1. Introduction
Central nervous system (CNS) cancers are rare and heterogeneous tumors with diverse biologies and genetics, which account for about 3% of all cancers in the world and are more common in men [
1]. Gliomas constitute the most common type of brain tumors, comprising around 24.7% of all primary tumors in the brain and other CNS tumors, and 74.6% of malignant cases [
2]. The most prevalent malignant histology is the glioblastoma, making up 14.2% of all tumors and constituting 50.1% of all malignant CNS tumors [
3]. The new 2021 WHO CNS 5 classification has divided diffuse gliomas into adult-type and pediatric-type. The basic molecules for the integrated diagnosis of adult diffuse gliomas are
IDH (Isocitrate dehydrogenase),
p53,
ATRX (alpha-thalassemia/mental retardation, X-linked), and
1p/19q co-deletion. Adult diffuse gliomas (IDH-mutant astrocytomas, IDH-mutant, 1p/19q co-deleted oligodendroglioma, and IDH-wildtype glioblastoma) are diffusely infiltrating brain tumors [
4]. High-grade gliomas, predominantly wild-type glioblastomas, are extremely lethal neoplasms with a poor prognosis. Despite maximum neurosurgical resection and adjuvant therapy; temozolamide (TMZ), median survival barely extends to approximately 12 months [
5]. Unfortunately, as with other solid tumors, chemo-resistance is one of the major challenges in this regard. Various mechanisms have been described for chemo-resistance. The evasion of apoptosis is one of the mechanisms of tumor progression and chemo-resistance in gliomas [
6], probably through either deficiency in BAX or BAK or a gain of Bcl2 or BclX [
7]. Some studies have also revealed that the resistance of cancer cells to chemotherapeutic drugs may be due to the up-regulation of autophagy, thereby avoiding apoptosis [
8,
9].
Autophagy is a cellular degradation pathway for the breakdown and removal of impaired long-lived proteins, as well as the elimination of organelles and pathogens [
10]. Moreover, it has an important homeostatic role in maintaining cell viability in stressed or nutritionally deprived states through recycling the cytoplasmic constituent [
11]. Thus, it controls the quality and quantity of proteins and organelles. Dysfunctional autophagy contributes to various diseases—cancer being one of them. Nevertheless, in cancer, the role of autophagy is somewhat complicated and controversial. Autophagy is a double-edged sword as it has dual effects on cancer. On the one hand, it promotes tumor cell survival by breaking down macromolecules into smaller components like amino acids, fatty acids, and metabolic substances. Studies have suggested that autophagy is activated in glioblastoma (GBM) as a reaction to pathophysiological challenges like necrosis and an acidic milieu. Hypoxia, a characteristic feature of high-grade gliomas, is responsible for inducing autophagy in these tumors. As the tumor progresses and reacts to therapeutic interventions, cells must adjust their metabolism to endure in hypoxic and nutrient-deficient surroundings; this adjustment is commonly linked to chemotherapy and radiotherapy resistance. This process facilitates tumor growth and viability, contributing to the maintenance of intracellular metabolism. Under unfavorable hypoxic conditions, molecules such as hypoxia-induced factors (HIF2a and HIF2b), BECN1 (Beclin-1), and BNIP3 (BCL2/adenovirus E1B 19KDa interacting protein 2) serve as survival mechanisms, promoting the progression of GBM and its resistance to anticancer treatmentin vivo [
12,
13,
14,
15]. Hypoxia causes BECN1 phosphorylation via the HIF-1a/BECN1 signaling pathway [
16,
17]. Beclin-1 is a key regulatory protein in autophagosome formation, which binds to the class III PI–3 kinase Vps34, thus facilitating progression of the autophagic process [
18,
19]. Remarkably, in the presence of hypoxia, the initiation of autophagy through BNIP3/BNIP3L serves as a survival mechanism, fostering the progression of GBM and enhancing its resistance to anticancer treatments in vivo [
20]. A recent study showed that in U87 cells exposed to hypoxic conditions, the knockdown of HIF1A significantly reduced BNIP3 expression [
21]. This implies that tissues with low oxygen levels are prone to show heightened autophagy, indicating that an increased autophagy flux may serve an adaptive function [
15,
22]. On the other hand, autophagy can also suppress tumor progression in development, as impaired autophagy causes oxidative stress, triggers DNA damage responses, and results in genomic instability, a known cause of tumor initiation. Such dual effects on tumor behavior are most likely context-dependent [
23,
24,
25,
26]. Moreover, research has indicated that prolonged hypoxic stress in vitro up-regulates a pro-apoptotic Bcl-2 protein (BNIP3), leading to hypoxia-dependent autophagic cell death (ACD) in GBM cells [
27]. In terms of this mechanism, the elevated expression of BNIP3 leads to the release of BECN1 from complexes with Bcl-2 or Bcl-xL, thereby enabling BECN1 to activate autophagy [
28]. Hence, autophagy has been found to show conflicting functions in tumor onset and advancement. Thus, the manipulation of autophagy pathways as a means of cell death in cancer has led to the exploration of both inhibitors and inducers. Keeping in mind the contradictory results of autophagy in cancer, we investigated the expression of autophagy markers and autophagy-related genes (ATGs) in our cohort of low- and high-grade gliomas.
3. Discussion
Autophagy is primarily a stress response process, and most of body tissues need autophagy to eliminate accumulated damaged organelles and unfolded proteins to maintain normal homeostatic milieu. In cancer, autophagy plays both tumor-suppressing and tumor-promoting roles depending on the specific context; thus, it could influence the prognosis either favorably or adversely. We investigated whether autophagy plays anti-tumoral or pro-tumoral functions in various grades of gliomas.
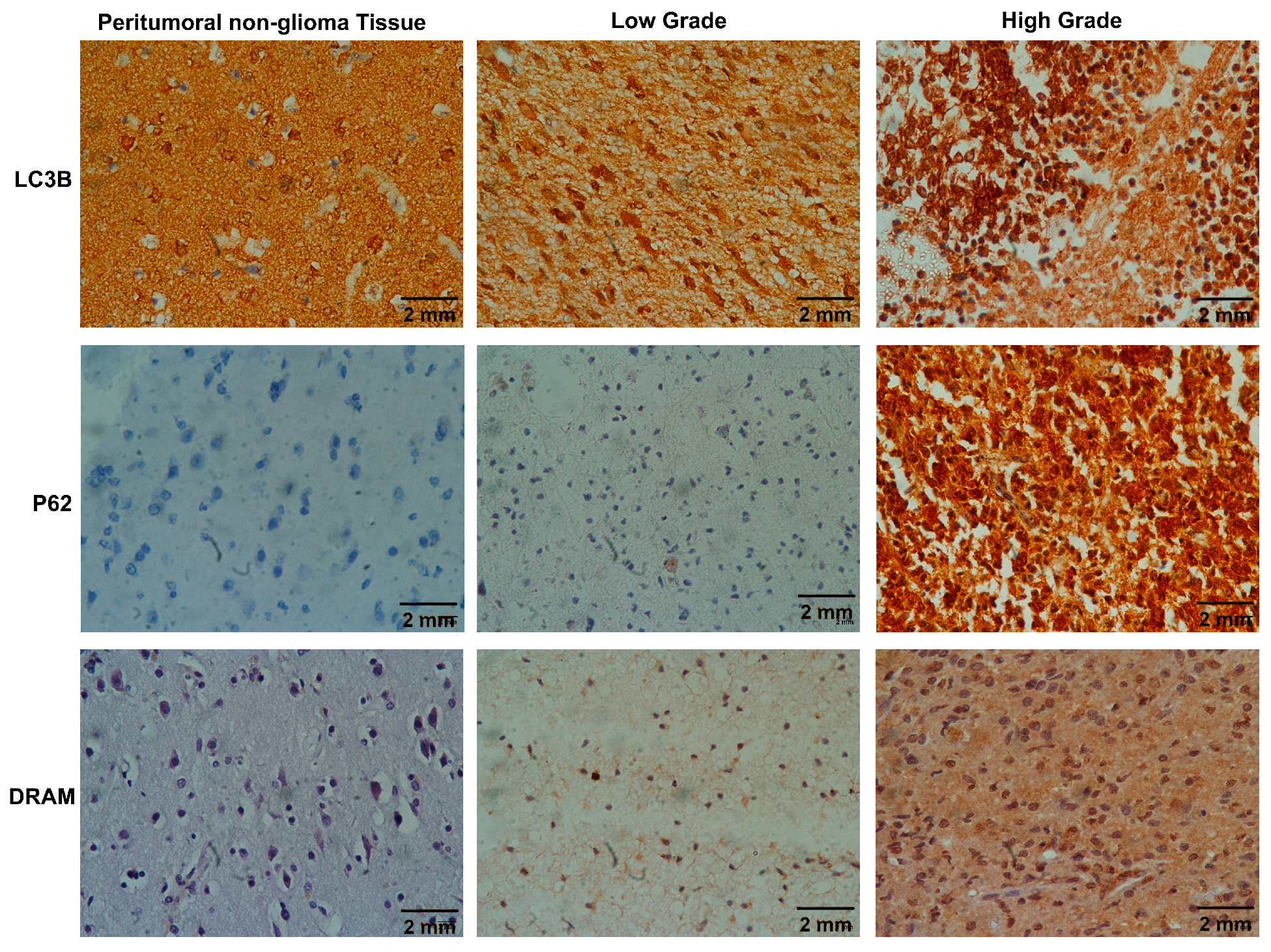
In this current research, we assessed the immunohistochemical expression of the autophagy markers and the mRNA levels of ATGs in different grades of adult-type diffuse gliomas while also assessing their correlation with clinicopathological parameters. Our investigation revealed a significant connection between the immunohistochemical expression of
LC3B,
SQSTM1/p62, and
DRAM, and the overall autophagy status, particularly tumor grades. Notably, among the 25 positive cases of
LC3B, only two were observed in the LGGs. Similarly, out of the 20 strongly positive cases of
SQSTM1/p62, none were found in the LGG category, and this difference achieved statistical significance (as indicated in
Table 3). These results are in accordance with other studies, which reported that
LC3B and
SQSTM1/p62 were highly expressed in high-grade gliomas [
29,
30,
31]. Remarkably, there is a statistically significant association between the autophagy markers
LC3B and
SQSTM1/p62 (
p = 0.000), highlighting the interaction between them. These results may be attributed to the active participation of
LC3B and
SQSTM1/p62 in the process of autophagy, given their roles as essential structural components of the autophagosome [
32], and recent literature also revealed that
SQSTM1/p62 overexpression is capable of promoting mitochondrial and classical macroautophagy [
33], which promote tumor progression and chemo-resistance. Furthermore, increased expression of
LC3B and
SQSTM1/p62 in HGGs compared with LGGs may be in response to conferring stress tolerance, which is greater in HGGs, and serves to maintain tumor cell survival [
34]. Hypoxia, which is a major hallmark of glioblastoma, is known to induce autophagy in these tumors, which eventually produces a survival mechanism facilitating the breakdown of various cellular components to generate ATP and metabolic precursors to cope with stress, highlighting the crucial role of autophagy in protecting cells against stressful conditions [
35]. According to a study conducted by Deng et al., both the mRNA and protein levels of SQSTM1/p62 were found to be elevated in human glioma tissues. Furthermore, it was discovered that the suppression of SQSTM1/p62 had an anti-tumor effect on glioma cells [
34]. These findings suggest that high levels of LC3B and SQSTM1/p62 expression, or prompted autophagy, are correlated with advanced tumor grade and aggressiveness. Additionally, these findings support the potential role of autophagy as a tumor enhancer, which further supports the results of the study.
We also looked at another important autophagy modulator,
DRAM1, in our study and found its overexpression in 28.2% of the cases; it was not significantly associated with tumor grades, but higher expression of
DRAM1 was present in high-grade gliomas when compared with LGGs.
DRAM1 primarily localizes to lysosomes and is frequently downregulated in various human cancers [
36].
DRAM1 enhances lysosomal acidification and facilitates the fusion of lysosomes with autophagosomes, thereby promoting autophagy. Moreover,
DRAM1 plays a pivotal role in governing the association of
SQSTM1/p62 with autophagosomes and its subsequent degradation through autophagy. Therefore, reduced
DRAM1 expression might be associated with decreased
SQSTM1/p62 localization within autophagosome, highlighting the role of
DRAM1 in
SQSTM1/p62-mediated autophagy. Our observations are in line with the research conducted by Geng et al., which also indicated decreased DRAM1 expression in non-small-cell lung carcinoma, linked with an unfavorable prognosis [
37]. However, another study showed high expression of both
DRAM1 and
SQSTM1/p62 in glioblastoma, where they regulate cell migration and invasion and are associated with shorter or poor overall survival [
38]. These conflicting results could arise from variations in demographics and sample sizes. Thus, utilizing the expression levels of
LC3B,
SQSTM1/p62, and
DRAM1 to assess autophagy status in glioma patients could be potential predictive markers.
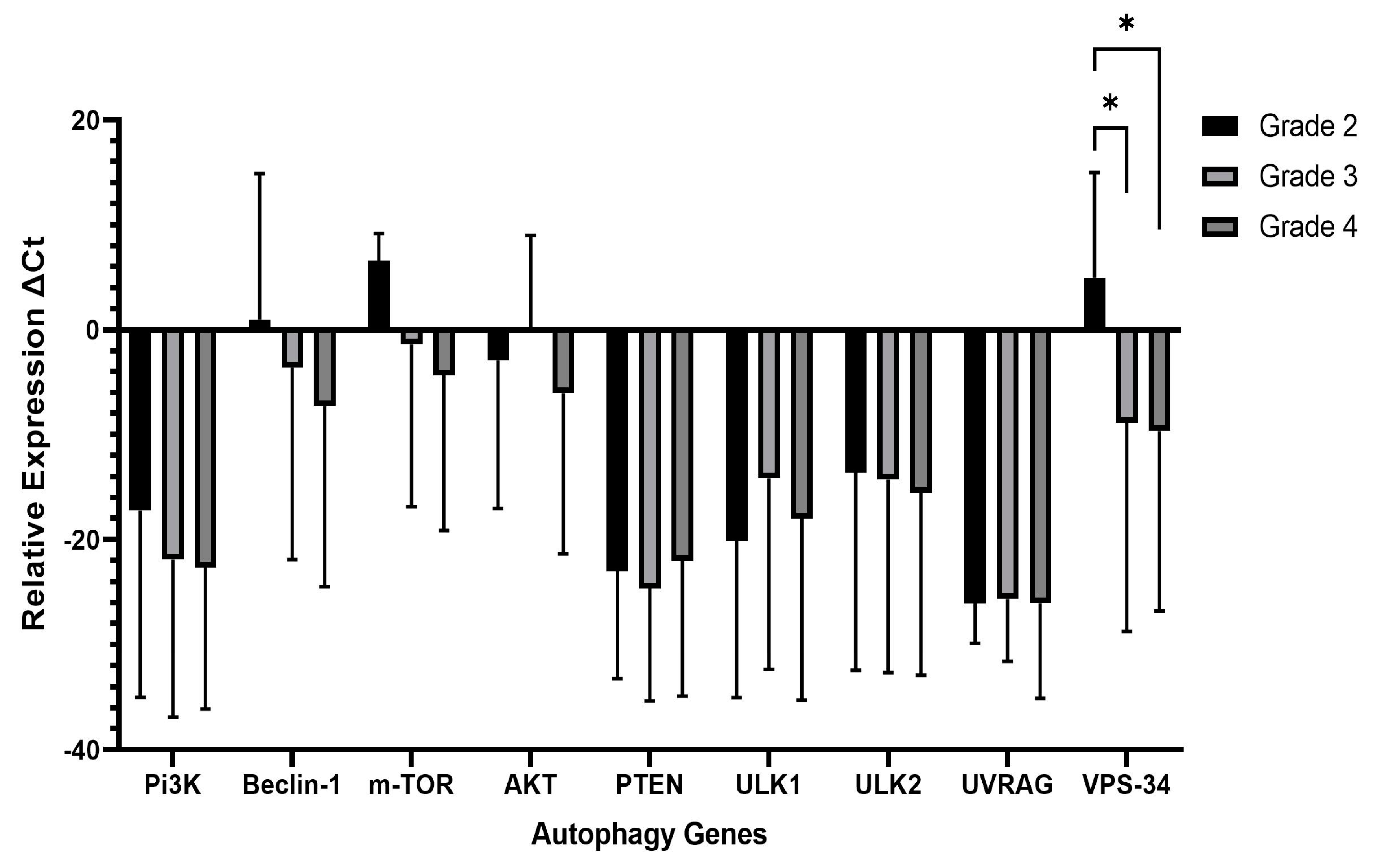
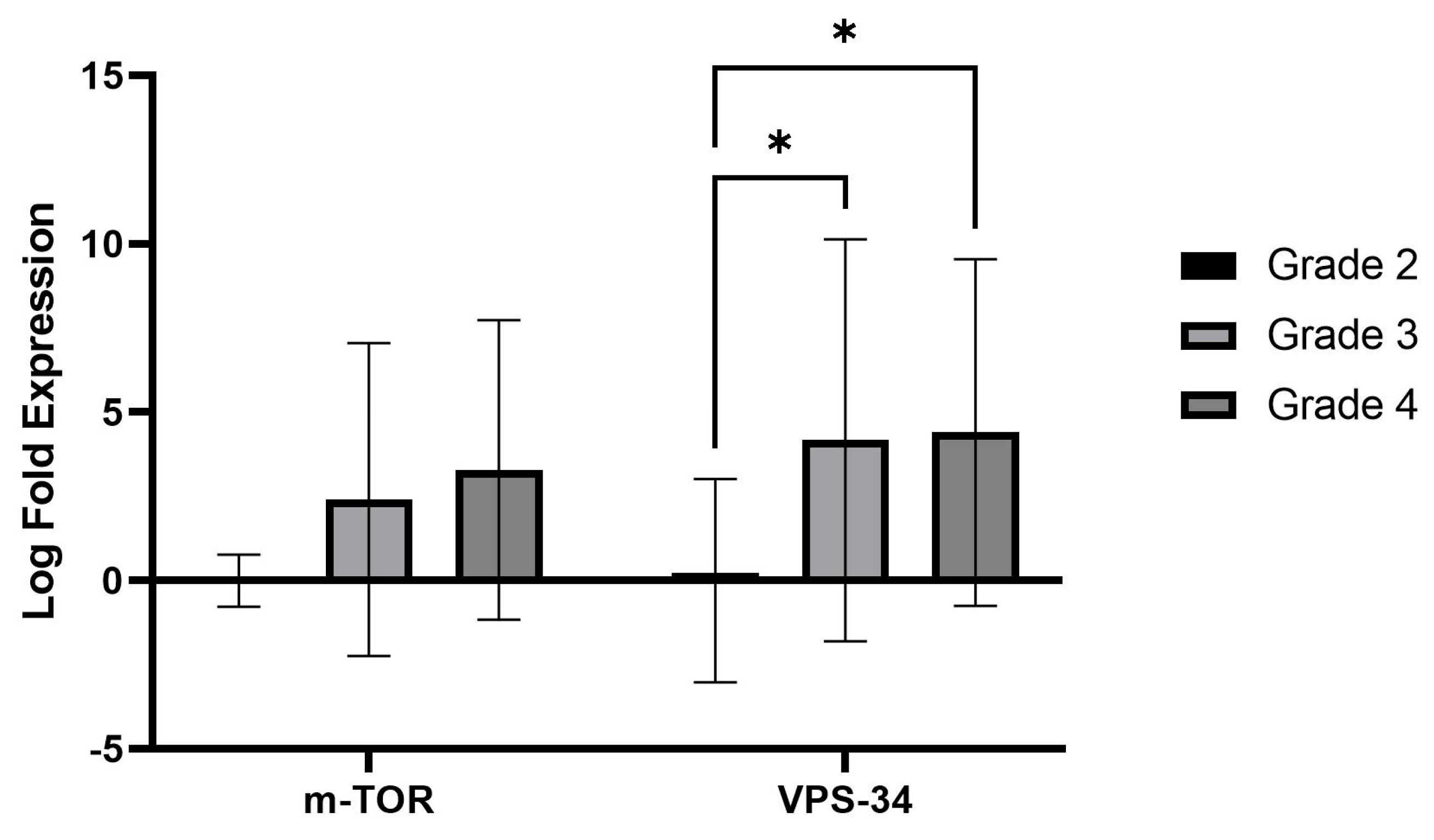
While investigating the autophagy gene expression patterns in both LGGs (grade 2) and HGGs (grades 3 and 4), we observed a remarkable and statistically significant up-regulation in the expression of vacuolar protein sorting 34 (
Vps34), a critical kinase in autophagy. This indicates that the VPS34 gene plays a role in promoting autophagy in gliomas. Previous research has shown that
VPS34 initiates autophagy by interacting with Vps15/Atg14/UVRAG/Beclin1 [
39]. Therefore, inhibiting VPS34 has been considered as a potential target for inhibiting autophagy [
40,
41].
We also evaluated the autophagy upstream pathway,
PI3K/AKT/mTOR. We recorded higher expression of
mTOR transcripts in HGGs than LGGs, a negative autophagy regulator, but it did not reach a significant value (
p = 0.07). The
PI3K/Akt/mTOR signaling pathway is a frequently disrupted pathway across different cancer types [
42,
43], and abnormal activation of this pathway has been associated with tumor development, progression, invasion, and metastasis [
44] and is indeed activated in glioma cells [
45]. Nonetheless, the
PTEN,
PI3K,
AKT,
mTOR,
Beclin-1,
UVRAG,
ULK1, and
ULK2 genes remained unchanged when compared in both grades. The significant increase in transcriptional expression of VPS34 implies an increased initiation of autophagy, as VPS34 plays a crucial role in the nucleation of autophagosomes. Conversely, the absence of substantial transcriptional alterations in other genes suggests that these components may not be profoundly influenced at the transcriptional level in high-grade gliomas. Nevertheless, it is important to consider the potential involvement of post-transcriptional, post-translational, or alternative regulatory mechanisms. The modulation of autophagy in high-grade gliomas depends on the unique context of these tumors, which are heterogeneous and have diverse genetic and epigenetic profiles. Therefore, autophagy signaling pathways may vary among individual tumors.
Spearman’s correlation between autophagy genes was examined in grades 2, 3, and 4 of glioma cases, and we found a significant positive correlation among different genes. It is essential to highlight that these observed correlations were significant, but whether they have a biological basis remains uncertain. For instance, in grade 2 gliomas,
PTEN showed a strong positive correlation with autophagy genes, which aligns with a study by Errafiy Rajaa [
46]. However, the current study’s absence of
PTEN correlation in grade 4 highlights the loss of
PTEN, a hallmark of GBM, and could be due to mutation or promoter methylation of
the PTEN gene [
47,
48].
Similarly, in HGGs, the significant positive correlation between
VPS34 and
ULK1,
ULK2,
UVRAG,
Beclin1, and
PI3K genes can also be justified biologically, as the literature reports that the
VPS34 kinase forms a stable complex with Beclin1 and p150, serving as a binding partner for
ATG14L,
UVRAG, and
AMBRA [
49], which are responsible for the promotion of autophagy. Thus, a significant positive correlation indicates that the expression of autophagy genes tends to change consistently, providing insights into the regulatory connections within autophagic pathways. This understanding may have implications for comprehending the underlying mechanisms influencing tumor progression.
Nevertheless, a limitation of our study is the absence of autophagy flux assessment in formalin-fixed, paraffin-embedded tissue block (FFPE) samples. Elevated expression levels of LC3B and SQSTM1/p62 do not consistently correlate with an overall augmentation in autophagy. They may be attributed to a potential hindrance in autophagy at the later stages of autophagosome processing. Thus, there is a need for a more comprehensive assessment of autophagy, particularly considering the dynamic nature of the process. Moreover, ATG5, ATG12, ATG 7, and ATG 4 are crucial for two conjugation systems (Atg8–Atg4 and Atg12–Atg5) involved in autophagy and would provide valuable information, as these processes contribute to the formation and elongation of autophagosomes, facilitating the degradation and recycling of cellular components. Unfortunately, due to financial constraints, we could not perform this analysis. However, future comprehensive studies utilizing diverse techniques will be invaluable in addressing these limitations.
4. Materials and Methods
4.1. Patients Selection
Patients with adult diffuse gliomas were enrolled from the Histopathology Department of the Dow Diagnostic Reference and Research Lab (DDRRL) at Dow University of Health Sciences (DUHS). The Institutional Review Board at Dow University of Health Sciences approved the research protocol (Ref: IRB-1150/DUHS/Approval/2018). Clinical information was recorded. The study included patients diagnosed with adult diffuse gliomas of all grades. A total of 50 adult diffuse gliomas were initially included in the study, excluding patients diagnosed with pediatric-type diffuse gliomas and astrocytomas with circumscribed morphology. However, due to the use of multiple immunohistochemical stains and the extraction of nucleic acids from tissue samples, some cases ran out of available tissue. As a result, only 39 patients were finally included in the study. The study aimed to determine the mRNA and protein expression of autophagy-related genes (ATGs) using Quantitative real-time PCR (qPCR) and IHC.
4.2. Tissue Processing for Histopathological Analysis
The histopathology department received biopsy specimens of brain tumors from various hospitals and centers in the city through its collection point service. For histopathological examination, every specimen was placed in an automated tissue processor (Thermo Scientific, model EXCELSIOR AS) for 12 hours, where they were exposed to a graded series of alcohol at 70%, 95%, and 100%; followed by xylene; and then paraffin. The paraffin blocks obtained were sliced into sections measuring 4–5 μm thickness for subsequent hematoxylin and eosin (H&E) staining. The slides were deparaffinized before staining, and the sections were treated with xylene, followed by hydration through decreasing concentration of ethanol of 100%, 70%, and 50%. Slides were then cleaned with water before being submerged in H&E for staining. To eliminate excess water from the slide after staining, the sections were submerged in 60%, 80%, and 100% ethanol. Slides were covered by a coverslip with Enthelan® after being submerged in xylene (Merck, Darmstadt, Germany). The Nikon Eclipse E200 optical microscope (Nikon Instruments Inc. in Tokyo, Japan) was used to perform the histopathological examination of the H&E stained slides, and the tumors characterized as adult diffuse gliomas were enrolled in the study. The cases of diffuse gliomas were categorized according to CNS5 classification into distinct subtypes and malignancy grades using histopathological features such as cellularity, atypia, necrosis, micro-vascular proliferation, and mitosis.
4.3. Tissue Processing for Immunohistochemical Examination
IHC was carried out using the Autostainer Link 48 (Dako North America Inc., USA, S no AS3006D1307, Carpinteria, CA, USA) on formalin-fixed, paraffin-embedded (FFPE) tissue blocks for LC3B, p62, and DRAM antibodies. Sections were sliced into 3–4 μm, affixed on charged slides (EnVision FLEX visualization systems), and dried for 60 min at 60–70 °C. The slides were then deparaffinized with xylene, rinsed in decreasing ethanol concentrations, and finally rehydrated in distilled H2O. In a preheated water bath, retrieval solution (EnVision Flex Target Retrieval solution, pH 9.0, TRIS HCL) was used for 20 min to facilitate antigen unmasking. To quench endogenous peroxidase activity, slides were immersed in peroxidase blocking solution (EnVision Flex Peroxidase blocking reagent, RTU) for 10 min. TBST (Tris Buffer saline with Tween 20, EnVision Flex wash Buffer) was used for washing; then, sections were incubated for 30–60 min at room temperature with primary antibodies LC3B (ABCAM, ab51520: 1: 800), p62 (Invitrogen, Waltham, MA, USA, clone: SOSTM1: 1: 25), and DRAM (Invitrogen, 1; 25). Subsequently, the sections were subjected to a 30 min treatment at room temperature with a secondary antibody (EnVision Flex/HRP, RTU) after washing with PBS buffer. DAB solution 3, 3-diaminobenzidine tetrahydrochloride solution (EnVision Flex DAB+ chromogen) was applied for 10 min to the sections on the slides to reveal the color of antibody staining and counterstained with hematoxylin. Slides were washed for 10 min in running water. Sections were then dehydrated (in graded alcohol of 80%, 90%, and 100%), cleaned, and cover-slipped using a DPX mounting solution.
4.4. Immunohistochemical Evaluation of Autophagy Markers
LC3B and
SQSTM1/p62 punctate/dotted cytoplasmic staining were considered as positive [
50,
51]. The immunopositivity was scored based on the intensity and percentage of positive glioma cells. Immunopositivity of >50% of tumor cells was considered as positive for
LC3B, whereas for
SQSTM1/p62, it was considered positive when it exceeded >30% [
52]. The intensity of DRAM1 staining was scored on a scale of 0 to 3, where 0 means no staining, 1 means weak staining, 2 means moderate staining, and 3 means strong staining. Percentage scores were assigned on a scale of 1 to 4, where 1 means 0–25% staining, 2 means 26–50% staining, 3 means 51–75% staining, and 4 means 71–100% staining. Each sample was given a score, which was then multiplied to calculate a total value that ranged from 0 to 12. A score of 4 or more was defined as DRAM1 overexpression, while a score of less than 4 was classified as weak or negative expression [
53]. The immunohistochemical staining was independently scored by two pathologists at 40× objective magnification; then, discrepancies were discussed on a multi-head microscope and final scores were determined. Autophagy status was considered positive when two out of the three autophagy-associated proteins were detected in each sample [
54].
4.5. RNA Extraction and cDNA Synthesis
For the RT-qPCR analysis, total RNA was isolated from FFPE blocks via Pure Link FFPE, total isolation (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) according to the manufacturer’s protocols [
55]. Subsequently, DNase treatment was carried out for any DNA contamination. This involved combining 1 μg of RNA template with 1 μL of reaction buffer containing MgCl
2; 1 μL of DNase-I, RNase-free (Thermo Fisher Scientific, Cat. No. EN0521); and nuclease-free water in a 0.2 mL tube. The mixture was incubated in a Master cycler X50a (Eppendorf, Hamburg, Germany) for 30 min at 37 °C. To prevent RNA degradation, following the DNase-I treatment, we introduced 1 μL of 50 mM EDTA, and the samples were incubated at 65 °C for 10 min. The RNA integrity was evaluated with a nanodrop and, subsequently, cDNA was generated according to the manufacturer’s instructions using the Revert-Aid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Cat. No. K1612). The resulting cDNA was stored at −20 °C for future applications.
4.6. Gene Expression Analysis via Quantitative Real-Time PCR (qPCR)
qPCR was conducted to assess the gene expression levels of
ULK1,
ULK2,
Beclin 1,
UVRAG,
VPS34,
PTEN,
PI3K,
AKT, and
mTOR using a PCR kit (PowerUp™ SYBR™ Green Master Mix (Thermo Fisher Scientific, Cat. No A25742) and primers (Eurofins, Barberton, OH, USA). β-actin, a housekeeping gene, was used for result normalization in the qPCR assay, utilizing the corresponding primer sets. To perform qPCR analysis, cDNA samples were used, and a 10 μL reaction mixture was prepared. The mixture comprised 2 microliter cDNA, one μL primers of both forward and reverse primers, 5 μL of PowerUp™ SYBR™ Green Master Mix, and 1–2 microliter of nuclease-free water. The thermal cycling conditions for the reaction were as follows: an initial 2 min hold at 50 °C, another 2 min hold at 95 °C, and then 40 cycles of denaturation for 15 sec at 95 °C followed by annealing at 60 °C for 1 min. 2
−ΔΔCt (Livak’s method) was used to analyze the relative changes in gene expression, and a multivariate ANOVA test was performed to determine statistical significance. Following the differential expression of ATGs, the log2 fold change was calculated. Grade 2 gliomas, considered low grade, were chosen as the baseline group for computing the log2 fold change (FC).
Table 8 shows the list of primers that were utilized.
4.7. Statistical Analysis
Descriptive statistics was used to express the means with standard deviation. Pearson’s Chi-square test was executed for the association of demographics, clinical pathologic parameters, and molecular markers with tumor type and grade. Multivariate ANOVA test was performed to identify any notable difference in the relative gene expression among all the examined ATGs in 2, 3, and 4 WHO grades of diffuse gliomas, and p < 0.05 was considered significant. Spearman’s correlation test was executed to determine the correlation between autophagy genes in grade 2, 3, and 4 gliomas. We performed all the analyses using IBM SPSS version 24 and used a significance threshold of p < 0.05 to determine statistical associations.

 ,
, 




{kind=link}
{kind=link}
{kind=link}