales, P.; Rodríguez-Cueto, C.; Fernández-Ruiz, J.; Jagerovic, N.; Franco, R. Targeting Cannabinoid CB2 Receptors in the Central Nervous System. Medicinal Chemistry Approaches with Focus on Neurodegenerative Disorders. Front. Neurosci. 2016, 10, 211100. [Google Scholar] [CrossRef]
- Bettica, P.; Squassante, L.; Zamuner, S.; Nucci, G.; Danker-Hopfe, H.; Ratti, E. The Orexin Antagonist SB-649868 Promotes and Maintains Sleep in Men with Primary Insomnia. Sleep 2012, 35, 1097–1104. [Google Scholar] [CrossRef]
- Kang, J.E.; Lim, M.M.; Bateman, R.J.; Lee, J.J.; Smyth, L.P.; Cirrito, J.R.; Fujiki, N.; Nishino, S.; Holtzman, D.M. Amyloid-β Dynamics Are Regulated by Orexin and the Sleep-Wake Cycle. Science 2009, 326, 1005–1007. [Google Scholar] [CrossRef] [PubMed]
- Berrendero, F.; Flores, Á.; Robledo, P. When Orexins Meet Cannabinoids: Bidirectional Functional Interactions. Biochem. Pharmacol. 2018, 157, 43–50. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebassa, J.B.; Capó, T.; Lillo, J.; Raïch, I.; Reyes-Resina, I.; Navarro, G. Cannabinoid and Orexigenic Systems Interplay as a New Focus of Research in Alzheimer’s Disease. Int. J. Mol. Sci. 2024, 25, 5378. https://doi.org/10.3390/ijms25105378
Rebassa JB, Capó T, Lillo J, Raïch I, Reyes-Resina I, Navarro G. Cannabinoid and Orexigenic Systems Interplay as a New Focus of Research in Alzheimer’s Disease. International Journal of Molecular Sciences. 2024; 25(10):5378. https://doi.org/10.3390/ijms25105378
Chicago/Turabian StyleRebassa, Joan Biel, Toni Capó, Jaume Lillo, Iu Raïch, Irene Reyes-Resina, and Gemma Navarro. 2024. "Cannabinoid and Orexigenic Systems Interplay as a New Focus of Research in Alzheimer’s Disease" International Journal of Molecular Sciences 25, no. 10: 5378. https://doi.org/10.3390/ijms25105378
APA StyleRebassa, J. B., Capó, T., Lillo, J., Raïch, I., Reyes-Resina, I., & Navarro, G. (2024). Cannabinoid and Orexigenic Systems Interplay as a New Focus of Research in Alzheimer’s Disease. International Journal of Molecular Sciences, 25(10), 5378. https://doi.org/10.3390/ijms25105378