

Sensitivity of Human Induced Pluripotent Stem Cells and Thereof Differentiated Kidney Proximal Tubular Cells towards Selected Nephrotoxins

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
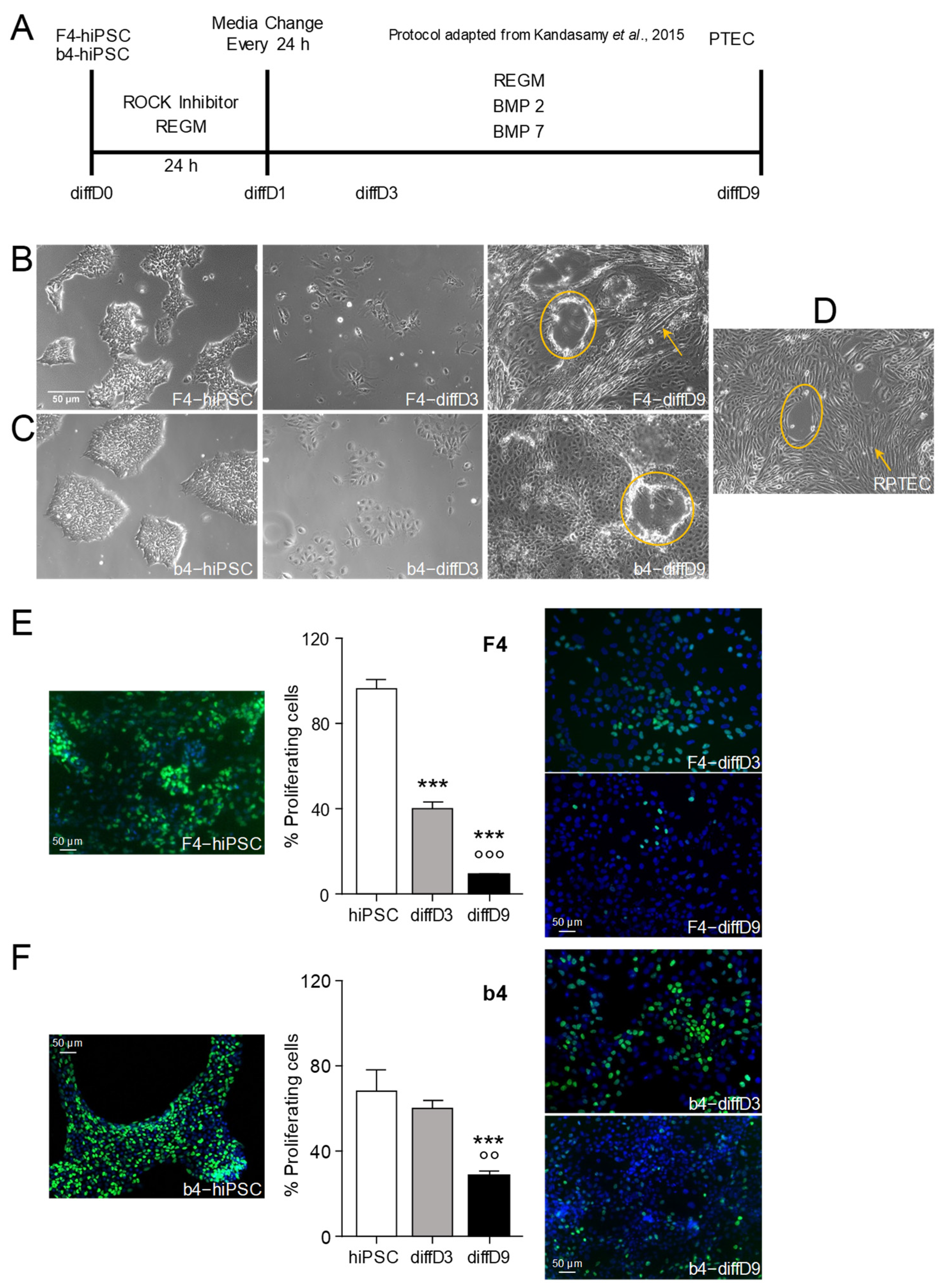
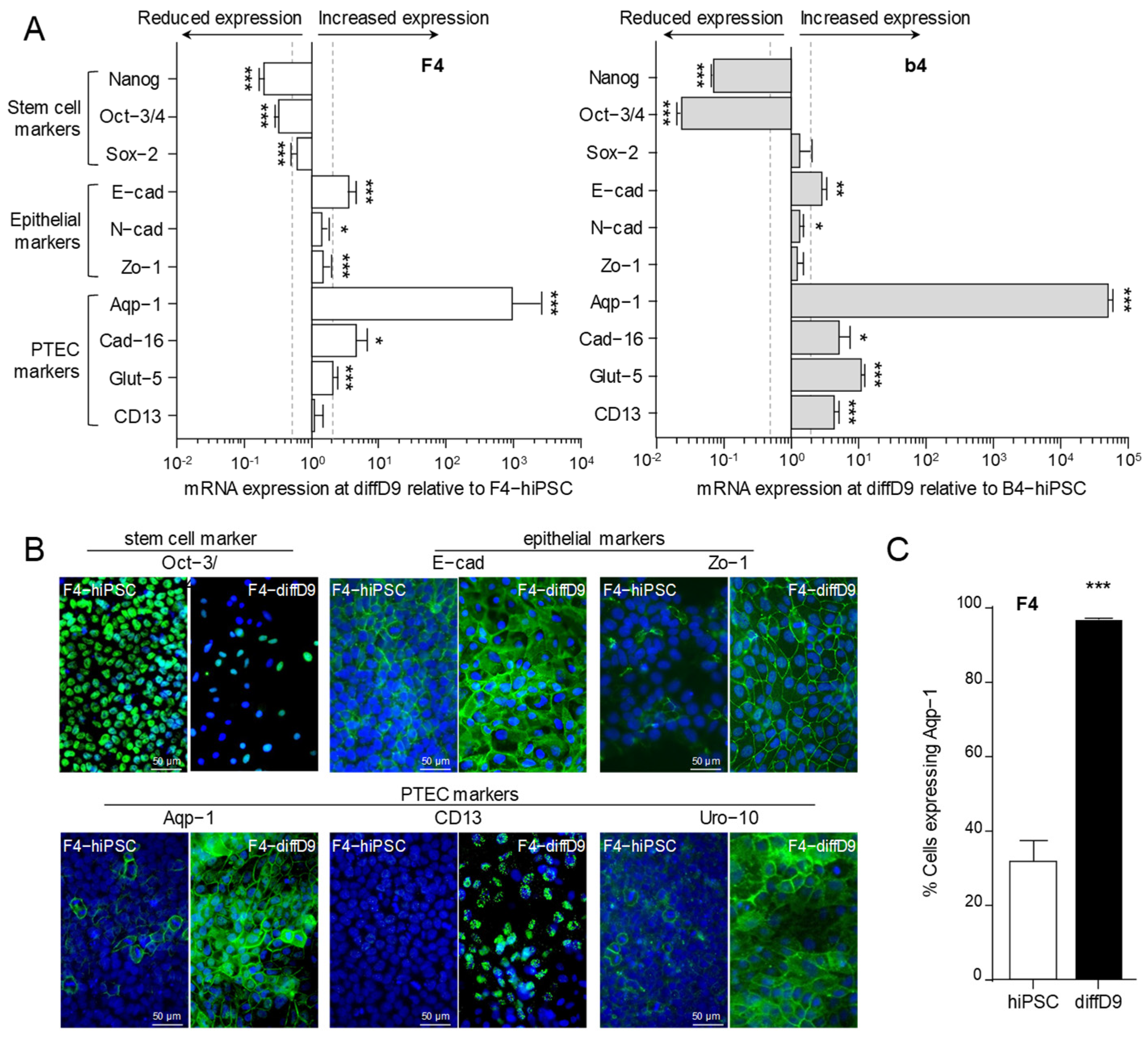
2.1. Differentiation of hiPSC into Proximal Tubular Epithelial-like Cells
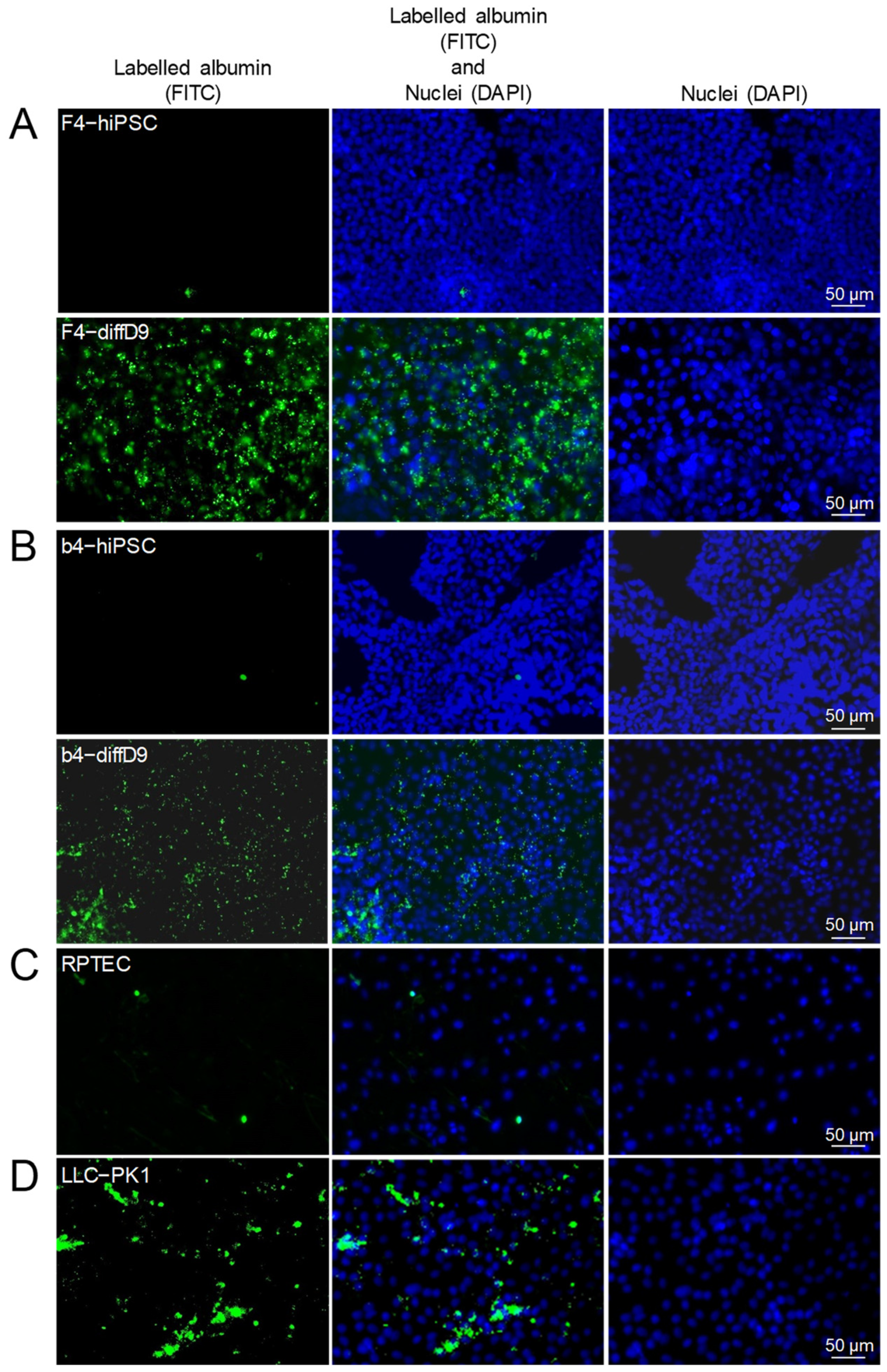
2.2. Transporter Expression and Transport Capacity of PTELC Differentiated out of hiPSC
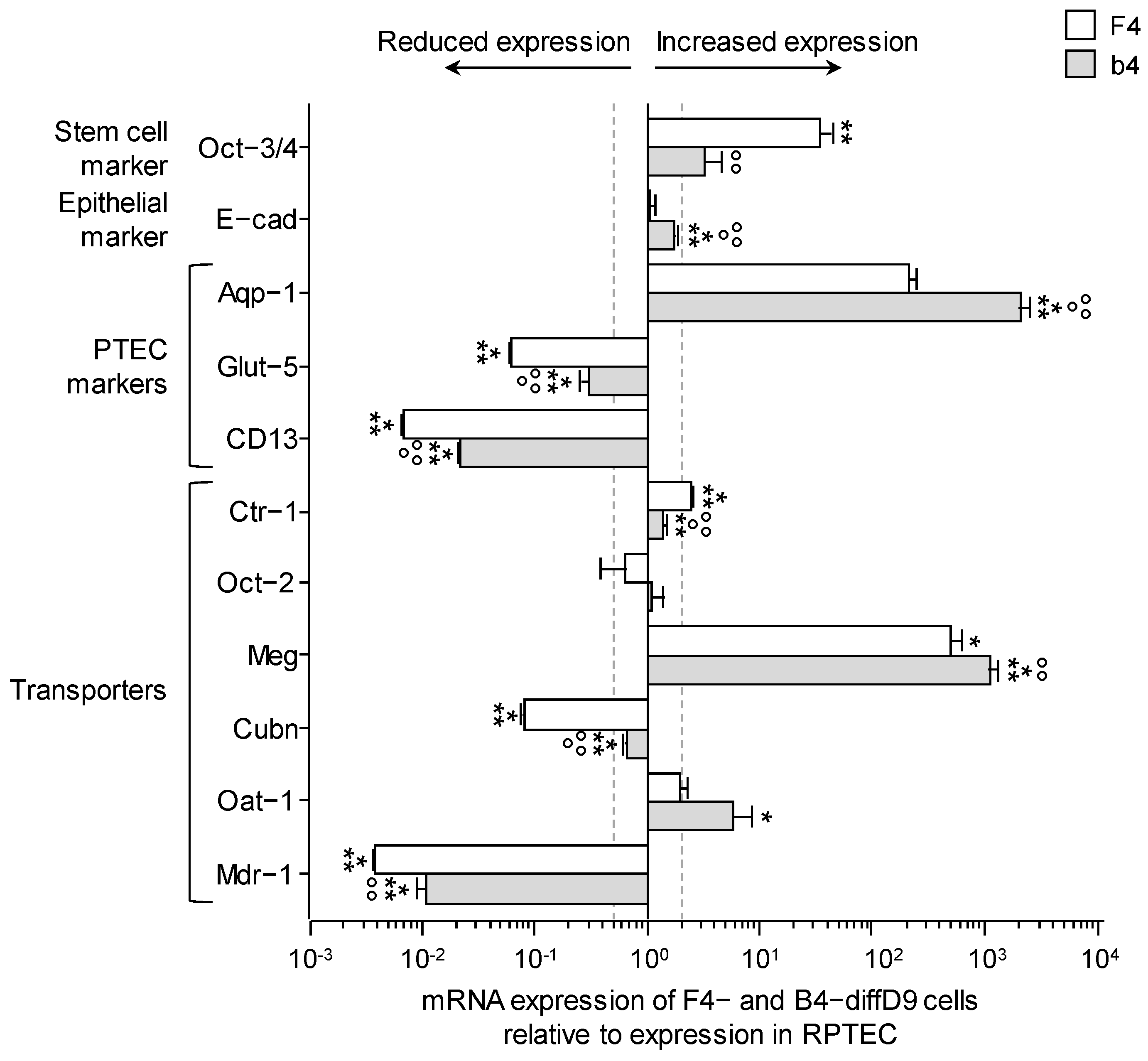
2.3. Comparison of Marker and Transporter Expression as Well as Transport Capacity of the PTELC with the Human Proximal Tubular Cell Line RPTEC/TERT1
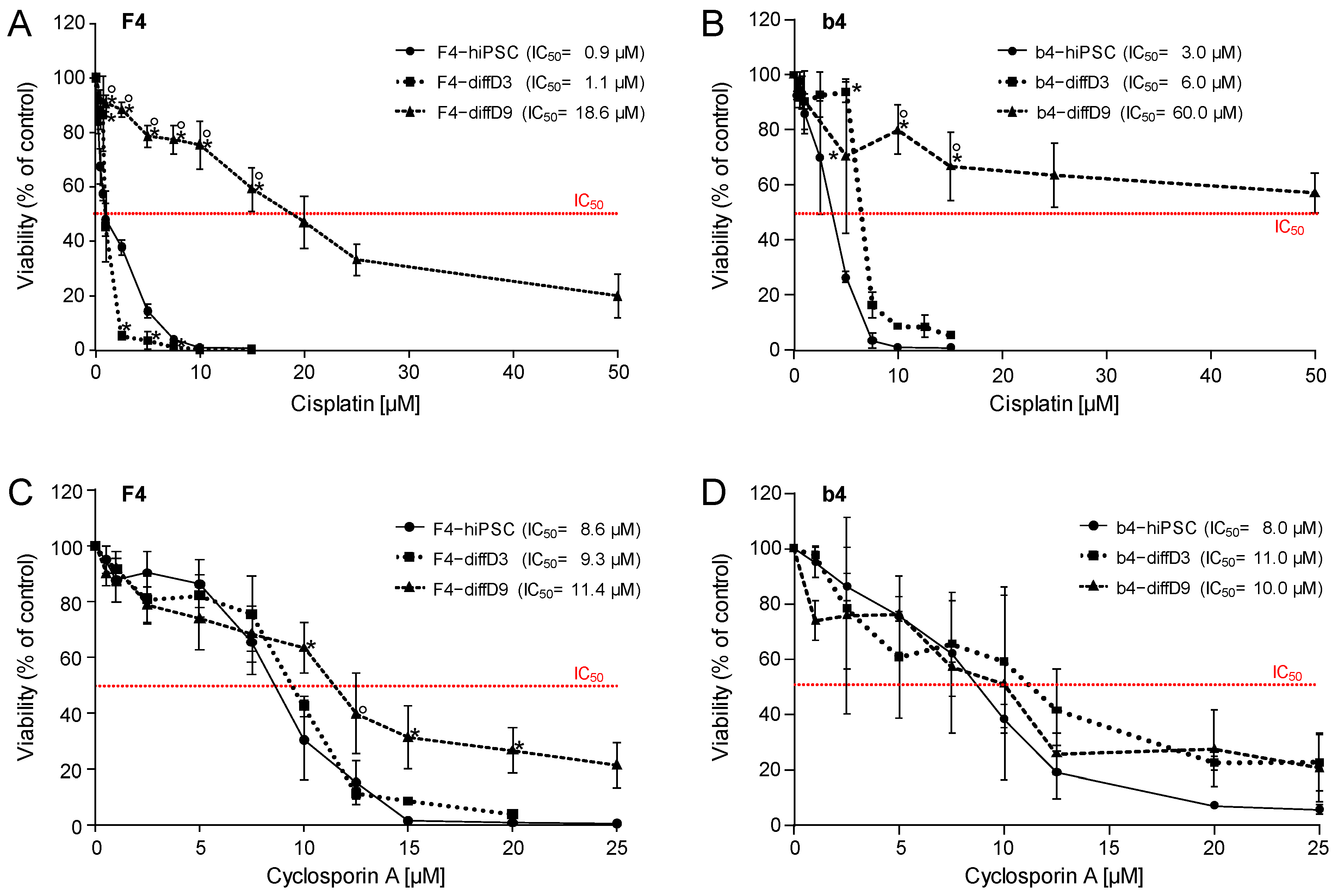
2.4. Sensitivity of hiPSC, hiPSC Differentiating into PTELC, and hiPSC Differentiated into PTELC towards a Genotoxic Nephrotoxin and a Non-Genotoxic Nephrotoxin
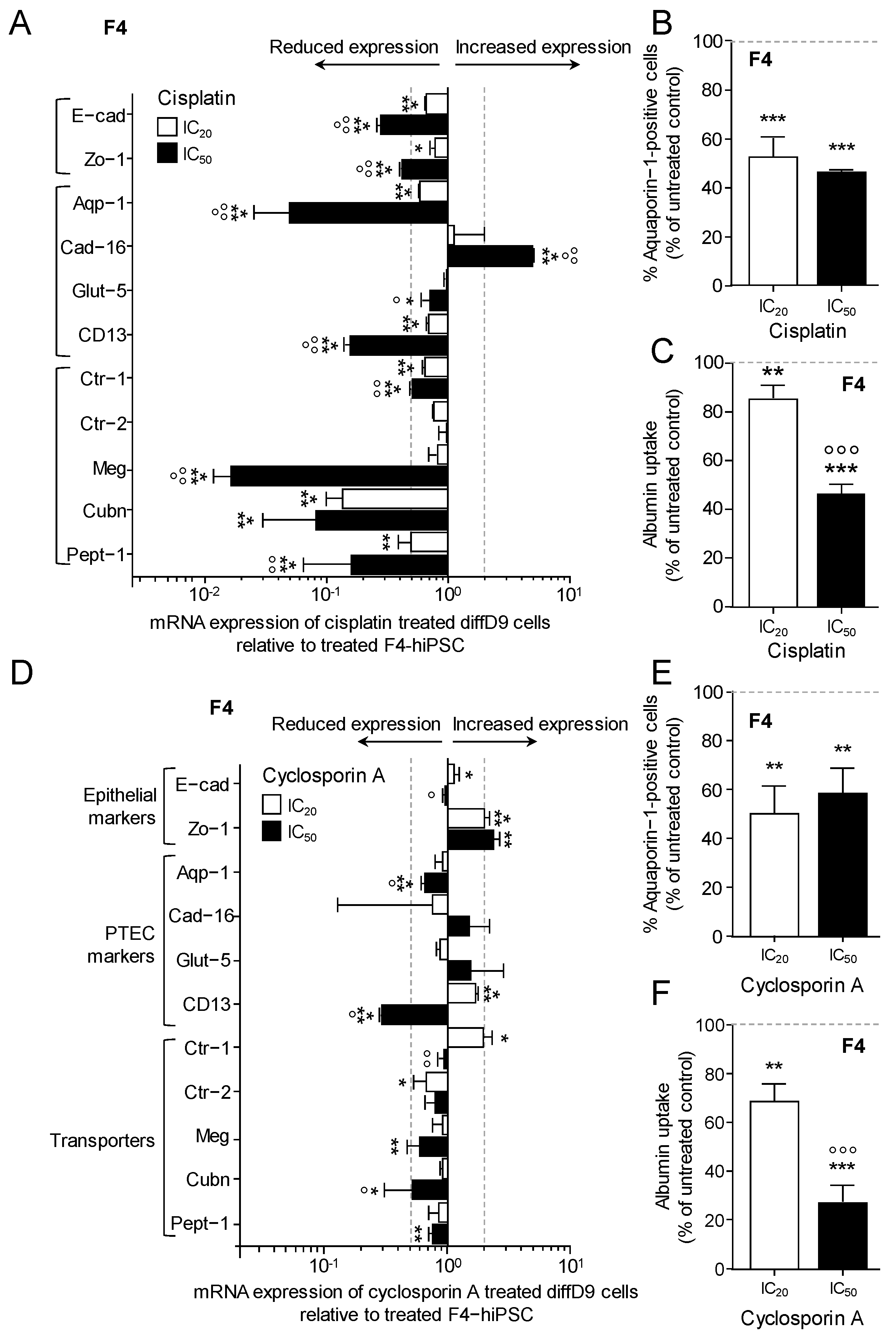
2.5. Impact of Toxin Treatment on Marker Expression and Transporter Function
3. Discussion
Limitations
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Cell Culture of RPTEC, F4-and b4-hiPSC, and In Vitro Differentiation Protocol
4.2.2. Analysis of Cell Viability
4.2.3. Analysis of Gene Expression (RT-PCR)
4.2.4. Immunofluorescence Analysis
4.2.5. Western Blot Analysis
4.2.6. Analysis of Cell Proliferation by Measuring EdU Incorporation
4.2.7. Flow Cytometry-Based Quantitative Detection of Aquaporin-1
4.2.8. Albumin Uptake Assay
4.2.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiong, H.Y.; Huang, P.; Xiong, S.; Li, Y.; Vathsala, A.; Zink, D. Drug-induced nephrotoxicity: Clinical impact and preclinical in vitro models. Mol. Pharm. 2014, 11, 1933–1948. [Google Scholar] [CrossRef] [PubMed]
- Nieskens, T.T.G.; Sjogren, A.K. Emerging In Vitro Systems to Screen and Predict Drug-Induced Kidney Toxicity. Semin. Nephrol. 2019, 39, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Naughton, C.A. Drug-induced nephrotoxicity. Am. Fam. Phys. 2008, 78, 743–750. [Google Scholar]
- Ameku, T.; Taura, D.; Sone, M.; Numata, T.; Nakamura, M.; Shiota, F.; Toyoda, T.; Matsui, S.; Araoka, T.; Yasuno, T.; et al. Identification of MMP1 as a novel risk factor for intracranial aneurysms in ADPKD using iPSC models. Sci. Rep. 2016, 6, 30013. [Google Scholar] [CrossRef] [PubMed]
- Berger, K.; Moeller, M.J. Mechanisms of epithelial repair and regeneration after acute kidney injury. Semin. Nephrol. 2014, 34, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Rangaswamy, D.; Sud, K. Acute kidney injury and disease: Long-term consequences and management. Nephrology (Carlton) 2018, 23, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.L.; Jaber, B.L.; Faubel, S.; Chawla, L.S.; Acute Kidney Injury Advisory Group of American Society of Nephrology. AKI transition of care: A potential opportunity to detect and prevent CKD. Clin. J. Am. Soc. Nephrol. 2013, 8, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Zink, D.; Chuah, J.K.C.; Ying, J.Y. Assessing Toxicity with Human Cell-Based In Vitro Methods. Trends Mol. Med. 2020, 26, 570–582. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Vinken, M.; Benfenati, E.; Busquet, F.; Castell, J.; Clevert, D.A.; de Kok, T.M.; Dirven, H.; Fritsche, E.; Geris, L.; Gozalbes, R.; et al. Safer chemicals using less animals: Kick-off of the European ONTOX project. Toxicology 2021, 458, 152846. [Google Scholar] [CrossRef]
- Chu, X.; Bleasby, K.; Evers, R. Species differences in drug transporters and implications for translating preclinical findings to humans. Expert. Opin. Drug Metab. Toxicol. 2013, 9, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Stecula, A.; Gupta, A.; Prasad, B.; Chien, H.C.; Yee, S.W.; Wang, L.; Unadkat, J.D.; Stahl, S.H.; Fenner, K.S.; et al. Molecular Mechanisms for Species Differences in Organic Anion Transporter 1, OAT1: Implications for Renal Drug Toxicity. Mol. Pharmacol. 2018, 94, 689–699. [Google Scholar] [CrossRef]
- Kandasamy, K.; Chuah, J.K.; Su, R.; Huang, P.; Eng, K.G.; Xiong, S.; Li, Y.; Chia, C.S.; Loo, L.H.; Zink, D. Prediction of drug-induced nephrotoxicity and injury mechanisms with human induced pluripotent stem cell-derived cells and machine learning methods. Sci. Rep. 2015, 5, 12337. [Google Scholar] [CrossRef] [PubMed]
- Morizane, R.; Miyoshi, T.; Bonventre, J.V. Concise Review: Kidney Generation with Human Pluripotent Stem Cells. Stem Cells 2017, 35, 2209–2217. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Rodrigues, A.D.; Steppan, C.M.; Engle, S.J.; Mathialagan, S.; Schroeter, T. Human Pluripotent Stem Cell-Derived Kidney Model for Nephrotoxicity Studies. Drug Metab. Dispos. 2018, 46, 1703–1711. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, V.; Carta, G.; da Costa Pereira, D.; Gupta, R.; Murphy, C.; Feifel, E.; Kern, G.; Lechner, J.; Cavallo, A.L.; Gupta, S.; et al. Generation and characterization of iPSC-derived renal proximal tubule-like cells with extended stability. Sci. Rep. 2021, 11, 11575. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.L.; Elhendawi, M.; Morlock, M.; Liu, W.; Liu, S.; Palakkan, A.; Seidl, L.F.; Hohenstein, P.; Sjogren, A.K.; Davies, J.A. Human iPSC-derived renal organoids engineered to report oxidative stress can predict drug-induced toxicity. iScience 2022, 25, 103884. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.T.T.; Rossbach, B.; Sebastien, I.; Neubauer, J.C.; Kurtz, A.; Hariharan, K. Functional differentiation and scalable production of renal proximal tubular epithelial cells from human pluripotent stem cells in a dynamic culture system. Cell Prolif. 2022, 55, e13190. [Google Scholar] [CrossRef]
- Valentich, J.D.; Tchao, R.; Leighton, J. Hemicyst formation stimulated by cyclic AMP in dog kidney cell line MDCK. J. Cell Physiol. 1979, 100, 291–304. [Google Scholar] [CrossRef]
- Wieser, M.; Stadler, G.; Jennings, P.; Streubel, B.; Pfaller, W.; Ambros, P.; Riedl, C.; Katinger, H.; Grillari, J.; Grillari-Voglauer, R. hTERT alone immortalizes epithelial cells of renal proximal tubules without changing their functional characteristics. Am. J. Physiol. Renal Physiol. 2008, 295, F1365–F1375. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, H.; Liu, F.; Dong, Z. Mitochondrial dysregulation and protection in cisplatin nephrotoxicity. Arch. Toxicol. 2014, 88, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Fukusumi, H.; Handa, Y.; Shofuda, T.; Kanemura, Y. Evaluation of the susceptibility of neurons and neural stem/progenitor cells derived from human induced pluripotent stem cells to anticancer drugs. J. Pharmacol. Sci. 2019, 140, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Peskova, L.; Vinarsky, V.; Barta, T.; Hampl, A. Human Embryonic Stem Cells Acquire Responsiveness to TRAIL upon Exposure to Cisplatin. Stem Cells Int. 2019, 2019, 4279481. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, P.; Di Serafino, A.; Sorino, L.; Ballerini, P.; Marchisio, M.; Pierdomenico, L.; Stuppia, L.; Antonucci, I. Genetic and epigenetic modifications induced by chemotherapeutic drugs: Human amniotic fluid stem cells as an in-vitro model. BMC Med. Genom. 2019, 12, 146. [Google Scholar] [CrossRef] [PubMed]
- Wing, C.; Komatsu, M.; Delaney, S.M.; Krause, M.; Wheeler, H.E.; Dolan, M.E. Application of stem cell derived neuronal cells to evaluate neurotoxic chemotherapy. Stem Cell Res. 2017, 22, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Tsui, A.Y.P.; Li, Z.; Zhang, Y.; Zhao, Q.; Xing, H.; Wang, X. Mechanisms of peripheral neurotoxicity associated with four chemotherapy drugs using human induced pluripotent stem cell-derived peripheral neurons. Toxicol. In Vitro 2021, 77, 105233. [Google Scholar] [CrossRef]
- Guo, H.; Deng, N.; Dou, L.; Ding, H.; Criswell, T.; Atala, A.; Furdui, C.M.; Zhang, Y. 3-D Human Renal Tubular Organoids Generated from Urine-Derived Stem Cells for Nephrotoxicity Screening. ACS Biomater. Sci. Eng. 2020, 6, 6701–6709. [Google Scholar] [CrossRef]
- Antonios, J.P.; Farah, G.J.; Cleary, D.R.; Martin, J.R.; Ciacci, J.D.; Pham, M.H. Immunosuppressive mechanisms for stem cell transplant survival in spinal cord injury. Neurosurg. Focus. 2019, 46, E9. [Google Scholar] [CrossRef]
- Wellens, S.; Dehouck, L.; Chandrasekaran, V.; Singh, P.; Loiola, R.A.; Sevin, E.; Exner, T.; Jennings, P.; Gosselet, F.; Culot, M. Evaluation of a human iPSC-derived BBB model for repeated dose toxicity testing with cyclosporine A as model compound. Toxicol. In Vitro 2021, 73, 105112. [Google Scholar] [CrossRef]
- Schultze, N.; Wanka, H.; Zwicker, P.; Lindequist, U.; Haertel, B. Mitochondrial functions of THP-1 monocytes following the exposure to selected natural compounds. Toxicology 2017, 377, 57–63. [Google Scholar] [CrossRef]
- de Arriba, G.; de Hornedo, J.P.; Rubio, S.R.; Fernandez, M.C.; Martinez, S.B.; Camarero, M.M.; Cid, T.P. Vitamin E protects against the mitochondrial damage caused by cyclosporin A in LLC-PK1 cells. Toxicol. Appl. Pharmacol. 2009, 239, 241–250. [Google Scholar] [CrossRef]
- Nagavally, R.R.; Sunilkumar, S.; Akhtar, M.; Trombetta, L.D.; Ford, S.M. Chrysin Ameliorates Cyclosporine-A-Induced Renal Fibrosis by Inhibiting TGF-beta1-Induced Epithelial-Mesenchymal Transition. Int. J. Mol. Sci. 2021, 22, 10252. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Xie, G.; Cui, H.; Yao, Z.; Shao, C.; Yuan, W.; Chen, B. Cyclosporin-A reduced the cytotoxicity of propranolol in HUVECs via p38 MAPK signaling. Medicine 2022, 101, e28329. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, N.; Jin, L.; Qi, X.; Zhang, C.; Hua, D. The calcium pump PMCA4 prevents epithelial-mesenchymal transition by inhibiting NFATc1-ZEB1 pathway in gastric cancer. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118833. [Google Scholar] [CrossRef] [PubMed]
- Hori, Y.; Aoki, N.; Kuwahara, S.; Hosojima, M.; Kaseda, R.; Goto, S.; Iida, T.; De, S.; Kabasawa, H.; Kaneko, R.; et al. Megalin Blockade with Cilastatin Suppresses Drug-Induced Nephrotoxicity. J. Am. Soc. Nephrol. 2017, 28, 1783–1791. [Google Scholar] [CrossRef] [PubMed]
- Mahadevappa, R.; Nielsen, R.; Christensen, E.I.; Birn, H. Megalin in acute kidney injury: Foe and friend. Am. J. Physiol. Renal Physiol. 2014, 306, F147–F154. [Google Scholar] [CrossRef]
- Arjumand, W.; Seth, A.; Sultana, S. Rutin attenuates cisplatin induced renal inflammation and apoptosis by reducing NFkappaB, TNF-alpha and caspase-3 expression in wistar rats. Food Chem. Toxicol. 2011, 49, 2013–2021. [Google Scholar] [CrossRef] [PubMed]
- Kishore, B.K.; Krane, C.M.; Di Iulio, D.; Menon, A.G.; Cacini, W. Expression of renal aquaporins 1, 2, and 3 in a rat model of cisplatin-induced polyuria. Kidney Int. 2000, 58, 701–711. [Google Scholar] [CrossRef]
- Afjal, M.A.; Goswami, P.; Ahmad, S.; Dabeer, S.; Akhter, J.; Salman, M.; Mangla, A.; Raisuddin, S. Tempol (4-hydroxy tempo) protects mice from cisplatin-induced acute kidney injury via modulation of expression of aquaporins and kidney injury molecule-1. Drug Chem. Toxicol. 2022, 45, 1355–1363. [Google Scholar] [CrossRef]
- Shitara, Y.; Itoh, T.; Sato, H.; Li, A.P.; Sugiyama, Y. Inhibition of transporter-mediated hepatic uptake as a mechanism for drug-drug interaction between cerivastatin and cyclosporin A. J. Pharmacol. Exp. Ther. 2003, 304, 610–616. [Google Scholar] [CrossRef]
- Nascimento, C.R.; Braga, F.; Capella, L.S.; Santos, O.R.; Lopes, A.G.; Capella, M.A. Comparative study on the effects of cyclosporin a in renal cells in culture. Nephron Exp. Nephrol. 2005, 99, e77–e86. [Google Scholar] [CrossRef]
- Motohashi, H.; Katsura, T.; Saito, H.; Inui, K. Effects of tacrolimus and cyclosporin A on peptide transporter PEPT1 in Caco-2 cells. Pharm. Res. 2001, 18, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Shitara, Y.; Takeuchi, K.; Nagamatsu, Y.; Wada, S.; Sugiyama, Y.; Horie, T. Long-lasting inhibitory effects of cyclosporin A, but not tacrolimus, on OATP1B1- and OATP1B3-mediated uptake. Drug Metab. Pharmacokinet. 2012, 27, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.W.; Ahn, K.O.; Sheen, M.R.; Jeon, U.S.; Kim, J.; Yang, C.W.; Kwon, H.M. Downregulation of renal sodium transporters and tonicity-responsive enhancer binding protein by long-term treatment with cyclosporin A. J. Am. Soc. Nephrol. 2007, 18, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Edemir, B.; Reuter, S.; Borgulya, R.; Schroter, R.; Neugebauer, U.; Gabriels, G.; Schlatter, E. Acute rejection modulates gene expression in the collecting duct. J. Am. Soc. Nephrol. 2008, 19, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Capolongo, G.; Damiano, S.; Suzumoto, Y.; Zacchia, M.; Rizzo, M.; Zona, E.; Pollastro, R.M.; Simeoni, M.; Ciarcia, R.; Trepiccione, F.; et al. Cyclosporin-induced hypertension is associated with the up-regulation of Na+-K+-2Cl− cotransporter (NKCC2). Nephrol. Dial. Transplant. 2023, gfad161. [Google Scholar] [CrossRef] [PubMed]
- Secker, P.F.; Schlichenmaier, N.; Beilmann, M.; Deschl, U.; Dietrich, D.R. Functional transepithelial transport measurements to detect nephrotoxicity in vitro using the RPTEC/TERT1 cell line. Arch. Toxicol. 2019, 93, 1965–1978. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Wang, Y.; Adjaye, J. A cyclic AMP analog, 8-Br-cAMP, enhances the induction of pluripotency in human fibroblast cells. Stem Cell Rev. Rep. 2011, 7, 331–341. [Google Scholar] [CrossRef]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mboni-Johnston, I.M.; Kouidrat, N.M.Z.; Hirsch, C.; Weber, A.G.; Meißner, A.; Adjaye, J.; Schupp, N. Sensitivity of Human Induced Pluripotent Stem Cells and Thereof Differentiated Kidney Proximal Tubular Cells towards Selected Nephrotoxins. Int. J. Mol. Sci. 2024, 25, 81. https://doi.org/10.3390/ijms25010081
Mboni-Johnston IM, Kouidrat NMZ, Hirsch C, Weber AG, Meißner A, Adjaye J, Schupp N. Sensitivity of Human Induced Pluripotent Stem Cells and Thereof Differentiated Kidney Proximal Tubular Cells towards Selected Nephrotoxins. International Journal of Molecular Sciences. 2024; 25(1):81. https://doi.org/10.3390/ijms25010081
Chicago/Turabian StyleMboni-Johnston, Isaac Musong, Nazih Mohamed Zakari Kouidrat, Cornelia Hirsch, Andreas Georg Weber, Alexander Meißner, James Adjaye, and Nicole Schupp. 2024. "Sensitivity of Human Induced Pluripotent Stem Cells and Thereof Differentiated Kidney Proximal Tubular Cells towards Selected Nephrotoxins" International Journal of Molecular Sciences 25, no. 1: 81. https://doi.org/10.3390/ijms25010081
APA StyleMboni-Johnston, I. M., Kouidrat, N. M. Z., Hirsch, C., Weber, A. G., Meißner, A., Adjaye, J., & Schupp, N. (2024). Sensitivity of Human Induced Pluripotent Stem Cells and Thereof Differentiated Kidney Proximal Tubular Cells towards Selected Nephrotoxins. International Journal of Molecular Sciences, 25(1), 81. https://doi.org/10.3390/ijms25010081